Introduction
Heparanase (HPSE) is an endoglycosidase and plays a
critical role in tumor metastasis and vascularization by
specifically degrading the heparan sulfate chains of proteoglycan,
a component of blood vessel walls and the extracellular matrix.
Previous studies demonstrated that HPSE expression is increased in
a variety of malignancies (1–8). The
human HPSE gene and its genomic structure were cloned and
characterized by several research teams simultaneously in 1999
(6–9). To understand the mechanisms of HPSE
gene expression and regulation, the HPSE promoter sequence has been
cloned and some transcription factor binding sites (TFBSs) have
also been identified. Sequence analysis revealed that the
TATA-less, GC-rich promoter of the HPSE gene belongs to the family
of housekeeping genes. There were three Sp1, four Ets-relevant
elements (ERE) and two early growth response gene-1 (EGR-1) binding
sites within the core promoter area (10,11).
Sp1, GA-binding protein (GABP) and EGR-1 were able to regulate HPSE
promoter activity. HPSE has therefore become a new intensive
research topic in the field of target gene therapy (12,13).
Targeted cancer therapy requires a targeted,
efficient and safe gene-transfer system. The application of a
tumor-specific promoter is a basic method. The tumor-specific
promoter plays a significant role in targeted gene therapy by
regulating the specific expression of downstream therapeutic genes
in tumor cells and reducing damage to normal tissues and cells
(14–16). However, the effectiveness of
available tumor-specific promoters is limited. Thus, it is
necessary to investigate a new tumor-specific promoter and identify
its activity and specificity.
Given that the HPSE promoter is also a
tumor-specific promoter and plays a significant role in targeted
gene therapy, it is crucial to understand its activity and
specificity in tumor cells. In this study, we cloned the core
fragment of the HPSE gene promoter, analyzed the TFBSs, constructed
a tumor cell expression vector using the HPSE gene promoter and for
the first time detected its transcriptional activity and
specificity in diverse cell types. The present study is likely to
provide an experimental basis for gene-targeted therapy using the
HPSE promoter.
Materials and methods
Materials
Normal human umbilical vein endothelial (ECV304),
human hepatocellular carcinoma (HepG2), laryngeal epithelial
(Hep2), erythromyeloblastoid leukemia (K562), E. coli
XL1-blue and BL21 cells were donated by Professor Xue-Long Wang
from the Department of Parasitology of Anhui Medical University,
China. The human genome extraction kit and PCR product purification
kit were purchased from Tiangen Biotech Company (Beijing, China).
The Taq enzyme, dNTP, cloning vector plasmid pMD18-T Simple,
DNA Marker DL 2000 and DL15000 were purchased from Takara
Biotechnology Company (Dalian, China). T4DNA ligase,
XhoI and HindIII were purchased from MBI Fermentas
(Burlington, ON, Canada). Tryptone, yeast extract and agar were
purchased from Oxoid Company (Hampshire, UK). Lipofectamine 2000
plasmid kit and DMEM medium were purchased from Invitrogen
Corporation (Carlsbad, CA, USA). Plasmid pEGFP-1 [multiple cloning
site (MCS) not containing promoter sequences] and pEGFP-N1 [MCS
containing cytomegalovirus promoter sequence] were purchased from
Yili company (Hefei, China), and HPSE primers were synthesized by
Sangon Biotech Company (Shanghai, China). The fluorescence
microscope was purchased from Nikon (Tokyo, Japan), and the flow
cytometer was from Beckman Coulter Inc. (Brea, CA, USA).
HPSE promoter PCR amplification and
purification
The experiment was conducted with the informed
consent of the volunteers and was approved by the Ethics Committee
of our institutes (the Affiliated Yijishan Hospital of Wannan
Medical College, Wuhu, Anhui and the Tongling People’s Hospital,
Anhui, China). Peripheral blood (20 μl) was collected from healthy,
normal volunteers. Genomic DNA was extracted according to the
conventional method, and concentration and purity were determined
by ultraviolet spectrophotometry. The primers were designed
according to the human HPSE promoter Genbank sequence (GenBank
Accession AF461265). The upstream primer (5′-GCTCAAGTGACAAGCAAGTGT
TTAT-3′) was synthesized using the only restriction enzyme cutting
site (RECS) XhoI in the HPSE DNA sequence and no additional
RECS was added. A HindIII restriction site (underlined) was
introduced into the downstream primer (5′-CCA
AGCTTGGGCTCACCTGGCTGCTC-3′). The core fragment of the HPSE
gene promoter was amplified by PCR using 10 μl genomic DNA as a
template. The PCR conditions included initial denaturation at 95°C
for 5 min, 35 cycles of amplification with subsequent denaturation
at 94°C for 55 sec, annealing at 57°C for 50 sec and extension at
72°C for 50 sec, followed by a final extension at 72°C for 10 min.
Electrophoresis was performed using 1% agarose gel. The PCR product
was purified according to the manufacturer’s instructions and
two-way sequencing was completed by Sangon Biotech Company.
Identification of the PCR products
XL1-blue competent cells were prepared using the
calcium chloride method. PCR products and pMD18-T Simple were
incubated overnight for ligation and then transformed into
competent cells. Positive colonies were identified and amplified,
plasmid DNA was extracted using the alkaline lysis method and
identified by double digestion with XhoI and HindIII.
Following 1% agarose gel electrophoresis, 1 ml bacterial suspension
containing the recombinant plasmid was sent to Sangon Biotech
Company for sequencing and the sequences were analyzed for putative
TFBS.
Construction of recombinant plasmid
vector pEGFP-Hp
The recombinant plasmid was amplified and the DNA
was extracted and digested by double digestion with XhoI and
HindIII. The digested PCR products were analysed using
agarose gel electrophoresis and then retrieved. The purified
product was a DNA fragment of the HPSE promoter. The plasmid
pEGFP-1 (4.2 kbp in length) was also digested by double digestion
and the large fragment was retrieved. During a ligation reaction,
both recycled products were incubated overnight at 4°C in order to
insert the DNA fragment of the HPSE core promoter into the pEGFP-1
vector. After the competent cell suspension was thawed at room
temperature, ligation products were added for transformation
analysis. The recombinant plasmid was screened and the plasmid DNA
was extracted by the alkaline lysis method, identified by double
digestion, PCR amplification and DNA sequencing. The positive
recombinant plasmid was designated as pEGFP-Hp.
Functional analysis of recombinant
plasmid pEGFP-Hp
Cells were cultured in 96-well plates in RPMI-1640
cell culture medium with 10% fetal calf serum (FCS) and incubated
at 37°C in a humidified atmosphere containing 5% CO2.
For each well, at 70–80% confluence, cells were inoculated into
three wells at a density of 5×105–1×106
cells/well and transfected with the pEGFP-Hp, pEGFP-1 and pEGFP-N1
plasmids. The procedure was performed according to the
manufacturer’s instructions for Lipofectamine 2000. The cells were
subsequently cultured for 48 h, the suspended cells were then
collected. The adherent cells were digested by trypsin (0.2%). The
digestion was terminated by adding blood serum culture medium. The
fluorescence intensity was observed using a fluorescence microscope
and the percentage of light-emitting cells and transfection
efficiency were determined by flow cytometry. All experiments were
repeated at least three times.
Results
Amplification, identification and
sequence analysis of HPSE core promoter
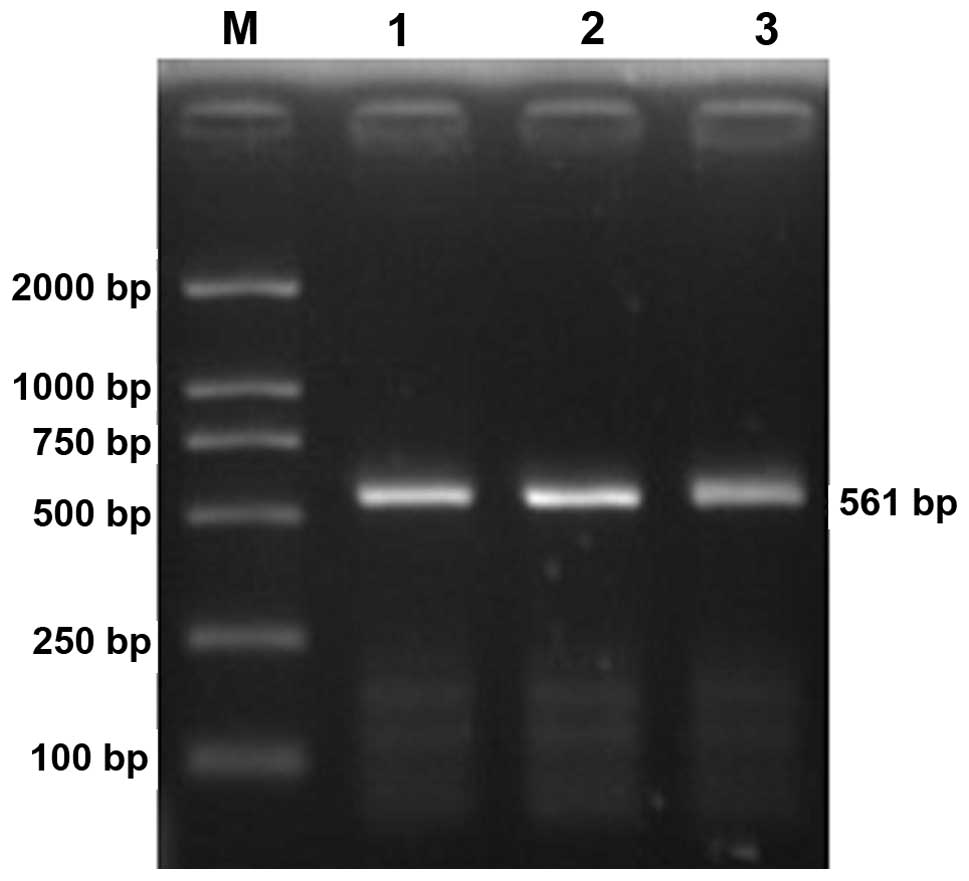
Electrophoresis of HPSE promoter PCR products
revealed a specific band between 500 and 750 bp, which was
consistent with the theoretical length of 561 bp (Fig. 1). After the PCR product was ligated
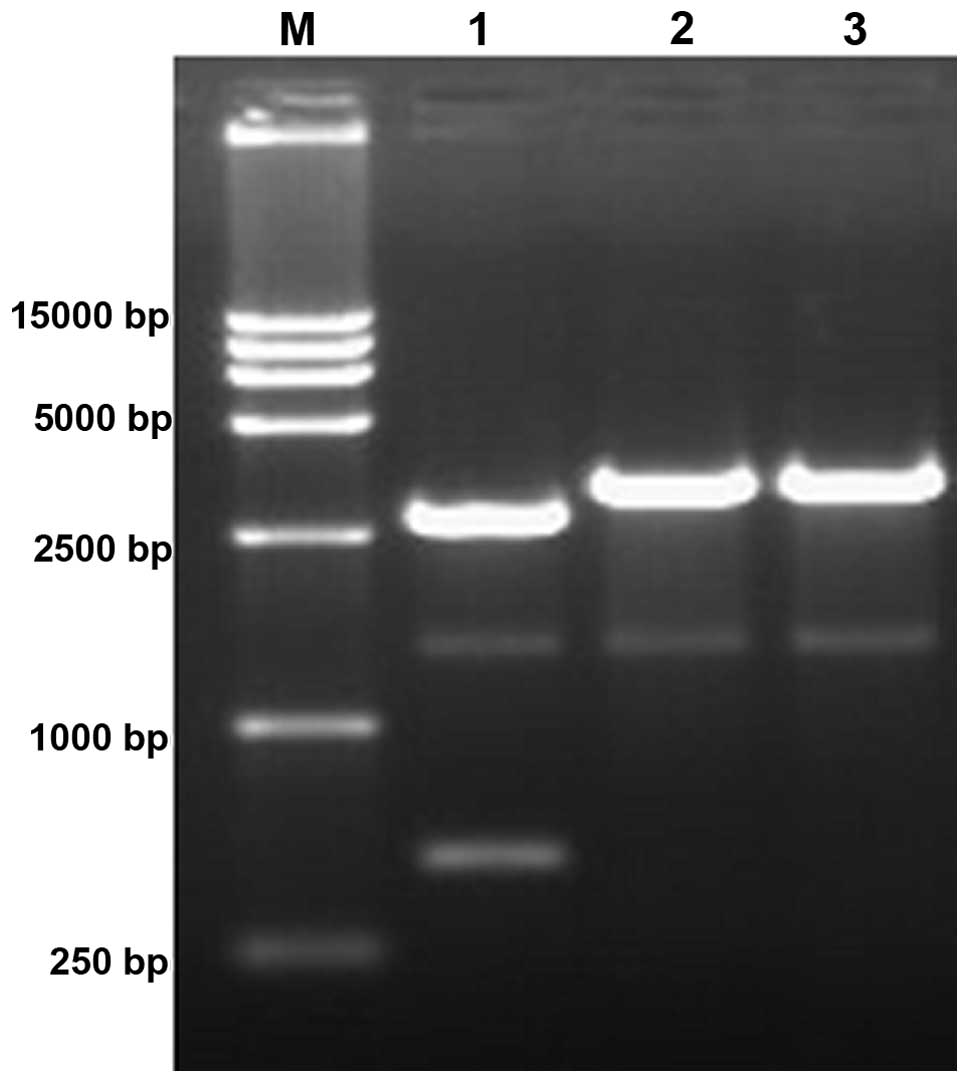
with the pMD18-T Simple and transformed into competent cells, the
plasmid was extracted. When the products were digested with
XhoI and HindIII, two fragments were generated, 2,771
and 482 bp in length. When the PCR product was digested with a
single enzyme, only a 3,253-bp fragment was produced (Fig. 2).
The sequencing results were consistent with the
GenBank database of human HPSE promoter nucleotide sequence
(Accession AF461265′s bl2seq). The 5′ and 3′ ends were located at
556 and 4 bp upstream of the transcription start site,
respectively. With the downstream primer carrying restriction sites
CCAAGCTT (8 bp), the total length was 561 bp. Sequence analysis
revealed that the fragment contained the following TFBSs, as
reported in previous studies (8,9):
three Sp1, four ERE, two EGR-1, one E47, one N-myc and one
NGFI-p300. In addition, there are six potential E-box sites located
at 553-548, 542-537, 402-397, 286-281, 34-29 and 14-9 bp upstream
of the transcription start site (Fig.
3).
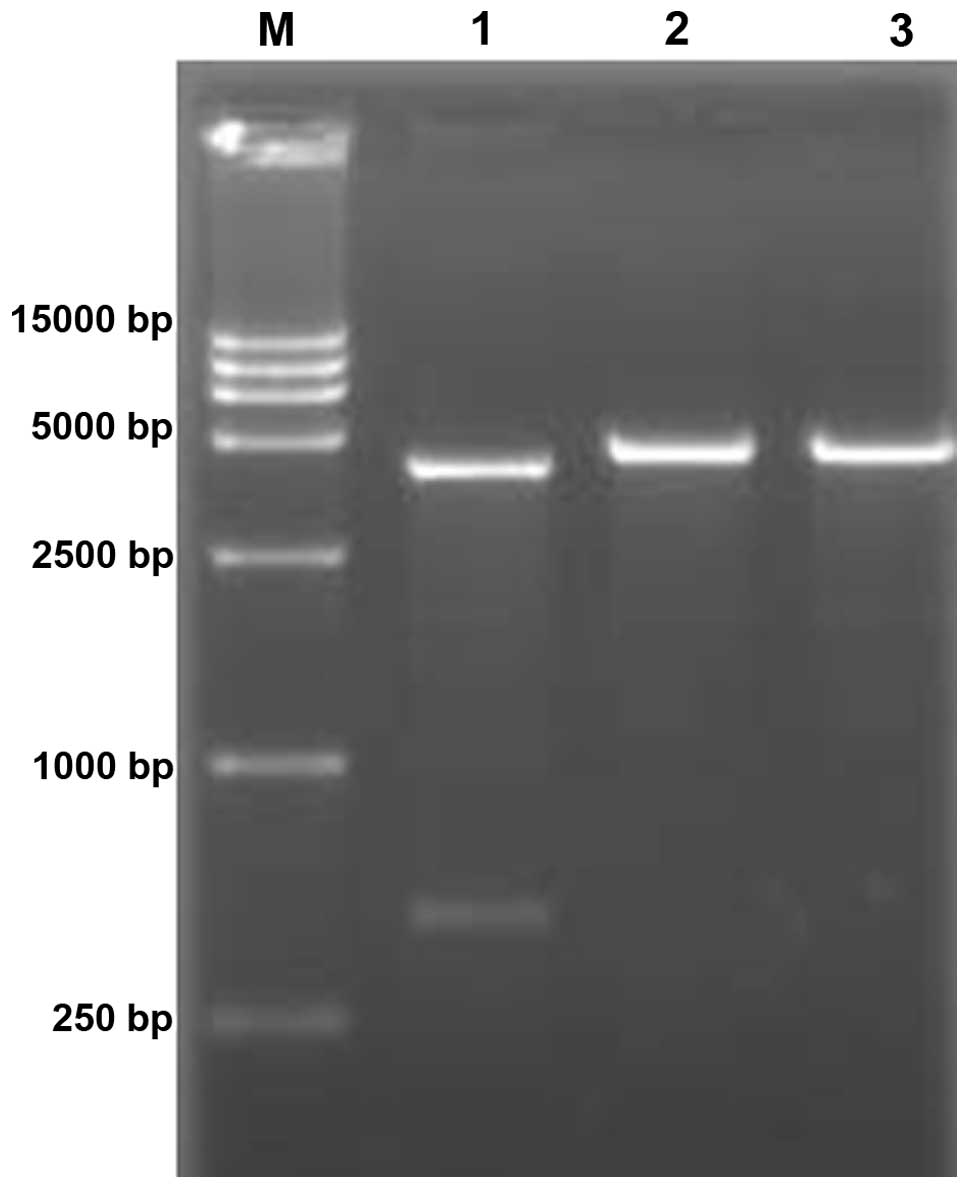
Identification of plasmid pEGFP-Hp
Two fragments were identified following
electrophoresis when the recombinant plasmid pEGFP-Hp was subjected
to double digestion with XhoI and HindIII. Only a
single fragment was observed when the plasmid was digested by
XhoI or HindIII. This proves that the recombinant
plasmid was correctly constructed (Fig. 4). When plasmid pEGFP-Hp was used as
a template, the HPSE promoter fragment was amplified, and the
sequencing results were consistent with the GenBank data.
The activity and specificity of HPSE
promoter
After cells were transfected with three types of
plasmids, the fluorescence analysis showed that there was no
fluorescence in any of the cells from the pEGFP-1 group, but a
strong fluorescence in all the cells from the pEGFP-N1 group. In
the pEGFP-Hp group, almost no fluorescence was detected in ECV304
cells, a degree of fluorescence was observed in all cancer cells
other than ECV304 cancer cells, a strong expression was observed in
the HepG2 and Hep2 cells and a relatively weak fluorescence was
evident in K562 cells (Fig. 5).
Flow cytometric analysis showed that the average transfecion
efficiencies of pEGFP-Hp were 3.9±0.3, 21.3±2.1, 10.8±1.1 and
6.5±0.6% in ECV304, HepG2, Hep2 and K562 cells, respectively. The
transfection efficiencies of pEGFP-Nl were 17.1±1.7, 24.0±2.5,
14.0±1.3 and 11.0±1.0%, respectively, which were all notably higher
than those of pEGFP-Hp. The ratios of transfection efficiency of
the two vectors in ECV304, HepG2, Hep2 and K562 cells were 0.23,
0.89, 0.77 and 0.59, respectively; all <1.
Discussion
The HPSE gene promoter is located in a region 2,300
bp upstream of the ATG translation initiation site. It lacks a TATA
box but contains GC-rich regions. There were three Sp1, four ERE
and two EGR-1 binding sites within the core promoter area (9,10).
In the present study, the HPSE promoter core fragment was cloned.
Sequence analysis showed that it contained the above TFBSs, as
described in the literature (9,10).
In addition, six E-box binding sites were identified, which were
located at 553-548, 542-537, 402-397, 286-281, 34-29, 14-9 bp
upstream of the ATG translation initiation site. These results
suggest that the expression of heparanase gene is likely affected
by the regulation of E-box sites.
One of the key factors of oncogene-targeted therapy
is the selective or specific expression of a therapeutic gene in
tumor cells. The application of tumor-specific promoters is one of
the basic methods. A tumor-specific promoter drives downstream
therapeutic genes that are only expressed in tumor cells, thereby
reducing damage to normal tissue cells (14–16).
At present, commonly used promoters include α-fetoprotein (17), carcinoembryonic antigen (18), prostate-specific antigen (19), human telomerase reverse
transcriptase (20) and multidrug
resistance gene promoters (21).
Tumor-specific promoters play a significant role in targeted gene
therapy by combining with suicide gene, antioncogene, antisense
nucleic acid, apoptosis gene and RNA interference.
Since HPSE mRNA and protein are expressed in the
majority of tumor cells, but are not expressed in normal cells
(1–8), the HPSE promoter may be used to
construct a tumor-specific expression vector to guide the
expression of a therapeutic gene. The vector may reduce or avoid
adverse effects and improve the specificity and safety of gene
therapy. In this study, for the first time, the HPSE promoter
fragment was inserted into the MCS of pEGFP-1 to construct the HPSE
promoter-driven eukaryotic expression vector pEGFP-Hp, in which the
HPSE promoter regulates the transcription and expression of the
downstream GFP gene. After the recombinant vectors were transfected
into normal and tumor cells, fluorescence observation and flow
cytometric analysis showed that pEGFP-Hp does not initiate the
transcription of the downstream GFP gene in normal cells. However,
pEGFP-Hp drove GFP expression in tumor cell lines, which was
consistent with the HPSE mRNA and/or protein expression levels in
these cells (1–8). Since the transcriptional activity of
pEGFP-N1 was strong in all the cells and pEGFP-1 demonstrated no
pEGFP-N1 activity, this suggests that the HPSE promoter within the
recombinant vector pEGFP-Hp regulates the tumor cell-specific
expression of downstream genes. The HPSE promoter was therefore
correlated with HPSE mRNA and/or the protein expression level.
These findings are likely to provide new methods and an
experimental basis for future targeted therapy for tumor-specific
genes. Further experimentation is necessary to determine whether
the HPSE promoter was tumor-specific.
In the same experimental conditions, the activity of
the promoter in the expression vector determines its transfection
efficiency in cells. Transfection efficiency, in turn, reflects the
promoter activity. In this study, the transfection efficiencies of
pEGFP-Hp in the four cell types were all lower than that of
pEGFP-N1. This demonstrates that HPSE promoter activity is weak.
Therefore, the efficiency of the HPSE promoter to drive therapeutic
gene expression in tumor cells may not be as effective as we
expect. Consequently, it exhibits drawbacks as an ideal gene
therapy. Thus, future investigations should be performed to
appropriately modify the HPSE promoter and increase its
activity.
In summary, we successfully amplified the core
fragment of the HPSE promoter and constructed the recombinant
plasmid pEGFP-Hp by inserting it into the plasmid pEGFP-1.
Following cell transfection using pEGFP-Hp, some degree of GFP
expression was observed in tumor cells and no expression was
identified in normal cells. The GFP expression level in tumor cells
was weaker than that in the plasmid pEGFP-N1. These results
demonstrate that the HPSE gene core promoter drives the vector
expression specifically in tumor cell lines, but that its activity
is relatively weak. The results may provide an experimental basis
for tumor-targeting therapy using a tumor-specific promoter.
Studies should be conducted to increase the activity of the HPSE
promoter and evaluate whether the HPSE promoter was
tumor-specific.
Acknowledgements
This study was supported by the Natural Science
foundation of Anhui Province, China (Grant no. 11040606M181) and
the Provincial Natural Science Foundation of College of Anhui
Province, China (Grant no. kj2010B239). We would like to thank
Professor Xue-Long Wang (Anhui Medical University, China) for
technical assistance and Dr Li Gui (Anhui Medical University) for
aid with flow cytometry.
Abbreviations:
|
ECV304
|
human umbilical vein endothelial cell
line
|
|
HepG2
|
hepatoma carcinoma cell line
|
|
Hep2
|
laryngocarcinoma cell line
|
|
K562
|
leukemia cell line
|
References
|
1
|
El-Assal ON, Yamanoi A, Ono T, Kohno H and
Nagasue N: The clinicopathological significance of heparanase and
basic fibroblast growth factor expressions in hepatocellular
carcinoma. Clin Cancer Res. 7:1299–1305. 2001.PubMed/NCBI
|
|
2
|
Chen XP, Liu YB, Rui J, Peng SY, Peng CH,
Zhou ZY, Shi LH, Shen HW and Xu B: Heparanase mRNA expression and
point mutation in hepatocellular carcinoma. World J Gastroenterol.
10:2795–2799. 2004.PubMed/NCBI
|
|
3
|
Komatsu N, Waki M, Sue M, Tokuda C,
Kasaoka T, Nakajima M, Higashi N and Irimura T: Heparanase
expression in B16 melanoma cells and peripheral blood neutrophils
before and after extravasation detected by novel anti-mouse
heparanase monoclonal antibodies. J Immunol Methods. 331:82–93.
2008. View Article : Google Scholar
|
|
4
|
Ilan N, Elkin M and Vlodavsky I:
Regulation, function and clinical significance of heparanase in
cancer metastasis and angiogenesis. Int J Biochem Cell Biol.
38:2018–2039. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yingying X, Yong Z, Zhenning W, Xue Z, Li
J, Yang L and Huimian X: Role of heparanase-1 in gastric carcinoma
invasion. Asian Pac J Cancer Prev. 10:151–154. 2009.PubMed/NCBI
|
|
6
|
Kussie PH, Hulmes JD, Ludwig DL, Patel S,
Navarro EC, Seddon AP, Giorgio NA and Bohlen P: Cloning and
functional expression of a human heparanase gene. Biochem Biophys
Res Commun. 261:183–187. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vlodavsky I, Friedmann Y, Elkin M, Aingorn
H, Atzmon R, Ishai-Michaeli R, Bitan M, Pappo O, Peretz T, Michal
I, et al: Mammalian heparanase: gene cloning, expression and
function in tumor progression and metastasis. Nat Med. 5:793–802.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Toyoshima M and Nakajima M: Human
heparanase. Purification, characterization, cloning and expression.
J Biol Chem. 274:24153–24160. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hulett MD, Freeman C, Hamdorf BJ, Baker
RT, Harris MJ and Parish CR: Cloning of mammalian heparanase, an
important enzyme in tumor invasion and metastasis. Nat Med.
5:803–809. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jiang P, Kumar A, Parrillo JE, Dempsey LA,
Platt JL, Prinz RA and Xu X: Cloning and characterization of the
human heparanase-1 (HPR1) gene promoter: role of GA-binding peotein
and Sp1 in regulating HPR1 BASAL promoter activity. J Biol Chem.
277:8989–8998. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
de Mestre AM, Rao S, Hornby JR, Soe-Htwe
T, Khachigian LM and Hulett MD: Early growth response gene 1 (EGR1)
regulates heparanase gene transcription in tumor cells. J Biol
Chem. 280:35136–35147. 2005.
|
|
12
|
Boyd DD and Nakajima M: Involvement of
heparanase in tumor metastases: A new target in cancer therapy? J
Natl Cancer Inst. 96:1194–1195. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang JM, Wang HJ, Du L, Han XM, Ye ZY,
Fang Y, Tao HQ, Zhao ZS and Zhou YL: Screening and identification
of novel B cell epitopes in human heparanase and their
anti-invasion property for hepatocellular carcinoma. Cancer Immunol
Immunother. 58:1387–1396. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Song MS, Jeong JS, Ban G, Lee JH, Won YS,
Cho KS, Kim IH and Lee SW: Validation of tissue-specific
promoter-driven tumor-targeting trans-splicing ribozyme system as a
multifunctional cancer gene therapy device in vivo. Cancer Gene
Ther. 16:113–125. 2009. View Article : Google Scholar
|
|
15
|
Fukazawa T, Maeda Y, Durbin ML, Nakai T,
Matsuoka J, Tanaka H, Naomoto Y and Tanaka N: Pulmonary
adenocarcinoma-targeted gene therapy by a cancer- and
tissue-specific promoter system. Mol Cancer Ther. 6:244–252. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huyn ST, Burton JB, Sato M, Carey M,
Gambhir SS and Wu L: A potent, imaging adenoviral vector driven by
the cancer-selective mucin-1 promoter that targets breast cancer
metastasis. Clin Cancer Res. 15:3126–3134. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Willhauck MJ, Sharif Samani BR, Klutz K,
Cengic N, Wolf I, Mohr L, Geissler M, Senekowitsch-Schmidtke R,
Göke B, et al: Alpha-fetoprotein promoter-targeted sodium iodide
symporter gene therapy of hepatocellular carcinoma. Gene Ther.
15:214–223. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qiao J, Doubrovin M, Sauter BV, Huang Y,
Guo ZS, Balatoni J, Akhurst T, Blasberg RG, Tjuvajev JG, Chen SH
and Woo SL: Tumor-specific transcriptional targeting of suicide
gene therapy. Gene Ther. 9:168–175. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kraaij R, van der Weel L, de Ridder CM,
van der Korput HA, Zweistra JL, van Rijswijk AL, Bangma CH and
Trapman J: A small chimeric promoter for high prostate-specific
transgene expression from adenoviral vectors. Prostate. 67:829–839.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Murofushi Y, Nagano S, Kamizono J,
Takahashi T, Fujiwara H, Komiya S, Matsuishi T and Kosai K: Cell
cycle-specific changes in hTERT promoter activity in normal and
cancerous cells in adenoviral gene therapy: A promising implication
of telomerase-dependent targeted cancer gene therapy. Int J Oncol.
29:681–688. 2006.
|
|
21
|
Wang X, Ji C, Ma D, Zhao J, Hou M, Yu H
and Zang S: Antitumor effects of cytosine deaminase and thymidine
kinase fusion suicide gene under the control of mdr1 promoter in
mdr1 positive leukemia cells. Leuk Lymphoma. 48:1600–1609. 2007.
View Article : Google Scholar : PubMed/NCBI
|