Introduction
According to Cancer Statistics In Japan 2012, it was
estimated that ~357,000 people died from cancer in 2011 in Japan
(1). The treatment strategies for
cancer involve surgical treatment, chemotherapy/molecular targeted
therapy and radiotherapy; however, cancer cells acquire resistance
when treatment is prolonged and hypoxic conditions serve a role in
this acquired resistance (2).
Furthermore, hypoxia has been previously reported to be involved in
metastasis and recurrence of cancer (3–10).
Therefore, hypoxia has become a key target of cancer therapy.
The transcription factor hypoxia-inducible factor-1α
(HIF-1α) is accumulated in tumor cells under hypoxic conditions and
is involved in the acquired resistance towards cancer therapy and
adaptation to hypoxia (2). HIF-1α
moves into the nucleus and promotes the expression of numerous
genes involved in angiogenesis, cell proliferation, glucose
metabolism and apoptosis (2).
Furthermore, the intracellular accumulation of HIF-1α inhibits the
production of reactive oxygen species (ROS) induced by hypoxic
stress, and HIF-1α serves an important part in the adaptation of
cells to hypoxia (11,12). In the presence of oxygen, HIF-1α is
degraded by the ubiquitin-proteasome system subsequent to
hydroxylation by the von Hippel-Lindau (VHL) protein. Conversely,
non-hydroxylated HIF-1α enters the nucleus to form a heterodimer
with the constitutively expressed HIF-1β (13). The HIF-1 heterodimer binds to the
hypoxia-response element (HRE), thereby activating the expression
of numerous hypoxia-response genes, including the pro-angiogenic
growth factor vascular endothelial growth factor (VEGF) (13).
Various types of small molecule inhibitors of HIF-1
have been developed and studied previously (14–16).
One example, LW6, has been reported to upregulate the VHL protein
(17). As a result, the
transcriptional activity of the hypoxia-response genes is
downregulated due to the marked degradation of HIF-1α. LW6 is
therefore hypothesized to improve resistance to cancer therapy in
hypoxia. Previous studies have demonstrated that LW6 exerts marked
anti-tumor efficacy in vivo and causes reductions in HIF-1α
expression levels in mice carrying xeno-grafts of HCT116 cells
(17). However, it is not clear
whether the difference of anti-tumor efficacy is associated with
the oxygen levels. The aim of the present study was to investigate
whether LW6 enhances cytotoxicity selectively in hypoxic cells
through depolarization of the mitochondrial membrane potential
(MMP). These results suggested that agents which are able to
depolarize the MMP, such as LW6, may represent a novel therapeutic
strategy to be used on hypoxic cells that survive other cancer
therapies.
Materials and methods
Materials
Dulbecco's modified Eagle's medium (DMEM) was
obtained from Sigma-Aldrich (St. Louis, MO, USA). Penicillin and
streptomycin were obtained from Gibco-BRL (Invitrogen Life
Technologies, Carlsbad, CA, USA) and fetal bovine serum (FBS) was
obtained from GE Healthcare (Little Chalfont, UK). LW6 was
purchased from Merck Millipore (Darmstadt, Germany) and diluted in
dimethyl sulfoxide (DMSO; Wako Pure Chemical Industries, Ltd.,
Osaka, Japan). Mouse monoclonal anti-HIF-1α antibody (ab1) was
obtained from Abcam (Cambridge, UK) and goat polyclonal anti-actin
antibody (sc-1615) was obtained from Santa Cruz Biotechnology
(Dallas, TX, USA).
Cell culture and growth conditions
The human lung adeno-carcinoma cell line A549 was
grown in DMEM supplemented with penicillin, streptomycin and 10%
heat-inactivated FBS at 37°C in a humidified atmosphere containing
5% CO2. Hypoxia was defined as 1% oxygen, which was
achieved by culturing cells in modular incubator chambers
(Billups-Rothenberg, Inc., Del Mar, CA, USA), which were flushed
with gas mixtures (95% nitrogen/5% carbon dioxide) and sealed to
maintain hypoxia.
Cells were seeded into 35-mm dishes (Iwaki, Chiba,
Japan) at 2×105 cells/dish with 1.5 ml medium containing
LW6 for 12 h. Cells were incubated under normoxia or hypoxia for 36
h and were then assessed for the expression of HIF-1α and the ratio
of apoptotic cells. To analyze active caspase-3, the cells treated
with LW6 for 12 h were exposed to hypoxia or normoxia for 48 h and
the cells were then analyzed.
Cell viability analysis
Cells were incubated in 96-well ELISA Plates (Iwaki)
with 100 μl culture medium at 2×105 cells/ml with
or without LW6 for 24 h. Cell viability was assessed by the
dimethyl thiazolcarboxy-methoxyphenylsulfophenyltetrazolium (MTS)
assay performed using a CellTiter 96® AQueous One
Solution Cell Proliferation Assay kit (Promega Corporation,
Madison, WI, USA) according to the manufacturer's instructions. The
absorbance was measured at 490 and 620 nm using a microplate reader
(iMark™; Bio-Rad Laboratories, Inc., Hercules, CA, USA). For the
trypan blue dye. exclusion test, cells were stained by
phosphate-buffered saline (PBS) containing 0.1% trypan blue
(Nacalai Tesque, Inc., Kyoto, Japan). Cell viability was assessed
by counting the number of unstained cells using the TC20™ automated
cell counter (Bio-Rad Laboratories, Inc.).
Western blot analysis
Cells were lysed using Cell Lysis Buffer (Cell
Signaling Technology, Inc., Danvers, MA, USA) and
phenylmethanesulfonylfluoride (Sigma-Aldrich). Cell lysates and
pre-stained molecular weight markers were separated by SDS-PAGE
with 12% Mini-PROTEAN® TGX™ precast gels (Bio-Rad
Laboratories, Inc.), followed by transfer onto polyvinylidene
fluoride membranes with Trans-Blot® Turbo™ (Bio-Rad
Laboratories, Inc.). The membranes were blocked in Tris-buffered
saline with 0.1% Tween-20 containing 5% milk and then incubated
with various primary antibodies diluted in blocking buffer. Mouse
monoclonal anti-HIF-1a antibody (cat. no. 610959; BD Biosciences)
at a dilution of 1:200, rabbit monoclonal anti-histone H1 antibody
(cat. no. ab125027; Abcam) at a dilution of 1:200, goat polyclonal
anti-actin antibody (cat. no. sc-1615; Santa Cruz Biotechnology) at
a dilution of 1:1,000, rabbit monoclonal anti-VEGF antibody (cat.
no. ab52917; Abcam) at a dilution of 1:200, rabbit polyclonal
anti-VHL antibody (cat. no. sc-5575; Santa Cruz Biotechnology) at a
dilution of 1:200 were incubated for 1 h. The blots were then
washed with Tris-buffered saline containing 0.1% Tween-20 (Bio-Rad
Laboratories, Inc.) three times and incubated with horseradish
peroxidase-conjugated donkey anti-mouse IgG (cat. no. sc-2314;
Santa Cruz Biotechnology) at a dilution of 1:5,000, donkey
anti-rabbit IgG (cat. no. sc-2313; Santa Cruz Biotechnology) at a
dilution of 1:5,000, or donkey anti-goat IgG (cat. no. sc-2056;
Santa Cruz Biotechnology) at a dilution of 1:5,000 in blocking
buffer for 1 h. Membranes were washed three times and
immunoreactivity was visualized using a chemiluminescence Molecular
Imager® ChemiDoc™ XRS+ system (Bio-Rad Laboratories,
Inc.) according to the manufacturer's instructions.
Detection of HIF-1α
HIF-1α was detected using the fluorescein
isothiocyanate (FITC)-conjugated monoclonal active HIF-1α antibody
(ab1; Abcam) according to the manufacturer's instructions. Briefly,
cells were collected with 0.1% trypsin under hypoxia. Following
incubation on ice for 20 min, the cells were washed with ice-cold
PBS and suspended in Cytofix/Cytoperm™ solution (BD Biosciences).
Subsequently, following incubation on ice for 20 min, the cells
were centrifuged for 3 min at 200 × g, pelletted and the
supernatant was aspirated and then washed with wash buffer at room
temperature. The cells were suspended in the wash buffer containing
the anti HIF-1α antibody at a dilution of 1:200 for 30 min on ice
in the dark. The cells were washed and incubated with secondary
donkey anti-mouse immuno-globulin G-FITC (sc-2099; Santa Cruz
Biotechnology, Inc.) antibody at a dilution of 1:200 for 30 min on
ice in the dark, and were then analyzed using a Cell Lab Quanta™ SC
flow cytometer (Beckman Coulter, Inc., Fullerton, CA, USA). The
results were analyzed using FlowJo software, version 7.6.5
(TreeStar, Inc., Ashland, OR, USA).
Detection of active caspase-3
The detection of active caspase-3 was performed
using the FITC-conjugated monoclonal active caspase-3 antibody
apoptosis kit I (BD Biosciences, Franklin Lakes, NJ, USA) according
to the manufacturer's instructions. Briefly, cells were washed with
ice-cold PBS and suspended in Cytofix/Cytoperm™ solution.
Subsequent to incubation on ice for 20 min, the cells were
pelleted, aspirated and then washed with wash buffer at room
temperature. The cells were suspended in the wash buffer containing
5% (v/v) FITC-conjugated anti-active caspase-3 antibody for 40 min
at room temperature in the dark. The cells were then washed with
the wash buffer and analyzed using a Cell Lab Quanta™ SC flow
cytometer.
Annexin V staining
The detection of apoptotic cells was performed using
the TACS Annexin V-FITC Apoptosis Detection kit (Trevigen, Inc.,
Helgerman Ct., MD, USA) according to the manufacturer's
instructions. Briefly, following a 12-h incubation in a 35-mm
culture dish (Iwaki) under hypoxia or normoxia, the cells were
harvested, washed and suspended with 100 μl binding buffer,
and were then stained with Annexin V-FITC for 10 min on ice in the
dark. Following washing with the binding buffer, the cells were
re-suspended in the buffer with propidium iodide (PI). Apoptotic
cells were determined using a Cell Lab Quanta™ SC flow cytometer.
Subsequent to excluding the PI-positive cells from gating, the
fraction of Annexin V-positive cells was evaluated.
Cell cycle analysis
The cell cycle phase distribution was analyzed using
PI staining according to the manufacturer's instructions. Briefly,
following a 12-h incubation in a 35-mm culture dish (Iwaki) under
hypoxia or normoxia, the cells were harvested. Subsequent to
washing with the binding buffer, the cells were re-suspended in the
buffer with PI. Stained cells were analyzed for PI fluorescence
using a Cell Lab Quanta™ SC flow cytometer.
Determination of mitochondrial membrane
potential (MMP)
The MMP was determined by staining with JC-1
(Setareh Biotech, Eugene, OR, USA) according to the manufacturer's
instructions. Briefly, cells were washed in PBS and incubated with
4 μM JC-1 at 37°C for 10 min. The cells were then washed in
PBS and were observed using a BZ-9000 fluorescence microscope
(Keyence, Osaka, Japan). Cells were then harvested and stained with
JC-1 as described above. The cells were then re-suspended in PBS
and analyzed using a Cell Lab Quanta™ SC flow cytometer.
Measurement of mitochondrial
O2•−
The MitoSOX™ RED Mitochondrial
O2•− Indicator (Invitrogen Life Technologies)
was used to detect mitochondrial O2•−.
Briefly, the cells were incubated with 20 μM LW6 under
normoxia or hypoxia, were washed in PBS and were then incubated
with 5 μM MitoSOX™ RED at 37°C for 30 min according to the
manufacturer's instructions. The cells were then washed,
re-suspended in 300 μl PBS and analyzed using a Cell Lab
Quanta™ SC flow cytometer.
Statistical analysis
The significance of the differences was determined
using Student's two-tailed t-test and Welch's t-test depending on
the data distribution. P<0.05 was considered to indicate a
statistically significant difference. Excel 2007 software
(Microsoft Corporation, Redmond, WA, USA) with the add-in software
Statcel 3 was used for statistical analysis.
Results
LW6 inhibits HIF-1α expression induced by
hypoxia
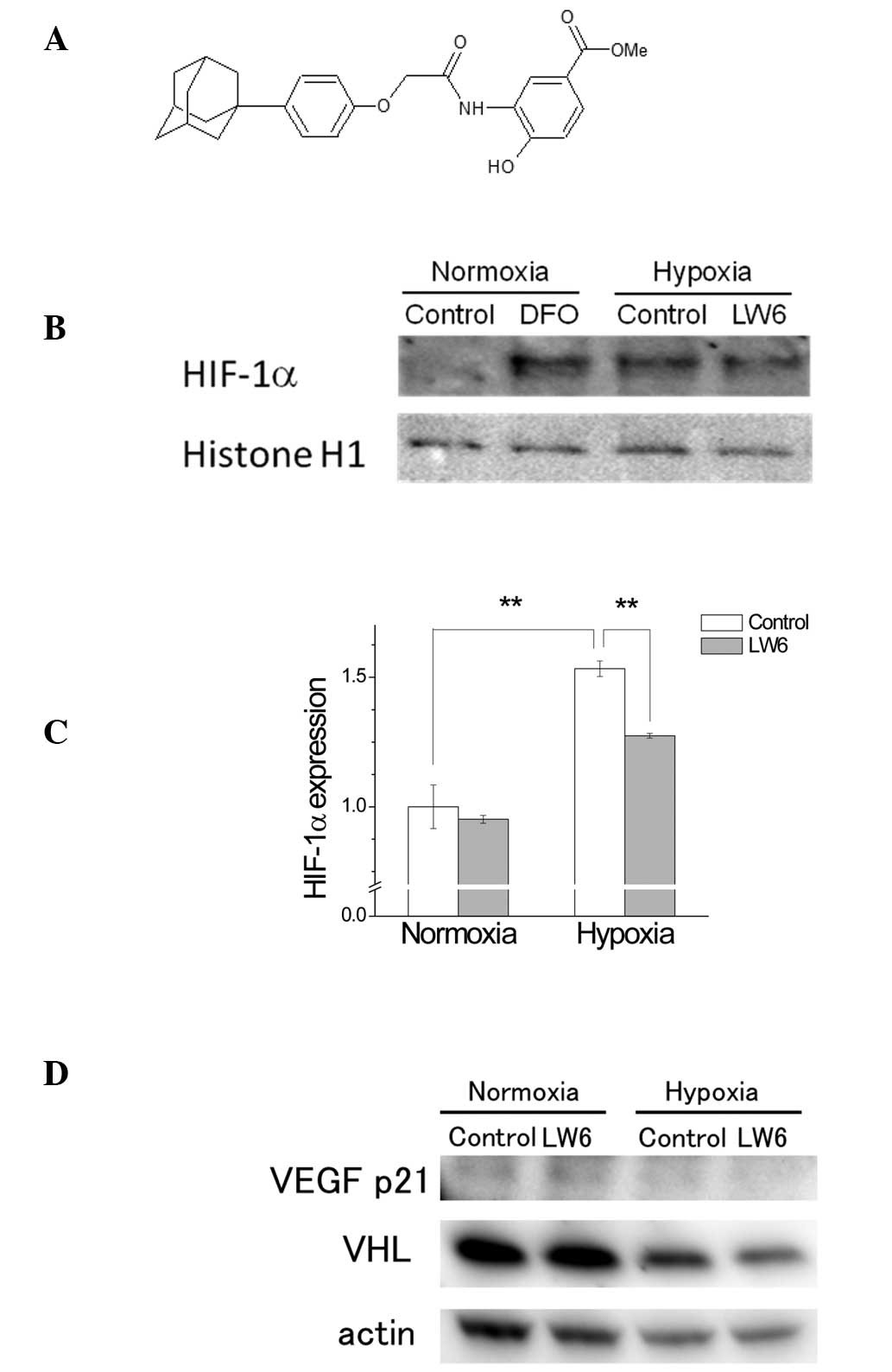
Although LW6 is synthesized as an
(aryloxyacetylamino)benzoic acid derivative (Fig. 1A) and has the potential to inhibit
the expression of HIF-1α in the HCT116 human colon cancer cell line
(17), its mechanism of action has
remained to be fully elucidated. A preliminarily investigation of
the cytotoxicity of different concentrations of LW6 against A549
cells was undertaken. A549 cells were incubated with 5–100
μM LW6 for 24 h and the cytotoxic concentration of LW6 was
determined by MTS assay. It was observed that 100 μM LW6
significantly reduced the cell viability (0.73±0.02; P<0.01). No
cytotoxic effects were observed with concentrations of up to 50 mM
in A549 cells (data not shown).
The increase of HIF-1α expression under hypoxia was
reported to be inhibited by LW6 through the overexpression of VHL,
leading to the inhibition of tumor angiogenesis in the HCT116 human
colon cancer cell line (17). The
potential inhibitory effect of LW6 on the expression of HIF-1α was
investigated in A549 cells. Although the cells incubated under
hypoxia for 36 h exhibited increased expression levels of HIF-1α,
the treatment with LW6 partially reversed this hypoxia-induced
HIF-1α expression (Fig. 1B and C).
LW6 had no effect on the expression of HIF-1α in the normoxic
group. The expression levels of VHL and VEGF were not attenuated by
LW6 treatment in either hypoxia or normoxia (Fig. 1D). These results suggested that LW6
had an inhibitory effect on HIF-1α expression independent of the
upregulation of VHL.
LW6 promotes apoptosis preferentially in
hypoxic cells
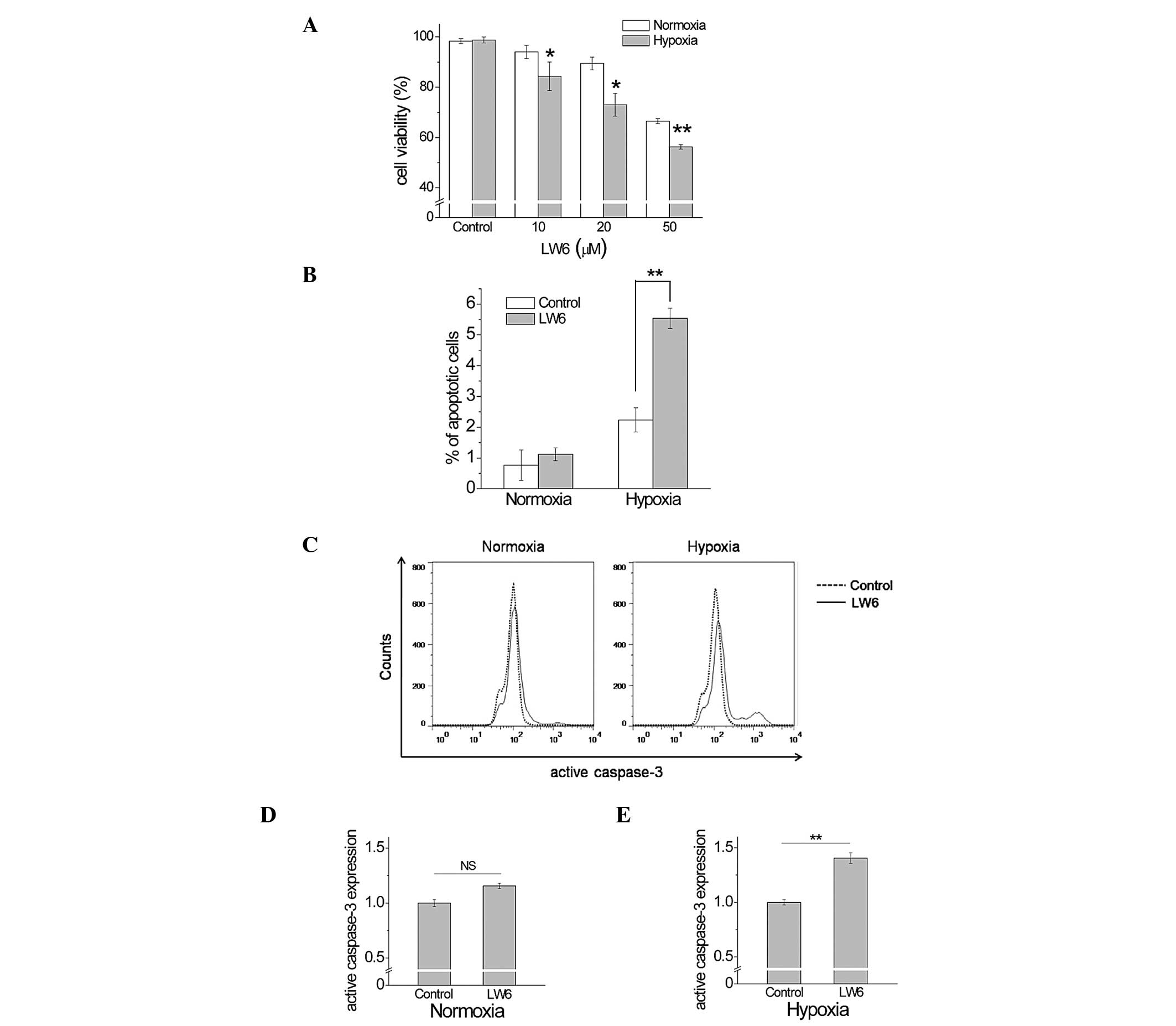
Next, the present study investigated whether the
cell death induced by LW6 is dependent on the oxygen levels that
may be modulated by mitochondrial respiration. Although the
exposure of cells to 20 μM LW6 for 24 h resulted in no
significant toxicity for the metabolic activity of the mitochondria
in the preliminarily MTS assay (data not shown), cell death was
induced by the 48-h exposure to LW6. The cell viability was
significantly reduced in a dose-dependent manner for LW6 under
hypoxia compared with normoxic conditions (Fig. 2A). The cells pre-treated with LW6
under hypoxic conditions exhibited a significant increase in the
percentage of apoptotic cells compared with that of non-treated
cells under hypoxic conditions (5.54±0.32% vs. 2.24±0.39%,
P<0.01, Fig. 2B). No
significant alterations were observed between pre-treated and
non-treated cells under normoxic conditions (1.12±0.20% vs.
0.77±0.49%, P>0.05). The expression levels of active caspase-3
were investigated in order to analyze whether active caspase-3 was
involved in the promotion of apoptosis by LW6. While no significant
difference was observed in active caspase-3 expression between
pre-treated and non-treated cells under normoxic conditions
(1.00±0.031 vs. 1.15±0.024; P>0.05; Fig. 2C and D), cells pre-treated with LW6
under hypoxic conditions displayed a significant increase in active
caspase-3 expression compared with that in non-treated cells
(1.00±0.024 vs. 1.40±0.048; P<0.01; Fig. 2C and E). These results suggested
that active caspase-3 expression participates in the promotion of
apoptosis selectively induced by LW6 in hypoxic A549 cells.
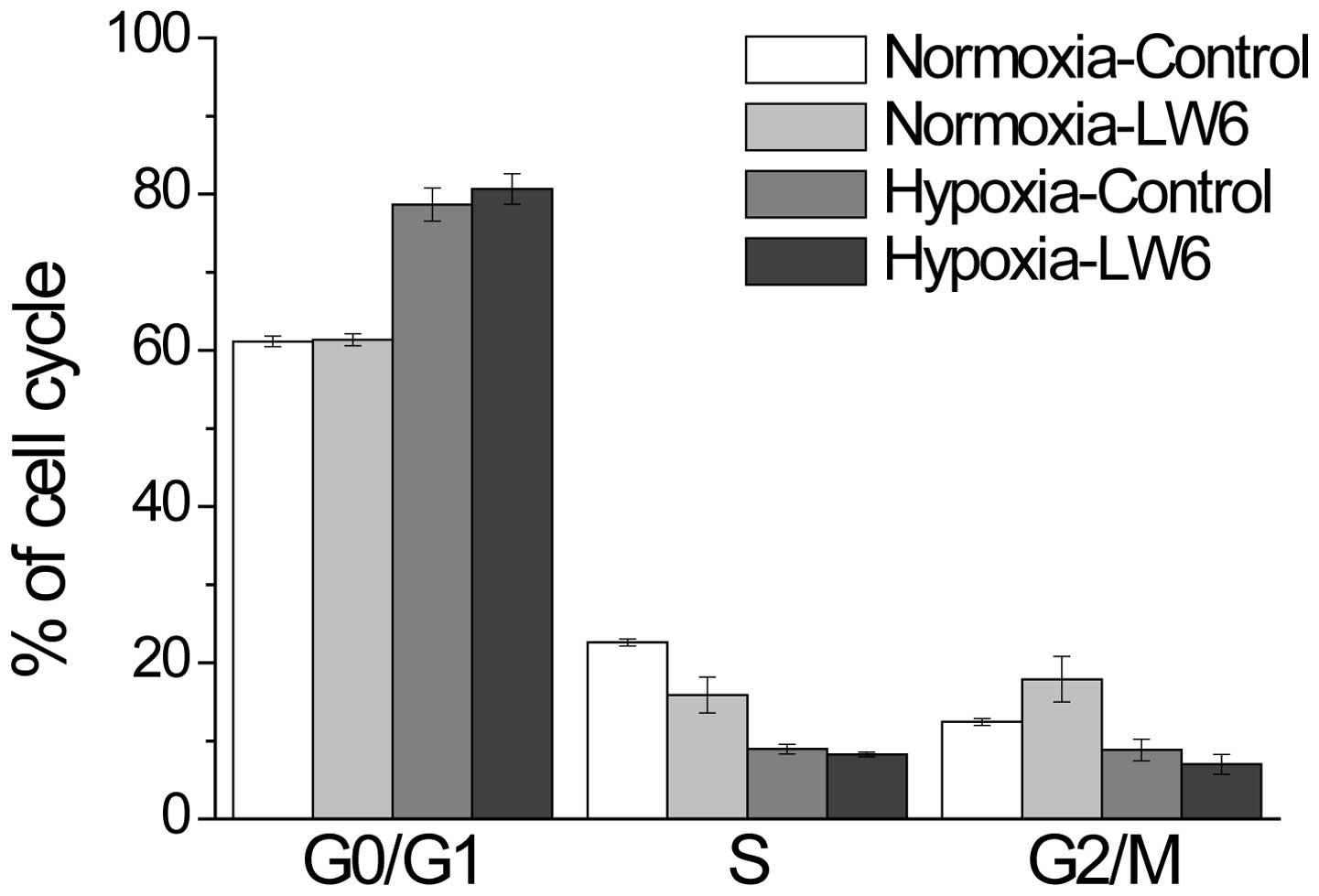
Although the proportion of G1 cells was marginally
increased under hypoxic conditions compared with that under
normoxic conditions, treatment with LW6 presented no additive
effect on cell cycle arrest (Fig.
3).
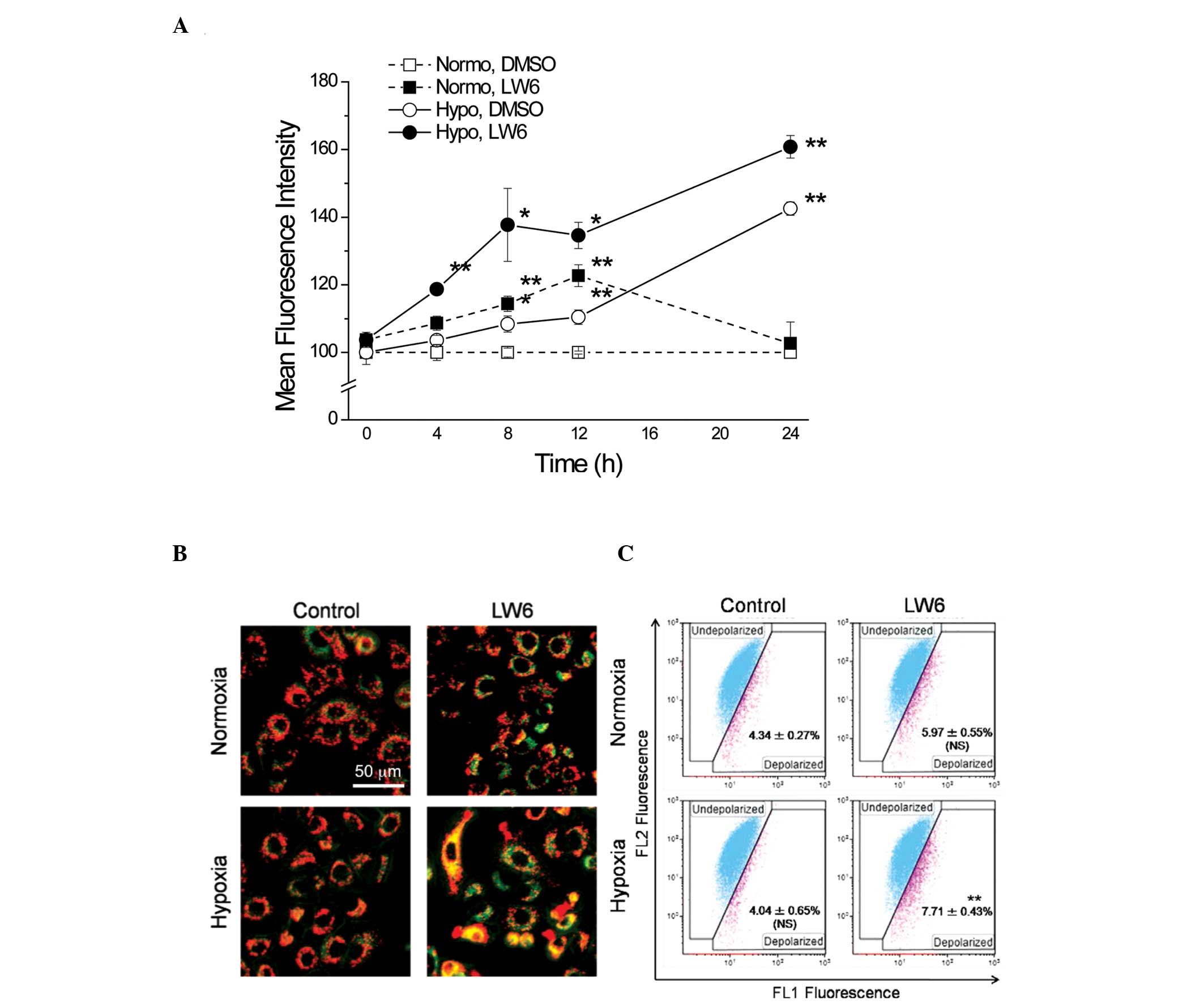
LW6 induces ROS formation through the
depolarization of MMP in hypoxic cells
The activity of the mitochondrial respiratory chain
has been previously reported to be inhibited by HIF-1α (18). Malate dehydrogenase 2 (MDH2), a
critical enzyme involved in the aerobic metabolism of the
mitochondria and the citric acid cycle, has been identified as a
target protein of the HIF-1 inhibitor LW6 (19). The overproduction of intracellular
ROS by mitochondria has been detected in almost all cancer cells,
and the production of ROS is directly associated with the efficacy
of mitochondrial oxygen utilization (20). Therefore, whether LW6 affects the
production of intracellular ROS via the mitochondria was evaluated.
The fluorescence intensity of MitoSOX™ RED was examined using flow
cytometry and subsequently, the time-dependent alterations in the
production of mitochondrial O2•− were assessed. The
exposure to hypoxia significantly increased mitochondrial ROS
production and maintained mitochondrial ROS at high levels.
Although LW6 increased mitochondrial ROS production, the
combination with hypoxia induced a marked increase in ROS
production and this high level was maintained up until 24 h
(Fig. 4A).
Subsequently, the MMP was measured with JC-1, which
also suggested that an alteration in oxygen utilization efficiency
between normoxia and hypoxia results in attenuation of MMP. As
presented in Fig. 4B, treatment 20
μM LW6 combined with hypoxia for 8 h induced a significant
reduction in MMP in A549 cells. Treatment with LW6 under normoxia
was not able to significantly attenuate the MMP, thus suggesting
that the effect of LW6 is dependent on hypoxic conditions (Fig. 4C).
Discussion
In the present study, the potential of LW6 for
inducing apoptosis in normoxic and hypoxic cells was examined. The
results suggest that LW6 preferably induces apoptosis in hypoxic
cells. LW6 was previously reported to inhibit the accumulation of
HIF-1α in hypoxic cells through the upregulation of VHL protein
(17). The VHL protein maintains a
low level of HIF-1α expression in normoxic cells through an
ubiquitin-dependent protein degradation mechanism. HIF-1α
accumulates in cells under hypoxia induced by the rapid growth and
increase in tumor oxygen consumption, enabling the cells to adjust
to the state of hypoxia through gene expression (21). Therefore, necrotic cell death and
apoptosis of cancer cells may be induced by the deletion of HIF-1α.
Supporting this hypothesis, the induction of apoptosis accompanied
by downregulation of HIF-1α was observed in hypoxic cells. In
agreement with the results of the present study, it has been
previously reported that reagents able to inhibit the expression of
HIF-1α in hypoxic cells promote apoptosis (22,23).
Conversely, HIF-1 is involved in hypoxia-induced apoptosis via the
stabilization of p53, through the combination of HIF-1 with p53
ubiquitin ligase mdm2, or by a direct interaction between HIF-1 and
p53 (24,25). An additional mechanism proposed for
HIF-1-mediated apoptosis is the induction of the expression of the
pro-apoptotic protein (B-cell lymphoma 2/adenovirus E1B 19-kDa
interacting protein 3 (BNIP3) gene by HIF-1, through the binding of
the transcription factor to the HRE sequence in the BNIP3 promoter,
thereby resulting in the BNIP3 protein initiating apoptosis and
inducing necrosis (26,27). However, high concentrations of
HIF-1α resulting from activation of the phosphoinositide
3-kinase/Akt pathway have been reported to potentiate resistance to
hypoxia-induced apoptosis in a pancreatic cancer cell line
(28). Previous studies, in
addition to the results of the present study, suggested that HIF-1
enables adaptation to the hypoxic conditions by maintaining the
balance between pro-apoptotic and anti-apoptotic factors.
Hypoxia results in hyperpermeability of the inner
mitochondrial membrane, which leads to the release of cytochrome C
(29). It was suggested that
inhibition of the electron transport chain at the inner
mitochondrial membrane induces apoptosis (29). Lee et al (17) revealed that LW6 is a specific
inhibitor of MDH2 (17). As MDH2
is known to serve a significant role in the citric acid cycle at
the mitochondrial membrane, LW6 indirectly reduces the activity of
the mitochondrial respiratory chain through the inhibition of MDH2.
It was hypothesized that the effect of LW6 on MDH2 activity
indirectly inhibits the electron transport chain, thus leading to
apoptosis. In addition, in the present study the intracellular ROS
levels in the hypoxic A549 cells treated with LW6 were
significantly increased. ROS production resulting from
mitochondrial dysfunction may induce the release of cytochrome C,
which subsequently leads to cell death. In accordance with this, it
was observed in the present study that the loss of MMP is
accompanied by the production of mitochondrial O2•− in
hypoxic cells treated with LW6. Although the influence of LW6 on
ROS production remains to be fully elucidated, the results of the
present study suggested that the hypoxia-selective apoptotic
effects are closely associated with the loss of MMP along with the
dysfunction of mitochondria and increased ROS levels.
In conclusion, LW6 was demonstrated to be able to
inhibit the accumulation of HIF-1α and induce apoptosis through
depolarization of the MMP in hypoxic cells. The present study
suggested that LW6 may be useful in the induction of cell death in
hypoxic cells that have developed resistance to chemotherapy and
radiotherapy. LW6 provides novel insight into cancer therapy
strategy, particularly for the hypoxic cancer cells commonly
observed in tumor tissues.
Acknowledgments
The present study was supported by JSPS KAKENHI,
Grant-in-Aid for Young Scientists (grant B; no. 25861056).
References
|
1
|
The Editorial Board of the Cancer
Statistics in Japan: Cancer Statistics in Japan 2012. Foundation
for Promotion of Cancer Research; Tokyo, Japan: 2012
|
|
2
|
Harris AL: Hypoxia - a key regulatory
factor in tumour growth. Nat Rev Cancer. 2:38–47. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ellis LM and Fidler IJ: Angiogenesis and
metastasis. Eur J Cancer. 32A:2451–2460. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhong H, De Marzo AM, Laughner E, Lim M,
Hilton DA, Zagzag D, Buechler P, Isaacs WB, Semenza GL and Simons
JW: Overexpression of hypoxia-inducible factor 1alpha in common
human cancers and their metastases. Cancer Res. 59:5830–5835.
1999.PubMed/NCBI
|
|
5
|
Brizel DM, Scully SP, Harrelson JM,
Layfield LJ, Bean JM, Prosnitz LR and Dewhirst MW: Tumor
oxygenation predicts for the likelihood of distant metastases in
human soft tissue sarcoma. Cancer Res. 56:941–943. 1996.PubMed/NCBI
|
|
6
|
Theodoropoulos VE, Lazaris AC, Sofras F,
Gerzelis I, Tsoukala V, Ghikonti I, Manikas K and Kastriotis I:
Hypoxia-inducible factor 1 alpha expression correlates with
angiogenesis and unfavorable prognosis in bladder cancer. Eur Urol.
46:200–208. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sundfør K, Lyng H and Rofstad EK: Tumour
hypoxia and vascular density as predictors of metastasis in
squamous cell carcinoma of the uterine cervix. Br J Cancer.
78:822–827. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Walenta S, Wetterling M, Lehrke M,
Schwickert G, Sundfør K, Rofstad EK and Mueller-Klieser W: High
lactate levels predict likelihood of metastases, tumor recurrence
and restricted patient survival in human cervical cancers. Cancer
Res. 60:916–921. 2000.PubMed/NCBI
|
|
9
|
Walenta S, Salameh A, Lyng H, Evensen JF,
Mitze M, Rofstad EK and Mueller-Klieser W: Correlation of high
lactate levels in head and neck tumors with incidence of
metastasis. Am J Pathol. 150:409–415. 1997.PubMed/NCBI
|
|
10
|
Pitson G, Fyles A, Milosevic M, Wylie J,
Pintilie M and Hill R: Tumor size and oxygenation are independent
predictors of nodal diseases in patients with cervix cancer. Int J
Radiat Oncol Biol Phys. 51:699–703. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Papandreou I, Cairns RA, Fontana L, Lim AL
and Denko NC: HIF-1 mediates adaptation to hypoxia by actively
downregulating mitochondrial oxygen consumption. Cell Metab.
3:187–197. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim JW, Tchernyshyov I, Semenza GL and
Dang CV: HIF-1-mediated expression of pyruvate dehydrogenase
kinase: A metabolic switch required for cellular adaptation to
hypoxia. Cell Metab. 3:177–185. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Giaccia A, Siim BG and Johnson RS: HIF-1
as a target for drug development. Nat Rev Drug Discov. 2:803–811.
2003. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Okamoto K, Ito D, Miyazaki K, Watanabe S,
Tohyama O, Yokoi A, Ozawa Y, Asano M, Kawamura T, Yamane Y, et al:
Microregional antitumor activity of a small-molecule
hypoxia-inducible factor 1 inhibitor. Int J Mol Med. 29:541–549.
2012.PubMed/NCBI
|
|
15
|
Welsh S, Williams R, Kirkpatrick L,
Paine-Murrieta G and Powis G: Antitumor activity and
pharmacodynamic properties of PX-478, an inhibitor of
hypoxia-inducible factor-1alpha. Mol Cancer Ther. 3:233–244.
2004.PubMed/NCBI
|
|
16
|
Yeo EJ, Chun YS, Cho YS, Kim J, Lee JC,
Kim MS and Park JW: YC-1: A potential anticancer drug targeting
hypoxia-inducible factor 1. J Natl Cancer Inst. 95:516–525. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee K, Kang JE, Park SK, Jin Y, Chung KS,
Kim HM, Lee K, Kang MR, Lee MK, Song KB, et al: LW6, a novel HIF-1
inhibitor, promotes proteasomal degradation of HIF-1alpha via
upregulation of VHL in a colon cancer cell line. Biochem Pharmacol.
80:982–989. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang H, Gao P, Fukuda R, Kumar G,
Krishnamachary B, Zeller KI, Dang CV and Semenza GL: HIF-1 inhibits
mitochondrial biogenesis and cellular respiration in VHL-deficient
renal cell carcinoma by repression of C-MYC activity. Cancer Cell.
11:407–420. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee K, Ban HS, Naik R, Hong YS, Son S, Kim
BK, Xia Y, Song KB, Lee HS and Won M: Identification of malate
dehydrogenase 2 as a target protein of the HIF-1 inhibitor LW6
using chemical probes. Angew Chem Int Ed Engl. 52:10286–10289.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Y, Fiskum G and Schubert D: Generation
of reactive oxygen species by the mitochondrial electron transport
chain. J Neurochem. 80:780–787. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Carmeliet P, Dor Y, Herbert JM, Fukumura
D, Brusselmans K, Dewerchin M, Neeman M, Bono F, Abramovitch R,
Maxwell P, et al: Role of HIF-1alpha in hypoxia-mediated apoptosis,
cell proliferation and tumour angiogenesis. Nature. 394:485–490.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yeo EJ, Ryu JH, Chun YS, Cho YS, Jang IJ,
Cho H, Kim J, Kim MS and Park JW: YC-1 induces S cell cycle arrest
and apoptosis by activating checkpoint kinases. Cancer Res.
66:6345–6352. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao Q, Du J, Gu H, Teng X, Zhang Q, Qin H
and Liu N: Effects of YC-1 on hypoxia-inducible factor 1-driven
transcription activity, cell proliferative vitality and apoptosis
in hypoxic human pancreatic cancer cells. Pancreas. 34:242–247.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen D, Li M, Luo J and Gu W: Direct
interactions between HIF-1 alpha and Mdm2 modulate p53 function. J
Biol Chem. 278:13595–13598. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hansson LO, Friedler A, Freund S, Rudiger
S and Fersht AR: Two sequence motifs from HIF-1alpha bind to the
DNA-binding site of p53. Proc Natl Acad Sci USA. 99:10305–10309.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kothari S, Cizeau J, McMillan-Ward E,
Israels SJ, Bailes M, Ens K, Kirshenbaum LA and Gibson SB: BNIP3
plays a role in hypoxic cell death in human epithelial cells that
is inhibited by growth factors EGF and IGF. Oncogene. 22:4734–4744.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vande Velde C, Cizeau J, Dubik D, Alimonti
J, Brown T, Israels S, Hakem R and Greenberg AH: BNIP3 and genetic
control of necrosis-like cell death through the mitochondrial
permeability transition pore. Mol Cell Biol. 20:5454–5468. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Akakura N, Kobayashi M, Horiuchi I, Suzuki
A, Wang J, Chen J, Niizeki H, Kawamura Ki, Hosokawa M and Asaka M:
Constitutive expression of hypoxia-inducible factor-1alpha renders
pancreatic cancer cells resistant to apoptosis induced by hypoxia
and nutrient deprivation. Cancer Res. 61:6548–6554. 2001.PubMed/NCBI
|
|
29
|
Tait SW and Green DR: Mitochondria and
cell death: Outer membrane permeabilization and beyond. Nat Rev Mol
Cell Biol. 11:621–632. 2010. View
Article : Google Scholar : PubMed/NCBI
|