Introduction
c-Jun is a member of the AP-1 family of leucine
zipper transcription factors, and it forms DNA-binding homodimers
with other family members, such as jun-D, c-Fos and ATF-2. The
transcription factor c-jun is a useful model used in the study of
the complexity and specificity of signaling. This inducible
transcription factor directs changes in gene expression in response
to various extracellular stimuli (1). c-Jun is involved in cell death and
survival. In particular, c-Jun has been termed ‘a killer protein’
for developing neurons and is regarded as essential for neuronal
apoptosis via the transcriptional activation of c-jun target genes
encoding pro-apoptotic proteins (2). However, c-jun also plays a protective
role against programmed cell death (3). Thus, the biological role of c-Jun in
regulating cell death or survival concerns the cell type and also
depends on the nature of the death-inducing signal. PC12 cells are
a model for dopaminergic cells with well-defined steps triggered by
apoptosis stimuli (in response to metabolic, oxidative stress or
apoptotic signals). Therefore, an advantage of the PC12 model over
other neuronal cell lines is that it has a markedly different
response to various stress stimuli. Previous studies on the
function of c-Jun in PC12 cells suggested c-jun as an executor of
death signals in neuronally differentiated PC12 cells, as well as
in fibroblasts (4). The molecular
basis for the role of c-jun in apoptosis is not fully understood.
However, regulation of the c-jun gene in differentiated PC12 cells
may be useful in the study of the molecular basis of the role of
c-jun in apoptosis.
Nitric oxide (NO) performs a variety of
physiological and pathological functions in the nervous system. NO
in cells is produced by nitric oxide synthase (NOS), of which there
are three isotypes: nNOS or bNOS (neuronal NOS), eNOS (endothelial
NOS) and iNOS (inducible NOS). In the nervous system, NO is
involved in learning and memory, neurotransmitter release,
development and neuronal damage or degeneration (5). It has also been postulated that NOS
levels are induced during the differentiation of PC12 cells by
nerve growth factor (NGF), and nNOS and NO are required for PC12
cell differentiation (6). In
NGF-treated PC12 cells, nNOS was induced at RNA and protein levels,
resulting in increased NOS activity (7). Mounting evidence suggests that the
crosstalk between NO/nNOS and the c-jun N-terminal kinase (JNK)
signaling pathway was present in the different neuronal cell lines
and in various physiological and pathological processes (8). Subsequent to inhibition of proteasome
activity, the increased nNOS protein levels correlated with
activation (phosphorylation) of JNK. JNK activation was suppressed
upon nNOS inhibition, thus establishing a pecking order in which
nNOS upregulation and formation of reactive oxygen species are
upstream of JNK phosphorylation (9). c-jun is also a substrate for a
related group of JNKs (10).
Therefore, a correlation has been suggested between c-jun and nNOS
expression in differentiated PC12 cells in vitro.
In the present study, we first detected the
expression of c-jun and nNOS in differentiated PC12 cells, and then
a c-jun siRNA transfection strategy was used to study whether the
nNOS levels in differentiated PC12 cells change following silencing
of the c-jun gene.
Materials and methods
Cell culture and transfection
The differentiated PC12 cell line was purchased from
the Shanghai Cellular Institute of China Scientific Academy
(Shanghai, China). Differentiated PC12 cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
heat-inactivated horse serum, 5% fetal bovine serum (FBS) and
antibiotics (100 U/ml penicillin A and 100 U/ml streptomycin).
Cells were grown to the subconfluent stage at 37°C in a humidified
atmosphere containing 5% CO2. Prior to siRNA
transfection manipulations, PC12 cells were plated
(2×105 cells/dish) onto 35-mm tissue culture plates and
grown in DMEM growth medium without antibiotics, since adding
antibiotics to the medium during transfection reduces transfection
efficiency. The PC12 cells were then randomly divided into two
sets. One set of the PC12 cells was used to detect the expression
of c-jun and nNOS by immunofluorescence double labeling, while the
other set was treated with growth medium only (normal control),
non-silencing siRNAs and c-jun siRNAs (Cell Signaling Technology,
Inc., Beverly, MA, USA), respectively, to investigate the
expression of c-jun and nNOS protein by western blot analysis. One
day after plating, differentiated PC12 cells were transfected for 6
h with siRNA duplexes (100 nmol/l of c-jun siRNA or control
non-silencing siRNA) using Lipofectamine 2000 (Invitrogen,
Carlsbad, CA, USA), according to the manufacturer's instructions,
and the transfection medium was changed after 6 h. Cells were
incubated at 37°C in a CO2 incubator for 72 h until they
were ready to be assayed for gene knockdown.
Immunofluorescence double labeling for
c-jun and nNOS in culture cells
For immunofluorescence double labeling, we followed
the procedures described in our previous studies (11). The cultured cells in the 96-well
plates were gently washed with PBS and fixed with 4%
paraformaldehyde in PBS at room temperature for 30 min. After
rinsing three times with PBS, the cells were treated with 0.3%
Triton X-100 at room temperature for 20 min. The cells were then
gently washed with PBS for 15 min, for 5 min each time.
Subsequently, the cells were treated with 0.1% BSA and 0.3% Triton
at room temperature for 30 min. The cells were then incubated with
rabbit anti-c-jun (1:400; Cell Signaling Technology, Inc.) and
mouse anti-nNOS (1:3,000; Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) at 4°C overnight. After washing three times with
PBS, the cells were incubated with TRITC-conjugated anti-rabbit IgG
(1:400; Sigma, St. Louis, MO, USA) and FITC-conjugated anti-mouse
IgG (1:200; Sigma) antibodies at room temperature for 2 h in the
dark. The cells were then washed extensively with PBS, and Hoechst
33258 was included in the last wash to stain the nuclei. Staining
without primary antibodies was used as a negative control. The
cells were examined and images were captured using fluorescence
microscopy (Zeiss, New York, NY, USA).
Western blot analysis for c-jun and
nNOS
To semi-quantify the c-jun and nNOS protein
expressed in differentiated PC12 cells after treatment, western
blot analysis was performed as previously described (11). Briefly, PC12 cells were grown to
subconfluence in growth medium in 60-mm dishes and then scraped
down gently by a cell slicker. The cell pellet was homogenized in
0.2 ml buffer (10 mM Tris-HCl, pH 7.4, 5 mM EDTA, 1 mM
phenylmethylsulfonyl fluoride, 20 μg/ml leupeptin, 0.1% aprotinin,
1 mM iodoacetamide, 200 μg/ml bacitracin, 20 μg/ml soybean trypsin
inhibitor, 10 mM NACl and 0.25% Triton X-100) for 10 min on ice,
followed by centrifugation at 15,000 rpm for 30 min. Protein
concentrations were determined using the Bradford protein assay
reagent (Bio-Rad, Hercules, CA, USA) with bovine serum albumin as a
standard. For western blotting, samples (20 μg protein/lane) were
separated on 8% sodium dodecyl sulphate-polyacrylamide gels
(SDS-PAGE), with a pre-stained protein ladder (5 μl) as a molecular
weight marker, and then transferred to polyvinylidene difluoride
(PVDF; Pall, Port Washington, NY, USA) membranes.
To detect c-jun and nNOS, the membranes were blocked
with 5% non-fat dry milk in Tris-buffered saline (TBS) containing
0.1% Tween-20 at room temperature for 2 h, and incubated overnight
at 4°C with c-jun polyclonal antibody (1:1,000; Cell Signaling
Technology, Inc.), nNOS polyclonal antibody (1:500; Santa Cruz
Biotechnology) or anti-β-actin (1:1,000; Boster, China). After
washing and incubation for 2 h at room temperature with a secondary
antibody conjugated with horseradish peroxidase (1:4,000), the
membranes were washed five times in 0.1% Tween-20 in TBS.
Immunoreactive bands were visualized by the supersignal west Pico
Trial kit (Pierce, Rockford, IL, USA) and blots were exposed to
X-ray film (Fujifilm, Japan). The intensity was quantified in both
bands. Relative levels of protein in the different lanes were
compared by analyzing scanned images using the NIH IMAGE program.
The studies were performed a minimum of three times using
independent cultures.
Statistical analysis
The statistical calculations and data handling were
performed using SPSS version 16.0. Variables were expressed as
medians or means ± standard deviation (χ̄ ± SD), including the
range. A one-way ANOVA was applied to detect differences among
groups followed by Tukey-Kramer multiple comparison tests.
Differences were considered significant at p<0.05.
Results
Coexpression of c-jun and nNOS in
differentiated PC12 cells
Cells were cultured as described in the experimental
procedures to examine whether there was a correlation between c-jun
and nNOS in differentiated PC12 cells. In the first series of
experiments, the expression of c-jun and nNOS in differentiated
PC12 cells was analyzed by immunofluorescent double labeling
detection. Due to their sympathetic neuronal-like phenotype,
differentiated PC12 cells are characterized by a large soma and two
unbranched processes, with the length of the processes being more
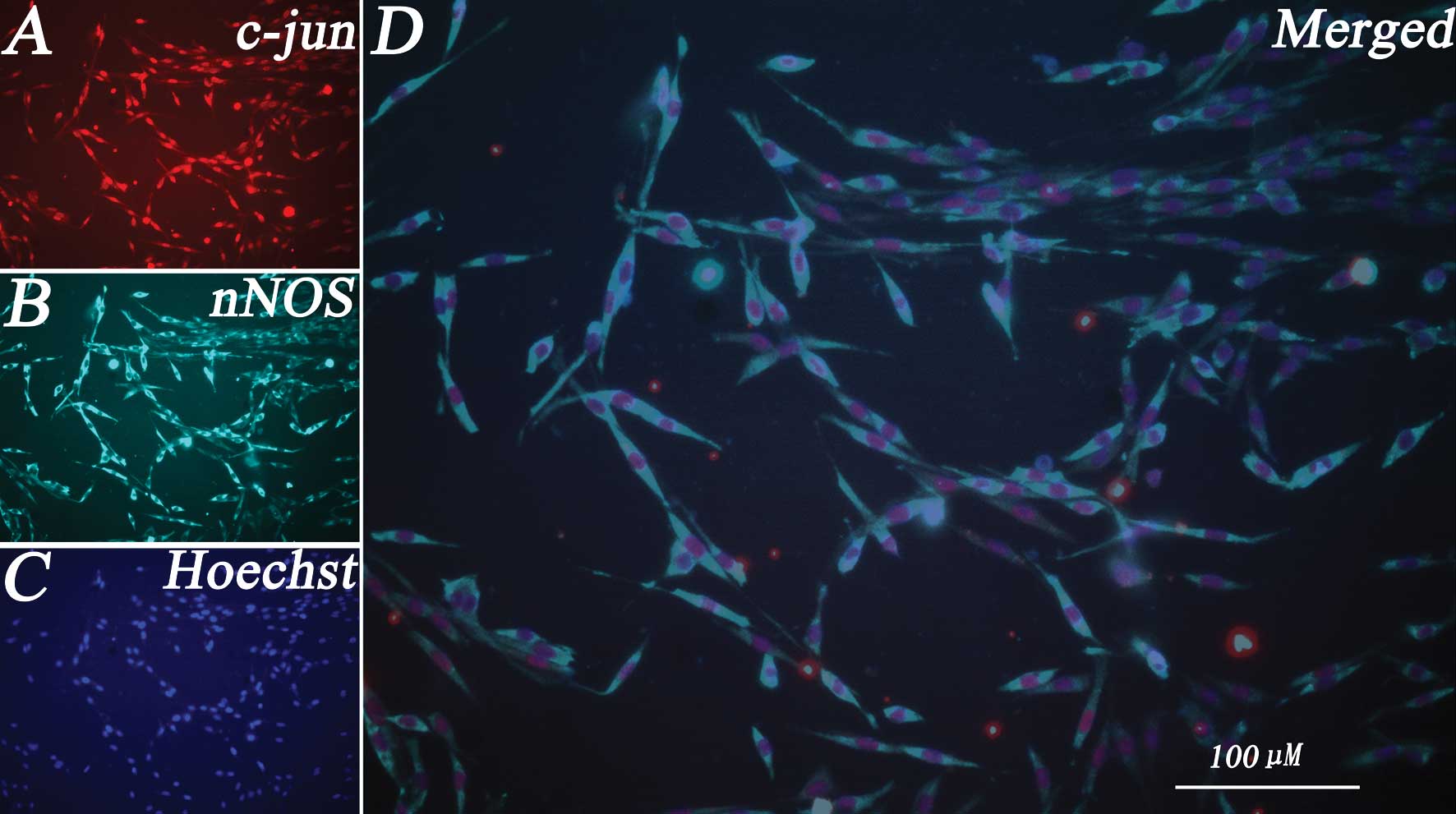
than twice the cell body (Fig. 1).
Our result showed that c-jun was clearly expressed in
differentiated PC12 cells, while c-jun immunoreactivity (ir) was
present in the nuclei (Fig. 1A).
nNOS was also expressed in differentiated PC12 cells, while nNOS-ir
was present in the cytoplasm (Fig.
1B). All the nuclei of the differentiated PC12 cells were
labeled with Hoechst 33258 (Fig.
1C). We then examined whether c-jun and nNOS expression were
colocalized inside the same cells. Immunofluorescence double
labeling further indicated that c-jun and nNOS were coexpressed in
the same differentiated PC12 cells. The results indicated that
almost all the differentiated PC12 cells coexpressed c-jun and nNOS
(Fig. 1D).
Transfection of siRNA knocks down the
c-jun gene in differentiated PC12 cells
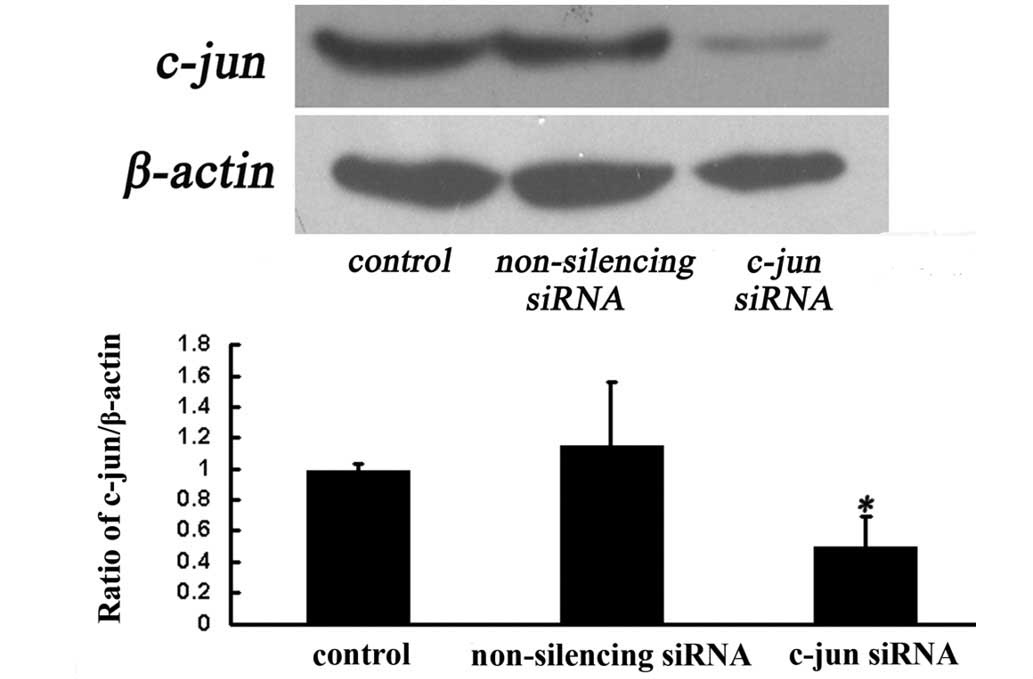
To investigate whether c-jun siRNA silences c-jun
expression in differentiated PC12 cells, we evaluated the protein
levels of c-jun in the cells after 72 h of c-jun-siRNA treatment by
western blotting. The result indicated that PC12 cells transfected
with c-jun siRNA exhibited significantly lower levels of c-jun
protein (Fig. 2A). Statistical
analysis showed that the c-jun protein in c-jun-siRNA-treated cells
was decreased by ~50% as compared to that in
non-silencing-siRNA-treated cells after 72 h of transfection
(Fig. 2B). This finding suggested
that the transient transfection of c-jun siRNA to cultured
differentiated PC12 cells was able to selectively knock down the
c-jun gene.
Suppression of c-jun expression affects
nNOS expression in differentiated PC12 cells
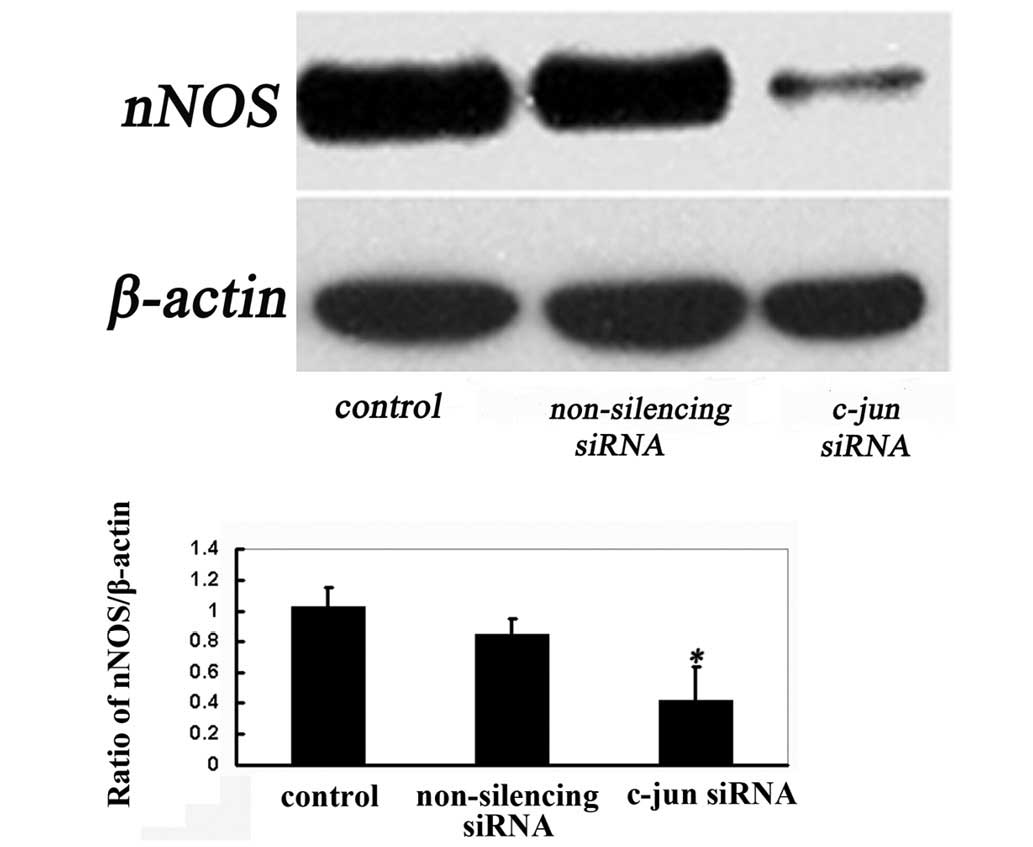
Findings of a previous study showed that the
impairment of proteasome activity and consequent increases in nNOS
levels lead to nitrative stress that involves the coordinated
response of the JNK/c-jun cytosolic signaling pathway (9). To investigate the impact on the nNOS
gene resulting from the downregulation of c-jun expression by the
siRNA transfection, nNOS protein expression was evaluated by
western blotting. Cells were harvested 72 h after transfection for
western blot analysis on nNOS protein. Our data indicated that such
silencing of c-jun siRNA resulted in a significant decrease in the
protein expression of nNOS after 72 h of transfection (Fig. 3). It is suggested that c-jun siRNA
knocks down the level of c-jun protein efficiently and affects nNOS
protein expression in differentiated PC12 cells.
Discussion
In the present study, we demonstrated that almost
all the differentiated PC12 cells coexpressed the c-jun and nNOS
genes. For the first time c-jun siRNA was shown to effectively
inhibit the expression of c-jun in vitro in differentiated
PC12 cells. We also found, for the first time, that suppression of
c-jun affected nNOS expression in differentiated PC12 cells using
the c-jun siRNA strategy.
In response to NGF treatment, PC12 cells adopt a
sympathetic neuronal phenotype (12). c-Jun is involved in neuronal
differentiation in response to NGF and in death upon NGF
withdrawal. In previously published studies, in which neuronally
differentiated PC12 cells or sympathetic neurons were used,
apoptosis induced by NGF withdrawal was prevented by
dominant-negative mutants or by microinjected antibodies specific
for c-Jun (2). In addition, the
constitutively active mutant of c-Jun was sufficient to trigger
apoptosis in cerebellar granule neurons (13). Taken together, these results
indicate that activation of c-Jun induces apoptosis in
differentiated PC12 and other neuronal cells. However, the
molecular basis for the role of c-jun in apoptosis is not fully
understood. It may depend on the target gene activated by c-jun in
the respective cell type, or the relevance or crosstalk of related
proteins. In various animal models of central neuronal diseases,
the expression of c-jun and nNOS genes is induced inside injured
neurons.
c-Jun is known to be expressed in NOS immunoreactive
neurons in the lateral geniculate nucleus of experimental
glaucomatous rats (14). Moreover,
NO-regulated activation of c-Jun N-terminal kinase 1/2 is
associated with neuronal survival in hippocampal neurons in a rat
model of ischemia (15).
Compromised proteasome degradation elevates nNOS levels and induces
programmed cell death in differentiated PC12 cells (9). In our previous study, there may have
been a functional crosstalk between c-jun and nNOS in motoneurons,
and this may have had an impact on motoneuron degeneration
(16). Our study also showed that
c-jun phosphorylation acts as the upstream molecule that inhibits
the nNOS expression in injured spinal cords within the first 2
weeks after root-avulsion injury. Additionally, a functional
relationship between c-jun and nNOS in motoneurons is possible
(17).
In the above-mentioned studies, c-jun expression
always occurred prior to expression of nNOS in motoneuron injuries.
In the present study, we investigated whether there is a
correlation between c-jun and nNOS and whether the crosstalk
between these two genes regulated the pathological progress of
injury-induced neuron degeneration. Using a c-jun siRNA strategy
for the first time, we found that suppression of c-jun affects nNOS
expression in differentiated PC12 cells. The results are in line
with the previous studies. Firstly, PC12 cell differentiation is
accompanied by the coexpression of c-jun and nNOS. Secondly, other
studies indicated c-jun as an executor of death signals (4), and that elevation of the nNOS levels
induces apoptotic cell death in neuronally differentiated PC12
cells (9). Thirdly, our data
demonstrated that the suppression of c-jun affects nNOS expression
in differentiated PC12 cells. The present findings suggest that we
cannot exclude the non-specific effects of c-jun siRNA on both the
c-jun and nNOS genes, however, we believe that the c-jun siRNA
purchased underwent specific verification after manufacture.
Therefore, based on the present study, the levels of c-jun and
other related genes should be investigated by regulating change of
the nNOS gene. Thus, investigation of the relationship between the
c-jun and nNOS genes is crucial.
The present study has shown that there is a
coexpression of c-jun and nNOS in differentiated PC12 cells. siRNA
transfection effectively knocks down the c-jun gene and suppression
of c-jun affects nNOS expression in differentiated PC12 cells. More
studies are required, however, to gain a better understanding of
the relationship between c-jun and nNOS and the underlying
molecular mechanism involved.
Acknowledgements
This study was supported by research grants from the
National Science Foundation Council of China (81070995; 31171290)
and the Chinese High Education Foundation (20110171110048).
References
|
1
|
Karin M, Liu Z and Zandi E: AP-1 function
and regulation. Curr Opin Cell Biol. 9:240–246. 1997. View Article : Google Scholar
|
|
2
|
Ham J, Babij C, Whitfield J, Pfarr CM,
Lallemand D, Yaniv M and Rubin LL: A c-Jun dominant negative mutant
protects sympathetic neurons against programmed cell death. Neuron.
14:927–939. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hilberg F, Aguzzi A, Howells N and Wagner
EF: c-jun is essential for normal mouse development and
hepatogenesis. Nature. 365:179–181. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tournier C, Hess P, Yang DD, Xu J, Turner
TK, Nimnual A, Bar-Sagi D, Jones SN, Flavell RA and Davis RJ:
Requirement of JNK for stress-induced activation of the cytochrome
c-mediated death pathway. Science. 288:870–874. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yun HY, Dawson VL and Dawson TM:
Neurobiology of nitric oxide. Crit Rev Neurobiol. 10:291–316. 1996.
View Article : Google Scholar
|
|
6
|
Phung YT, Bekker JM, Hallmark OG and Black
SM: Both neuronal NO synthase and nitric oxide are required for
PC12 cell differentiation: a cGMP independent pathway. Brain Res
Mol Brain Res. 64:165–178. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schonhoff CM, Bulseco DA, Brancho DM,
Parada LF and Ross AH: The Ras-ERK pathway is required for the
induction of neuronal nitric oxide synthase in differentiating PC12
cells. J Neurochem. 78:631–639. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pei DS, Song YJ, Yu HM, Hu WW, Du Y and
Zhang GY: Exogenous nitric oxide negatively regulates c-Jun
N-terminal kinase activation via inhibiting endogenous NO-induced
S-nitrosylation during cerebral ischemia and reperfusion in rat
hippocampus. J Neurochem. 106:1952–1963. 2008. View Article : Google Scholar
|
|
9
|
Lam PY and Cadenas E: Compromised
proteasome degradation elevates neuronal nitric oxide synthase
levels and induces apoptotic cell death. Arch Biochem Biophys.
478:181–186. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Treisman R: Regulation of transcription by
MAP kinase cascades. Curr Opin Cell Biol. 8:205–215. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li M, Wang L, Peng Y, Wang JC and Zhou LH:
Knockdown of the neuronal nitric oxide synthase gene retard the
development of the cerebellar granule neurons in vitro. Dev Dyn.
239:474–481. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sheng M and Greenberg ME: The regulation
and function of c-fos and other immediate early genes in the
nervous system. Neuron. 4:477–485. 1990. View Article : Google Scholar
|
|
13
|
Watson A, Eilers A, Lallemand D, Kyriakis
J, Rubin LL and Ham J: Phosphorylation of c-Jun is necessary for
apoptosis induced by survival signal withdrawal in cerebellar
granule neurons. J Neurosci. 18:751–762. 1998.PubMed/NCBI
|
|
14
|
Wang X, Tay SS and Ng YK: C-fos and c-jun
expressions in nitric oxide synthase immunoreactive neurons in the
lateral geniculate nucleus of experimental glaucomatous rats. Exp
Brain Res. 144:365–372. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zeng XW, Li MW, Pan J, Ji TL, Yang B,
Zhang B and Wang XQ: Activation of c-Jun N-terminal kinase 1/2
regulated by nitric oxide is associated with neuronal survival in
hippocampal neurons in a rat model of ischemia. Chin Med J (Engl).
124:3367–3372. 2011.PubMed/NCBI
|
|
16
|
Zhou LH, Han S, Xie YY, Wang LL and Yao
ZB: Differences in c-jun and nNOS expression levels in motoneurons
following different kinds of axonal injury in adult rats. Brain
Cell Biol. 36:213–227. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang LL, Zhao XC, Yan LF, Wang YQ, Cheng
X, Fu R and Zhou LH: C-jun phosphorylation contributes to down
regulation of neuronal nitric oxide synthase protein and
motoneurons death in injured spinal cords following root-avulsion
of the brachial plexus. Neuroscience. 189:397–407. 2011. View Article : Google Scholar
|