Introduction
As the most common primary liver cancer, and the
second most common cause of cancer-related deaths worldwide
(1), hepatocellular carcinoma
(HCC) is one of the most common cancers (2). In Asia, HCC is the second leading
cause of cancer related deaths (3). Chronic infection with hepatitis B and
C, exposure to aflatoxin, smoking, excessive alcohol use and
metabolic syndromes are the main etiological factors (4). Of these, HBV infection is the most
common cause, especially in Asia (5). A high propensity to metastasize,
active angiogenesis and rapid proliferation are the common causes
of rapid recurrence and poor prognoses in HCC patients (6).
Fatty acid synthase (FASN) is a key enzyme for
catalyzing endogenous fatty acid synthesis, a precursor of some
biologically important lipids (7).
Since the original determination that FASN is highly expressed in
breast cancer and is related to tumor stage and prognosis,
increasing number of studies have found that it has effect on tumor
occurrence and development in various cancers, such as esophageal,
lung, ovarian, prostate, nasopharyngeal and gastric cancers and
malignant melanoma (8–11). As shown in a previous study, FASN
contributed to migration and invasion in HCC cells (12) and osteosarcoma (OS) cells (13). However, few studies have focused on
the mechanism by which FASN promotes migration and invasion of
tumor cells.
In the present study, we took advantage of isobaric
tags for relative and absolute quantitation (iTRAQ) method to
identify the differentially expressed secretory proteins (DEPs)
between HepG2215 cells with or without silencing of the FASN gene.
After verification and further analysis by RT-PCR and western blot
analysis, we identified a novel pathway which may explicate the
promotional effect of FASN on metastasis of HCC.
Materials and methods
Reagents
The eight-plex iTraq kits were acquired from Applied
Biosystems (Foster City, CA, USA). All electrophoresis reagents
were purchased from Bio-Rad (Hercules, CA, USA). CytoSelect™
24-well Cell Migration and Invasion assay kits (8 µm,
colorimetric format) were purchased from Cell Biolabs (San Diego,
CA, USA). Opti-MEM was purchase from Gibco (San Diego, CA, USA).
Lipofectamine 2000, FASN specific siRNA oligonucleotides
(HSS103565, HSS176712, HSS176713) and a negative control
(12935–400) were from Invitrogen (Carlsbad, CA, USA). Monoclonal
antibodies against fatty acid synthase (FASN, ab128870),
antithrombin-III (SERPINC1, ab126598), proprotein convertase
subtilisin/kexin type 9 (PCSK9, ab181142), insulin-like growth
factor binding protein 1 (IGFBP1, ab180948), hypoxia-inducible
factor-1α (HIF-1α, ab51608), ubiquitin (ab134953) were obtained
from Abcam (Cambridge, MA, USA). Apolipoprotein E (APOE, TA805358),
neutrophil gelatinase-associated lipocalin (LCN2, TA322583),
translationally-controlled tumor protein (TPT1, TA314640) were from
OriGene (Rockville, MD, USA). CoCl2 (S2619) was from
Sigma (St. Louis, MO, USA), and MG132 (C8661) was purchased from
Selleck Chemicals (Houston, TX, USA).
Cell lines
Human liver cancer cell lines, HepG2, HepG2215 and
Bel7402, were acquired from ATCC and cultured in high-glucose DMEM
with 10% fetal bovine serum (FBS; Gibco), containing 1% penicillin
and streptomycin. Cells were incubated in an atmosphere of 5.0%
CO2 at 37°C.
Protein collection and iTRAQ
labeling
Total secretory protein was collected, filtered by
needle filter, and concentrated by centrifugation with an
Amicon® centrifugal filter (Billerica, MA, USA). The
protein concentration was determined using a 2-D Quant kit. For
each sample, 100 µg of protein was precipitated, denatured,
cysteine blocked, and digested with sequencing-grade modified
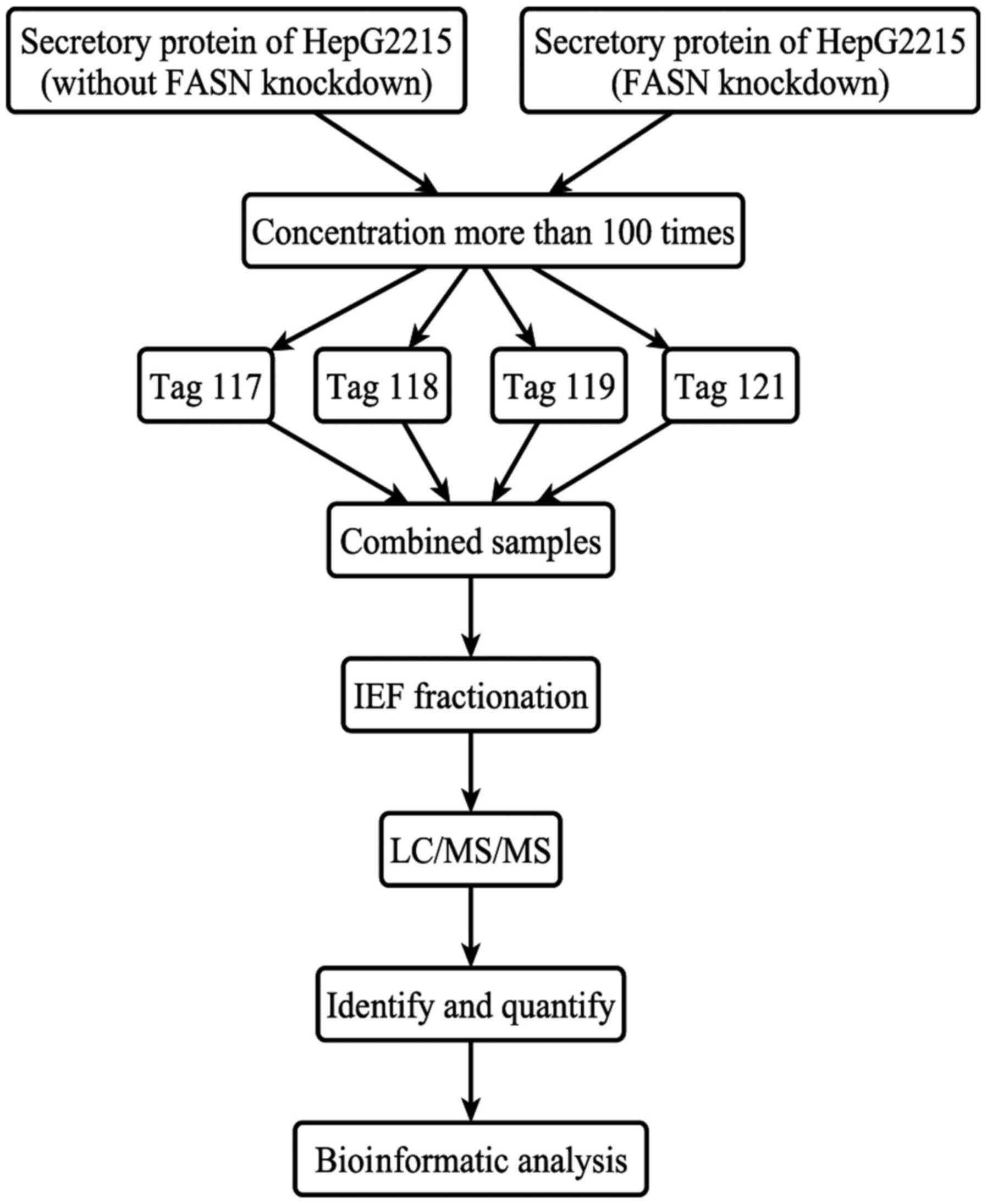
trypsin, according to the manufacturer's instructions. Labeling was
performed as follows: pooled secretory protein samples without FASN
knockdown were labeled with 117 and 118 tags; pooled secretory
protein samples with FASN knockdown were labeled with 119 and 121
tags. Prior to analysis, the labeled samples were pooled.
Fractionation of peptides
Fractions of the iTRAQ-labeled peptides were
solubilized with a Pharmalyte and urea (Amersham Biosciences)
solution. After dissolution, the samples were applied to pH
3.0–10.0 IPG gel strips (Amersham Biosciences) for 14 h at 30 V.
The peptides were subsequently focused successively for 1 h at 500
V, 1 h 1,000 V, and, finally, at 8,000 V for 8.5 h on IPG phor
(Amersham Biosciences). Peptides were extracted from the gel by use
of a solution containing 0.1% formic acid and 2% acetonitrile. The
fractions were lyophilized in a vacuum concentrator and purified on
a SPE C18 column. Prior to mass spectrometric analysis, the
purified fractions were lyophilized and stored at −20°C.
Mass spectrometry
After purification, each iTRAQ-labeled sample was
resuspended in a 2% acetonitrile solution containing 0.1% formic
acid, and injected into a QStar ESI Quadrupole time-of-flight mass
spectrometer (Applied Biosystems) coupled to online capillary LC
system (Amsterdam, The Netherlands). A gradient series for each
analysis was loaded on the C18-PepMap column (Dionex) to separate
the peptide mixture at a flow rate of 0.3 µl/min. For data
acquisition, positive ion mode was used with a 300–1,800 m/z mass
range. We selected the two most abundantly charged peptides >20
counts, and excluded peptides with a mass tolerance of 30 sec ± 50
mDa.
Data were processed using ProteinPilot v.2.0
(Applied Biosystems, Waltham, MA, USA) and searching in the
Uniprotkb database. MMTS modification of cysteine was specified as
a fixed modification. Protein identification was based on selection
thresholds of ProtScore >1.3 or ProtScore <0.77, and false
discovery rate P-values <0.05.
Immunohistochemistry (IHC) and tissue
microarrays (TMA)
Tissue microarrays containing 70 HCC samples from
different cases and 70 matched HCC adjacent normal tissue were
obtained from US Biomax Inc. (Rockville, MD, USA). After being
fixed with 1% formalin, the tissue samples were embedded in
paraffin and cut into sections. The sections were then warmed in a
60°C oven, deparaffinized and rehydrated. After 3 washes with PBS
(5 min per wash), the sections were subjected to heat-induced
antigen retrieval in 0.01 M citrate solution for 5 min in a
microwave. Endogenous peroxidase activity was quenched by
incubating the sections in 3% H2O2 for 10
min. The sections were blocked with bovine serum albumin (BSA) for
30 min and were incubated overnight with antibodies against FASN
(1:200) at 4°C. Detection was performed on an Envision/Horseradish
Peroxidase system (Dako-Cytomation, Glostrup, Denmark), and all
slides were counterstained with Gill's hematoxylin for evaluation
under a light microscope.
For further analysis by tissue microarray, TMAs were
baked at 60°C, and immunohistochemical procedures were performed as
above. The expression of FASN was assessed by semi-quantitative
scoring of the staining intensity (scale 0–3) and the percentage of
positive cells (0–100%). The staining intensity and positive cell
scores were then multiplied, generating a score ranging from 0 to
300. In order to maintain the consistency of the measurements, the
same qualified pathologist assessed each sample and interpreted the
hematoxylin and eosin sections and analysis/scoring of IHC
data.
FASN siRNA transfection, Transwell assay
and wound healing assay
Cells were transfected with 50 nM of FASN-specific
siRNA or a negative control siRNA (12935–400) or blank control
(Lipo2000) using Lipofectamine 2000 (Thermo Fisher Scientific,
Carlsbad, CA, USA) and opti-MEM (Gibco). The cells were cultured in
high-glucose DMEM with 10% fetal bovine serum after transfection.
Wound healing and Transwell assays were conducted 2 days after
transfection. When the cells cultured in 6-well plates reached 100%
confluence, a sterile p200 pipette tip was used to incise a wound
in the cell monolayer, and the debris was removed by gently washing
with PBS. The resulting gap was examined at 0 and 24 h under a
microscope. The capacity of cell migration was determined by the
closure of the wound channel. The Transwell invasion was performed
using a Cell Invasion assay kit (Cell Biolabs, Inc., Beijing,
China), according to the manufacturer's protocol. As determined by
trypan blue exclusion, ~1×105 viable and
siRNA-transfected cells were seeded in the upper chamber, separated
from a lower chamber by an 8-µm pore size membrane. ECM
Matrix gel was used to determine the number of invading cells by
CyQuant GR fluorescent dye (560 nm) at 24 h after seeding. The
determination of FASN downregulation was made via western blot
analysis. All experiments were performed in triplicate.
Western blot analysis
Cells were lysed with lysis buffer, and protein
concentration was determined via an Enhanced BCA Protein assay kit
(Beyotime, Jiangsu, China). The proteins were separated by SDS-PAGE
and subsequently transferred to PVDF membranes. The membranes were
blocked with BSA for 1 h at room temperature, and incubated with
the primary antibodies (1:500–1:2,000) overnight at 4°C. The
membranes were washed three times with Tris-buffered saline
containing Tween-20 (TBST) buffer and incubated with HRP-conjugated
secondary antibodies at a dilution of 1:5,000. The membranes were
washed again with TBST and visualized with an ECL detection system
(Bio-Rad Laboratories). Western blot analyses were performed in
triplicate.
RNA extraction and quantitative
RT-PCR
Total RNA was extracted using TRIzol reagent
(Invitrogen), according to the manufacturer's instructions.
First-strand cDNA was synthesized from 2 µg of total RNA,
using a Reverse Transcription kit (Thermo Fisher Scientific,
Waltham, MA, USA). A Fast PCR kit (KAPA SYBR, MA, USA) was used to
perform RT-PCR with primers for SERPINC1 (Hs00379789_CE), APOE
(Hs00185983_CE), LCN2 (Hs00761477_CE), PCSK9 (Hs00358728_CE), APOA1
(Hs00609868_CE), IGFBP1 (Hs00287487_CE), GAPDH (Hs00115502_CE),
TPT1 (Hs00806865_CE), GDF15 (Hs00783664_CE). Quantification of gene
expression was calculated according to the 2−ΔΔCT
method. RT-PCR analyses were conducted in triplicate.
Detection of HIF-1α activity
All reagents and equipment used in the
Dual-Luciferase Reporter assay system were acquired from Promega
Corp. (Madison, WI, USA). Detection of HIF-1α activity was
performed as described previously (14). The luciferase vector pGL3-HRE
(encoding firefly luciferase) was used to monitor promoter activity
and the pRL-SV40 plasmid (encoding Renilla luciferase) was
used as normalization control. Each sample was set up with 3
parallel expriments.
HepG2215 cells were transiently transfected with
FASN siRNA or control siRNA. After transfection, the cells were
seeded into 24-well plates and cultured in DMEM supplemented with
10% fetal bovine serum in 5% CO2 at 37°C. After 24 h,
all cells were transiently transfected with 500 ng pGL3-HRE and 15
ng pRL-SV40 constructs using Lipofectamine 2000. At 24 h
post-transfection of plasmid, the promoter activities were
determined via the Dual-luciferase assay system, and normalized
against the internal control activity of Renilla luciferase.
Each experiment was done in triplicate.
Transient transfection of
pCMV-HIF-1α
The pCMV- HIF-1α plasmid was acquired from The
Institute for Viral Hepatitis, The Second Affiliated Hospital of
Chongqing Medical University, Chongqing, China. Transfection was
performed using Lipofectamine 2000, according to the manufacturer's
instructions. Twenty-four hours before transfection,
5×105 HepG2212 cells were seeded into a 6-well plate and
transfected with 1 µg plasmid. Cells were treated with other
conditions after cultured 48 h post transfection.
Immunoprecipitation (IP)
Cells cultured under hypoxic conditions for 6 h were
collected for immunoprecipitation. Briefly, the cells were lysed
with IP lysis buffer (Beyotime) and the lysate was centrifuged to
pellet the cellular debris. The supernatant was transferred to a
separate microfuge tube, precleared with protein A/G agarose beads
and centrifuged to pellet the beads and removed protein impurity.
Again, the pellet was discarded and the supernatant was transferred
to a sterile microfuge tube and incubated with mouse or rabbit IgG
overnight at 4°C. The following day, the supernatant was incubated
with protein A/G agarose beads coupled with the indicated
antibodies for >2 h. The suspension was centrifuged to
concentrate the beads, which then were washed thrice with IP lysis
buffer (3 min per wash). The final pellet was resuspended with the
2X SDS loading buffer used for western blot analysis.
Statistical analysis
All experiments were conducted at least three times.
Continuous variables are presented as the mean ± standard deviation
(SD). Comparisons between groups were analyzed by the Student's
t-test or a Mann-Whitney U test. Qualitative variables are
presented as counts and percentages, which were analyzed with the
χ2 test. Differences in the data were considered to be
statistically significant when the P-value was <0.05.
Results
Differential expression of FASN in
tissues
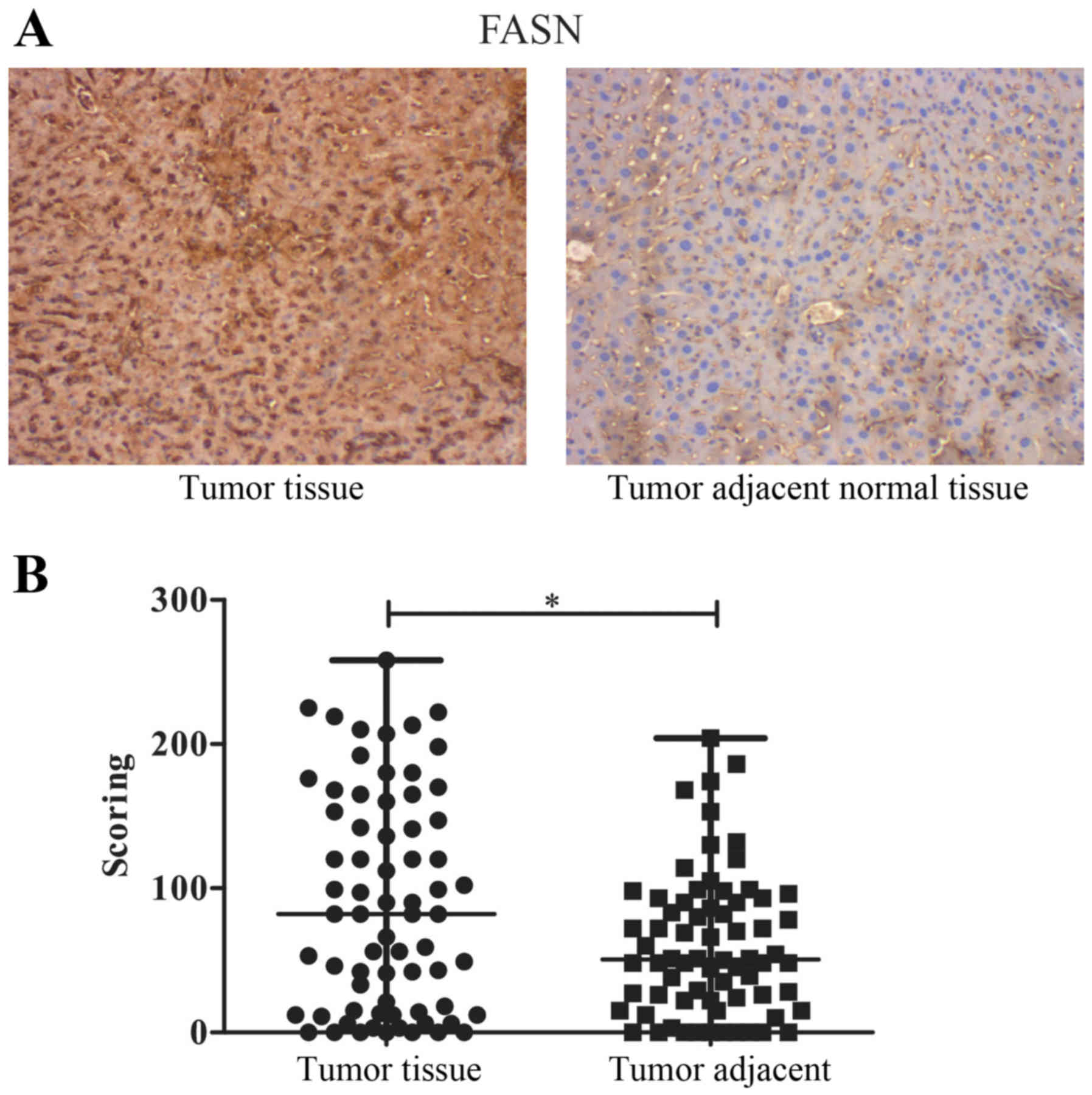
To verify the previous determination that the
expression of FASN was higher in HCC tissue, compared to normal
tissue adjacent to HCC tissue, IHC was performed on tissue
microarrays of hepatocellular carcinoma and non-cancerous tissues
to identify the differences in FASN expression. The results showed
that FASN was expressed higher in cancer samples than in
non-cancerous samples (Fig. 1A).
IHC score values of FASN were significantly higher in the HCC
tissue group than in the HCC adjacent normal tissue group (Fig. 1B), similar to previous results.
Effect of FASN on cell migration and
invasion in HCC
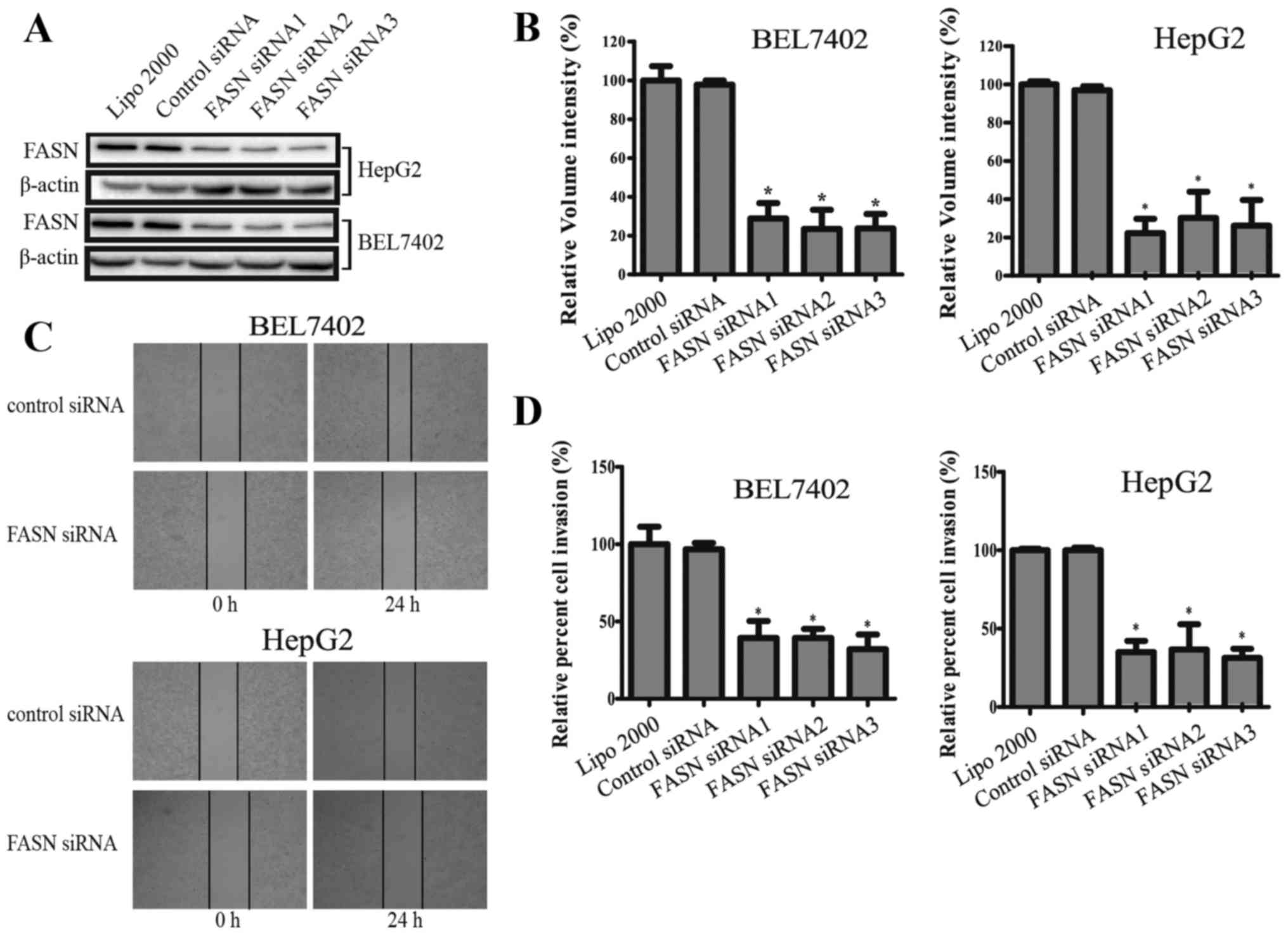
To validate the association of FASN to HCC migration
and invasion, three FASN siRNA and control siRNA were used to
transfect HepG2 and BEL7402 cell lines. The FASN siRNA
significantly downregulated the expression of FASN (Fig. 2A and B). According to the results,
it was found that the invasive ability of HepG2 and BEL7402 was
significantly decreased after RNAi mediated silencing of FASN
(Fig. 2D). According to the wound
healing test, FASN knockdown resulted in a significant decrease in
the migration ability of HepG2 and BEL7402 cells (Fig. 2C).
iTRAQ analysis of differentially
expressed proteins
In order to study the mechanism by which FASN
influences cell migration and invasion, iTRAQ-based MS was
performed on proteins isolated from HepG2215 after siRNA-mediated
FASN knockdown and from unsilenced control cells (Fig. 3). For ProteinPilot-based database
searching and identification, the threshold was set to achieve 95%
confidence at 5% false discovery rate. The iTRAQ results revealed
that the protein profiles differed significantly between the FASN
knockdown and unsilenced control samples. The ratios of 119:117 and
121:118 identified the differential protein expression and,
according to the commonly adopted iTRAQ-based MS conventions
(15), a ratio <0.77 (1/1.3) or
>1.3 (1.3/1) was considered as significantly downregulated or
upregulated, respectively. According to previous studies, technical
variation <30% can be achieved by repeat iTRAQ experiments.
A total of 30 proteins were identified with a
confidence of 95%, of which 8 were significantly upregulated and 22
down-regulated (Table I). To
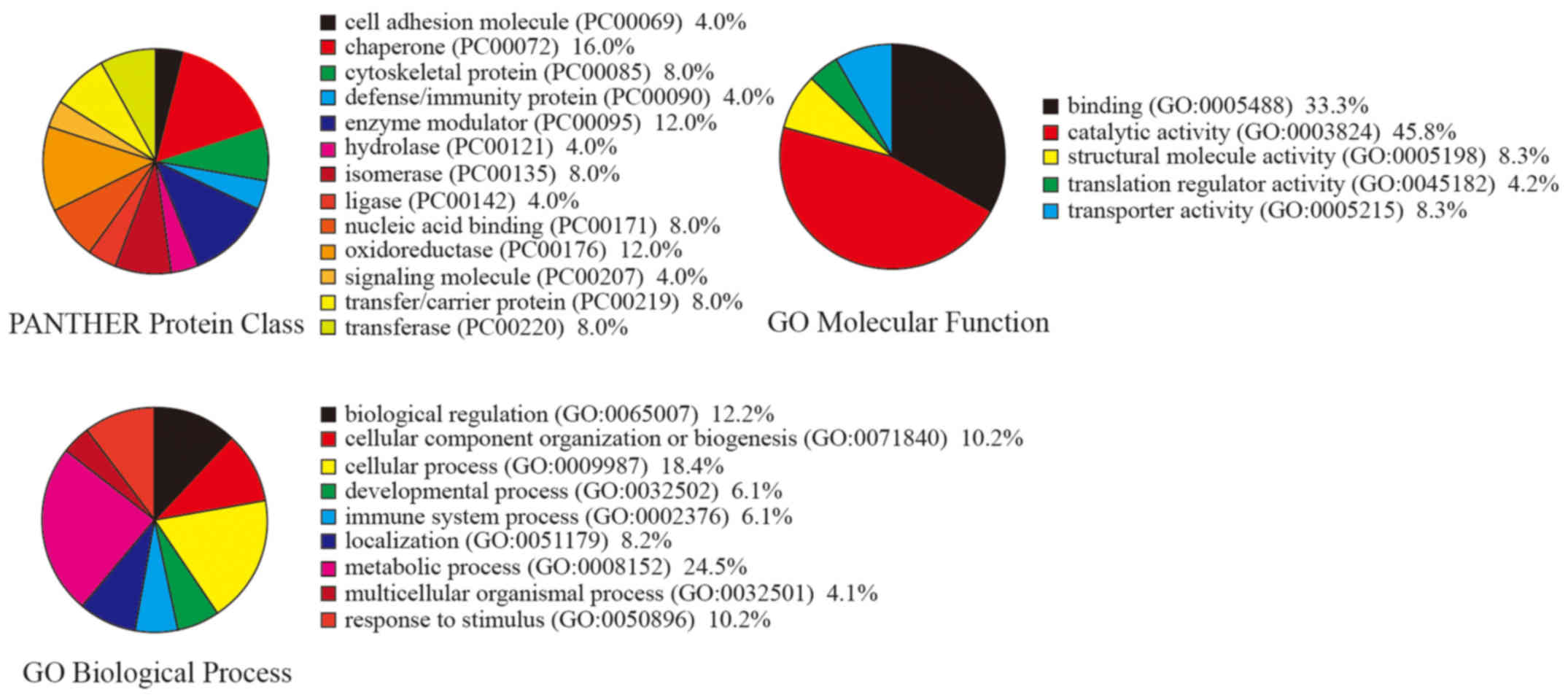
identify the cellular and molecular characteristics of these
proteins, the differentially expressed proteins were grouped
according to their reported molecular functions and biological
process using the PANTHER (www.pantherdb.org/) Classification System. The 30 DEPs
hold 13 protein classes, 9 biological processes and 5 molecular
functions (Fig. 4). The results
showed that 'catalytic activity' and 'binding' were the most common
molecular functions while 'metabolic process' was the most common
biological process.
 | Table IThe proteins observed to be
differentially expressed by iTRAQ analysis in supernatant of
hepG2215: FASN knockdown vs control. |
Table I
The proteins observed to be
differentially expressed by iTRAQ analysis in supernatant of
hepG2215: FASN knockdown vs control.
| N | Accession no. | Gene symbol | Protein name | Peptides (95%) | FASN knockdown :
control
119:117 | Pval
119:117 | FASN knockdown :
control
(121:118) | Pval
121:118 |
|---|
| 1 |
sp|P01008|ANT3_HUMAN | SERPINC1 |
Antithrombin-III | 19 | 1.739886999 | 0.00141695 | 2.154124975 | 5.13E-06 |
| 2 |
sp|P02647|APOA1_HUMAN | APOA1 | Apolipoprotein
A-I |
102 | 1.489493012 | 0.000435383 | 1.488209963 | 0.005546236 |
| 3 |
sp|P02649|APOE_HUMAN | APOE | Apolipoprotein
E | 37 | 1.790249944 | 0.000148533 | 2.041884899 | 0.000425205 |
| 4 |
sp|Q06520|ST2A1_HUMAN | SULT2A1 | Bile salt
sulfotransferase | 8 | 1.346820951 | 0.026328281 | 1.469063997 | 0.01441405 |
| 5 |
sp|P05546|HEP2_HUMAN | SERPIND1 | Heparin cofactor
2 | 41 | 1.430474043 | 0.00015355 | 1.555734038 | 2.40E-05 |
| 6 |
tr|X6R8F3|X6R8F3_HUMAN | LCN2 | Neutrophil
gelatinase-associated lipocalin | 7 | 1.391461015 | 0.04503623 | 1.689599037 | 0.0504102 |
| 7 |
sp|Q8NBP7|PCSK9_HUMAN | PCSK9 | Proprotein
convertase subtilisin/kexin type 9 | 34 | 1.556053996 | 0.008803246 | 1.62815702 | 0.000942904 |
| 8 |
sp|P04004|VTNC_HUMAN | VTN | Vitronectin | 16 | 1.437466025 | 0.004682186 | 1.433408022 | 0.006844658 |
| 9 |
sp|P61604|CH10_HUMAN | HSPE1 | 10 kDa heat shock
protein, mitochondrial | 26 | 0.633759379 | 0.002095495 | 0.667803884 | 0.040668011 |
| 10 |
sp|P11021|GRP78_HUMAN | HSPA5 | 78 kDa
glucose-regulated protein | 64 | 0.686024487 | 6.97E-06 | 0.7461766 | 0.001131944 |
| 11 |
sp|Q14008|CKAP5_HUMAN | CKAP5 |
Cytoskeleton-associated protein 5 | 11 | 0.729114115 | 0.00480286 | 0.737102091 | 0.000435989 |
| 12 |
tr|Q6FI51|Q6FI51_HUMAN | DNAJB1 | DNAJB1 protein | 10 | 0.717387795 | 0.0145011 | 0.679285109 | 0.028635601 |
| 13 |
sp|P24534|EF1B_HUMAN | EEF1B2 | Elongation factor
1-β | 10 | 0.66290468 | 0.003484956 | 0.641378522 | 0.002671605 |
| 14 |
tr|V9HW77|V9HW77_HUMAN | HEL-211 | Epididymis luminal
protein 211 | 11 | 0.605548799 | 0.001777139 | 0.621318579 | 0.009196805 |
| 15 |
sp|P49327|FAS_HUMAN | FASN | Fatty acid
synthase |
111 | 0.567631781 | 4.05E-13 | 0.518709183 | 5.75E-15 |
| 16 |
sp|P04406|G3P_HUMAN | GAPDH |
Glyceraldehyde-3-phosphate
dehydrogenase |
114 | 0.508620799 | 1.46E-05 | 0.534764171 | 5.27E-05 |
| 17 |
sp|Q99988|GDF15_HUMAN | GDF15 |
Growth/differentiation factor 15 | 49 | 0.562119782 | 0.001072533 | 0.568296909 | 2.78E-05 |
| 18 |
sp|P08107|HSP71_HUMAN | HSPA1A | Heat shock 70 kDa
protein 1A/1B | 70 | 0.669435024 | 0.002287141 | 0.655011415 | 0.006298876 |
| 19 |
sp|P11142|HSP7C_HUMAN | HSPA8 | Heat shock cognate
71 kDa protein |
118 | 0.680737495 | 8.89E-05 | 0.686902404 | 0.000633161 |
| 20 |
sp|P08238|HS90B_HUMAN | HSP90AB1 | Heat shock protein
HSP 90-β | 85 | 0.763628185 | 0.008333246 | 0.768081725 | 0.01902041 |
| 21 |
tr|Q6PEY6|Q6PEY6_HUMAN | IGFBP1 | Insulin-like growth
factor binding protein 1 | 29 | 0.482746691 | 0.00062962 | 0.422192693 | 0.007676132 |
| 22 |
tr|V9HW62|V9HW62_HUMAN | HEL-S-74 | Lactoylglutathione
lyase | 13 | 0.760211587 | 0.04173319 | 0.745779395 | 0.001756667 |
| 23 |
tr|Q68CK4|Q68CK4_HUMAN | HMFT1766 | Leucine-rich
α-2-glycoprotein | 29 | 0.631778002 | 3.93E-05 | 0.641239882 | 9.37E-05 |
| 24 |
sp|Q6UWW0|LCN15_HUMAN | LCN15 | Lipocalin-15 | 9 | 0.592697024 | 0.043171048 | 0.543676019 | 0.04871738 |
| 25 |
sp|P19338|NUCL_HUMAN | NCL | Nucleolin | 43 | 0.691573083 | 1.00E-06 | 0.724589288 | 6.62E-06 |
| 26 |
sp|P06748|NPM_HUMAN | NPM1 | Nucleophosmin | 41 | 0.6987679 | 0.004307589 | 0.74495542 | 0.029937331 |
| 27 |
sp|P05121|PAI1_HUMAN | SERPINE1 | Plasminogen
activator inhibitor 1 | 82 | 0.577512681 | 1.92E-07 | 0.583439708 | 3.19E-05 |
| 28 |
sp|O60701|UGDH_HUMAN | UGDH | UDP-glucose
6-dehydrogenase | 46 | 0.685960412 | 4.82E-05 | 0.688110411 | 0.0026483 |
| 29 |
sp|P25311|ZA2G_HUMAN | AZGP1 |
Zinc-α-2-glycoprotein | 22 | 0.723056316 | 0.025207611 | 0.69239378 | 0.001940342 |
| 30 |
tr|Q5W0H4|Q5W0H4_HUMAN | TPT1 |
Translationally-controlled tumor
protein | 10 | 0.54772937 | 0.00283738 | 0.562642515 | 0.025747631 |
Validation of differentially expressed
proteins
RT-PCR and western blot analyses were used to
validate the differentially expressed proteins identified by iTRAQ.
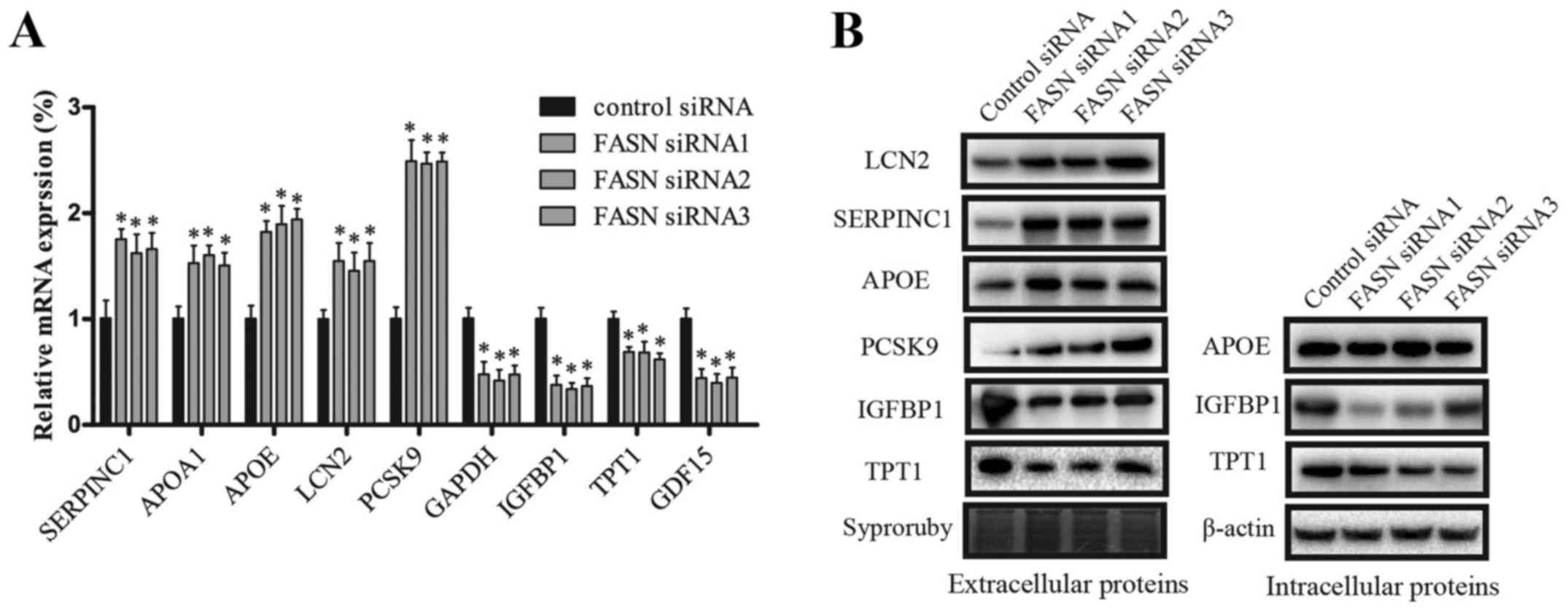
RT-PCR showed the mRNA expression levels of SERPINC1, APOA1, APOE,
LCN2, PCSK9, GAPDH, IGFBP1, TPT1, GDF15, relative to 18S. The
RT-PCR results were consistent with the results obtained via iTRAQ.
After knockdown of FASN, the mRNA expression levels of SERPINC1,
APOA1, APOE, LCN2, and PCSK9 were significantly increased, while
GAPDH, IGFBP1, TPT1 and GDF15 were decreased (Fig. 5A). Western blot analysis was
utilized to quantify the expression levels of the identified
proteins (Fig. 5B), and the
results of the extracellular LCN2, SERPINC1, APOE, PCSK9, IGFBP1,
TPT1, were consistent with the results achieved through iTRAQ
analysis. The intracellular IGFBP1 and TPT1 also followed the above
trend. However, the expression of intracellular IGFBP1 was
significantly lower than extracellular levels. After silencing
FASN, the intracellular APOE displayed no significant difference,
and the intracellular levels of SERPINC1, LCN2, and PCSK9 were
expressed so weakly that they could not be identified by western
blot analysis.
FASN promotes the development of HCC by
the effect of HIF-1α to IGFBP1
Because IGFBP1 affects the growth, proliferation and
metastasis of tumors (16), we
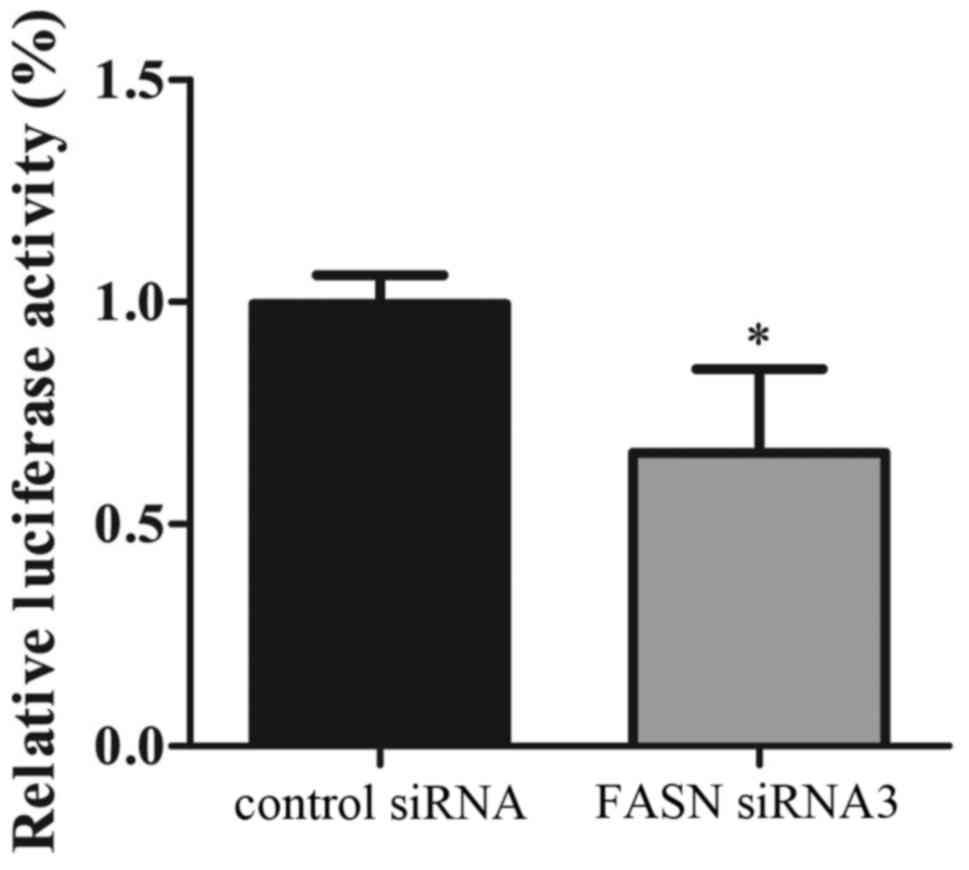
investigated the expression of HIF-1α, which is one of the upstream
precursors of IGFBP1. HIF-1α activity was determined using the
Dual-Luciferase Reporter assay system (Fig. 6). The results showed that the
activity of HIF-1α in the FASN-silenced group was decreased
significantly compared to the control group (P=0.0008). To
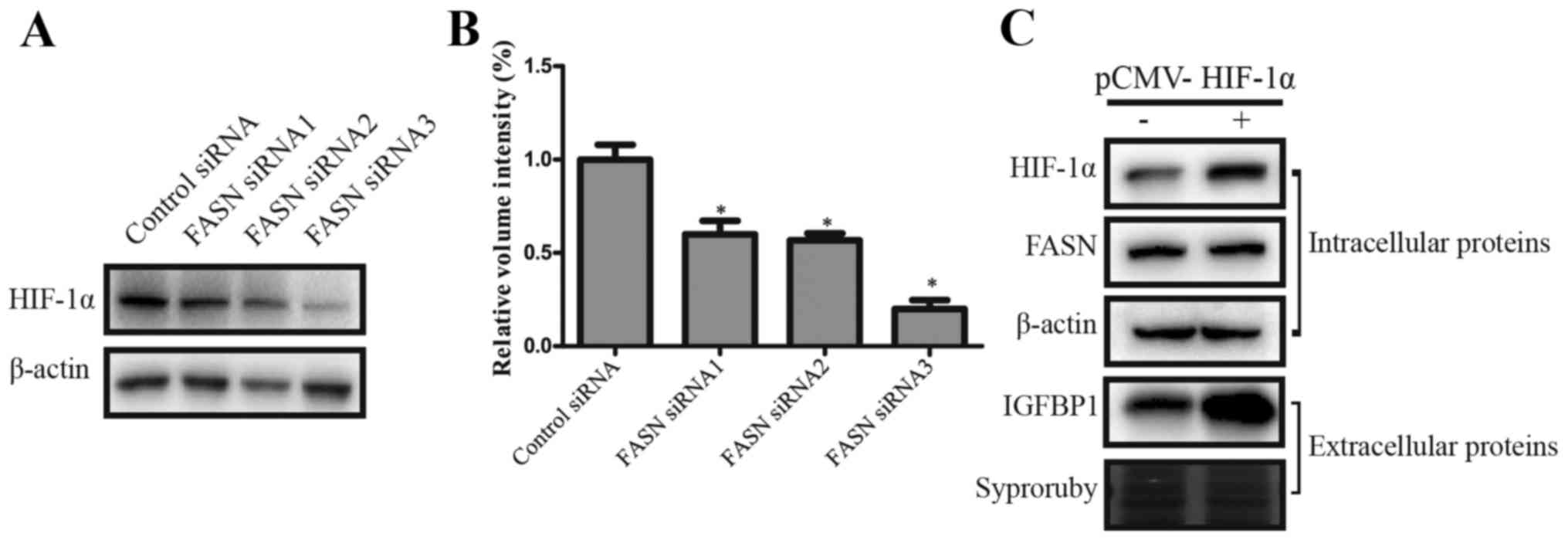
determine whether FASN regulated IGFBP1 by influencing HIF-1α, we
conducted western blot analyses to monitor the expression of HIF-1α
under FASN knockdown conditions (Fig.
7A and B). In addition, western blot analyses monitored the
expression of FASN when HIF-1α overexpression was induced by
pCMV-HIF-1α (Fig. 7C), and these
results were compared to control siRNA or plasmid control
conditions. The results suggested that FASN may regulate HIF-1α and
therefore influence IGFBP1. This pathway might be the mechanism by
which FASN affects proliferation and metastasis of HCC.
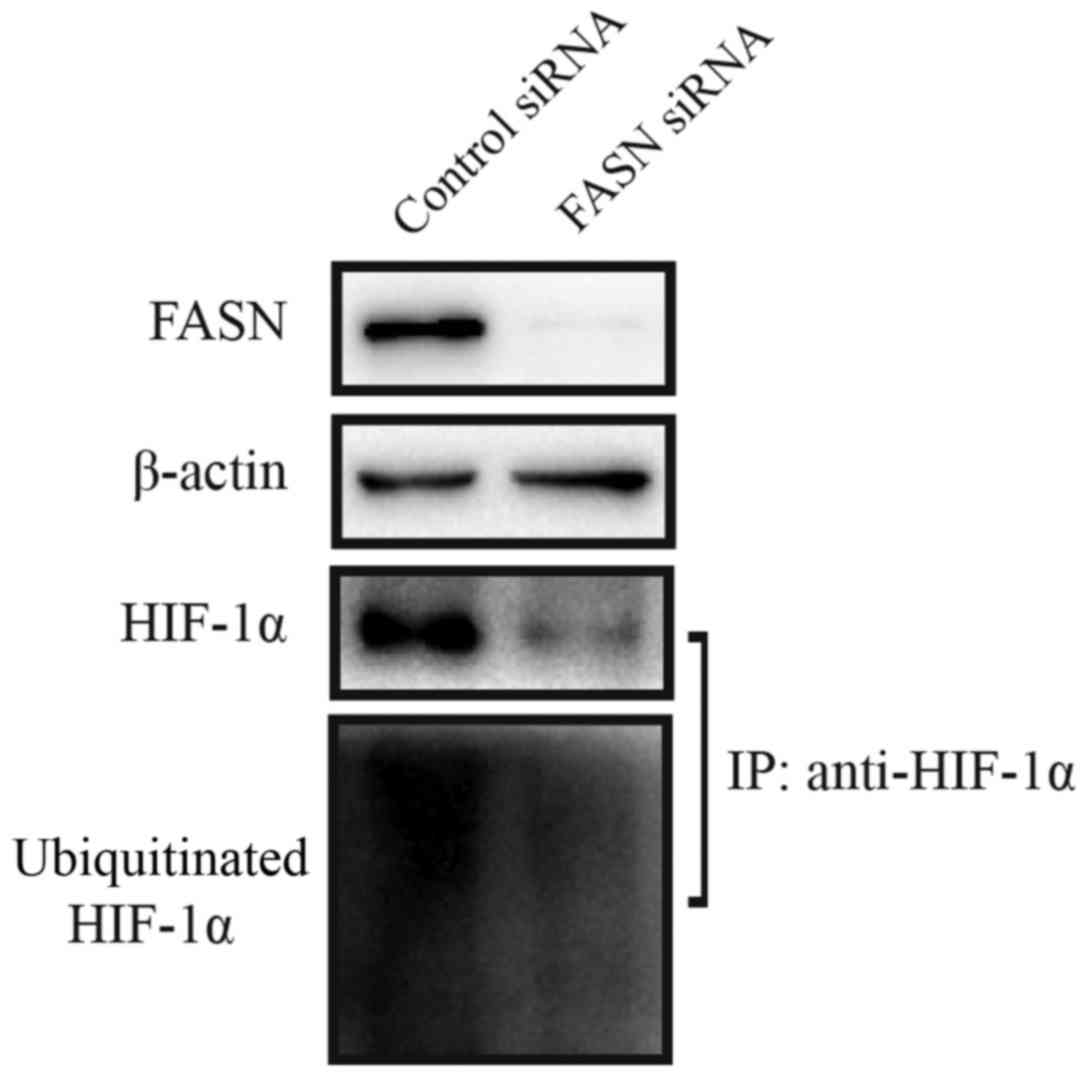
FASN affects HIF-1a ubiquitination
It is known that many pathways regulate HIF-1 α
(17), but the degradation pathway
through ubiquitination-mediated proteasome is the most important
regulator for cellular HIF-1α levels (18). We identified the influence of FASN
for HIF-1α ubiquitination by western blot analysis after
immunoprecipitation (Fig. 8).
After FASN knockdown, the HepG2215 cells were treated with
CoCl2 (100 µM), to induce a hypoxic response, and
MG132 (10 µM), to inhibit proteasome directed HIF-1α
degradation. The results revealed that inhibiting FASN could
downregulate HIF-1α ubiquitination.
Discussion
The incidence of HCC is increasing continuously, and
diagnosis is frequently made only during the late stages of disease
(19). Although surgical resection
and liver transplantation are the most successful treatments
(20), patients suffering from
advanced stage HCC have a high mortality rate. Metastasis and
recurrence are the major obstacles to improvement of survival rates
and quality of life in HCC patients (21). In recent years, many studies have
confirmed that the occurrence of tumors is closely related to
abnormal cell metabolism (22).
FASN, as the key molecule for catalyzing fatty acid synthesis, is
highly expressed in a number of malignant tumors. In this study,
the results of the comparison of FASN expression between liver
tumor tissue and adjacent non-cancerous tissue were in accordance
with previous reports (23). We
further found that FASN could significantly inhibit cell migration
and invasion in Transwell assays and in the wound healing test.
According to the iTRAQ results, the majority of the
DEPs were involved in metabolic and cellular processes. We
identified many of them, including SERPINC1, APOA1, APOE, LCN2,
PCSK9, GAPDH, IGFBP1, TPT1 and GDF15 using RT-PCR. Some of them
were identified by western blot analysis. According to the results,
the iTRAQ labeling method is both reliable and powerful for protein
quantification. Based on protein classes, biological processes and
molecular functions, we further classified the DEPs into different
types, and noted the major changes after silencing FASN.
IGFBP1 belongs to the insulin-like growth factor
binding protein gene family, which consists of six
well-characterized members that encode a family of homologous
multifunctional proteins, IGFBP-1 to IGFBP-6. However, in contrast
to other proteins of this family, the expression of IGFBP-1 is
markedly altered by changes in metabolic state (24). Jones et al have reported
that IGFBP1-stimulated CHO cell migration is mediated by the
α5β1 integrin, but is independent of IGF-I
(25). Several studies have
reported that HIF-1α-responsive genes in mammals include the
IGFBP-1 (26–28). HIF-1, consisting of HIF-1α and
HIF-1β, and HIF-2 play a pivotal role in cellular adaptation to
hypoxia and regulate the expression of genes responsible for
glucose metabolism, angiogenesis, and cell survival (29). Based on the close relationship
between HIF-1α and IGFBP-1, and the observation that IGFBP-1 was
significantly decreased when FASN was silenced, we explored further
to identify the mechanism by which silencing FASN reduced
expression of HIF-1α.
Studies have reported that FASN inhibition regulates
the expression of HIF-1α to various degrees in cancer cells
(30–32). However, few studies have reported
on the relationship of FASN and HIF-1α in HCC. Our results revealed
that the level of FASN expression can influence the level of
HIF-1α, but HIF-1α did not significantly regulate the expression of
FASN. Additionally, measurements of HIF-1α activity, after
knockdown of FASN, showed that FASN promoted HIF-1α transcriptional
activity. When silencing FASN, the decrease in IGFBP1 may be
through this pathway.
The ubiquitin-proteasome is an important pathway for
protein degradation, and is the major regulator for HIF-1α s in
cells. After treating cells with CoCl2, to mimic a
hypoxic environment, and MG132, to block HIF-1a degradation, the
level of HIF-1α and ubiquitination were significantly decreased in
the FASN knockdown group, compared to the control group. The
results indicated that FASN has an effect on ubiquitination of
HIF-1α.
Lu et al considered that if FASN is a target
gene of HIF, FASN levels would change with the change of HIF levels
(33). However, Zhou et al
reported inhibition of FASN blocked HIF-1α in glioma cells
(32). In different tissues, the
relationship of FASN and HIF-1α is different. In the present study,
we determined that inhibition of FASN decreased the level of
HIF-1α, IGFBP1, HIF-1α activity and ubiquitination, in the HepG2215
cell line. As is known, FASN and HIF-1α could promoted the
occurrence, development and metastasis of tumors (13,23,33–39).
Especially for the promotional effect of HIF-1α on tumor cell
metastasis, many previous studies have indicated this conclusion,
and reported some relevant pathways (14,40–43).
It is reasonable, then, to conclude that the effect of FASN on the
proliferation, migration, invasion, and metastasis of hepatoma
carcinoma cells, and angiogenesis within tumor tissues, is based on
the role of HIF-1α. In future, these findings can bring some new
insights for the treatment of HCC. FASN may be a new target for
inhibiting tumor development and metastasis, and for improving the
survival rate and quality of life of patients with hepatocellular
carcinoma.
In conclusion, the present study provides a novel
mechanism by which FASN promotes the proliferation, migration,
invasion, metastasis of hepatocellular carcinoma cells, and
angiogenesis in tumor tissues. Inhibition of FASN suppresses
migration, invasion and healing of hepatoma carcinoma cells by
decreasing HIF-1α activity and IGFBP1.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (81171560), National Key Technology
Support Program (2012BAI35B03), the 'Par-Eu Scholars Program' of
Chongqing City, and the National Science and Technology Major
Project of China (2012ZX10002007001).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
3
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Colombo M and Sangiovanni A:
Hepatocellular carcinoma. Recenti Prog Med. 107:386–394. 2016.In
Italian. PubMed/NCBI
|
|
5
|
Dondeti MF, El-Maadawy EA and Talaat RM:
Hepatitis-related hepatocellular carcinoma: Insights into cytokine
gene polymorphisms. World J Gastroenterol. 22:6800–6816. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sucandy I, Cheek S, Golas BJ, Tsung A,
Geller DA and Marsh JW: Longterm survival outcomes of patients
undergoing treatment with radiofrequency ablation for
hepatocellular carcinoma and metastatic colorectal cancer liver
tumors. HPB Oxf. 18:756–763. 2016. View Article : Google Scholar
|
|
7
|
Liu H, Liu JY, Wu X and Zhang JT:
Biochemistry, molecular biology, and pharmacology of fatty acid
synthase, an emerging therapeutic target and diagnosis/prognosis
marker. Int J Biochem Mol Biol. 1:69–89. 2010.PubMed/NCBI
|
|
8
|
Cerne D, Zitnik IP and Sok M: Increased
fatty acid synthase activity in non-small cell lung cancer tissue
is a weaker predictor of shorter patient survival than increased
lipoprotein lipase activity. Arch Med Res. 41:405–409. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Daker M, Bhuvanendran S, Ahmad M, Takada K
and Khoo AS: Deregulation of lipid metabolism pathway genes in
nasopharyngeal carcinoma cells. Mol Med Rep. 7:731–741.
2013.PubMed/NCBI
|
|
10
|
Grunt TW, Wagner R, Grusch M, Berger W,
Singer CF, Marian B, Zielinski CC and Lupu R: Interaction between
fatty acid synthase- and ErbB-systems in ovarian cancer cells.
Biochem Biophys Res Commun. 385:454–459. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou Y, Niu C, Li Y, Gao B, Zheng J, Guo X
and Ma W: Fatty acid synthase expression and esophageal cancer. Mol
Biol Rep. 39:9733–9739. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hao Q, Li T, Zhang X, Gao P, Qiao P, Li S
and Geng Z: Expression and roles of fatty acid synthase in
hepatocellular carcinoma. Oncol Rep. 32:2471–2476. 2014.PubMed/NCBI
|
|
13
|
Long XH, Mao JH, Peng AF, Zhou Y, Huang SH
and Liu ZL: Tumor suppressive microRNA-424 inhibits osteosarcoma
cell migration and invasion via targeting fatty acid synthase. Exp
Ther Med. 5:1048–1052. 2013.PubMed/NCBI
|
|
14
|
Shi T, Dong Y, Li J, Gao P, Fu D and Ma D:
High-throughput screening identifies CHMP4A associated with
hypoxia-inducible factor 1. Life Sci. 87:604–608. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gan CS, Chong PK, Pham TK and Wright PC:
Technical, experimental, and biological variations in isobaric tags
for relative and absolute quantitation (iTRAQ). J Proteome Res.
6:821–827. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Firth SM and Baxter RC: Cellular actions
of the insulin-like growth factor binding proteins. Endocr Rev.
23:824–854. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Frede S, Berchner-Pfannschmidt U and
Fandrey J: Regulation of hypoxia-inducible factors during
inflammation. Methods Enzymol. 435:405–419. 2007.PubMed/NCBI
|
|
18
|
Richard DE, Berra E, Gothié E, Roux D and
Pouysségur J: p42/p44 mitogen-activated protein kinases
phosphorylate hypoxia-inducible factor 1alpha (HIF-1alpha) and
enhance the transcriptional activity of HIF-1. J Biol Chem.
274:32631–32637. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qiao M, Sheng S and Pardee AB: Metastasis
and AKT activation. Cell Cycle. 7:2991–2996. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Belghiti J and Fuks D: Liver resection and
transplantation in hepatocellular carcinoma. Liver Cancer. 1:71–82.
2012. View Article : Google Scholar
|
|
21
|
Schwartz M, Roayaie S and Llovet J: How
should patients with hepatocellular carcinoma recurrence after
liver transplantation be treated? J Hepatol. 43:584–589. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cairns RA, Harris I, McCracken S and Mak
TW: Cancer cell metabolism. Cold Spring Harb Symp Quant Biol.
76:299–311. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang H, Li H, Yang Y, Li S, Ren H, Zhang
D and Hu H: Differential regulation of host genes including hepatic
fatty acid synthase in HBV-transgenic mice. J Proteome Res.
12:2967–2979. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee PD, Giudice LC, Conover CA and Powell
DR: Insulin-like growth factor binding protein-1: Recent findings
and new directions. Proc Soc Exp Biol Med. 216:319–357. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jones JI, Gockerman A, Busby WH Jr, Wright
G and Clemmons DR: Insulin-like growth factor binding protein 1
stimulates cell migration and binds to the alpha 5 beta 1 integrin
by means of its Arg-Gly-Asp sequence. Proc Natl Acad Sci USA.
90:10553–10557. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Semenza GL: Oxygen sensing, homeostasis,
and disease. N Engl J Med. 365:537–547. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sugawara J, Tazuke SI, Suen LF, Powell DR,
Kaper F, Giaccia AJ and Giudice LC: Regulation of insulin-like
growth factor-binding protein 1 by hypoxia and 3′,5′-cyclic
adenosine monophosphate is additive in HepG2 cells. J Clin
Endocrinol Metab. 85:3821–3827. 2000.PubMed/NCBI
|
|
28
|
Tuomisto TT, Rissanen TT, Vajanto I,
Korkeela A, Rutanen J and Ylä-Herttuala S: HIF-VEGF-VEGFR-2,
TNF-alpha and IGF pathways are upregulated in critical human
skeletal muscle ischemia as studied with DNA array.
Atherosclerosis. 174:111–120. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bertout JA, Patel SA and Simon MC: The
impact of O2 availability on human cancer. Nat Rev Cancer.
8:967–975. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Menendez JA, Vellon L, Oza BP and Lupu R:
Does endogenous fatty acid metabolism allow cancer cells to sense
hypoxia and mediate hypoxic vasodilatation? Characterization of a
novel molecular connection between fatty acid synthase (FAS) and
hypoxia-inducible factor-1alpha (HIF-1alpha)-related expression of
vascular endothelial growth factor (VEGF) in cancer cells
overexpressing her-2/neu oncogene. J Cell Biochem. 94:857–863.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Seguin F, Carvalho MA, Bastos DC, Agostini
M, Zecchin KG, Alvarez-Flores MP, Chudzinski-Tavassi AM, Coletta RD
and Graner E: The fatty acid synthase inhibitor orlistat reduces
experimental metastases and angiogenesis in B16-F10 melanomas. Br J
Cancer. 107:977–987. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou Y, Jin G, Mi R, Zhang J, Zhang J, Xu
H, Cheng S, Zhang Y, Song W and Liu F: Inhibition of fatty acid
synthase suppresses neovascularization via regulating the
expression of VEGF-A in glioma. J Cancer Res Clin Oncol.
142:2447–2459. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lu GY, Huang SM, Liu ST, Liu PY, Chou WY
and Lin WS: Caffeine induces tumor cytotoxicity via the regulation
of alternative splicing in subsets of cancer-associated genes. Int
J Biochem Cell Biol. 47:83–92. 2014. View Article : Google Scholar
|
|
34
|
Chang L, Wu P, Senthilkumar R, Tian X, Liu
H, Shen X, Tao Z and Huang P: Loss of fatty acid synthase
suppresses the malignant phenotype of colorectal cancer cells by
down-regulating energy metabolism and mTOR signaling pathway. J
Cancer Res Clin Oncol. 142:59–72. 2016. View Article : Google Scholar
|
|
35
|
Liu ZL, Mao JH, Peng AF, Yin QS, Zhou Y,
Long XH and Huang SH: Inhibition of fatty acid synthase suppresses
osteosarcoma cell invasion and migration via downregulation of the
PI3K/Akt signaling pathway in vitro. Mol Med Rep. 7:608–612.
2013.
|
|
36
|
Mao JH, Zhou RP, Peng AF, Liu ZL, Huang
SH, Long XH and Shu Y: microRNA-195 suppresses osteosarcoma cell
invasion and migration in vitro by targeting FASN. Oncol Lett.
4:1125–1129. 2012.PubMed/NCBI
|
|
37
|
Singh R, Yadav V, Kumar S and Saini N:
MicroRNA-195 inhibits proliferation, invasion and metastasis in
breast cancer cells by targeting FASN, HMGCR, ACACA and CYP27B1.
Sci Rep. 5:174542015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang TF, Wang H, Peng AF, Luo QF, Liu ZL,
Zhou RP, Gao S, Zhou Y and Chen WZ: Inhibition of fatty acid
synthase suppresses U-2 OS cell invasion and migration via
down-regulating the activity of HER2/PI3K/AKT signaling pathway in
vitro. Biochem Biophys Res Commun. 440:229–234. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yu B, Miao ZH, Jiang Y, Li MH, Yang N, Li
T and Ding J: c-Jun protects hypoxia-inducible factor-1alpha from
degradation via its oxygen-dependent degradation domain in a
nontranscriptional manner. Cancer Res. 69:7704–7712. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cai Q, Wang Z, Wang S, Weng M, Zhou D, Li
C, Wang J, Chen E and Quan Z: Long non-coding RNA LINC00152
promotes gallbladder cancer metastasis and epithelial-mesenchymal
transition by regulating HIF-1alpha via miR-138. Open Biol.
7:1602472017. View Article : Google Scholar
|
|
41
|
Maroni P, Matteucci E, Bendinelli P and
Desiderio MA: Functions and epigenetic regulation of Wwox in bone
metastasis from breast carcinoma: Comparison with primary tumors.
Int J Mol Sci. 18:182017. View Article : Google Scholar
|
|
42
|
Sun LL, Song Z, Li WZ and Tang SY: Hypoxia
facilitates epithelial-mesenchymal transition-mediated rectal
cancer progress. Genet Mol Res. 15:152016. View Article : Google Scholar
|
|
43
|
Yan X, Jiao SC, Zhang GQ, Guan Y and Wang
JL: Tumor-associated immune factors are associated with recurrence
and metastasis in non-small cell lung cancer. Cancer Gene Ther.
2017. View Article : Google Scholar : PubMed/NCBI
|