Introduction
An interferon-like virus inhibitor, later designated
as IFN-γ, was identified in phytohemagglutinin (PHA)-stimulated
human leukocyte cultures (1).
IFN-γ is secreted by NK cells, γδT cells and
CD8+/CD4+ T cells, which undergo activation
by antigens, mitogens or alloantigens. IFN-γ is important in the
control of intracellular infection, immunoregulation, tumor
development, hypersensitivity reactions, and autoimmune diseases
(2,3). Due to its numerous biological and
pathological functions, human IFN-γ (hIFN-γ) is used extensively to
treat drug-resistant tuberculosis (4), atopic dermatitis (5) and hepatitis B infections (6).
Obtaining large quantities of pure hIFN-γ is
important for clinical studies and mechanistic investigations.
Previous studies have reported that recombinant hIFN-γ (rhIFN-γ)
was produced by an adenovirus (7),
Escherichia coli (E. coli) (8–10),
Saccharomyces cerevisiae (11), bicistronic baculovirus insects
(12) and Chinese hamster ovary
(CHO) cells (13,14). However, complex manipulations, high
culture expenses, low yields or low biological activity limit the
use of these expression systems. The methylotrophic yeast,
Pichia pastoris (P. pastoris), is frequently used as
an expression system for the production of proteins. P.
pastoris possesses prokaryotic features including clear genetic
background, easy manipulation, high cell densities and low cost,
and eukaryotic features including protein folding,
post-translational modification and secretion.
In general, the secretion of a protein of interest
into the culture medium is preceded by an N-terminal extension of
3–60 amino acids, forming a signal peptide, which targets the
propeptide of the endoplasmic reticulum followed by cleavage using
membrane bound signal peptidases. hIFN-γ has a native signal
sequence of 23 amino acids, which directs its secretion from human
lymphocyte cells. The Saccharomyces cerevisiae α-mating
factor (MF-α) prepro sequence is widely used in P. pastoris
cells to secrete heterologous proteins. MF-α consists of a signal
sequence of 19 amino acids followed by a 66-residue sequence. The
heterologous protein, fused with the signal sequence, is cleaved by
the KEX2 or STE13 gene product, at the site following
the dibasic paired residues Lys-Arg or Glu-Ala, respectively. In
this study, we compared the secretion level of rhIFN-γ under the
native secretion signal of hIFN-γ versus the MF-α signal peptide,
to achieve a native N-terminal, and a higher quantity of a
biologically active rhIFN-γ.
Materials and methods
Materials
P. pastoris X-33, pPICZaC vector, and Zeocin™
antibiotic were purchased from Invitrogen (Carlsbad, CA, USA).
E. coli XL-Blue cells were purchased from Beijing Dingguo
Biotechnology Company (Beijing, China). Restriction enzymes, DNA
markers and protein markers were purchased from Takara (Dalian,
China). The plasmid extraction kit was purchased from Feyou
Biotechnology Co., Ltd. (Guangzhou, China). Primers and the coding
sequence of hIFN-γ were synthesized by Sangon Biotechnology Co.
(Shanghai, China). The rabbit anti-IFN-γ polyclonal antibody was
purchased from Abcam (Cambridge, UK). Goat anti-rabbit IgG labeled
with HRP was purchased from Beijing Dingguo Biotechnology Company.
Vivaflow 200 polyethersulfone (PES) 50,000 molecular weight
cut-offs (MWCO) and PES 10,000 MWCO and Vivaspin 2 ml were
purchased from Sartorius Stedim Biotech GmbH (Goettingen, Germany).
SP Sepharose XL and Superdex G75 were purchased from GE Healthcare
Bio-Sciences Co. (Piscataway, NJ, USA). Standard hIFN-γ was
purchased from Novoprotein (Shanghai, China). A Bradford protein
assay kit was purchased from Beyotime (Shanghai, China).
Construction of expression vectors
The coding sequence of hIFN-γ, flanked with
BspT1041 and KpnI, was synthesized by Sangon
Biotechnology Co. based on the codon preference in P.
pastoris, and the fragment between the BspT1041 and
KpnI sites was inserted into pPICZαC at the same sites to
generate pNhIFN. The optimized sequence encoding hIFN-γ
flanked with BspT1041 and KpnI was as follows:
TTCGAAACGATGAAGTATACTAGTTAC ATCTTAGCCTTTCAATTGTGCATTGTTCTTGGTTC
TTTGGGATGTTATTGTCAAGATCCATACGTTAAGGA
GGCCGAGAACCTAAAGAAATACTTTAACGCCGGCCA
TGACGTGGCAGATAATGGGACTTTCTCGTTTTTGGG
TATTCTGAAAAATTGGAAGGAAGAGTCCGATAGAAA
GATTATGCAATCCCAGATCGTAAGTTTCTACTTCAAG
CTGTTTAAAAATTTCAAGGACGATCAGTCTATACAA
AAATCAGTGGAGACCATTAAAGAAGACATGAACGTC
AAGTTCTTCAACTCTAACAAAAAGAAGAGAGATGAT
TTTGAAAAACTAACCAATTATTCTGTTACAGACCTGA
ATGTTCAGAGAAAAGCTATACATGAATTAATCCAGG
TCATGGCTGAACTTTCACCTGCTGCTAAAACGGGTA
AACGAAAGAGATCACAAATGTTGTTTCGTGGAAGGA GAGCATCCCAATAAGGTACC. The
recognition sequences for restriction enzymes BspT1041 and
KpnI are underlined, and the bold shows the native secretion
signal peptide of hIFN-γ.
To construct a plasmid containing the hIFN-γ
gene with an MF-α signal sequence containing a Kex2 cleavage site
(Lys-Arg), primers containing XhoI and XbaI
recognition sites were used. In order to create a Kex2 cleavage
site upstream of the coding sequence of the mature hIFN-γ, bases
coding for Lys-Arg were incorporated in the forward primer. Forward
primer, 5′-GCCTCGAGAAGAGACAAGATCCATACG-3′; the recognition
site for XhoI is underlined, and the bold letters signify
the encoded region of the MF leader peptide of the Kex2 cleavage
site (Lys-Arg). Reverse primer,
5′-CCGTCTAGATTATTGGGATGCTCTCC-3′; the recognition site for
XbaI is underlined, and the stop codon is in bold.
The polymerase chain reaction (PCR) was performed
using the vector pNhIFN as a template, using 33 cycles of 94ºC for
30 sec, 55ºC for 40 sec and a final extension of 5 min at 72ºC. The
product was digested with XhoI and XbaI and ligated
into pPICZαC at the same sites, generating pαIFN. pNhIFN and pαIFN
were verified by DNA sequencing.
Vector transformation into P. pastoris
and screening with genomic PCR
pNhIFN and pαIFN were isolated using an Omega
Bio-Tek plasmid mini kit (Norcross, GA, USA), linearized with
SacI and transformed into the X-33 strain of P.
pastoris according to the transformation protocol (Invitrogen).
The transformed cells were selected on YPD (yeast extract, peptone
and dextrose) containing Zeocin (100 μg/ml). Zeocin-resistant
clones were isolated and the integration of hIFN-γ was
confirmed by PCR using AOX1 universal primers: forward,
5′-GACTGGTTCCAATTGACAAGC-3′ and reverse,
5′-GCAAATGGCATTCTGACATCC-3′. Thirty amplification cycles were
performed at 94ºC for 30 sec, 55ºC for 30 sec, and a final
extension for 5 min at 72ºC.
Detection of the insertion copy
number
It has been reported that multiple copies of the
transgene may integrate following electroporation and that the gene
dosage may affect the expression levels of the recombinant protein
(15). Therefore, in order to
achieve a more physiologically comparable result, quantitative PCR
(qPCR) was used to determine the copy number of the transgenes in
the positive transformants. The hIFN-γ insertion copy number
was examined three times for each positive clone. Data were
normalized against the ACT1 gene as an endogenous control
(housekeeping gene). The primers used were: hIFN-γ, forward:
5′-AGGAAGAGTCCGATAGAAAG-3′ and reverse: 5′-TCAGCCATGACCTGGATTAA-3′;
ACT1, forward: 5′-TCATGGTCGGTATGGGTCAA-3′ and reverse:
5′-ACGATACCGTGCTCGATTGG-3′.
Genomic DNA was used as the template for qPCR
analysis. Reaction conditions were established according to the
SYBR® Premix Ex Taq™ II manual (Takara). Each 20-μl
reaction contained 10 μl SYBR Premix Ex Taq II (2×), 10 μM forward
and reverse primers (1 μl each), 2 μl sample genomic DNA and 6 μl
sterile double deionized water. qPCR reactions were run in
triplicate on ABI7000 (Applied Biosystems, Foster City, CA, USA)
under the following conditions: 95ºC for 30 sec, followed by 40
cycles of 95ºC for 5 sec and 60ºC for 30 sec. The amplicon
specificity was verified by analyzing the melting curve after 40
cycles and by agarose gel electrophoresis.
Expression of hIFN-γ in P. pastoris
Single copy clones were placed in 10 ml buffered
glycerol-complex (BMGY) medium (1.0% yeast extract, 2.0% peptone,
1.34% yeast nitrogen base, 4×10−5 biotin, 100 mM
potassium phosphate, pH 6.0, and 1.0% glycerol) and incubated at
30ºC for 24 h. Subsequently, the cells were harvested by
centrifugation for 5 min at 1500 × g and resuspended in 10 ml
buffered methanol-complex (BMMY) medium (BMGY medium with 0.5%
methanol instead of 1.0% glycerol). Methanol was added to a final
concentration of 0.5% (v/v) every 24 h. To detect the expression of
rhIFN-γ in the supernatant, 12% (w/v) Tricine-SDS-PAGE, western
blot analysis and amino-terminal sequencing were performed.
Tricine-SDS-PAGE analysis was performed using a 12% gel according
to the protocol proposed by Schägger (16).
For western blot analysis, proteins were
transblotted onto a polyvinylidene fluoride (PVDF) membrane. The
membrane was blocked using 5% BSA in Tris-buffered saline with
Tween (TBST) (150 mM NaCl, 25 mM Tris, and 0.05% Tween-20, pH 7.5)
for 2 h at room temperature, and subsequently incubated with mouse
anti-HSA monoclonal antibody at 4ºC overnight. After being washed
three times with TBST, the membrane was incubated with goat
anti-mouse IgG conjugated to horseradish peroxidase (HRP) (Beijing
Dingguo Biotechnology Company) for 2 h at room temperature.
Finally, the membrane was washed with TBST and proteins were
detected with 3,3′-diaminobenzidine (DAB).
The amino-terminal sequence of rhIFN-γ was
determined using automated Edman degradation performed on a model
PPSQ-21A protein sequencer (Shimadzu, Japan).
Purification of rhIFN-γ
To purify rhIFN-γ from the culture supernatant, 5
liters of P. pastoris culture were centrifuged at 1,500 × g
for 15 min. As the molecular weight of rhIFN-γ is 17 kDa, proteins
in the supernatant with a molecular weight of 10–50 kDa were
isolated and concentrated using Vivaflow 200 PES 50,000 MWCO and
PES 10,000 MWCO. The concentrated sample was loaded onto the SP
Sepharose XL for further separation, as the isoelectric point of
hIFN-γ is ~8.1–9.1. The rhIFN-γ fraction was concentrated by
Vivaspin 2 ml. Fractions of each step were analyzed on 15%
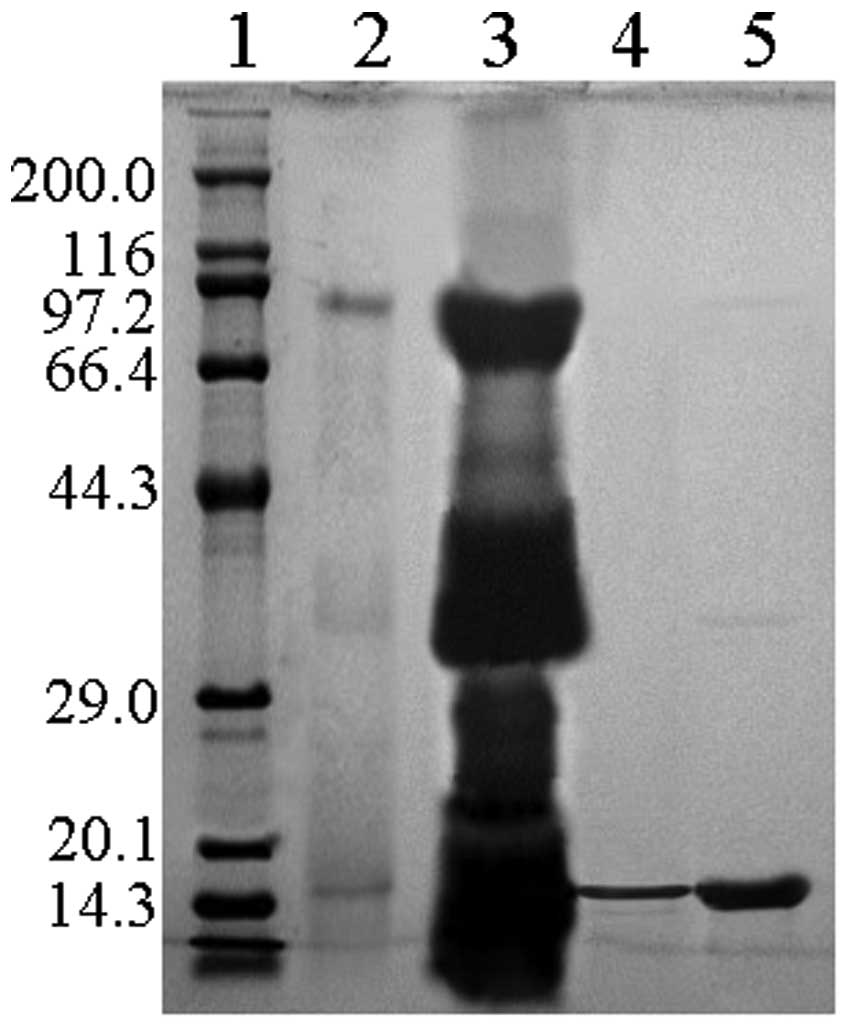
Tricine-SDS-PAGE. The rhIFN-γ fraction was stored at −20ºC. Protein
concentration was determined using a Bradford protein assay kit.
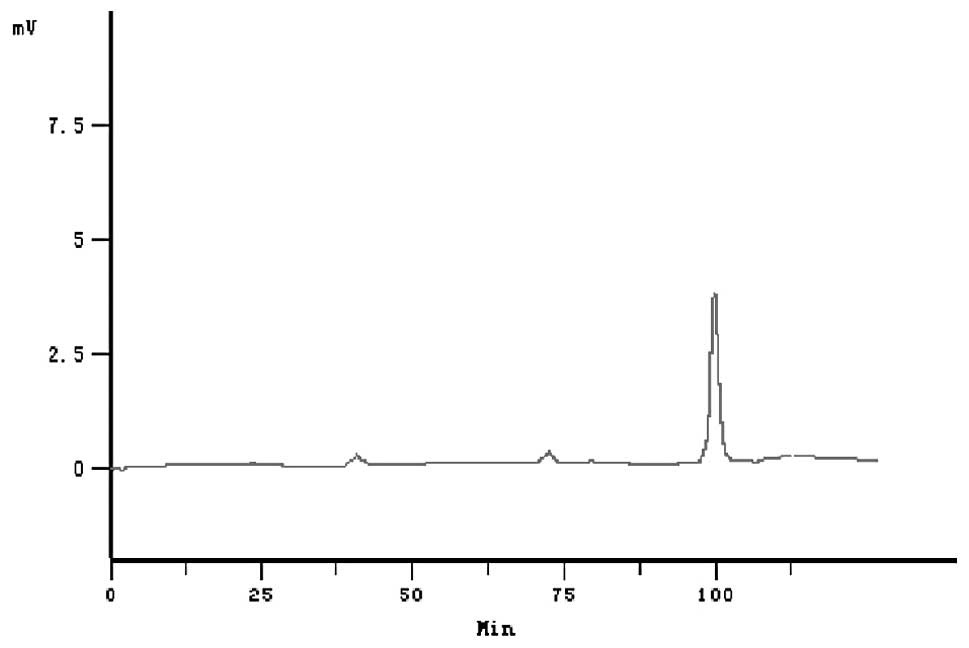
The purity of rhIFN-γ was determined by HPLC analysis.
Activity assay of hIFN-γ
The antiviral activity of rhIFN-γ was evaluated as
previously described (17). The
human carcinoma cell line, HEP2C, was challenged with the
encephalomyocarditis virus (EMCV) in the presence or absence of
hIFN-γ. Antiviral activity was calculated in comparison with the
standard obtained from Novoprotein.
Results
Construction of expression plasmids and
transformation of P. pastoris
The 516-bp gene fragment encoding hIFN-γ in addition
to the BspT1041 and KpnI sites was synthesized by
Sangon Biotechnology Co. and subcloned into pPICZαC in order to
generate the recombinant plasmid, pNhIFN. The 451-bp gene fragment,
encoding the mature hIFN-γ in addition to the XhoI and
XbaI sites, was amplified by PCR from pNhIFN and subcloned
into pPICZαC to generate the recombinant plasmid, pαIFN. Nucleotide
sequencing analysis confirmed that the plasmids contained the
correct hIFN-γ gene (data not shown).
In order to integrate hIFN-γ into the genome
of P. pastoris by homologous recombination, the plasmids
were linearized with SacI and transformed into
electrocompetent P. pastoris cells. Successful integration
was confirmed by PCR using AOX1 universal primers. The PCR results
revealed that in positive yeast transformants there was one band of
~787 bp (pNhIFN) or 948 bp (pαIFN). The cells transformed with a
pPICZαC empty plasmid were used as a negative control, and in these
cells one band of ~588 bp was identified. qPCR data indicated that
90% of transformants contained a single copy of the gene, with only
single-copy integrants being selected for the comparison
experiments.
Expression and detection of rhIFN-γ in P.
pastoris
Single-copy clones of each strain were inoculated in
BMGY medium for 24 h and induced in BMMY medium for the expression
of rhIFN-γ. Each strain was analyzed for rhIFN-γ expression on
Tricine-SDS-PAGE following 96 h of induction. Fig. 1A shows a Coomassie-stained
Tricine-SDS-PAGE of supernatants collected from induced P.
pastoris cultures transformed with the expression plasmids of
the pPICZαC control, pNhIFN or pαIFN. The supernatant of the
control strain (lane 3) did not reveal any band corresponding to
the standard rhIFN-γ (lane 2). The supernatant of the P.
pastoris strain transformed with pNhIFN (lanes 4 and 5),
containing the hIFN-γ gene with the native secretion signal
peptide, did not reveal a rhIFN-γ protein band. The supernatant of
the P. pastoris strain transformed with pαIFN (lanes 6 and
7), containing the hIFN-γ gene with the MF-α signal peptide,
showed one protein band that was slightly higher than the hIFN-γ
band.
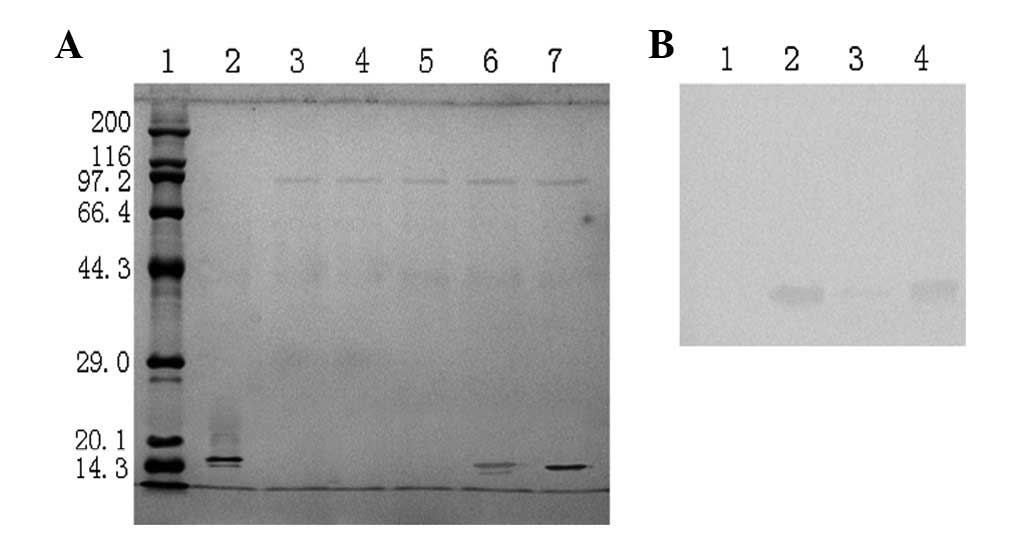 | Figure 1(A) Coomassie-stained Tricine-SDS-PAGE
profile of Pichia pastoris (P. pastoris) clones expressing
recombinant human interferon-γ (rhIFN-γ). Lane 1, broad molecular
weight protein marker (6.5–200 kDa); lane 2, standard human
interferon-γ (hIFN-γ); lane 3, supernatant of the P.
pastoris strain transformed with pPICZαC; lanes 4 and 5,
supernatant of the P. pastoris strain transformed with
pNhIFN, containing the IFN-γ gene with the native secretion
signal peptide; lanes 6 and 7, supernatant of the P.
pastoris strain transformed with pαIFN, containing the
IFN-γ gene with the MF-α signal peptide. (B) Western blot
analysis of the recombinant P. pastoris supernatant
expressing hIFN-γ. Lane 1, supernatant of the P. pastoris
strain transformed with pPICZαC; lane 2, standard hIFN-γ; lane 3,
supernatant of the P. pastoris strain transformed with
pNhIFN; lane 4, supernatant of the P. pastoris strain
transformed with pαIFN. The Coomassie staining profile of the P.
pastoris strain transformed with pNhIFN did not present a band
at 15 kDa, but a faint band was detected by western blotting. |
Western blot analysis using an anti-human IFN-γ
antibody confirmed that the protein band from the supernatant of
the pαIFN-transformed strain was hIFN-γ (Fig. 1B, lane 4). However, it showed a
faint band in the supernatant of the pNhIFN-transformed P.
pastoris strain (Fig. 1B, lane
3). These results suggest that the native hIFN-γ secretion signal
peptide is capable of secreting particularly low levels of hIFN-γ
that may only be detected by western blot analysis, as it is more
sensitive than Coomassie staining Tricine-SDS-PAGE.
Therefore, only the MF-α signal peptide cleavage
site of the recombinant rhIFN-γ was examined, and N-terminal
sequencing revealed that the first 14 amino acids of rhIFN-γ were
QDPYVKEAENLKKY, which were identical to the N-terminal sequence of
hIFN-γ, thus confirming the successful expression of the
protein.
Purification and biological activities of
rhIFN-γ in P. pastoris
Following the isolation and concentration of rhIFN-γ
from the P. pastoris cultures, rhIFN-γ was then purified
with cation exchange chromatography (SP Sepharose XL), using an
AKTA Explorer 100 chromatography system. The optimal pH of the 50
mM acetate buffer for binding was 5.0 and the optimal NaCl (Beijing
Dingguo Biotechnology Company) concentration for elution was 0.4 M.
The rhIFN-γ fraction was concentrated by Vivaspin 2 ml. Following
these procedures, from 5 liters of culture medium, we obtained a
total of 800 mg of purified rhIFN-γ. rhIFN-γ achieved a purity of
96.7% as revealed by Tricine-SDS-PAGE and HPLC (Figs. 2 and 3). Protein recovery and purity of rhIFN-γ
at the various purification steps are shown in Table I.
 | Table ISummary of purification steps of
recombinant human interferon-γ (rhIFN-γ) from Pichia
pastoris. |
Table I
Summary of purification steps of
recombinant human interferon-γ (rhIFN-γ) from Pichia
pastoris.
| Purification
steps | Volume (ml) | Total protein
(mg) | rhIFN-γ (mg) | Recovery (%) | Purity (%) |
|---|
| Supernant | 5000 | 4500 | 1500 | | 33 |
| Vivaflow 200 | 100 | 2415 | 1208 | 80.5 | 50 |
| SP Sepharose
XL | 50 | 751 | 676 | 56 | 90 |
| Vivaspin 2 ml | 44 | 601 | 574.6 | 85 | 95.6 |
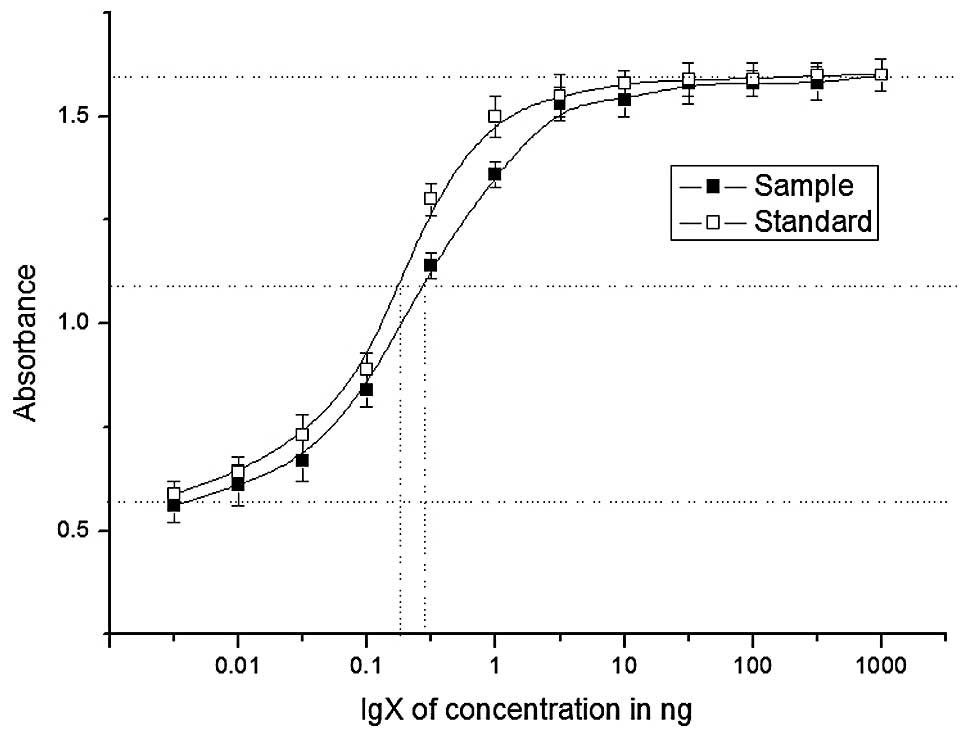
To determine the antiviral effects of purified
rhIFN-γ, an antiviral assay was performed by challenging HEP2C
cells with EMCV in the presence of varying concentrations of
rhIFN-γ. In three determinations, the calculated specific activity
of rhIFN-γ ranged from 1×107 to 1.6×107 IU/mg
protein, as assessed against the standard (Fig. 4). The results demonstrated that
rhIFN-γ produced by P. pastoris had lower levels of
biological activity compared with the standard obtained from
Novoprotein, however, the simpler purification strategy makes this
a useful method.
Discussion
hIFN-γ is a cytokine with multiple biological and
pharmaceutical functions (2,18),
thus it is expressed in a variety of systems, particularly in E.
coli and CHO cells. The disadvantage of E. coli is that
the accumulation of rhIFN-γ forms inactive and insoluble aggregates
known as inclusion bodies. Although a variety of techniques have
been developed to produce active recombinant proteins (dilution,
diafiltration, urea, combined urea-ammonium chloride, chaperones,
detergent-assisted and chromatography refolding), the complex
procedures may lead to a low yield of the active product (10,19–21).
Furthermore, rhIFN-γ expressed in E. coli is different from
native mature hIFN-γ, as its N-terminal amino acid is methionine,
which may have clinical and pharmaceutical side effects. The
disadvantage of the CHO cell system is the low productivity of
rhIFN-γ. Although culturing recombinant CHO cells at 32ºC may
increase the productivity of rhIFN-γ, it is <10 mg/l. In our
study, we used the P. pastoris system to express rhIFN-γ due
to of the following features: low cost, high productivity, a strong
regulated AOX1 promoter, growth to a high cell density in a simple
medium and stable integration of heterogeneous genes into the
genome. Furthermore, P. pastoris secretes few endogenous
proteins to the culture medium, making purification of recombinant
proteins easier.
In the P. pastoris expression system, the
secretion signal peptide is essential for the successful expression
of heterologous proteins. For IFN-α2b expression, the native
secretion signal of IFN-α2b did not secrete the protein, but the
α-prepro sequence without the EAEA repeats secreted 200 mg/l
IFN-α2b into the culture medium (22). Expression of human serum albumin,
using its native signal peptide, has been reported to produce >1
g/l (23). For the secretory
efficiency of recombinant xylanase xynB, the bovine β-casein signal
peptide was less efficient than the α-factor prepro sequence
(24). Polygalacturonase genes
were expressed in P. pastoris with the native signal peptide
or the α-factor secretion signal peptide, but only slight
differences in expression were observed (25). As signal peptides secrete
recombinant proteins with varying efficiencies, it is important to
examine different signal peptides that secrete the same protein in
P. pastoris. In this study, hIFN-γ was successfully
expressed in P. pastoris using either the native signal
peptide of hIFN-γ or the α-factor signal peptide, although the
former mediated trace levels of rhIFN-γ secretion, while the latter
mediated secretion of 300 mg/l rhIFN-γ into the culture medium with
the same amino terminal sequence as that of the native protein.
The biologically active form of hIFN-γ is a
homodimer that interacts with a heterodimeric receptor consisting
of INF-γ receptor 1 and 2 (26,27).
In our study, the activity of rhIFN-γ was only slightly lower than
that of the hIFN-γ standard, suggesting that the majority of the
rhIFN-γ dimerized following the correct secretion into the culture,
while small quantities of the rhIFN-γ may have remained in the
monomer form or formed other complex structures. In conclusion,
results of this study demonstrate that large quantities of
biologically active rhIFN-γ may be produced from P.
pastoris.
Acknowledgements
The authors would like to thank Jilin Department of
Health, Jilin, China, for their financial support (no.
2011I087).
References
|
1
|
Wheelock EF: Interferon-like
virus-inhibitor induced in human leukocytes by phytohemagglutinin.
Science. 149:310–311. 1965. View Article : Google Scholar
|
|
2
|
Chen J and Liu X: The role of interferon
gamma in regulation of CD4+ T-cells and its clinical
implications. Cell Immunol. 254:85–90. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Billiau A and Matthys P: Interferon-gamma:
a historical perspective. Cytokine Growth Factor Rev. 20:97–113.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Suárez-Méndez R, García-García I,
Fernández-Olivera N, Valdés-Quintana M, Milanés-Virelles MT,
Carbonell D, Machado-Molina D, Valenzuela-Silva CM and López-Saura
PA: Adjuvant interferon gamma in patients with drug-resistant
pulmonary tuberculosis: a pilot study. BMC Infect Dis.
4:442004.PubMed/NCBI
|
|
5
|
Chang TT and Stevens SR: Atopic
dermatitis: the role of recombinant interferon-gamma therapy. Am J
Clin Dermatol. 3:175–183. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Parvez MK, Sehgal D, Sarin SK, Basir SF
and Jameel S: Inhibition of hepatitis B virus DNA replicative
intermediate forms by recombinant interferon-gamma. World J
Gastroenterol. 12:3006–3014. 2006.PubMed/NCBI
|
|
7
|
Xu R, Ying B, Zhao S, Li C and Wang Y:
Construction and identification of a recombinant adenovirus which
expresses human interferon-gamma. Chin J Biotechnol. 13:1–8.
1997.PubMed/NCBI
|
|
8
|
Arakawa T, Hsu YR and Yphantis DA: Acid
unfolding and self-association of recombinant Escherichia
coli derived human interferon gamma. Biochemistry.
26:5428–5432. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Z, Tong KT, Belew M, Pettersson T
and Janson JC: Production, purification and characterization of
recombinant human interferon gamma. J Chromatogr. 604:143–155.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mohammadian-Mosaabadi J, Naderi-Manesh H,
Maghsoudi N, Nassiri-Khalili MA, Masoumian MR and Malek-Sabet N:
Improving purification of recombinant human interferon gamma
expressed in Escherichia coli; effect of removal of impurity
on the process yield. Protein Expr Purif. 51:147–156. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fieschko JC, Egan KM, Ritch T, Koski RA,
Jones M and Bitter GA: Controlled expression and purification of
human immune interferon from high-cell-density fermentations of
Saccharomyces cerevisiae. Biotechnol Bioeng. 29:1113–1121.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen YJ, Chen WS and Wu TY: Development of
a bi-cistronic baculovirus expression vector by the
Rhopalosiphum padi virus 5′ internal ribosome entry site.
Biochem Biophys Res Commun. 335:616–623. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Devos R, Opsomer C, Scahill SJ, Van der
Heyden J and Fiers W: Purification of recombinant glycosylated
human gamma interferon expressed in transformed Chinese hamster
ovary cells. J Interferon Res. 4:461–468. 1984. View Article : Google Scholar
|
|
14
|
Tan HK, Lee MM, Yap MG and Wang DI:
Overexpression of cold-inducible RNA-binding protein increases
interferon-gamma production in Chinese-hamster ovary cells.
Biotechnol Appl Biochem. 49:247–257. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li K, Gao H, Gao L, Qi X, Gao Y, Qin L,
Wang Y and Wang X: Recombinant gp90 protein expressed in Pichia
pastoris induces a protective immune response against
reticuloendotheliosis virus in chickens. Vaccine. 30:2273–2281.
2012.PubMed/NCBI
|
|
16
|
Schägger H: Tricine-SDS-PAGE. Nat Protoc.
1:16–22. 2006.
|
|
17
|
Reddy PK, Reddy SG, Narala VR, Majee SS,
Konda S, Gunwar S and Reddy RC: Increased yield of high purity
recombinant human interferon-gamma utilizing reversed phase column
chromatography. Protein Expr Purif. 52:123–130. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zaidi MR and Merlino G: The two faces of
interferon-gamma in cancer. Clin Cancer Res. 17:6118–6124. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yan X, Hu S, Guan YX and Yao SJ:
Coexpression of chaperonin GroEL/GroES markedly enhanced soluble
and functional expression of recombinant human interferon-gamma in
Escherichia coli. Appl Microbiol Biotechnol. 93:1065–1074.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Petrov S, Nacheva G and Ivanov I:
Purification and refolding of recombinant human interferon-gamma in
urea-ammonium chloride solution. Protein Expr Purif. 73:70–73.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jin T, Guan YX, Yao SJ, Lin DQ and Cho MG:
On-column refolding of recombinant human interferon-gamma inclusion
bodies by expanded bed adsorption chromatography. Biotechnol
Bioeng. 93:755–760. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ghosalkar A, Sahai V and Srivastava A:
Secretory expression of interferon-alpha 2b in recombinant
Pichia pastoris using three different secretion signals.
Protein Expr Purif. 60:103–109. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kobayashi K, Kuwae S, Ohya T, Ohda T,
Ohyama M, Ohi H, Tomomitsu K and Ohmura T: High-level expression of
recombinant human serum albumin from the methylotrophic yeast
Pichia pastoris with minimal protease production and
activation. J Biosci Bioeng. 89:55–61. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
He Z, Huang Y, Qin Y, Liu Z, Mo D, Cong P
and Chen Y: Comparison of alpha-factor preprosequence and a
classical mammalian signal peptide for secretion of recombinant
xylanase xynB from yeast Pichia pastoris. J Microbiol
Biotechnol. 22:479–483. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cho IJ, Yeo IC, Lee NK, Jung SH and Hahm
YT: Heterologous expression of polygalacturonase genes isolated
from Galactomyces citri-aurantii IJ-1 in Pichia
pastoris. J Microbiol. 50:332–340. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ealick SE, Cook WJ, Vijay-Kumar S, Carson
M, Nagabhushan TL, Trotta PP and Bugg CE: Three-dimensional
structure of recombinant human interferon-gamma. Science.
252:698–702. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sadir R, Forest E and Lortat-Jacob H: The
heparan sulfate binding sequence of interferon-gamma increased the
on rate of the interferon-gamma-interferon-gamma receptor complex
formation. J Biol Chem. 273:10919–10925. 1998. View Article : Google Scholar
|