Introduction
Diseases of the lumbar intervertebral disc (IVD)
affect the majority of the adult population, causing back pain,
sciatica, disc herniation and spinal stenosis, in addition to other
distressing and disabling spinal symptoms (1–3). The
IVD contains two sections: The annulus fibrosus (AF) and the
nucleus pulposus (NP) (4). The AF
is a ring of flexible fibrocartilage, which surrounds the soft
center consisting of the NP. The IVD acts as a shock buffer when
the spine encounters compression (5). It has been reported that IVD
degeneration initiates in the NP with cellular loss, proteoglycan
content breakdown and decreased water-binding capacity (6).
Current therapies for the treatment of IVD disease
include non-invasive and invasive therapies, which are able to
relieve the symptoms, but not resolve the condition completely
(7,8). Methods for increasing the
proliferative capacity and matrix synthesis of NP cells (NPCs)
include gene therapy and cell therapy (9,10).
Among them, regenerative medicine based on cell therapy is one of
the most promising approaches in the treatment of IVD disease
(11). Gruber et al
(12) attempted to use autologous
disc cell implantation to treat the disc degeneration in a sand rat
model. Okuma et al (13)
reported that the re-insertion of stimulated NPCs retards IVD
degeneration. However, autolo-gous disc cell transplantation is
difficult, as it is, at present, not possible to harvest a high
quantity of autologous disc cells from a single IVD, which would
cause damage to the donor site. Recent advances in cellular and
molecular biology indicated that stem cells exhibit great potential
to be applied in cell therapy (14,15).
Stem cells are able to differentiate into different types of cells,
including osteoblasts, chondrocytes, adipocytes, cardiomyocytes,
hepatocytes and epithelial cells (16). By contrast, stem cells secrete a
large quantity of growth factors, which are vital in cell therapy
(17–19). A number of studies have suggested
that mesenchymal stem cells (MSCs) may be used in the treatment of
IVD diseases (20). Yamamoto et
al (21) observed that bone
marrow-derived stromal cells may improve the viability of NPCs and
direct cell-to-cell contact in a co-culture system between NPCs and
bone marrow MSCs (BMSCs), accomplishing a significant increase in
the viability of NPCs. Wei et al (22) demonstrated that rodent BMSCs were
able to differentiate into IVD-like cells following co-culture with
rat disc tissue. Sun et al (23) noted that ADSCs protect compressive
load-induced NP cell death and degradation by inhibition of
activated caspase-9 and -3 activity in the co-culture system.
Therefore, in the present study, the effect of
adipose-derived stem cells (ADSCs) on degenerated NPCs was
investigated using a co-culture system, and the viability of NPCs
following co-culture with BMSCs was determined, in order to
evaluate the potential application of BMSCs in the treatment of IVD
diseases.
Materials and methods
The present study was approved by the Institutional
Review Board of the General Hospital of Chinese People's Liberation
Army (Beijing, China) and informed consent was obtained from all
patients.
Cell isolation and culture
The human adipose tissue was harvested from patients
who had undergone lipoaspirate procedures. To harvest ADSCs, the
obtained tissue was washed with phosphate-buffered saline (Gibco
Life Technologies, Carlsbad, CA, USA) to remove red blood cells,
cut into small sections and then digested with 0.075% collagenase
(NB4; SERVA Electrophoresis GmbH, Heidelberg, Germany) for 60 min
at 37°C. The digested tissues were filtered using a 200-µm
filter to remove tissue debris and obtain a single-cell suspension.
The cell suspension was centrifuged at 250 × g for 10 min, and the
pellet was then re-suspended in Dulbecco's modified Eagle's medium
containing 10% (v/v) fetal bovine serum (Hyclone, Logan, UT, USA).
The cells were seeded in 100-mm tissue culture dishes at a density
of 2×104 cells/cm2 and cultured at 37°C in a
humidified incubator with 5% CO2. After 7–10 days,
adherent cells were trypsinized (Gibco Life Technologies) and
sub-cultured as described previously (24).
Degenerated NPCs were harvested from NP tissues,
which were obtained from the DDD patients undergoing surgical
procedures as described previously (24). NPCs at passage 2 were used in the
present study.
Groups
A six-well culture plate (Costar®,
Corning, Inc., Corning, NY, USA) and Transwell® inserts
(Corning, Inc.) were used for the co-culture. The Transwell insert
used in the present study consisted of a polyethylene terephthalate
track-etched membrane with 0.4-µm pores at the bottom, which
prevented cell migration. The study groups consisted of the control
group (NPCs seeded onto the Transwell) and the experimental group
(NPCs seeded onto the Transwell and ADSCs seeded onto a six-well
plate).
A total of 1×104 NPCs or ADSCs at passage
2 were seeded in each group. The medium was changed every 2
days.
Cell proliferation
Cell proliferation was determined using the Cell
Counting Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan). A
total of 103 NPCs from the two groups in 100 µl
were seeded into every well of the 96-well plate. After 12 h, 10
µl CCK-8 solution was added into each well. After a further
4 h, the absorbance of the supernatant was measured
spectrophotometrically at 450 nm (Evolution™ 201 UV-visible
Spectrophotometer; Thermo Fisher Scientific, Waltham, MA, USA). The
level of cell proliferation at days 1, 3, 5 and 7 was assessed.
Cell counts were determined using a calibration curve.
DNA and glycosaminoglycan (GAG)
synthesis
The DNA content was quantified using a Qubit dsDNA
HS assay kit (Invitrogen Life Technologies, Carlsbad, CA, USA)
following the manufacturer's instructions. The DNA content was
quantified using the Qubit dsDNA HS assay kit (cat. no. Q32854,
Life Technologies, Grand Island, NY, USA) and the Qubit 2.0
Fluorometer (Life Technologies) according to the manufacturer's
instructions. Briefly, 1 µl DNA sample at 5 ng/µl was
diluted 200-fold in Qubit dsDNA HS buffer in clear plastic Qubit
assay tubes (cat. no. Q32856, Life Technologies) and measured on
the fluorometer. Prior to taking the measurements, a two-point
calibration curve was established using the supplied standards with
the kit, at 0 ng/µl and 10 ng/µl. Samples that fell
below the limit of quantification of 0.5 ng/ml (0.1 ng/µl,
diluted 200-fold) were not reported by Qubit. The contents of GAG
were measured using a Blyscan assay kit (Biocolor, Carrickfergus,
Northern Ireland) as described previously (25).
Telomerase activity
The telomerase activity was determined using a
telomerase polymerase chain reaction (PCR) ELISA kit as described
previously (26). Following
incubation for 7 days, the NPCs from the two groups were
homogenized in a lysis buffer [(0.5% CHAPS, 10 mM Tris-HC1 (pH
7.5), 1 mM MgCl2, 1 mM EGTA, 5 mM β-mercaptoethanol, 0.1
mM AEBSF and 10% glycerol)] and centrifuged at 3,000 × g for 10
min. The extracts were subsequently subjected to a telomeric repeat
amplification protocol. The elongated fragments were then amplified
using PCR. The PCR products were detected and quantified by
performing ELISA according to the manufacturer's instructions and
the results were normalized to those obtained for a standard.
Differentiation characteristics of
NPCs
The relative expression of sex determining region
Y-box 9 (SOX-9), aggrecan and type II collagen was measured in the
two groups at days 7, 14 and 21 to evaluate the differentiation
characteristics of NPCs. RNA was extracted from co-cultured NPCs
and control groups (NPC mono-culture) using TRIzol reagent
(Invitrogen Life Technologies), according to the manufacturer's
instructions, and reverse transcribed (5 µg) into cDNA using
BioScript™ reverse transcriptase (Bioline, Taunton, MA, USA),
according to the manufacturer's instructions. Reverse
transcription-quantitative PCR (RT-qPCR) was used to evaluate the
expression of the cartilage-specific genes SOX-9, aggrecan and type
II collagen. The primer sequences (Beijing Sunbiotech Co., Ltd.,
Beijing, China) are listed in Table
I and the cycling parameters were as follows: Denaturation at
94°C for 2 min; 40 cycles at 94°C for 15 sec and 60°C (or 5°C below
melting temperature) for 1 min. Amplification reactions were
performed in duplicate and the quantity of cDNA in the reactions
was normalized with an internal control, the constitutively
expressed gene GAPDH. The specificity of the amplification of the
expected DNA fragments was confirmed on 2% agarose gel
electrophoresis and by analysis of the melting curves. An
amplification reaction control with no reverse transcriptase
enzyme, termed RT-, was performed in order to assess the
interference of potential genomic DNA in the RNA solution. Relative
gene expression was calculated using the formula: ΔCT =
CTGAPDH - CTtarget.
 | Table IPrimer sequences for reverse
transcription-quantitative polymerase chain reaction. |
Table I
Primer sequences for reverse
transcription-quantitative polymerase chain reaction.
| Gene symbol | Primer | Product (bp) |
|---|
| Type II | F,
5′-CGAAAGGTCAGACGGGTGAA-3′ | 373 |
| Collagen | R,
5′-GGCATTCCCTGAAGACCTGG-3′ | |
| Aggrecan | F,
5′-ACCTCACCATGCCTTCACTG-3′ | 316 |
| R,
5′-GCTCTCACCTTTCACCACGA-3′ | |
| Sox-9 | F,
5′-AGGAGAACCCCAAGATGCAC-3′ | 110 |
| R,
5′-GAGGCGTTTTGCTTCGTCAA-3′ | |
| GAPDH | F,
5′-CCACATCGCTGAGACACCAT-3′ | 353 |
| R,
5′-AAATGAGCCCCAGCCTTCTC-3′ | |
Level of growth factors in
supernatants
ELISA was used to quantitatively determine the
levels of transforming growth factor (TGF)-β1 and insulin-like
growth factor (IGF) in the supernatant in the two groups at
different time-points (27) using
the TGF-β1 human ELISA kit (cat. no. ab100647, Abcam, Cambridge,
UK) and the IGF1 human ELISA kit (cat. no. ab100545, Abcam).
Statistical analysis
One-way analysis of variance was performed using
SPSS 12.0 (SPSS, Inc., Chicago, IL, USA). If the analysis of
variance indicated a significant difference (P<0.05) between the
groups, the difference was evaluated using the least significant
difference test. Values are expressed as the mean ± standard
deviation.
Results
Co-culture with ADSCs enhances
proliferation of NPCs
The number of cells increased gradually in the two
groups with increasing culture time. The cell yields in cultured
NPCs exhibited an increase in the control and experimental groups.
No significant difference was identified between the two groups on
the first day; however, a significant difference appeared as the
time period increased (days 3, 5 and 7; P<0.05) (Fig. 1).
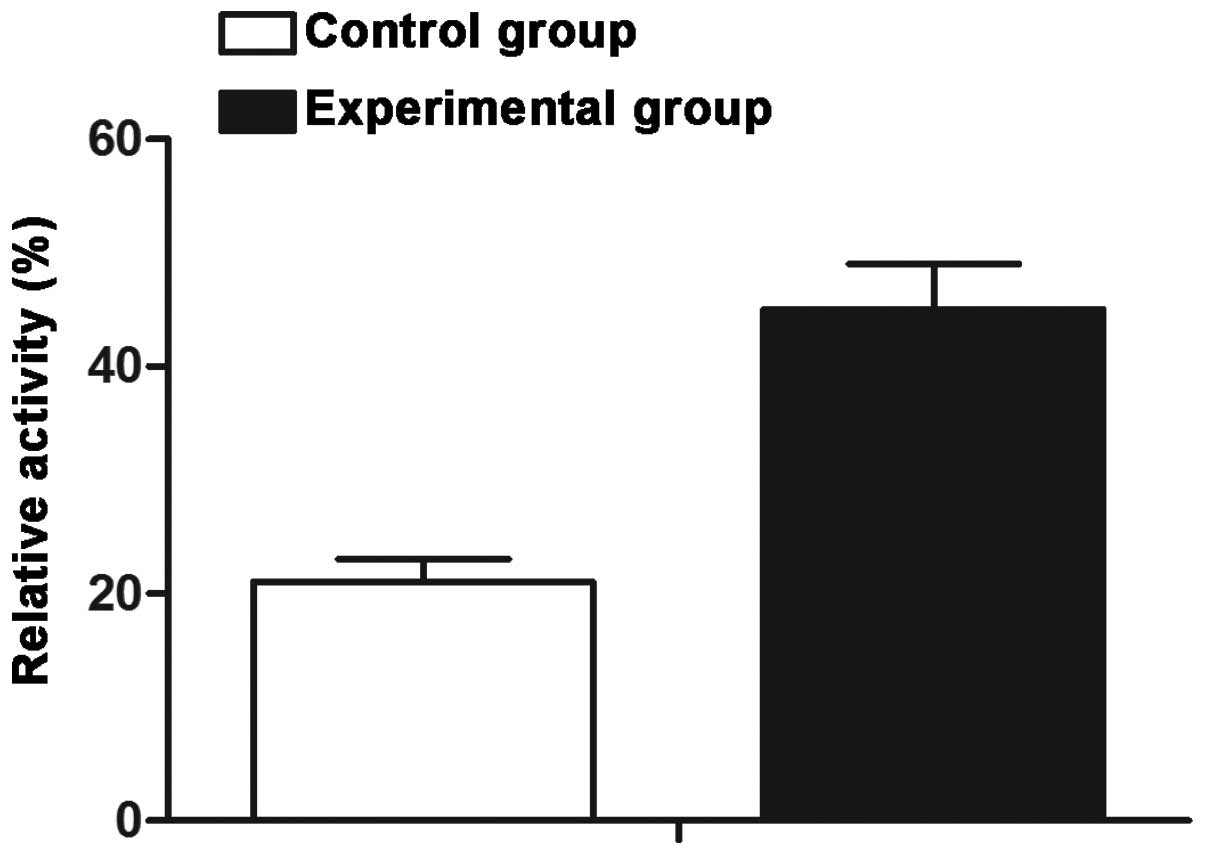
ADSCs enhance telomerase activity in
NPCs
NPCs cultured in the co-culture group exhibited a
higher level of relative telom-erase activity (44.9%) as compared
with that in NPCs in the mono-culture group (21.1%), and a
significant difference was identified between the two groups
(P<0.05) (Fig. 2).
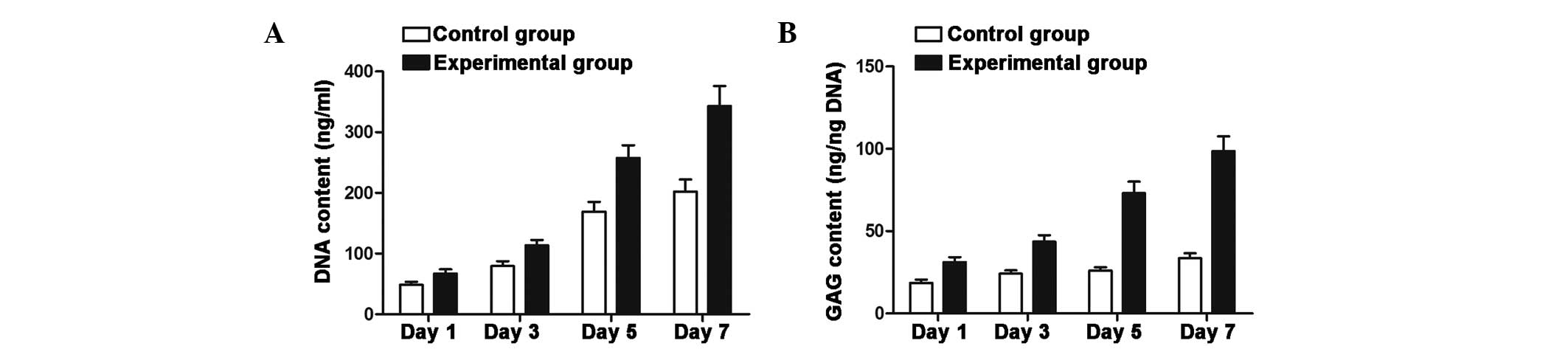
GAG and DNA content of NPCs are increased
in co-culture with ADSCs
The results from the DNA content analysis showed a
marked increase in DNA content in the co-culture group following 5
and 7 days of culture, and were therefore consistent with the
results regarding the level of proliferation (Fig. 3A). Co-culturing of NPCs and ADSCs
resulted in a significant increase in total GAG as compared with
that in the NPC mono-culture (P<0.05) (Fig. 3B).
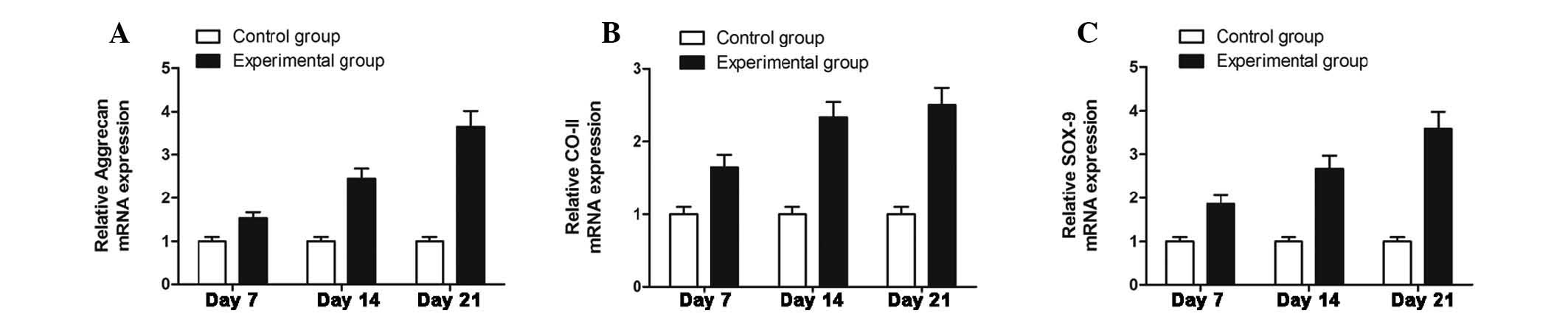
Expression of genes associated with
differentiation is enhanced in NPCs in co-culture with ADSCs
The gene expression of type II collagen, aggrecan
and SOX-9 reflects the chondrogenic differentiation of NPCs.
Fig. 4 demonstrates that the
expression of type II collagen, aggrecan and SOX-9 was upregulated
with increased culture time. Compared with the mRNA levels in NPC
mono-culture, the NPCs in the co-culture groups exhibited a
significantly higher gene expression (P<0.05).
ADSCs secrete growth factors during
co-culture with NPCs
The levels of TGF-β1 and IGF-1 in the supernatant in
the co-culture group was higher than that in the NPC mono-culture
group, which indicated that ADSCs secrete growth factors during the
co-culture process.
Discussion
Stem cell-based cell therapy provides a novel
promising treatment for IVD disorders. In previous years, there
have been an increasing number of studies focusing on the treatment
of IVD disorders, particularly those using BMSCs. BMSCs have been
observed to have the capability of differentiating into
mesen-chymal tissues, including chondrocytes, adipocytes,
osteoblasts, hepatocytes and epithelial cells, among others
(28,29). In addition, BMSCs may secrete
growth factors, which would feed or nurse other cells. Certain
studies have demonstrated that the restoration of BMSCs may improve
IVD degeneration (30). Therefore,
in the present study, the improvement of the biological and
metabolic viability of degenerated NPCs following co-culture with
BMSCs in a co-culture system was evaluated.
The results of the CCK assay indicated that
degenerated NPCs in the co-culture system exhibited a greater level
of proliferation than those in the NPC mono-culture. Certain
studies have revealed that stem cells may promote the proliferation
of other cells (26,31). This may be explained by the finding
that stem cells secrete various growth factors, chemokines and
cytokines, which have paracrine and autocrine activities, and these
secreted bioactive factors suppress the local immune system,
inhibit fibrosis (scar formation) and apoptosis, enhance
angiogenesis, and stimulate mitosis and differentiation of
tissue-intrinsic reparative or stem cells (18). Umeda et al (32) suggested that bone marrow cells are
effective for increasing the proliferative and matrix synthesis
capacity of NPCs. It is known that telomerase is induced in a
primitive subset of progenitor cells and is downregulated upon
further proliferation and differentiation of these cells (33). This provided an explanation as to
why the telomerase activity level of NPCs in the co-culture system
was higher than that in the mono-culture group, as it was
demonstrated that stem cell activity enhanced the viability of
NPCs. Nelson and Chen (34)
proposed the theory that cell-cell signaling by direct contact
increases cell proliferation via phosphoinositide
3-kinase-dependent signaling. The co-culture may promote the
proliferation, viability and phenotypic expression of cells by cell
signaling pathways and the expression of specific adhesion
molecules. This results in cell proliferation and an increase in
cell viability, which was consistent with the results regarding the
GAG and DNA content. The above results indicated that ADSCs may be
ideal cells in the treatment of IVD degeneration, and the
co-culture of NPCs with ADSCs significantly enhanced the biological
activity of NPCs, including the enhancement of cell proliferation,
DNA synthesis and GAG content.
In addition, ADSCs also affected the differentiation
of NPCs. NPCs have a chondrocyte-like appearance and express marked
levels of Sox9, type II collagen mRNA and aggrecan, appearing
rounded and enclosed within a lacuna (35). However, degenerated NPCs lose the
chondrocyte sub-type with the downregulation of
chondrocyte-associated genes. It has been reported that BMSCs are
able to promote the restoration of the lost chondrocyte sub-type in
de-differentiated chondrocytes. Therefore, the chondrogenic
differentiation of degenerated NPCs was evaluated in the co-culture
system, and the results were consistent with those of previous
studies (36). The expression of
SOX-9, collagen type II and aggrecan in the co-culture group was
higher than that in the control group at different time-points. The
differentiation of NPCs was determined by ADSCs co-culture system,
and TGF-β1 and IGF-1, which are considered major regulatory
cytokines, secreted by ADSCs, are able to promote the chondrogenic
differentiation of degenerated NPCs. Therefore, the levels of
growth factors in the supernatant were assessed in the two groups,
and the results showed that the growth factors secreted by ADSCs
were present in the co-culture medium and therefore had an
important role in the chondrogenic differentiation of NPCs. Sun
et al (37) demonstrated
that direct co-cultures of ADSCs and NP cells stimulated ADSCs
differentiation into the NP cell phenotype.
In conclusion, the present study confirmed that
co-culture of ADSCs and NPCs not only resulted in the increased
proliferation and viability of NPCs, but also promoted the
expression of type II collagen, SOX-9 and aggrecan genes,
indicating that the injection of ADSCs into degenerative IVD may be
a feasible and promising therapy in the treatment of IVD
disorders.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant no. 30872607).
References
|
1
|
Adams MA, Lama P, Zehra U and Dolan P: Why
do some intervertebral discs degenerate, when others (in the same
spine) do not? Clin Anat. 28:195–204. 2015. View Article : Google Scholar
|
|
2
|
Ohtori S, Inoue G, Miyagi M and Takahashi
K: Pathomechanisms of discogenic low back pain in humans and animal
models. Spine J. pii(Suppl): S1529–S9430. 2014.
|
|
3
|
Parker SL, Godil SS, Mendenhall SK,
Zuckerman SL, Shau DN and McGirt MJ: Two-year comprehensive medical
management of degenerative lumbar spine disease (lumbar
spondylolisthesis, stenosis, or disc herniation): a value analysis
of cost, pain, disability and quality of life: clinical article. J
Neurosurg Spine. 21:143–149. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim SH, Song JE, Lee D and Khang G:
Demineralized bone particle impregnated poly
(l-Lactide-co-Glycolide) scaffold for application in
tissue-engineered intervertebral discs. J Biomater Sci Polym Ed.
Nov 29–2011.Epub ahead of print.
|
|
5
|
Bron JL, Helder MN, Meisel HJ, Van Royen
BJ and Smit TH: Repair, regenerative and supportive therapies of
the annulus fibrosus: achievements and challenges. Eur Spine J.
18:301–313. 2009. View Article : Google Scholar
|
|
6
|
Tow BP, Hsu WK and Wang JC: Disc
regeneration: a glimpse of the future. Clin Neurosurg. 54:122–128.
2007.
|
|
7
|
Thacher C: Neuroanatomic and
pathophysiologic aspects of intervertebral disc disease in the dog.
Probl Vet Med. 1:337–357. 1989.PubMed/NCBI
|
|
8
|
Jezussek D, Schuh A, Hönle W and Janka M:
Conservative therapeutic options in intervertebral disc disease.
MMW Fortschr Med. 152:36–39. 2010.PubMed/NCBI
|
|
9
|
Sakai D: Future perspectives of cell-based
therapy for intervertebral disc disease. Eur Spine J. 17(Suppl 4):
452–458. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Johnson WE and Roberts S: 'Rumours of my
death may have been greatly exaggerated': a brief review of cell
death in human intervertebral disc disease and implications for
cell transplantation therapy. Biochem Soc Trans. 35:680–682. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sakai D, Nakamura Y, Nakai T, et al:
Exhaustion of nucleus pulposus progenitor cells with ageing and
degeneration of the intervertebral disc. Nat Commun. 3:12642012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gruber HE, Johnson TL, Leslie K, et al:
Autologous intervertebral disc cell implantation: a model using
Psammomys obesus, the sand rat. Spine (Phila Pa 1976).
27:1626–1633. 2002. View Article : Google Scholar
|
|
13
|
Okuma M, Mochida J, Nishimura K, Sakabe K
and Seiki K: Reinsertion of stimulated nucleus pulposus cells
retards intervertebral disc degeneration: an in vitro and in vivo
experimental study. J Orthop Res. 18:988–997. 2000. View Article : Google Scholar
|
|
14
|
Martinez-Morales PL, Revilla A, Ocaña I,
et al: Progress in stem cell therapy for major human neurological
disorders. Stem Cell Rev. 9:685–699. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang J, Liao L, Wang S and Tan J: Cell
therapy with autologous mesenchymal stem cells-how the disease
process impacts clinical considerations. Cytotherapy. 15:893–904.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wagner W, Wein F, Seckinger A, et al:
Comparative characteristics of mesenchymal stem cells from human
bone marrow, adipose tissue and umbilical cord blood. Exp Hematol.
33:1402–1416. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kilroy GE, Foster SJ, Wu X, et al:
Cytokine profile of human adipose-derived stem cells: expression of
angiogenic, hematopoietic and pro-inflammatory factors. J Cell
Physiol. 212:702–709. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Caplan AI and Dennis JE: Mesenchymal stem
cells as trophic mediators. J Cell Biochem. 98:1076–1084. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Boomsma RA and Geenen DL: Mesenchymal stem
cells secrete multiple cytokines that promote angiogenesis and have
contrasting effects on chemotaxis and apoptosis. PLoS One.
7:e356852012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mwale F, Wang HT, Roughly P, Antoniou J
and Haglund L: Link N and mesenchymal stem cells can induce
regeneration of the early degenerate intervertebral disc. Tissue
Eng Part A. 20:2942–2949. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yamamoto Y, Mochida J, Sakai D, et al:
Upregulation of the viability of nucleus pulposus cells by bone
marrow-derived stromal cells: significance of direct cell-to-cell
contact in coculture system. Spine (phila pa 1976). 29:1508–1514.
2004. View Article : Google Scholar
|
|
22
|
Wei A, Chung SA, Tao H, et al:
Differentiation of rodent bone marrow mesenchymal stem cells into
intervertebral disc-like cells following coculture with rat disc
tissue. Tissue Eng Part A. 15:2581–2595. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sun Z, Luo B, Liu ZH, Samartzis D, Liu Z,
Gao B, Huang L and Luo ZJ: Adipose-derived stromal cells protect
intervertebral disc cells in compression: Implications for stem
cell regenerative disc therapy. Int J Biol Sci. 11:133–143. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lu ZF, Zandieh Doulabi B, Wuisman PI, Bank
RA and Helder MN: Differentiation of adipose stem cells by nucleus
pulposus cells: configuration effect. Biochem Biophys Res Commun.
359:991–996. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim JY, Choeng HC, Ahn C and Cho SH: Early
and late changes of MMP-2 and MMP-9 in bleomycin-induced pulmonary
fibrosis. Yonsei Med J. 50:68–77. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Niu CC, Yuan LJ, Lin SS, Chen LH and Chen
WJ: Mesenchymal stem cell and nucleus pulposus cell coculture
modulates cell profile. Clin Orthop Relat Res. 467:3263–3272. 2009.
View Article : Google Scholar :
|
|
27
|
Schmidmaier G, Herrmann S, Green J, et al:
Quantitative assessment of growth factors in reaming aspirate,
iliac crest and platelet preparation. Bone. 39:1156–1163. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hass R, Kasper C, Böhm S and Jacobs R:
Different populations and sources of human mesenchymal stem cells
(MSC): A comparison of adult and neonatal tissue-derived MSC. Cell
Commun Signal. 9:122011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pittenger MF, Mackay AM, Beck SC, et al:
Multilineage potential of adult human mesenchymal stem cells.
Science. 284:143–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sobajima S, Vadala G, Shimer A, Kim JS,
Gilbertson LG and Kang JD: Feasibility of a stem cell therapy for
intervertebral disc degeneration. Spine J. 8:888–896. 2008.
View Article : Google Scholar
|
|
31
|
Watanabe T, Sakai D, Yamamoto Y, et al:
Human nucleus pulposus cells significantly enhanced biological
properties in a coculture system with direct cell-to-cell contact
with autologous mesenchymal stem cells. J Orthop Res. 28:623–630.
2010.
|
|
32
|
Umeda M, Kushida T, Sasai K, et al:
Activation of rat nucleus pulposus cells by coculture with whole
bone marrow cells collected by the perfusion method. J Orthop Res.
27:222–228. 2009. View Article : Google Scholar
|
|
33
|
Chiu CP, Dragowska W, Kim NW, et al:
Differential expression of telomerase activity in hematopoietic
progenitors from adult human bone marrow. Stem Cells. 14:239–248.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nelson CM and Chen CS: Cell-cell signaling
by direct contact increases cell proliferation via a PI3K-dependent
signal. FEBS Lett. 514:238–242. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sive JI, Baird P, Jeziorsk M, Watkins A,
Hoyland JA and Freemont AJ: Expression of chondrocyte markers by
cells of normal and degenerate intervertebral discs. Mol Pathol.
55:91–97. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Maidhof R, Alipui DO, Rafiuddin A, Levine
M, Grande DA and Chahine NO: Emerging trends in biological therapy
for intervertebral disc degeneration. Discov Med. 14:401–411.
2012.
|
|
37
|
Sun Z, Liu ZH, Zhao XH, Sun L, Chen YF,
Zhang WL, Gao Y, Zhang YZ, Wan ZY, Samartzis D, Wang HQ and Luo ZJ:
Impact of direct cell co-cultures on human adipose-derived stromal
cells and nucleus pulposuscells. J Orthop Res. 31:1804–1813.
2013.PubMed/NCBI
|