Introduction
Rheumatoid arthritis (RA) is a common inflammatory
autoimmune disease and the associated inflammation has been
extensively studied. RA patients suffer from pain (1–3),
which affects their daily lives (4). RA pain is frequently characterized by
neuropathic pain. The dorsal root ganglion (DRG) is a place where
primary sensory neurons accumulate in the pain conduction pathway
(5), which may selectively
perceive noxious stimulation and potential tissue damage. The
sensory neuron sensitization of the DRG is one of the principal
mechanisms underlying pain responses that are affected by the
interaction of inflammatory cells with neurons in the DRG and the
central nervous system. In addition, the release of inflammatory
mediators (e.g., various prostaglandins, leukotrienes and
interleukins) serves a crucial role in mediating the chronic pain
of RA. The inflammatory mediators lead to peripheral sensitization
of sensory nerves, and alterations in nerve growth and gene
expression. Therefore, RA pain originates from joint inflammation
and pain signal transduction processes, with complex regulatory
mechanisms. It is, therefore, of great interest to identify whether
there is a modulator that simultaneously influences the release of
inflammatory pain mediators and pain conduction.
Previously, microRNAs (miRs) have been associated
with certain pathological mechanisms for chronic inflammatory pain
and neuropathic pain (6,7). The expression of miRs is
significantly altered in different nerve tissues of previous pain
models. In addition, selective accumulation or deletion of miRs in
local synapses can regulate gene expression, protein synthesis and
inflammatory pain-associated transcription factors (8), being closely associated with the
occurrence and maintenance of pain. The authors' previous studies
have also confirmed abnormalities in miR expression and alterations
in miR-143 expression altered in RA patients and animal models
(9,10). Certain reports also demonstrated
that miR-143 had an association with inflammatory pain responses.
For example, the expression of hsa-miR-143-3p is reduced in
patients with fibromyalgia (11);
miR-143 degrades cyclooxygenase (COX)-2 mRNA and regulates pain
mediator production in pancreatic cancer (12). Tam et al (13) demonstrated that the expression
levels of miR-143 were significantly lower in DRGs ipsilateral to
complete Freund's adjuvant (CFA) injection. Therefore, the present
study aimed to investigate the role of miR-143 in the complex
mechanism of RA pain and to identify key pain targets for
regulating pain responses.
In the present study, collagen-induced arthritis
(CIA) was first compared by observing pain responses and detecting
the levels of inflammatory pain mediators in the DBA/1 and C57BL/6
mice. miR-143 expression in the peripheral blood and DRG of CIA
mice was further investigated, and the target genes that regulated
RA pain responses were clarified. The results may provide novel
insights into the development of strategies to prevent or alleviate
RA pain.
Materials and methods
Animals
A total of 18 DBA/1 and 12 C57BL/6 male mice aged
6–8 weeks (16–20 g) were provided by the Laboratory Animal Center
of the Academy of Military Medical Sciences (Beijing, China). The
experiment was performed in the Experimental Animal Center of
Nanjing University of Chinese Medicine (Jiangsu, China). Mice were
housed in a temperature- (22±3°C) and humidity-controlled (40–70%)
animal room under a 12-h light/dark cycle, with free access to food
and water. All experimental protocols performed in this study were
approved by the Ethics Review Committee for Animal Experimentation
of Nanjing University of Chinese Medicine and were in accordance
with the Declaration of the National Institutes of Health Guide for
Care and Use of Laboratory Animals (14).
Establishment of the DBA/1 or C57BL/6
mouse model of CIA (15)
Bovine type II collagen (CII; Chondrex, Inc.,
Redmond, WA, USA) was dissolved in 0.01 mol/l acetic acid into a 2
g/l solution and shaken overnight at 4°C. The solution was mixed
with CFA (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at a ratio
of 1:1 and emulsified. A total of 12 DBA/1 or C57BL/6 mice were
separately divided into two groups (n=6). In the model group, a
mixed emulsion (100 µl) of CII and CFA was intradermally injected
at the tail root of mice for primary immunization, and this day was
recorded as d1. On d21, CII was mixed evenly with incomplete
Freund's adjuvant (Sigma-Aldrich; Merck KGaA) at a ratio of 1:1,
100 µl of which was injected at the tail root to boost
immunization. The mice in the control group were intradermally
injected with normal saline at the tail root on d1 and d21,
respectively.
Detection of inflammatory responses
and pain mediator release in DBA/1 or C57BL/6 mice with CIA
The paw thickness of DBA/1 mice was measured on d21,
d28, d35, d41 and d49, and the mice were sacrificed by anesthesia
on d49. The paw thickness of C57BL/6 mice was measured on d5, d12,
d19, d26, d33, d40, d47 and d54, and the mice were sacrificed by
anesthesia on d54. The sera of all mice were collected and the
levels of tumor necrosis factor (TNF)-α and prostaglandin
(PG)E2 were measured using mouse TNF-α (cat. no.
YY02868B) and PGE2 ELISA kits (cat. no. YY02798B;
Shanghai Yuanye Biotechnology Co., Ltd., Shanghai, China).
Detection of pain responses in DBA/1
or C57BL/6 mice with CIA
Pain responses, including the mechanical withdrawal
threshold (MWT) and thermal withdrawal latency (TWL) were measured
on d20, d24, d30, d36, d42 and d48 in DBA/1 mice, and on d5, d12,
d19, d26, d33, d40, d47 and d54 in C57BL/6 mice. The tests were
performed as follows. The experimental environment was kept quiet.
The mice were placed in a plexiglass box with metal mesh bottom.
Following >30 min of adaptation, the experiment was begun when
the exploration activities had stopped. Von Frey filaments
(0.008–4.0 g) were used to stimulate the mouse paw vertically into
a slight S shape. Obvious paw withdrawal, lifting or licking was
regarded as a positive response, and no response was negative. MWT
was measured and calculated using the ‘up and down’ method
described by Chaplan et al (16).
Under the same conditions described above, the mouse
paw was irradiated with a radiation source using a 37370 thermal
plantar analgesia instrument and the avoidance time, from the
initiation of irradiation to leg lifting and paw withdrawal, was
recorded. Each mouse was tested three times with a 5-min interval,
and the average was recorded as TWL.
Detection of miR-143-3p expression in
the peripheral blood and DRG using reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
A total of 6 DBA/1 mice were equally divided into a
control group and a CIA group, and the CIA model was established
with the method described above. Peripheral blood and DRG tissues
were obtained from the control and CIA mice on d49, from which the
RNA was extracted with ice-cold TRIzol® (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The RNA concentration and
purity were spectrophotometrically determined at 260 and 280 nm.
cDNA was synthesized using a Transcriptor First-Strand cDNA
Synthesis kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China).
The reverse transcription reaction was performed under the
following conditions: 16°C for 30 min, 42°C for 40 min and 85°C for
5 min. The target miR of each sample and the internal reference
(mmu-miR-425-5p) were detected using an Applied Biosystems
detection instrument (Thermo Fisher Scientific, Inc.).
mmu-miR-425-5p was used for normalization. The SYBR®
Green qPCR Master Mix (Arraystar Inc, Rockville, MD, USA) was used
and the PCR was performed using the following cycling conditions:
95°C for 10 min, followed by 40 cycles of 95°C for 10 sec and 60°C
for 60 sec. The data were analyzed using the 2−ΔΔCq
method (17). A total of three
replicate wells were set up and the results were averaged. The
primers were as follows: mmu-miR-143-3p gene specific primer (GSP),
5′-GGGATGAGATGAAGCACT-3′ and reverse (R), 5′-TGCGTGTCGTGGAGTC-3′;
mmu-miR-425-5p GSP, 5′-GGGGAATGACACGATCACTC-3′ and R,
5′-GTGCGTGTCGTGGAGTCG-3′.
Bioinformatics analysis of
pain-associated target genes of miR-143
The target genes of miR-143 were predicted using
bioinformatics databases, including miRanda (http://www.mirbase.org), miRDB v. 4.0 (http://mirdb.org/miRDB), TargetScan v. 6.2 (http://www.targetscan.org), miRWalk v. 2.0 (http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2)
and RNA22 v. 2 (https://cm.jefferson.edu/rna22/Interactive). The
target genes of the three intersects were selected to minimize the
false positive rate. All the target genes were analyzed by
Cytoscape software v. 3.5.0 (http://www.cytoscape.org/) to obtain the microRNA-gene
regulatory network. Using the gene database from the Pain Genetics
Lab (http://www.psych.mcgill.ca/labs/mogillab/paingenetics/lab/)
of McGill University (Montreal, Canada), the predicted target genes
of miR-143 were analyzed comprehensively to determine potential
pain-associated genes.
Gene ontology (GO) analysis and
pathway analysis of miR-143 target genes
GO functional annotation enrichment analysis of the
predicted miR-143 target genes was conducted using the GO database
(http://www.geneontology.org), including
cell components and molecular function. Using the Kyoto
Encyclopedia of Gene and Genomes (KEGG) database (http://www.genome.jp/kegg) in the DAVID database,
biological pathway enrichment analysis was performed on the
predicted target genes.
Isolation, culture and transfection of
DRG cells
In an ice bath, the spinal canal of a normal mouse
was cut open under a dissecting microscope to remove the spinal
cord and other nerve fibers to isolate the DRG. Subsequently, 1 ml
mixed digestive enzyme was added, digestion was performed in a 37°C
water bath for 25–30 min and the resulting tissue was pipetted
repeatedly into a single cell suspension. The supernatant was
discarded following centrifugation (4°C, 12,000 × g, 15 min) and 1
ml DH10 medium (WISENT Inc., Quebec, QC, Canada) and 1% nerve
growth factor (NGF; R&D Systems, Inc., Minneapolis, MN, USA)
were added to the precipitate. The cell suspension was pipetted
into a single cell suspension and grown in a pre-coated petri dish.
When the density of DRG cells reached 50–60% confluence, the cell
culture medium was replaced prior to transfection.
The mmu-miR-143-3p sequence (Shanghai GenePharma
Co., Ltd., Shanghai, China) was dissolved into a 10 µM stock
solution, aliquoted and stored at-20°C. When the density of DRG
cells reached 50–60%, the culture medium was refreshed prior to
transfection and 2 ml antibiotic-free DH10 medium was added into
each dish. A total of 3 µl miR mimic, inhibitor or miR negative
control (NC) (Shanghai GenePharma Co., Ltd.) was mixed gently with
150 µl Opti-MEM reduced serum medium (Gibco; Thermo Fisher
Scientific, Inc.). A total of 9 µl Lipofectamine®
RNAiMAX (Invitrogen; Thermo Fisher Scientific, Inc.) was diluted
with 150 µl Opti-MEM reduced serum medium. Diluted
Lipofectamine® RNAiMAX was mixed gently with diluted
miR. miR-143 mimic NC-FAM (carboxyfluorescein): Sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′. miR-143 mimic: Sense,
5′-UGAGAUGAAGCACUGUAGCUC-3′ and antisense,
5′-GCUACAGUGCUUCAUCUCAUU-3′. miR-143 inhibitor: Sense,
5′-GAGCUACAGUGCUUCAUCUCA-3′. miR-143 inhibitor NC: Sense,
5′-CAGUACUUUUGUGUAGUACAA-3′. Following incubation at room
temperature for 5 min, the mixture was added into the dish
containing the DRG cells and culture medium, mixed by gentle
agitation and cultured in a CO2 incubator at 37°C. The
cells were observed using fluorescence microscopy at 6, 12, 24, 48
and 72 h following transfection to determine the suitable
transfection conditions.
Detection of the expression of
miR-143-3p target genes in transfected DRG cells and DRG tissues of
CIA mice
DRG cells were cultured and transfected with the
method described above. miR-143-3p mimic, inhibitor and miR NCs
were transfected into DRG cells for 48 h. RNA was extracted from
DRG cells and the expression of mmu-miR-143, MAS related GPR family
member E (Mrgpre), prostaglandin endoperoxide synthase 2 (Ptgs2,
also known as COX-2), prostaglandin D2 receptor (Ptgdr) and Tnf
were detected using qPCR.
A total of 6 DBA/1 mice were divided into a control
group and a CIA group. The CIA model was established by the method
described above. DRG tissues were obtained from the control and CIA
mice on d49, from which RNA was extracted. The expression of
Mrgpre, Ptgs2 and Tnf were detected using RT-qPCR, as described
above.
The primers were as follows: Mrgpre forward (F),
5′-CAGAACCACCCGTAGGCAAAT-3′ and R, 5′-TTCCAGGTCACCGTCCATCAC-3′;
Ptgs2 F, 5′-CAGATGACTGCCCAACTCCCA-3′ and R,
5′-GTGAACCCAGGTCCTCGCTTA-3′; Ptgdr F, 5′-AACACCGTCTCACTGTAGGCTT-3′
and R, 5′-CTGGTTTCCCAACTCATTTCTC-3′; Tnf F,
5′-GAGTCCGGGCAGGTCTACTTT-3′ and R, 5′-CAGGTCACTGTCCCAGCATCT-3′; U6
F, 5′-GCTTCGGCAGCACATATACTAAAAT-3′ and R,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; and GAPDH F,
5′-CACTGAGCAAGAGAGGCCCTAT-3′ and R,
5′-GCAGCGAACTTTATTGATGGTATT-3′.
Data processing and statistics
The experiments were repeated three times.
Experimental data are expressed as the means ± standard deviation.
T-test was used for the comparison of two groups and analysis of
variance followed by Tukey's test was used for the comparison of
multiple groups. Data processing was performed using the SPSS v.
13.0 statistical software (SPSS, Inc., Chicago, IL, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
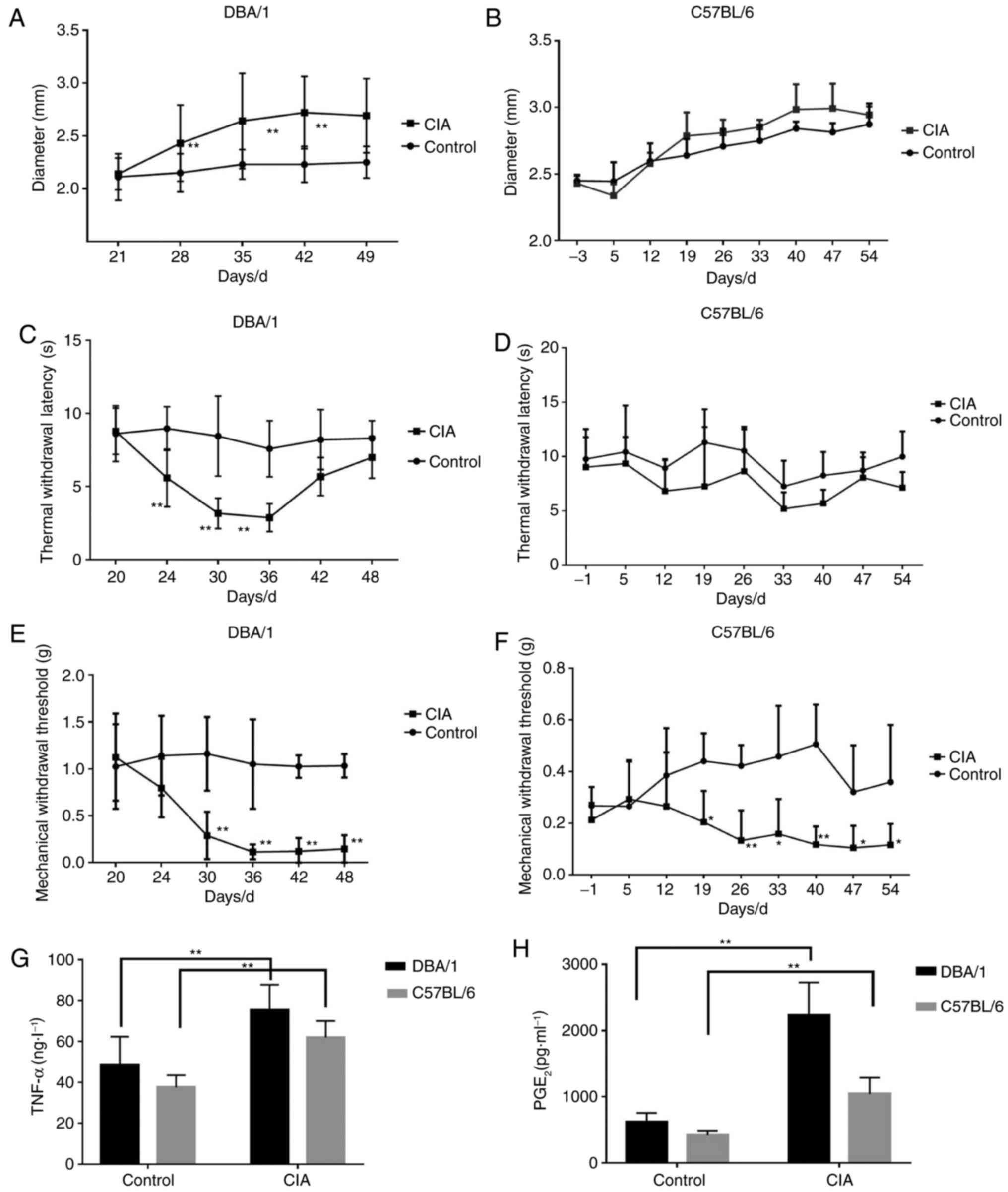
Comparison of inflammatory pain
responses and pain mediator release in DBA/1 or C57BL/6 mice with
CIA
DBA/1 and C57BL/6 mouse models of CIA were
constructed, and the alterations of CIA-associated indices were
investigated. The forelimb and hindlimb became red and swollen from
d24 (DBA/1 mice) and d33 (C57BL/6 mice), and their paws and ankle
joints were also notably affected on d28 (DBA/1 mice) and d40
(C57BL/6 mice). The paw thickness markedly increased with the
progression of arthritis in the CIA group compared with the control
group (P<0.01; Fig. 1A and
B).
MWT and TWL, which indicate pain responses, were
also detected. For DBA/1 mice, the TWL of the CIA model group
decreased significantly from d24 (P<0.01; Fig. 1C) and gradually recovered from d42,
and MWT dropped significantly from d30 (P<0.01; Fig. 1E). For C57BL/6 mice, TWL was
slightly lower compared with the control group (P>0.05; Fig. 1D) and the MWT of the CIA model
group significantly decreased (P<0.05; Fig. 1F).
Additionally, the levels of inflammatory pain
mediators, including TNF-α and PGE2, significantly
increased in the CIA model group compared with the control group
for the two strains of mice (P<0.01; Fig. 1G and H).
The results indicated that DBA/1 mice underwent more
marked hyperalgesia and had increased levels of inflammatory
mediators compared with the C57BL/6 mice, thus they were selected
for the following experiments.
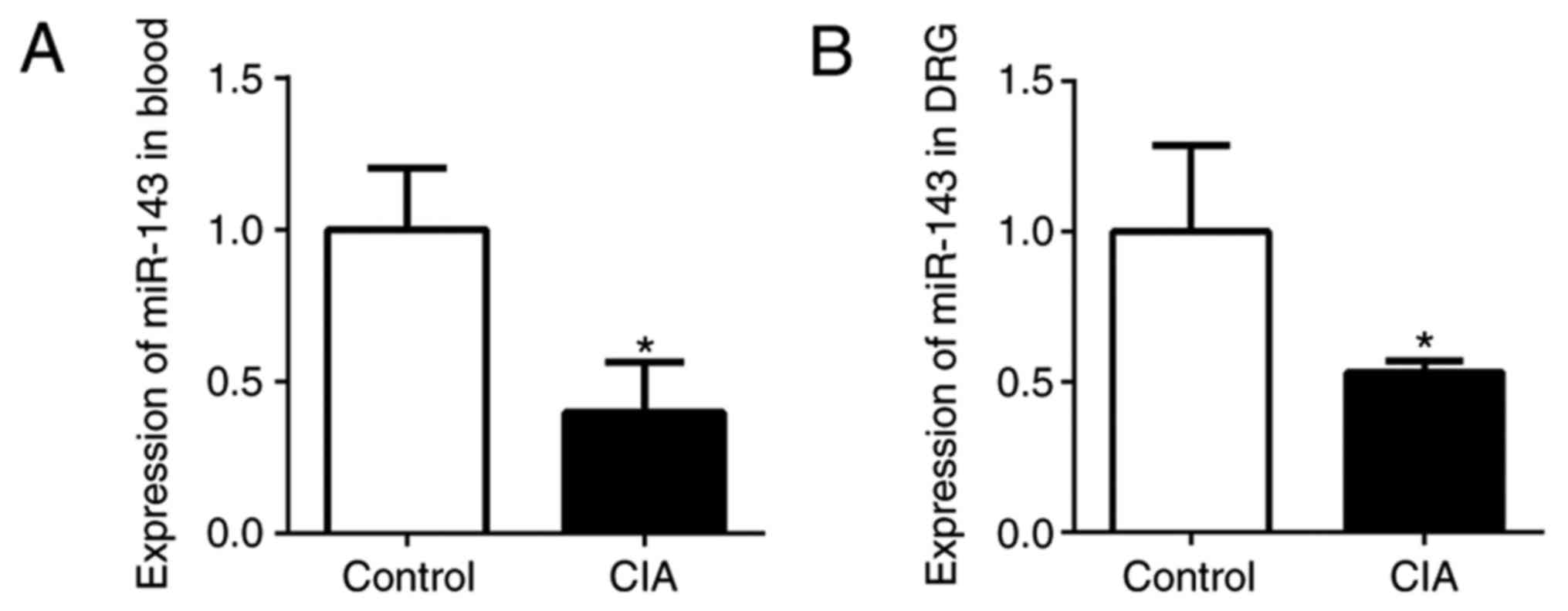
miR-143-3p expression in the
peripheral blood and DRG
In the authors' previous studies, miRs in CIA mice
and normal mice were screened, revealing that miR-143-3p was
expressed at a low level (9,10).
In the present study, miR-143-3p expression in the peripheral blood
and DRG was detected by RT-qPCR. The relative expression of
miR-143-3p in the peripheral blood of CIA mice was significantly
lower compared with the normal control mice (P<0.05; Fig. 2A). In addition, miR-143-3p
expression in the DRG tissue of the CIA mice was also significantly
downregulated (P<0.05; Fig.
2B). These results further confirmed that low miR-143-3p
expression was an important feature of the CIA mouse model.
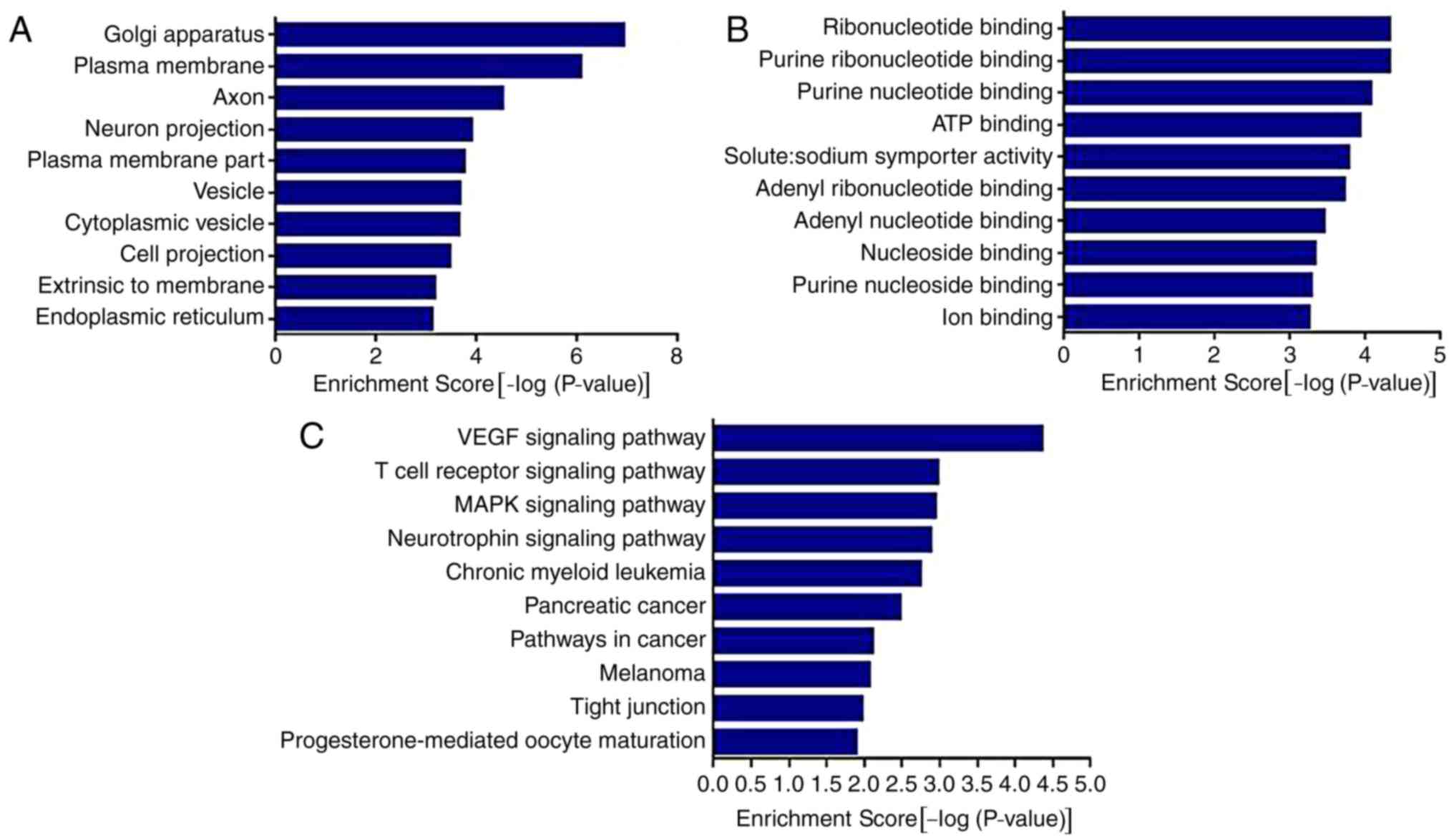
Prediction of pain-associated target
genes of miR-143
The target genes of miR-143 were predicted and
analyzed. The miR-target gene regulatory network is exhibited in
Fig. 3. A total of 1,305 genes,
including mitogen-activated protein kinase (MAPK)7, MAPK3, COX2,
matrix metalloproteinase and TNF, were predicted to be the target
genes of miR-143. Using the gene database of the Pain Genetics Lab,
55 pain-associated genes were obtained as potential target genes of
miR-143 (Table I).
 | Table I.Potential pain-associated target genes
of miR-143. |
Table I.
Potential pain-associated target genes
of miR-143.
| Gene | Protein name | Protein acronym |
|---|
| Accn1 | Amiloride-sensitive
cation channel 1, neuronal (degenerin) | ASIC2 |
| Adcy1 | Adenylatecyclase
1 | AC1 |
| Adra1d | Adrenergic
receptor, α1d | a1D-AR |
| Adra2c | Adrenergic
receptor, α2c | a2C-AR |
| Adrbk2 | Adrenergic receptor
kinase, β2 | GRK3 |
| Agtr2 | Angiotensin II
receptor, type 2 | AT2R |
| Bace1 | β-site APP cleaving
enzyme 1 | – |
| Cacnb3 | Calcium channel,
voltage-dependent, β3 subunit | b3 |
| Calb1 | Calbindin1 | Calb-1 |
| Ccr5 | Chemokine (C-C
motif) receptor 5 | CCR5 |
| Cd274 | CD274 antigen | B7-H1 |
| Cd40 | CD40 antigen | CD40 |
| Chrna4 | Cholinergic
receptor, nicotinic, α polypeptide 4 | Acra4 |
| Clock | Circadian locomoter
output cycles kaput | CLOCK |
| Cnr1 | Cannabinoid
receptor 1 (brain) | CB1 |
| Cxcr3 | Chemokine (C-C-C
motif) receptor 3 | CXCR3 |
| Dicer1 | Dcr-1 homolog
(Drosophila) | – |
| Dlg2 | Discs, large
homolog 2 (Drosophila) | – |
| Drd1a | Dopamine receptor
D1A | DRD1 |
| Foxn1 | Forkhead box
N1 | nu |
| Gabbr1 | γ-aminobutyric acid
(GABA-B) receptor, 1 | GABA-B(1) |
| Gad2 | Glutamic acid
decarboxylase 2 | GAD65 |
| Gja1 | Gap junction
protein, α1 | Cx43 |
| Gria1 | Glutamate receptor,
ionotropic, AMPA1 (α1) | GluR-1 |
| Ifngr1 | Interferon γ
receptor 1 | IFNgR |
| Ikbke | Inhibitor of κB
kinase ε | IKKepsilon |
| Il1rap | Interleukin 1
receptor accessory protein | IL-1RAcP |
| Kcna1 | Potassium
voltage-gated channel, shaker-related subfamily, member 1 | Kv1.1 |
| L1cam | L1 cell adhesion
molecule | L1 |
| Mgll | Monoglyceride
lipase | MAGL |
| Mmp24 | Matrix
metalloproteinase 24 | MT5-MMP |
| Mrgpre | MAS related GPR,
member E | MrgE |
| Nf1 | Neurofibromatosis
1 | Nf-1 |
| Ngfr | Nerve growth factor
receptor (TNFR superfamily, member 16) | p75 |
| Nlgn2 | Neuroligin 2 | NL2 |
| Nptx1 | Neuronal pentraxin
1 | NP1 |
| Npy | Neuropeptide Y | NPY |
| Nts | Neurotensin | NT |
| Ntsr1 | Neurotensin
receptor 1 | NT1R |
| Pcsk2 | Proprotein
convertase subtilisin/kexin type 2 | PC2 |
| Pik3cg |
Phosphoinositide-3-kinase, catalytic, γ
polypeptide | PI3Kg |
| Plp1 | Proteolipid protein
(myelin) 1 | Plp |
| Ppp1r9b | Protein phosphatase
1, regulatory subunit 9B | – |
| Ptgdr | Prostaglandin D
receptor | DP |
| Ptgfr | Prostaglandin F
receptor | FP |
| Ptgs2 |
Prostaglandin-endoperoxide synthase 2 | COX-2 |
| Rabggta | Rab geranylgeranyl
transferase, a subunit | gm |
| Slc12a6 | Solute carrier
family 12, member 6 | KCC3 |
| Slc6a2 | Solute carrier
family 6 (neurotransmitter transporter, noradrenalin), member
2 | NET |
| Slc6a4 | Solute carrier
family 6, member 4 | 5-HTT |
| Stx1a | Syntaxin 1A
(brain) | HPC-1 |
| Tacr1 | Tachykinin receptor
1 | NK1R |
| Tnf | Tumor necrosis
factor | TNF-α |
| Vip | Vasoactive
intestinal polypeptide | VIP |
The results of the GO analysis demonstrated that the
genes were associated with ‘Golgi apparatus’, ‘plasma membrane’,
‘axon’, ‘neuron projection’, ‘vesicle’, ‘cytoplasmic vesicle’ and
‘endoplasmic reticulum’. The results also demonstrated that the
genes were associated with various molecular functions (Fig. 4A), including ‘ribonucleotide
binding’, ‘purine ribonucleotide binding’, ‘ATP binding’, ‘solute:
sodium symporter activity’ and ‘ion binding’.
The results of the KEGG analysis demonstrated that
miR-143 was involved in the regulation of multiple pathways and its
target genes were significantly enriched in ‘VEGF signaling
pathway’, ‘T cell receptor signaling pathway’, ‘MAPK signaling
pathway’, ‘neurotrophin signaling pathway’, ‘pathways in cancer’
and other signaling pathways (P<0.01).
Following this, Ptgs2, Mrgpre, Ptgdr and Tnf were
selected as associated target genes for further verification, based
on the results of the GO and KEGG analysis and the factors in RA
inflammatory pain.
Effects of miR-143 on pain-associated
target genes
To determine the association between miR-143 and
target genes, DRG cells were cultured and transfected with
Lipofectamine RNAiMAX transfection reagent and the miR-143-3p mimic
NC-FAM. The transfected cells were observed using microscopy at 6,
12, 24, 48 and 72 h. As demonstrated in Fig. 5A and B, DRG cells were successfully
transfected, and the transfection efficiency peaked (~70%) at 48 h.
miR-143-3p expression significantly decreased following miR-143
inhibitor transfection for 48 h (P<0.01), whereas it increased
3.5-fold following miR-143 mimic transfection (P<0.01; Fig. 5C and D), verifying the effects of
inhibitor or mimic transfection.
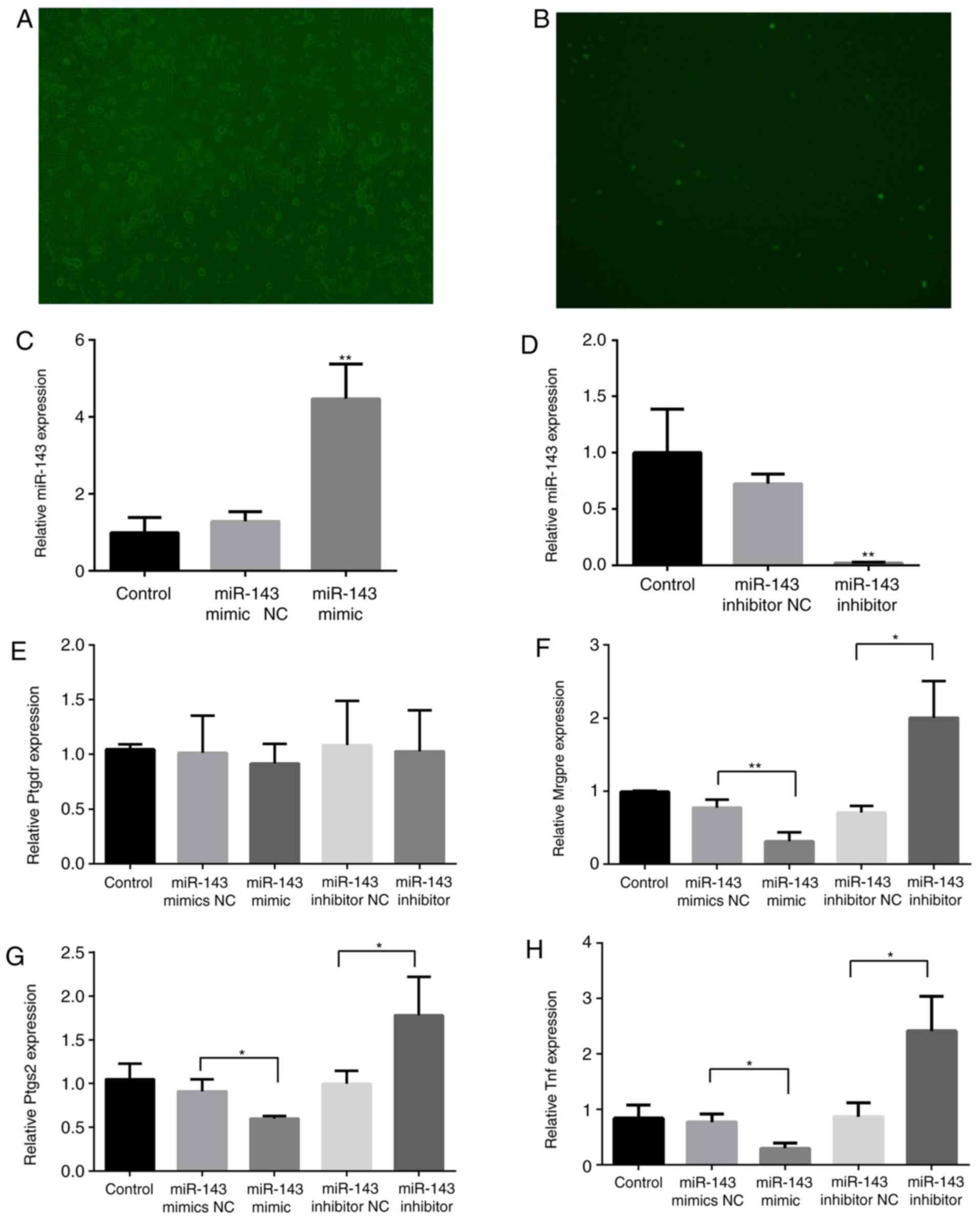 | Figure 5.Effects of miR-143 on pain-associated
target genes. Mouse DRG cells were transfected with Lipofectamine
RNAiMAX transfection reagent and the miR-143-3p mimic
NC-carboxyfluorescein. (A) Bright field (magnification, ×100). (B)
Fluorescence microscopy (magnification, ×100). RNA was extracted
from the DRG cells 48 h following transfection with an miR-143-3p
inhibitor or mimic. (C) miR-143-3p expression following
transfection with the mimic. (D) miR-143-3p expression following
transfection with the inhibitor. (E) Ptgdr expression, (F) Mrgpre
expression, (G) Ptgs2 expression and (H) Tnf expression.
*P<0.05, **P<0.01 vs. NC group. miR, microRNA; TNF, tumor
necrosis factor; DRG, dorsal root ganglion; Mrgpre, MAS related GPR
family member E; Ptgdr, prostaglandin D2 receptor; Ptgs2,
prostaglandin-endoperoxide synthase 2; NC, negative control. |
Subsequently, the expression of Mrgpre, Ptgs2, Ptgdr
and Tnf in DRG cells was detected. As illustrated in Fig. 5E-H, the expression of Mrgpre, Ptgs2
and Tnf was significantly inhibited (P<0.05) following
miR-143-3p mimic transfection, although it was significantly
promoted following miR-143-3p inhibitor transfection (P<0.05).
The mimic and inhibitor transfections did not markedly affect Ptgdr
expression.
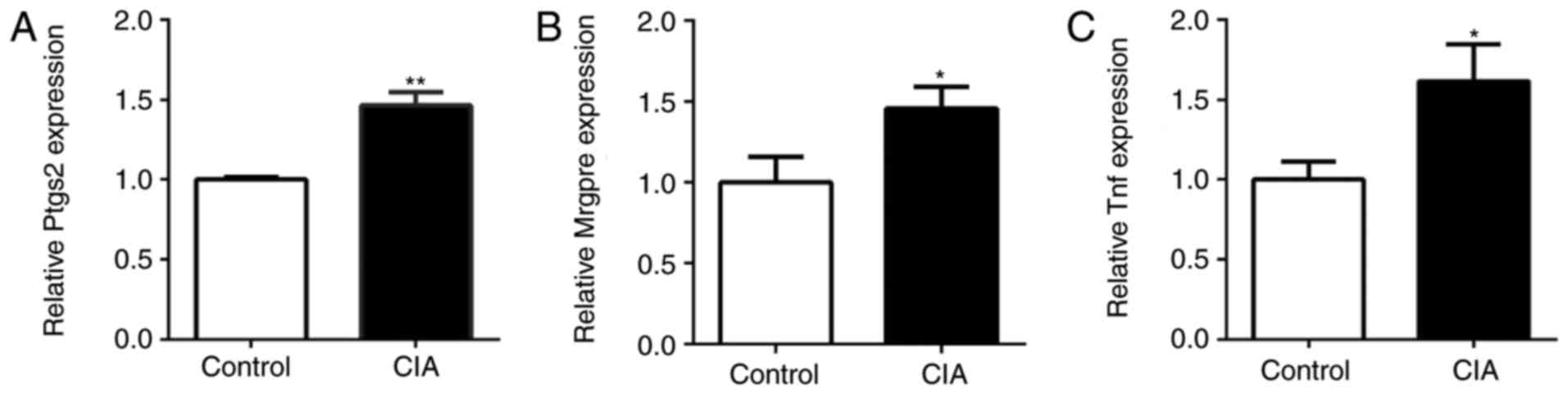
Expression of pain-associated genes in
the DRG of CIA mice
The expression of Ptgs2, Mrgpre and Tnf in the DRG
of CIA and control mice was also detected by RT-qPCR. Compared with
the control mice, the expression of Ptgs2, Mrgpre and Tnf in the
DRG of CIA mice was significantly upregulated (P<0.05; Fig. 6). The expression of miR-143-3p was
negatively associated with the expression of Mrgpre, Ptgs2 and Tnf
in CIA mice, being consistent with the results of the in
vitro experiments.
Discussion
The pain of RA patients is associated with multiple
complex mechanisms, including inflammation, peripheral and central
pain processing, and joint structure damage (18). Therefore, it may be useful for
effective treatment to investigate the mechanisms of pain. In this
study, the role of miR-143 in RA pain was investigated and the
mechanism by which miR-143 may regulate the pain of CIA is
described for the first time, to the best of the authors'
knowledge.
The CIA model has a similar pathological mechanism
to that of human RA (19) and it
is the most widely used mouse model for RA (20). CII is the principal constituent
collagen form in articular cartilage, and immunoreactivity to CII
maybe identified in certain patients with RA. Immunization with CII
leads to the development of severe polyarticular arthritis mediated
by immune mechanisms. The autoimmune response involves CII-specific
T cells and B cells and their products, proinflammatory cytokines
and anti-CII antibodies (21).
Thus, the CIA model may mimic the responses of RA pain more
accurately compared with other pain models. Considering that
different strains of mice may have a number of different symptoms,
two strains of mice were tested, and it was demonstrated that they
were subjected to apparent CIA. MWT and TWL are the common indices
that reflect pain sensitivity to mechanical and thermal stimuli. In
the present study, the MWT and TWL values of DBA/1 and C57BL/6 CIA
mice decreased, demonstrating that the CIA animal models exhibited
pain behaviors. Furthermore, DBA/1 CIA mice underwent increased
hyperalgesia compared with the C57BL/6 mice. Also, the levels of
PGE2 and TNF-α increased in the CIA mice. The results of
the present study also confirmed the increase in PGE2
and TNF-α levels in CIA mice. As a well-known mediator of
inflammation and pain, PGE2 serves a key role in the
occurrence and maintenance of RA pain and hyperalgesia (22). TNF-α, an essential inflammatory
factor for RA (23), is able to
induce the release of bradykinin, promote the release of substance
P from nerve terminal receptors and accelerate the synthesis of
PGE2 through COX activation (24,25).
Therefore, increasing inflammatory mediators may be one of the
mechanisms underlying the pain response. DBA/1 CIA mice exhibited
increased levels of inflammatory mediators compared with C57BL/6
mice. Accordingly, DBA/1 mice were selected to construct the CIA
model for further investigation.
Low-level expression of miR-143 in the peripheral
blood of patients with RA and CIA mice has been previously
demonstrated (9). In addition,
miR-143 has been associated with inflammatory pain responses
(11,12). In the present study, RT-qPCR
demonstrated that miR-143 was expressed at a low level in the
peripheral blood of CIA mice. Considering the role of the DRG in
pain conduction, miR-143 expression was also measured, and the
results demonstrated that miR-143 may be an important modulator of
the responses to CIA pain.
Bioinformatics analysis also revealed that certain
potential target genes of miR-143 were pain-associated and others
were associated with the production of inflammatory mediators. As
suggested by GO and KEGG analysis, the cytokines may be synthesized
in the Golgi apparatus and may be transported in a soluble form via
the endoplasmic reticulum. Alternatively, they may be present in a
membrane-bound form or processed into an intracellular form and
transported in the cell. Given that Ptgs2, Ptgdr and Tnf have been
closely associated with the release of inflammatory pain mediators
in RA, they may be the miR-143-3p target genes for pain regulation.
In addition, pain conduction is also worthy of attention according
to the results of the GO analysis. For example, target-gene
associated axons, the output channels of the neurons, are the
principal signal transmission channels in the nervous system. The
analysis of the molecular functions also indicated that miR-143 may
regulate gene expression by influencing nucleotide binding. It may
also regulate the selective transport of sodium ions, which
maintain the cell excitability and signal transduction, and affect
peripheral and central nervous system sensitization. miR-143 may
also influence adenosine triphosphate (ATP) release and its
combination with the ATP receptor in the primary afferent, and
subsequently modulate the excitement of the DRG nociceptive
neurons. Among the potential genes, Mrgpre is selectively highly
expressed in the DRG and small neurons of the trigeminal ganglion,
which are closely associated with pain conduction, being involved
in the onset of chronic pain and hyperalgesia (26). Therefore, Mrgpre may be the target
gene of miR-143-3p.
To further clarify the target genes of miR-143-3p in
CIA pain responses, mouse DRG cells were cultured and transfected
with an miR-143-3p inhibitor or mimic. The expression of certain
potential genes in the transfected cells was compared, and it was
demonstrated that the expression of Mrgpre, Ptgs2 and Tnf was
negatively associated with miR-143-3p, although Ptgdr expression
was not notably affected. Therefore, Mrgpre, Ptgs2 and Tnf may be
the target genes of miR-143-3p. The results of the in vivo
experiment were similar. In detail, the expression of Mrgpre, Ptgs2
and Tnf increased in the DRG of CIA mice. Low miR-143-3p expression
in RA may induce upregulation of Mrgpre in nervous tissues,
therefore affecting the onset of hyperalgesia and pain conduction.
In addition, Ptgs2 and Tnf expression were also upregulated. Ptgs2
in DRG cells may facilitate the synthesis of PGE2
(27), activate the corresponding
receptors in the DRG and a variety of cellular signaling pathways,
augment the sensitivity of nociceptors to pain stimulation, and
finally cause hyperalgesia and allodynia (18,28).
Tnf mRNA may further regulate the release of TNF-α and subsequently
activate COX to enhance the regulation of PGEs (23,25).
Pham et al (12) also
reported that miR-143 degraded COX-2 mRNA and regulated PGE
production. Therefore, miR-143-3p may modulate the levels of
inflammatory algogenic substances (PGE2 and TNF-α) by
regulating its target genes. Similarly, the increasing levels of
PGE2 and TNF-α in CIA mice concurred with the above
analysis. Taken together, miR-143-3p regulated pain transduction
and the release of inflammatory pain mediators, thereby regulating
the complex mechanisms of RA pain.
In conclusion, CIA mice were subjected to
hyperalgesia and the release of inflammatory pain mediators. Low
expression of miR-143-3p negatively regulated pain-associated
target genes, including Mrgpre, Ptgs2 and Tnf, thereby affecting
the chronic inflammatory pain and neuropathic pain of RA. Seeking
drugs that regulate miR-143 may be useful for the treatment of RA
pain.
Acknowledgements
The authors would like to express their sincere
gratitude to Professor Zongxiang Tang for his constructive comments
and useful suggestions as well as his technical assistance for this
project.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81673937,
81573869 and 81573929) and the National Natural Science Foundation
for the Youth of Jiangsu Province (grant no. BK20161043).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LLZ and XPZ conceived the project and designed the
experiments. LLZ, YMZ, FYQ, CCY and DPY performed the experiments.
LLZ and YMZ analyzed the data and wrote the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All experimental protocols performed in this study
were approved by the Ethics Review Committee for Animal
Experimentation of Nanjing University of Chinese Medicine and were
in accordance with the Declaration of the National Institutes of
Health Guide for Care and Use of Laboratory Animals (14).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Heiberg T and Kvien TK: Preferences for
improved health examined in 1,024 patients with rheumatoid
arthritis: pain has highest priority. Arthritis Rheum. 47:391–397.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Walsh DA and Mcwilliams DF: Pain in
rheumatoid arthritis. Curr Pain Headache Rep. 16:509–517. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mcwilliams DF, Zhang W, Mansell JS, Kiely
PD, Young A and Walsh DA: Predictors of change in bodily pain in
early rheumatoid arthritis: An inception cohort study. Arthrit Care
Res (Hoboken). 64:1505–1513. 2012. View Article : Google Scholar
|
|
4
|
Leffler AS, Kosek E, Lerndal T, Nordmark B
and Hansson P: Somatosensory perception and function of diffuse
noxious inhibitory controls (DNIC) in patients suffering from
rheumatoid arthritis. Eur J Pain. 6:161–176. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gold MS and Gebhart GF: Nociceptor
sensitization in pain pathogenesis. Nat Med. 16:1248–1257. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bai G, Ambalavanar R, Wei D and Dessem D:
Downregulation of selective microRNAs in trigeminal ganglion
neurons following inflammatory muscle pain. Mol Pain. 3:152007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Aldrich BT, Frakes EP, Kasuya J, Hammond
DL and Kitamoto T: Changes in expression of sensoryorgan-specific
microRNAs in rat dorsal root ganglia in association with mechanical
hypersensitivity induced by spinal nerve ligation. Neuroscience.
164:711–723. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao J, Lee MC, Momin A, Cendan CM,
Shepherd ST, Baker MD, Asante C, Bee L, Bethry A, Perkins JR, et
al: Small RNAs control sodium channel expression, nociceptor
excitability, and pain thresholds. J Neurosci. 30:10860–10871.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu YM, Zhou LL, Peng XW, Geng S and Zhou
XP: Study of Qingluo Tongbi Compound treating rheumatoid arthritis
based on miRNA network. Chin J Immunol. 32:495–499. 2016.(In
Chinese).
|
|
10
|
Yuan CC, Geng SA, Zhu YM, et al:
Comparision of abnormal expression of miRNAsin peripheral blood of
rheumatoid arthritis patients and osteoclasts in rat and analysis
of MiRNAs. Chin J Immunol. 33:715–720. 2017.(In Chinese).
|
|
11
|
Cerdá-olmedo G, Mena-durán AV, Monsalve V
and Oltra E: Identification of a MicroRNA Signature for the
Diagnosis of Fibromyalgia. PLoS One. 10:e01219032015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pham H, Rodriguez CE, Donald GW, Hertzer
KM, Jung XS, Chang HH, Moro A, Reber HA, Hines OJ and Eibl G:
miR-143 decreases COX-2 mRNA stability and expression in pancreatic
cancer cells. Biochem Biophys Res Commun. 439:6–11. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tam Tam S, Bastian I, Zhou XF, Vander Hoek
M, Michael MZ, Gibbins IL and Haberberger RV: MicroRNA-143
expression in dorsal root ganglion neurons. Cell Tissue Res.
346:163–173. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals.
Publication no. 85-23 (rev.). 8th edition. The National Academies
Press; 327. pp. 963–965. Washington, DC: 2011
|
|
15
|
Brand DD, Kang AH and Rosloniec EF: The
mouse model of collagen-induced arthritis. Methods Mol Med.
102:295–312. 2004.PubMed/NCBI
|
|
16
|
Chaplan SR, Bach FW, Pogrel JW, Chung JM
and Yaksh TL: Quantitative assessment of tactile allodynia in the
rat paw. J Neurosci Meth. 53:55–63. 1994. View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schweinhardt P, Kalk N, Wartolowska K,
Chessell I, Wordsworth P and Tracey I: Corrigendum to Investigation
into the neural correlates of emotional augmentation of clinical
pain. Neuroimage. 56:3842011. View Article : Google Scholar
|
|
19
|
Billiau A and Matthys P: Collagen-induced
arthritis and related animal models: How much of their pathogenesis
is auto-immune, how much is auto-inflammatory. Cytokine Growth F R.
22:339–344. 2011. View Article : Google Scholar
|
|
20
|
Luross JA and Williams NA: The genetic and
immunopathological processes underlying collagen-induced arthritis.
Immunology. 103:407–416. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cook AD, Rowley MJ, Stockman A, Muirden KD
and Mackay IR: Specificity of antibodies to type II collagen in
early rheumatoid arthritis. J Rheumatol. 21:1186–1191.
1994.PubMed/NCBI
|
|
22
|
Walsh DA and Mcwilliams DF: Mechanisms,
impact and management of pain in rheumatoid arthritis. Nat Rev
Rheumatol. 10:581–592. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Junger H and Sorkin LS: Nociceptive and
inflammatory effects of subcutaneous TNFalpha. Pain. 85:145–151.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Weinblatt ME, Kremer JM, Bankhurst AD,
Bulpitt KJ, Fleischmann RM, Fox RI, Jackson CG, Lange M and Burge
DJ: A trial of etanercept, a recombinant tumor necrosis factor
receptor: Fc fusion protein, in patients with rheumatoid arthritis
receiving methotrexate. N Engl J Med. 340:253–259. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Boettger MK, Weber K, Grossmann D, Gajda
M, Bauer R, Bär KJ, Schulz S, Voss A, Geis C, Bräuer R and Schaible
HG: Spinal tumor necrosis factor α neutralization reduces
peripheral inflammation and hyperalgesia and suppresses autonomic
responses in experimental arthritis: A role for spinal tumor
necrosis factor α during induction and maintenance of peripheral
inflamma. Arthritis Rheum. 62:1308–1318. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cox PJ, Pitcher T, Trim SA, Bell CH, Qin W
and Kinloch RA: The effect of deletion of the orphan G- protein
coupled receptor (GPCR) gene MrgE on pain-like behaviours in mice.
Mol Pain. 4:22008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Basbaum AI, Bautista DM, Scherrer G and
Julius D: Cellular and molecular mechanisms of pain. Cell.
139:267–284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Julius D and Basbaum AI: Molecular
Mechanisms of nociception. Nature. 413:203–210. 2001. View Article : Google Scholar : PubMed/NCBI
|