Introduction
Bile acids are synthesized from cholesterol and are
conjugated to either taurine or glycine. The primary bile acids are
dehydroxylated by intestinal bacteria, which gives rise to
secondary bile acids, including deoxycholic acid (DCA). Most bile
acids are absorbed from the terminal ileum and are returned to the
liver by enterohepatic circulation. Several bile acids, malabsorbed
by the small intestine, are transported to the colon where they
affect colonic motility. In certain pathological conditions, for
example, irritable bowel syndrome and ileal surgical resection,
bile acid reabsorption is compromised. Previous studies suggested
that bile acids affect gastrointestinal motility (1–3).
Bile acids have been reported to inhibit ileal motility in the
isolated perfused rabbit terminal ileum and guinea pig gallbladder
smooth muscle strips (1,2). Moreover, DCA has been shown to
inhibit spontaneous mechanical contractions in canine colonic
smooth muscle (3).
In non-muscle cells, bile acids act through a
variety of signaling mechanisms, including the activation of
protein kinase C (PKC) or cyclooxygenase, or the generation of
reactive oxygen species (ROS) (4–6). DCA
inhibits the pacemaker currents of the interstitial cells of Cajal
(ICC) by activating KATP channels through the production
of PGE2(7). L-type
Ca2+ channels in intestinal smooth muscle are thought to
trigger contractile activity. Intestinal smooth muscle
contractility is dependent on the activation of L-type
Ca2+ channels, which result in Ca2+ entry and
the initiation of contractions (8). The effects and mechanism of DCA
activity on the contraction of colon smooth muscles are not
completely understood. Our aim was to investigate the possible
mechanism by which DCA affects colon motility in vitro.
Materials and methods
Animals
Male Wistar rats weighing 200–250 g were used in
this study. Rats were given free access to food and water in a
temperature- and relative humidity-controlled room (23–25°C,
relative humidity 55–60%) with a 12-h light/dark cycle for 1 week.
Animals were maintained according to animal care guidelines from
the Institutional Animal Care and Use Committee of Wuhan
University. Approval was obtained from the ethics committee of
Wuhan University.
Preparation of colonic strips
Rats were anesthetized with ethyl ether prior to
cervical exsanguination. The proximal colon was cut along the
mesenteric border and cleaned with Ca2+-free
physiological saline solution (PSS). Muscle strips (~3×10 mm) were
removed and placed in Ca2+-free PSS, which was
oxygenated continuously. Following the careful removal of the
mucosa and submucosa by dissection, smooth muscle strips were
obtained.
Contraction recording of colonic smooth
muscle strips
Following the method described in our previous study
(9), the muscle strips were
mounted in 7 ml Tyrode’s solution and connected to an isometric
force transducer (JZJOIH, Chengdu, China). The Tyrode’s solution
was kept at 37°C and bubbled with carbogen (95% O2 + 5%
CO2). A tension of 1.0 g was slowly applied to the
muscle strips prior to drug treatment. The muscle strips were
allowed to equilibrate for 30–60 min, with the solution being
changed every 20 min. The strips were washed at least three times
with Tyrode’s solution between each experimental condition. The
mean contractile amplitude and frequency were recorded as control
values and the effects of drugs as response values. The results
were presented as the percentage change, calculated as: percentage
change = (response value - control value)/control value × 100%.
Cell isolation
Colon smooth muscle cells (SMCs) were dissociated by
enzymatic digestion according to a previously described method
(10). The colon tissue was pinned
to the Sylgard surface of a Petri dish and the mucosa was carefully
dissected away under an anatomical microscope. The smooth muscle
strips were then cut into small strips (2×2 mm) and placed in 2 ml
calcium-free PSS supplemented with 0.12% (w/v) collagenase (type
II), 0.2% soybean trypsin inhibitor and 0.2% bovine serum albumin
(BSA), and incubated for 20–30 min at 37°C. Pieces of tissue were
rinsed in Ca2+-free PSS solution five times to remove
the enzyme, and were maintained at 4°C for 6 h until use. Single
SMCs were isolated by several gentle triturations through the tip
of a fire-polished Pasteur pipette.
Whole-cell patch clamp recordings
For patch-clamp recordings, colon SMCs were placed
onto glass coverslips. After being allowed to adhere to the glass
for 10 min, the cells were washed with bath solution at a rate of 3
ml/min. The ion channel currents were recorded using a conventional
whole-cell patch clamp method with an EPC-10 amplifier (HEKA
Instruments Inc., Bellmore, NY, USA). Typical pipette resistances
were 3–5 MΩ. Data were filtered at 200 Hz and digitized at 10 KHz
(filter 1) and 2.9 KHz (filter 2). The experiments were performed
at 24±1°C, and the temperature of the bath solution ranged from 20
to 22°C. L-type Ca2+ currents were activated by step
depolarisation every 10 sec from a holding potential of −80 or −60
mV, between −40 and +30 mV for 400 ms in 10-mV increments to obtain
current-voltage (I-V) relationships.
Solutions and drugs
The cells were bathed in a solution (Tyrode’s
solution) containing (mM): NaCl, 147.0; KCl, 4.0; CaCl2,
2.0; NaH2PO4, 0.42;
Na2HPO4, 1.81; MgCl2, 1.05; and
glucose, 5.5 (adjusted to pH 7.35 with NaOH). The pipette solution
contained (mM): CsCl, 135; MgCl2, 4; HEPES, 10;
Na2-ATP, 2; EGTA, 10; and tetraethylammonium (TEA), 20
(adjusted to pH 7.3 with Tris base). The Ca2+-free PSS
contained (mM): NaCl, 135; KCl, 5.0; MgCl2, 1.2;
glucose, 10.0; and HEPES, 10 (adjusted to pH 7.4 with NaOH). The
drugs used were: acetylcholine (Ach), DCA, nifedipine,
chelerythrine and phorbol 12,13-dibutyrate (PDBu), and were
purchased from Sigma (St. Louis, MO, USA). Collagenase was
purchased from Gibco (Invitrogen Life Technologies, Carlsbad, CA,
USA). Nifedipine, chelerythrine and PDBu were prepared as stock
solutions in dimethylsulfoxide (DMSO) and stored at −20°C. The
final concentration of DMSO used in the experiments was <0.03%,
which had no effect on the strips and cells.
Statistical analysis
The drug-induced changes in contractile response
relative to the basal levels were calculated as percentages, and
all values were expressed as the mean ± standard error (SE).
Differences in the data were evaluated by the Student’s t-test.
P<0.05 was considered to indicate a statistically significant
result. The n values reported in the text refer to the number of
cells used in patch-clamp experiments.
Results
Effects of DCA on contractions of
proximal colon smooth muscle strips
DCA decreased the amplitude of spontaneous
contractions of the proximal colonic smooth muscle (PCSM) strips in
a concentration-dependent manner, but not the frequency of the
spontaneous contractions. DCA affected the contraction of the
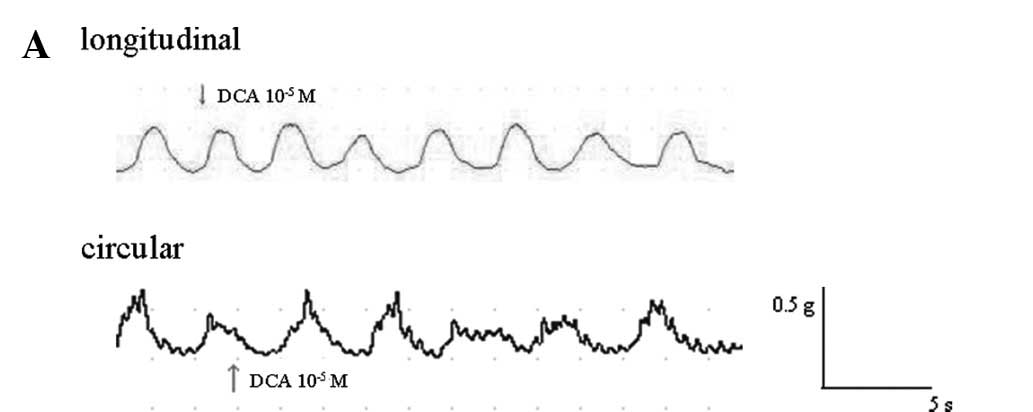
strips when applied for 2–3 min. Fig.
1A shows representative traces which demonstrate the effect of
DCA (10−5 M) on the resting tension of longitudinal and
circular muscle strips.
The mean contractile amplitude of longitudinal
muscle was reduced by 5.63±2.77, 10.95±2.76 and 19.89±3.33% at DCA
concentrations of 10−6, 10−5 and
10−4 M, respectively (all n=6; p<0.05). The
contractile amplitude of circular muscle was reduced by 5.22±1.63,
14.84±2.09 and 21.15±3.28%, respectively (all n=6; p<0.05)
(Fig. 1B).
When the contraction evoked by Ach or KCl had
reached a plateau, DCA (10−5 M) was administered. The
administration of DCA induced the relaxation of PCSM (followed by
circular muscle) strips pre-contracted with Ach (10 μM) or KCl (80
mM) by 12.2±1.5%, p<0.05 and 16.3±6.9%, p<0.05, respectively.
Data were expressed as the percentage changes in tension from the
contraction induced by Ach or KCl, respectively.
Increasing the concentration of CaCl2
resulted in a concentration-dependent contraction of the strips.
Incubation with DCA (10−5 M) for 20 min shifted the
concentration-response curves of CaCl2 to the right and
decreased the maximal response (Fig.
1C).
Pre-treatment of proximal colonic strips with the
PKC inhibitor chelerythrine for 20 min significantly attenuated the
effects of DCA (10−5 M) on the Ach-induced contractions
(Fig. 1D).
Effects of DCA on L-type Ca2+
currents in colonic smooth muscle
L-type calcium current (ICa-L) was
recorded with calcium as the current carrier when the pipette
contained a high-Cs solution. The maximum current reached ~0 mV
under these conditions, and was depressed by nifedipine (10 μM) by
~80% (n=6; p<0.05). The average cell capacitance (Cm) was
60.007±20.17 pF (n=60) and the access resistance was 5–10 MΩ. A
series resistance compensation (lag of 60 us) of 70–75% was applied
during each recording.
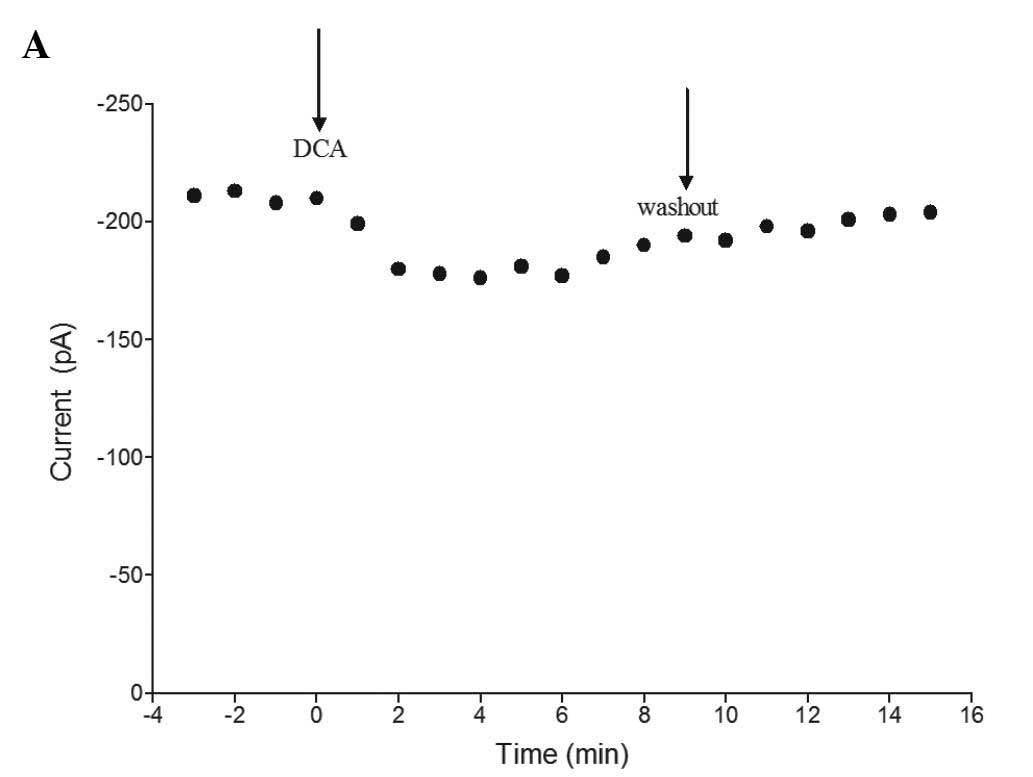
When DCA (10−5 M) was applied for 2–3
min, the peak ICa-L was reduced. As time increased,
ICa-L reduced to a steady state level and was partially
restored following a long-time washout. Fig. 2A shows the time-dependent effect of
DCA in a representative cell. The currents were evoked by a step to
0 mV from a holding potential of −40 mV.
As shown in Fig.
2B, current traces were obtained under control conditions and
during superfusion with DCA at concentrations from 10−6
to 10−4 M. Fig. 2C
shows that DCA suppressed the ICa-L at 0 mV by
6.02±0.87, 15.02±1.73 and 47.14±3.79% at concentrations of
10−6-, 10−5 and 10−4 M,
respectively (all n=6; p<0.05).
Fig. 2D shows the
average steady-state I-V relationships in the presence and absence
of 10−5 M DCA, indicating that the drug did not change
the shape of the I-V curve. When DCA was applied, the amplitude of
the peak current was decreased from 213±4.6 to 181±4.9 pA compared
with the control group (at 0 mV; n=6, p<0.05).
Effects of PKC on the action of DCA on
ICa-L in colonic smooth muscle cells
To determine whether PKC was involved in the action
of DCA, we used PDBu, a PKC activator, and chelerythrine, a potent
PKC inhibitor.
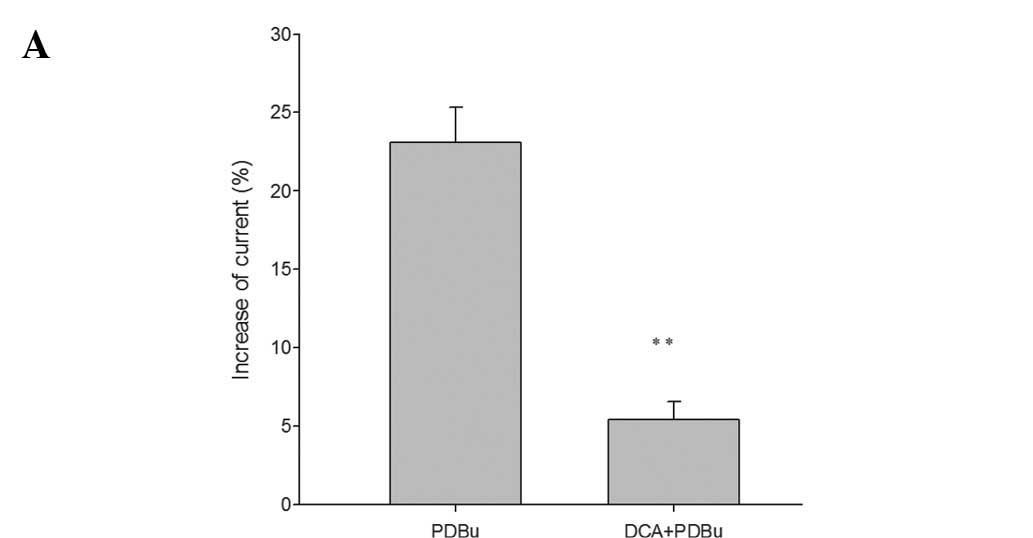
Firstly, we tested the effects of PDBu (100 nM) and
found that it promoted the ICa-L by 23.11±2.2% at 0 mV
(n=6; p<0.05). The results demonstrated that PKC enhanced the
L-type calcium currents in colonic SMCs. We then tested the effects
of PDBu on the ICa-L in cells treated with DCA
(10−5 M) for 3 min. We found that the promoting effect
of PDBu on the ICa-L was reduced to 5.45±1.12% in the
presence of DCA (Fig. 3A).
Secondly, pre-treatment of the colonic SMCs with the
PKC inhibitor chelerythrine (1 μM) greatly reduced the decrease of
ICa-L caused by DCA (10−5 M) (Fig. 3B). DCA decreased the
ICa-L by only 3.57±0.35% in the presence of
chelerythrine, compared with 15.09±1.65% in the absence of
chelerythrine at 0 mV (n=6; p<0.05) (Fig. 1D). These results suggest that the
inhibitory effects of DCA on the colonic SMC ICa-L are
mediated by PKC.
Dissussion
In the present study, we investigated whether DCA
inhibited the contraction of colonic smooth muscle strips in
vitro and decreased ICa-L, possibly by the PKC
pathway, in colonic SMCs.
It has been previously reported that concentrations
of biliary DCA are increased in subjects with cholesterol
gallstones, and that they alter gallbladder motor function
(11). Therefore, it appears
likely that DCA produced in the colon affects colonic motility. DCA
has been shown to impair esophageal mucosal integrity via the
induction of apoptosis (12). In
our experiment, the effects of DCA were tested at concentrations of
10−6–10−4 M. KCl- and Ach-induced
contractions of colonic smooth muscle strips were relaxed following
the administration of DCA. The concentration-response curve of
CaCl2 was shifted to the right by incubation with DCA.
These results indicate that DCA substantially inhibits extracelluar
Ca2+ influx.
The activities of ion channels closely correlate
with the movement of smooth muscle. In the present study, we found
that the ICa-L in colonic SMC was decreased by DCA in a
concentration-dependent manner. In addition, the characteristics of
the I-V curve were not significantly altered. The effects of DCA on
calcium have been demonstrated in a previous study. For example,
bile acids were shown to stimulate calcium mobilization through
extracellular calcium influx and/or calcium release from
intracellular organelles (as reviewed in ref. 13). Our patch clamp results directly
demonstrate that DCA inhibits the contraction of smooth muscles
through the reduction of calcium entry through L-type calcium
channels. The possible effect of DCA on the release of calcium from
intracellular organelles needs to be further investigated. Previous
studies have indicated that DCA inhibits the pacemaker currents
generated by interstitial cells of Cajal (ICC); interaction with
the activating KATP channels in the ICC resulted in the
suppression of intestinal motility (7). Variations in the channel suggest that
DCA may possess differing tissue and channel effects. The activity
of DCA on ICC is consistent with its effects on smooth muscles.
Thus, we presume that DCA activates KATP channels in
colonic smooth muscles, but further investigation is required.
Inhibition was observed when applying DCA for 2–4 min, and partly
recovered following washout; thus, we suggest that the activity of
DCA is non-genomic.
PKC, which is involved in cell responses to various
stimuli, including neurotransmitters, growth factors and hormones,
is essential for cell proliferation, differentiation and the
regulation of ion channels. In smooth muscles, PKC is important for
the regulation of movement. It is well known that activation of PKC
enhances ICa-L in a variety of smooth muscles (14). For example, cholecystokinin (CCK)
increases the ICa-L of gastric abtrum via the PKC
pathway (15). Results showed that
incubation with chelerythrine significantly affected the action of
DCA on colonic smooth muscle strips pre-contracted by Ach, while
the promoting effects of PDBu on the ICa-L were reduced
in the presence of DCA. The ICa-L in the presence of
chelerythrine was not greatly reduced by DCA. The results indicate
that DCA regulates the PKC pathway. We conclude that DCA reduces
PCSM contraction and inhibits L-type calcium channels in smooth
muscle cells. This activity appears to be mediated through PKC.
Furthermore, this study suggests another possible mechanism for the
bile acid-related modulation of gastrointestinal motility.
Acknowledgements
This study was supported by the Fundamental Research
Funds for the Central Universities (Grant no. 20103020101000201).
The authors also thank the Key Laboratory of Hubei Province for
Digestive System Disease.
References
|
1
|
Armstrong DN, Krenz HK, Modlin IM and
Ballantyne GH: Bile salt inhibition of motility in the isolated
perfused rabbit terminal ileum. Gut. 34:483–488. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xu QW, Freedman SM and Shaffer EA:
Inhibitory effect of bile salts on gallbladder smooth muscle
contractility in the guinea pig in vitro. Gastroenterology.
112:1699–1706. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee HK and Lee KH: Bile acid modulation of
gastrointestinal smooth muscle contraction and ionic currents. Kor
J Physiol Pharmacol. 4:333–338. 2000.
|
|
4
|
Sokol RJ, Winklhofer-Roob BM, Devereaux MW
and McKim JM Jr: Generation of hydroperoxides in isolated rat
hepatocytes and hepatic mitochondria exposed to hydrophobic bile
acids. Gastroenterology. 109:249–256. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang F, Subraramaiah K, Altorki N and
Dannenberg AJ: Dihydroxy bile acids activate the transcription of
cyclooxygenase-2. J Biol Chem. 273:2424–2428. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zuh Y, Hua P, Rafiq S, et al:
Ca2+- and PKC-dependent stimulation of PGE2
synthesis by deoxycholic acid in human colonic fibroblasts. Am J
Physiol Gastrointest Liver Physiol. 283:G503–G510. 2002.
|
|
7
|
Jun JY, Choi S, Chang IY, et al:
Deoxycholic acid inhibits pacemaker currents by activating
ATP-dependent K+ channels through prostaglandin E2 in
interstitial cells of Cajal from the murine small intestine. Br J
Pharmacol. 144:242–251. 2005.PubMed/NCBI
|
|
8
|
Farrugia G, Rae JL and Szurszewski JH:
Characterization of an outward potassium current in canine jejunal
circular smooth muscle and its activation by fenamates. J Physiol.
468:297–310. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu L, Chen J, Yu BP, et al: Effect of
progesterone on calcium activated potassium currents and
intracellular calcium in guinea pig colon myocytes. Methods Find
Exp Clin Pharmacol. 27:475–482. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu L, Yu BP, Chen JG and Luo HS:
Mechanisms mediating serotonin-induced contraction of colonic
myocytes. Clin Exp Pharmacol Physiol. 34:120–128. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Azzaroli F, Mazzella GF, Mazzeo C, et al:
Sluggish small bowel motility is involved in determining increased
biliary deoxycholic acid in cholesterol gallstone patients. Am J
Gastroenterol. 94:2453–2459. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huo X, Juergens S, Zhang X, et al:
Deoxycholic acid causes DNA damage while inducing apoptotic
resistance through NF-κB activation in benign Barrett’s epithelial
cells. Am J Physiol Gastrointest Liver Physiol. 301:G278–286.
2011.PubMed/NCBI
|
|
13
|
Nguyen A and Bouscarel B: Bile acids and
signal transduction: role in glucose homeostasis. Cell Signal.
20:2180–2197. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cobine CA, Callaghan BP and Keef KD: Role
of L-type calcium channels and PKC in active tone development in
rabbit coronary artery. Am J Physiol Heart Circ Physiol.
292:H3079–H3088. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Si XM, Huang L, Paul SC, An P and Luo HS:
Signal transduction pathways mediating CCK-8S-induced gastric
antral smooth muscle contraction. Digestion. 73:249–258. 2006.
View Article : Google Scholar : PubMed/NCBI
|