Introduction
Breast cancer is mortally harmful to females
globally. Triple negative breast cancer (TNBC) exhibits poor
prognosis, including poor distant free survival and overall
survival rates (1). Given the lack of
specific treatment for patients with TNBC, individuals are usually
treated with conventional adjuvant chemotherapy (2), and do not experience sufficient curative
effects (2). The lack of estrogen,
progesterone and human endothelial growth factor receptor 2
receptors makes it difficult to optimize therapeutic management
(2). Investigation into novel and
specific agents is required to combat these problems.
The Janus kinase (JAK)2/signal transducer and
activator of transcription (STAT)3 signaling pathway mediates the
expression of various cytokines (3).
Normally, this pathway is activated in response to certain
cytokines and then regresses rapidly; however, in tumor cells,
including breast cancer, lung cancer, hepatocellular carcinoma,
pancreatic cancer, colorectal cancer, prostate cancer, ovarian
carcinoma and gastric cancer, it is constitutively activated in a
high level (4–11). Furthermore, the JAK2/STAT3 signaling
pathway may modulate the expression of numerous genes and regulate
various physiological functions. It targets a number of
anti-apoptotic proteins, namely, B cell lymphoma-extra-large
(Bcl-xL) and myeloid cell leukemia 1 (Mcl-1), in addition to cell
cycle regulatory molecules and proteins associated with
mitochondrial apoptosis [Bcl2 associated X protein (Bax), Bcl2
antagonist/killer (Bak) and Caspase 3] (12). Mushroom Ganoderma lucidum, a
traditional Chinese medicine, has been demonstrated to exhibit
anti-cancer effects (13). However,
the effective components have not been well studied. Ganoderic acid
A (GA-A) is one of the major bioactive Ganoderma triterpenoids
isolated from Ganoderma and has been revealed to have an
effect on lymphoma growth (14), as
well as in the promotion of cisplatin-induced cell death by
inhibiting the JAK-STAT3 signaling pathway (15). In addition, it suppresses cancer cell
proliferation and invasion and induces apoptosis in osteosarcoma
(16). However, the effects and
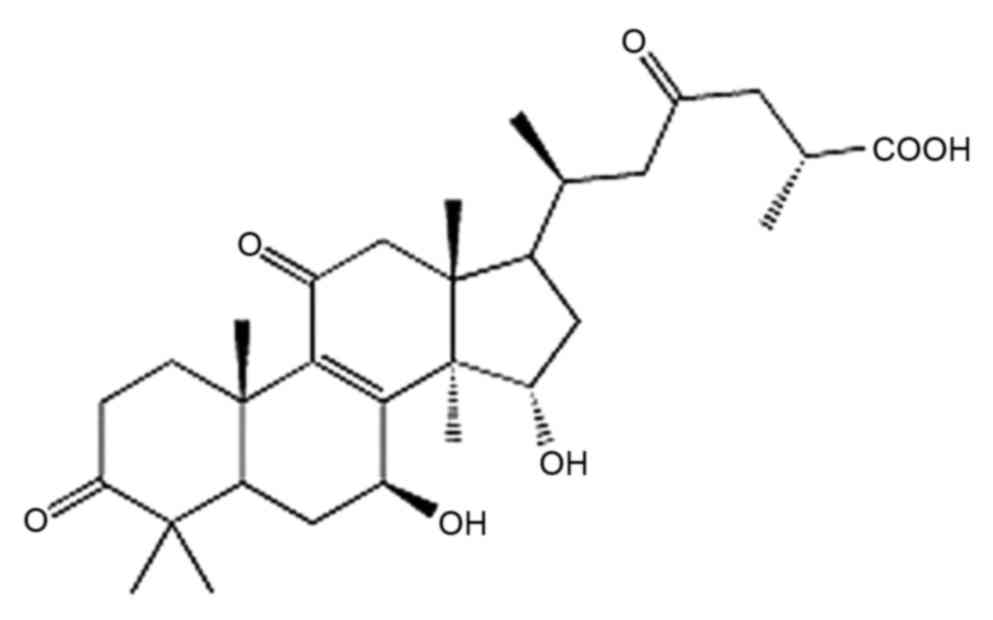
mechanisms of GA-A (Fig. 1) in human
breast cancer require further investigation.
The present study aims to investigate the anticancer
activity of GA-A in the human breast cancer cell line MDA-MB-231.
The function of the JAK2-STAT3 signaling pathway was the focus of
the present study and investigated the effects of GA-A in
mitochondrial apoptosis and the cell cycle.
Materials and methods
Materials
GA-A (96%) was purchased from Chengdu Must
Bio-technology Co., Ltd., (Chengdu, China), dissolved in
dimethylsulfoxide (DMSO) and maintained at 4°C. AG490, MTT, and
dichloro-dihydro-fluorescein diacetate (DCFH-DA) were obtained from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). The propidium
iodide (PI) staining kit and the JC-1 fluorescent dye were
purchased from Beyotime Institute of Biotechnology (Nanjing,
China).
Cell culture and reagents
MDA-MB-231 breast cancer cells were obtained from
the American Type Culture Collection (Manassas, VA, USA) and
maintained with Leibovitz L-15 (L-15) medium (pH=7.3; Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 100
mg/ml streptomycin, 100 IU/ml penicillin and 10% fetal bovine serum
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China). Cells were maintained at 37°C and 5% CO2, and
the medium was refreshed every 3 days. The GA-A was diluted in L-15
immediately prior to experiments. The control group was treated
with L-15 (0.01%) alone. Cells were first treated with GA-A (0.1,
0.2 and 0.4 mmol/l), then with 0.4 mmol/l GA-A and 20 µM AG490 at
37°C for 24 h. Cells were harvested at 24 or 48 h.
Cell viability
Cell viability was investigated using an MTT assay.
Cells (5×103) were treated with GA-A (0.01, 0.02, 0.05,
0.1, 0.2, 0.4, 0.6 and 0.8 mmol/l) for exactly 24 h then 20 ml MTT
at 5 mg/ml for 4 h at 37°C. The supernatant was then removed and
150 ml DMSO was added. After 15 min, the 490 nm optical density
value was measured. The results were presented as the ratio between
the control group and research group.
Transwell invasion assay
A Transwell chamber (8 µm pore polycarbonate;
Corning Incorporated, Corning, NY, USA) with Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA) was used in this experiment.
Cells were seeded (1×106) into the upper chambers of
24-transwell chamber wells and treated with GA-A (0.1, 0.2 and 0.4
mmol/l) at 37°C for 24 h. For the invasion assay, the cells were
plated in the upper chamber in the serum-free L-15 medium. The
medium containing 20% of FBS in the lower chamber served as a
chemoattractant. Following incubation for 24 h, cells in the upper
chambers were removed with a cotton swab. Subsequently, cells were
fixed with 100% methanol and then stained with 1% crystal violet in
2% ethanol for 10 min at room temperature. Under a Nikon light
microscope (×100), migrated cells were counted and images were
captured at five different fields of the chamber.
Cell apoptosis detection
Cells were seeded into 6-well plates
(5×105 cell/well) and incubated for 24 h prior to
treatment. GA-A (0, 0.1, 0.2 and 0.4 mmol/l, respectively) or DMSO
were added to the plate and incubated at 37°C for 1 day.
Subsequently, adherent cells and non-adherent cells were digested
in trypsin at 37°C for 2 min. The cell suspension was incubated
with 10 ml PI and 5 ml annexin V-fluorescein isothiocyanate (from
the kit aforementioned) for 10 min in the dark at 37°C and then
analyzed using a flow cytometer (BD Canto II) and Diva Software
v7.0 (BD Biosciences). The apoptotic index in Fig. 3 was calculated as: Apoptotic
cells/total cells ×100%.
Analysis of reactive oxygen species
(ROS)
Intracellular ROS production was investigated using
DCFH-DA. Cells were seeded at a density of 1×106
cells/well and cultured for 24 h. Cells were then harvested in
trypsin at 37°C for 2 min, washed with PBS for 5 min and
re-suspended in 500 µl PBS containing 20 µM DCFH-DA then incubated
at 37°C for 20 min in the dark. The cells were harvested and
analyzed by flow cytometry (BD Canto II) and Diva Software
v7.0.
Western blot analysis
The treated cells (1×107 cells/6 ml L-15
with 10% FBS in a 90-mm dish) were collected and washed twice using
cold PBS for 5 min. Cells were lysed in 200 µl lysis
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology). The lysate was incubated on ice for 30 min,
vortexed and centrifuged at 14,000 × g for 15 min at 4°C. The
supernatant was collected and protein concentration was determined
using a Bradford Assay. Following the addition of SDS-PAGE sample
loading buffer (Beyotime Institute of Biotechnology), the protein
samples (30 µg) underwent electrophoresis using a 10% SDS-PAGE and
were then transferred to a polyvinylidene fluoride membrane (EMD
Millipore, Billerica, MA, USA). After blocking for 4 h at room
temperature in a solution of 5% non-fat dry milk in Tris-buffered
saline containing 0.1% Tween-20, the membranes were incubated
overnight at 4°C with the primary antibodies against phosphorylated
(p)-JAK2 (catalog no. 3771), JAK2 (catalog no. 3230), p-STAT3
(catalog no. 9145), STAT3 (catalog no. 9139), Bcl-xL (catalog no.
sc-8392), Bak (catalog no. sc-517390), Mcl-1 (catalog no.
sc-12756), Bax (catalog no. sc-7480), Cytochrome c (catalog no.
sc-13156) and β-actin (catalog no. sc-47778) at a concentration of
1:1,000 in Tris-buffered saline with 0.1% Tween-20 containing 5%
non-fat dry milk. JAK2, p-JAK2, p-STAT3, STAT3 primary antibodies
were obtained from Cell Signaling Technology, Inc. (Danvers, MA,
USA). Bcl-xL, Bak, Mcl-1, Bax, Cytochrome c and β-actin primary
antibodies were purchased from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). Following washing four times for 5 min, the
membranes were incubated with a horseradish peroxidase-conjugated
secondary anti-rabbit IgG antibody (1:5,000 dilution; catalog no.
7074; Cell Signaling Technology, Inc.) at room temperature for 1 h
and were then washed six times for 10 min. Signals were detected
with an enhanced chemiluminescence detection kit (Applygen
Technologies, Inc., Beijing, China).
Statistical analysis
Statistical analysis was performed using SPSS 14.0
software (SPSS Inc., Chicago, IL, USA). Values presented as the
mean ± standard deviation and were analyzed using one-way analysis
of variance, followed by Tukey's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
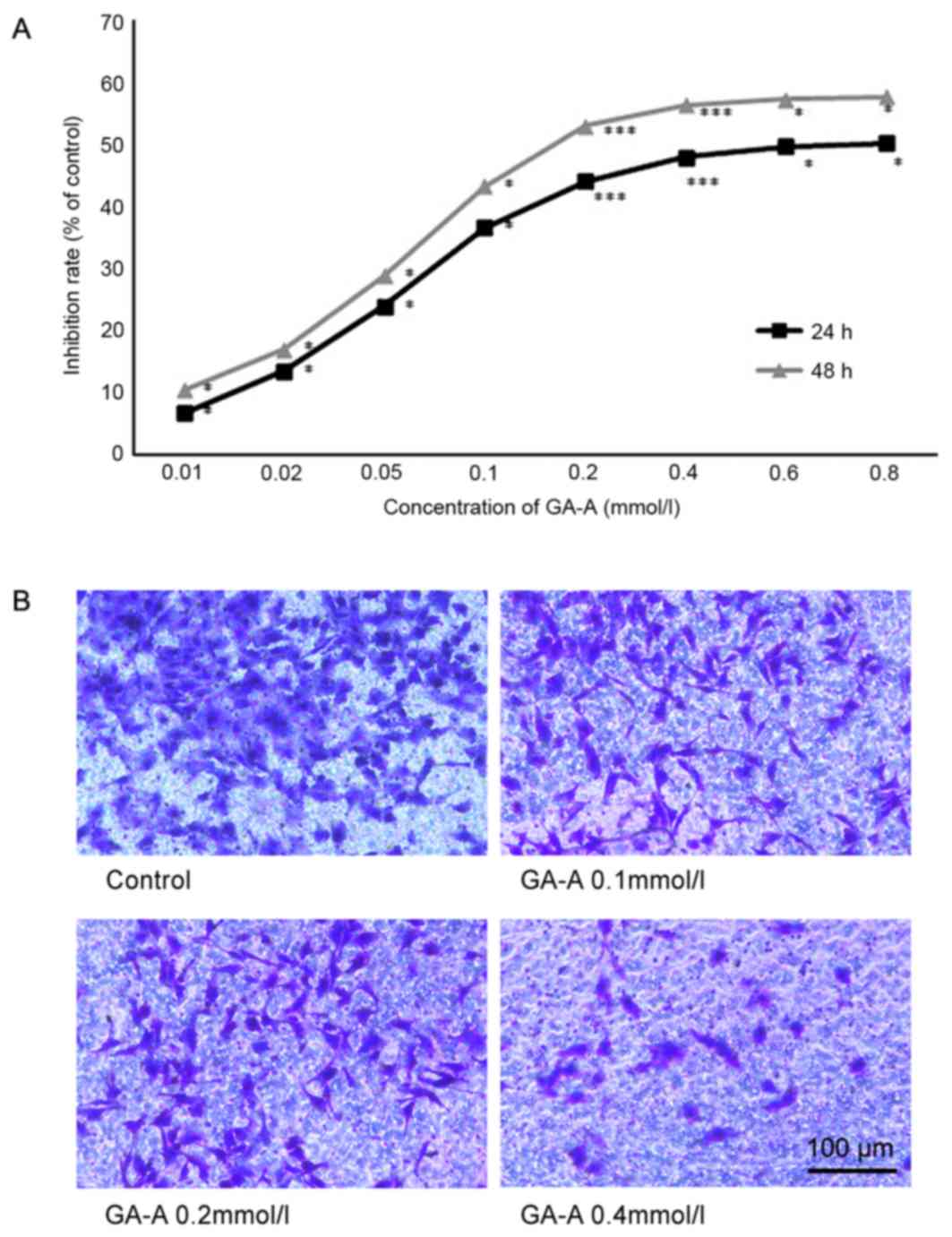
GA-A inhibits MDA-MB-231 cell
viability
MDA-MB-231 cells were stimulated by GA-A (0.1–0.8
mmol/l) to determine its effect on cell viability. GA-A
significantly decreased cell viability compared with the control
group, and the effects were not only dose dependent but also time
dependent (P<0.05; Fig. 2A). The
half-maximal inhibitory concentrations of GA-A at 24 and 48 h were
0.707 and 0.163 mmol/l, respectively.
GA-A inhibits MDA-MB-231 cell invasive
capacity
Based on the results of the Transwell invasion
assay, GA-A demonstrated an inhibitory effect on the invasion of
MDA-MB-231 cells. Fig. 2B reveals
that following incubation for 24 h, GA-A (0.1, 0.2 and 0.4 mmol/l)
markedly decreased the number of invasive cells. As GA-A
concentration increased, the MDA-MB-231 cells became less invasive.
These results were consistent with the Transwell invasion assay,
which indicated that GA-A suppressed the invasion of MDA-MB-231
cells.
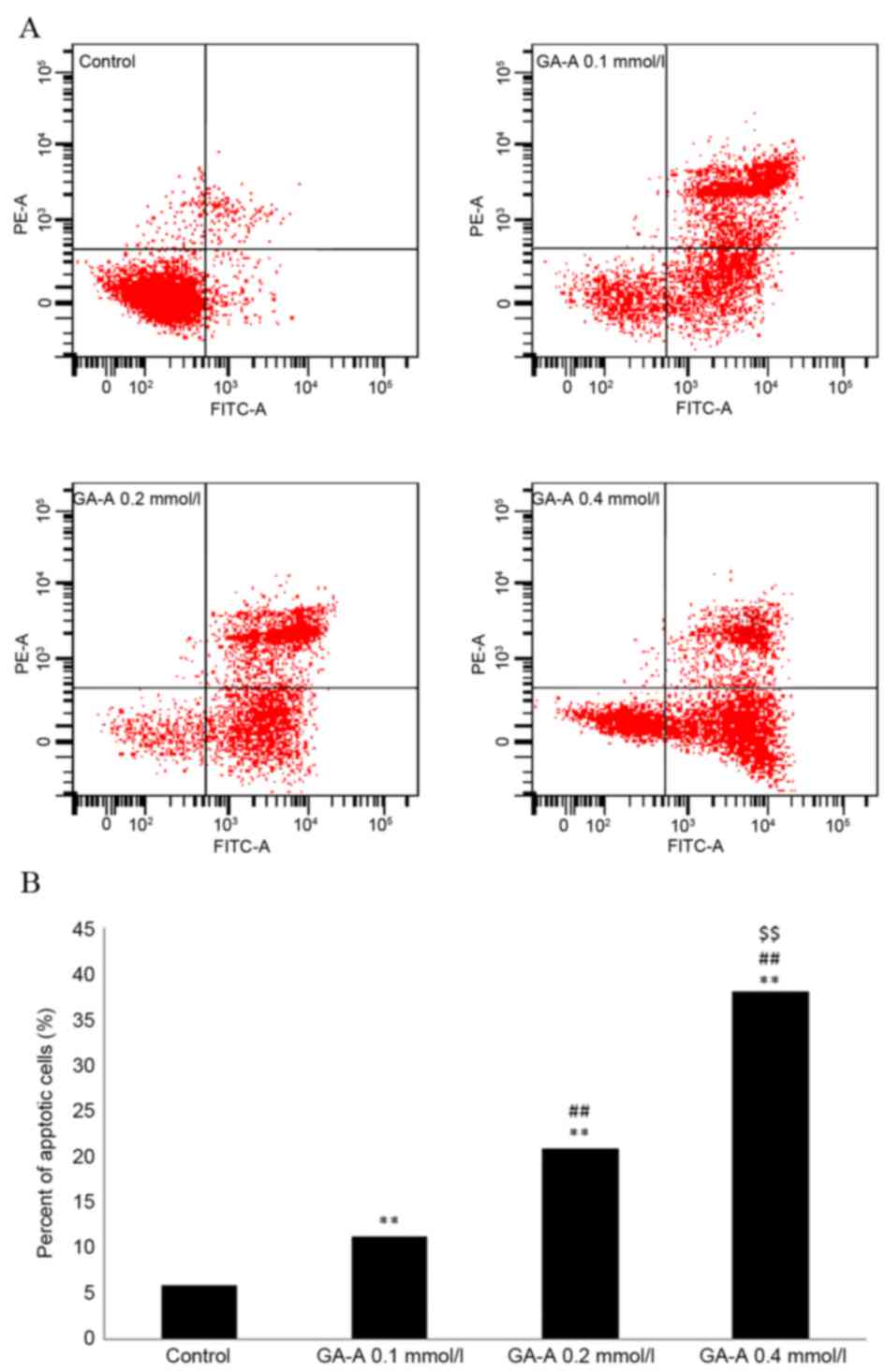
GA-A induces breast cancer cell
apoptosis
MDA-MB-231 cells were untreated or treated with GA-A
(0.1, 0.2 and 0.4 mmol/l) for 24 h. Following treatment with 0.1,
0.2 and 0.4 mmol/l of GA-A for 24 h, the apoptotic index increased
by 11.34±3.41, 20.89±3.32 and 38.13±3.91%, respectively (P<0.01,
vs. control). The effect of GA-A on apoptosis was dose-dependent.
These results indicate that GA-A induces apoptosis of MDA-MB-231
cells (Fig. 3B).
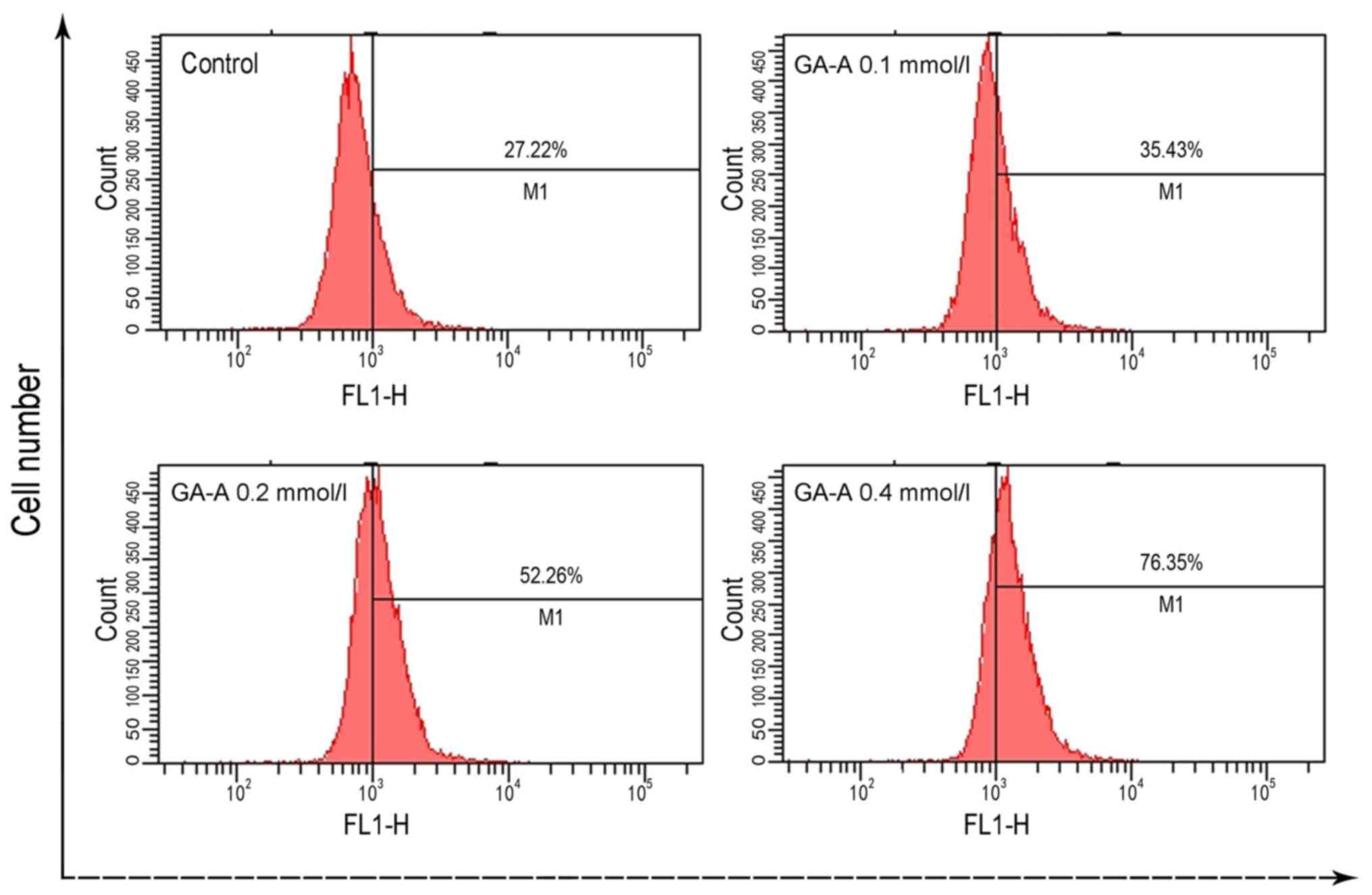
GA-A leads to ROS accumulation
As presented in Fig.
4, ROS level markedly increased following GA-A treatment. ROS
production following 0.4 mmol/l GA-A increased markedly from 27.22%
in the control group to 76.35%.
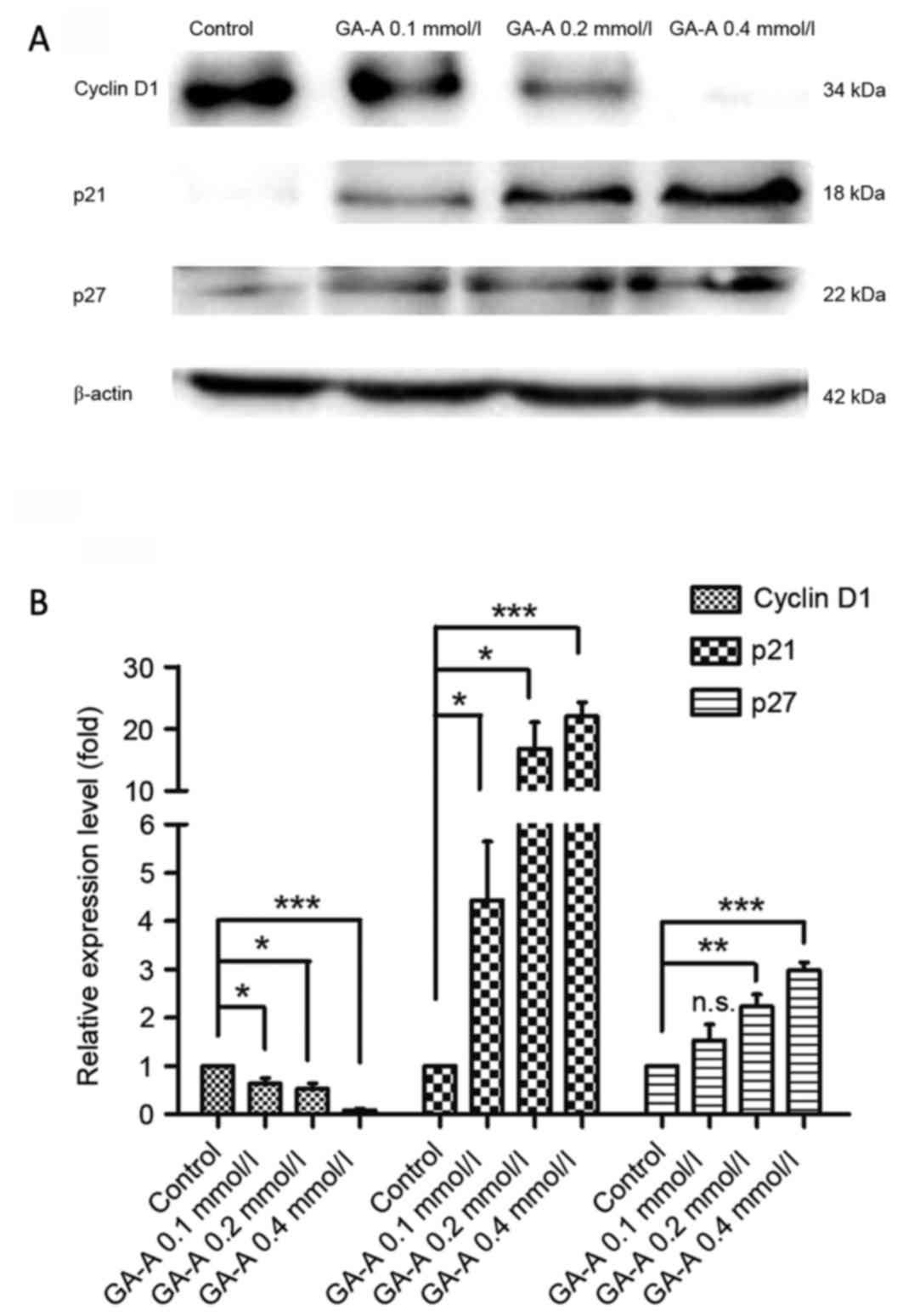
Effects of GA-A on cell
cycle-regulated protein expression
Western blot analysis was applied to determine
cyclin D1, p21 and p27 protein expression to further investigate
the characteristics of the observed G0-G1 phase control. It was
revealed that cyclin D1 expression significantly decreased compared
with the control, whereas p21 and p27 expression significantly
increased (P<0.05; Fig. 5), which
indicates that distinctions exist between cell-cycle-associated
protein, at least partially, leading to G0-G1 phase arrest by
GA-A.
Effects of GA-A on the JAK2/STAT3
signaling pathway and mitochondrial apoptosis
Western blot analysis was adopted to assess the
phosphorylated forms of JAK2 and STAT3 in breast cancer cells
stimulated by GA-A, from which the phosphorylation of these factors
was revealed to significantly decrease following GA-A treatment
(with the exception of p-JAK2 at 0.1 mmol/l GA-A; P<0.05). In
addition, the expression levels of Mcl-1 and Bcl-xL were
significantly downregulated by GA-A dose-dependently compared with
the control, in accordance with decreased phosphorylated JAK2 and
STAT3 (P<0.05; Fig. 6).
Additionally, GA-A treatment significantly enhanced the levels of
mitochondrial apoptotic pathway-associated proteins (Bak, Bax, and
cytosolic cytochrome C) compared with the control and in a dose
dependent manner (P<0.05), suggesting the mitochondrial
apoptosis occurred.
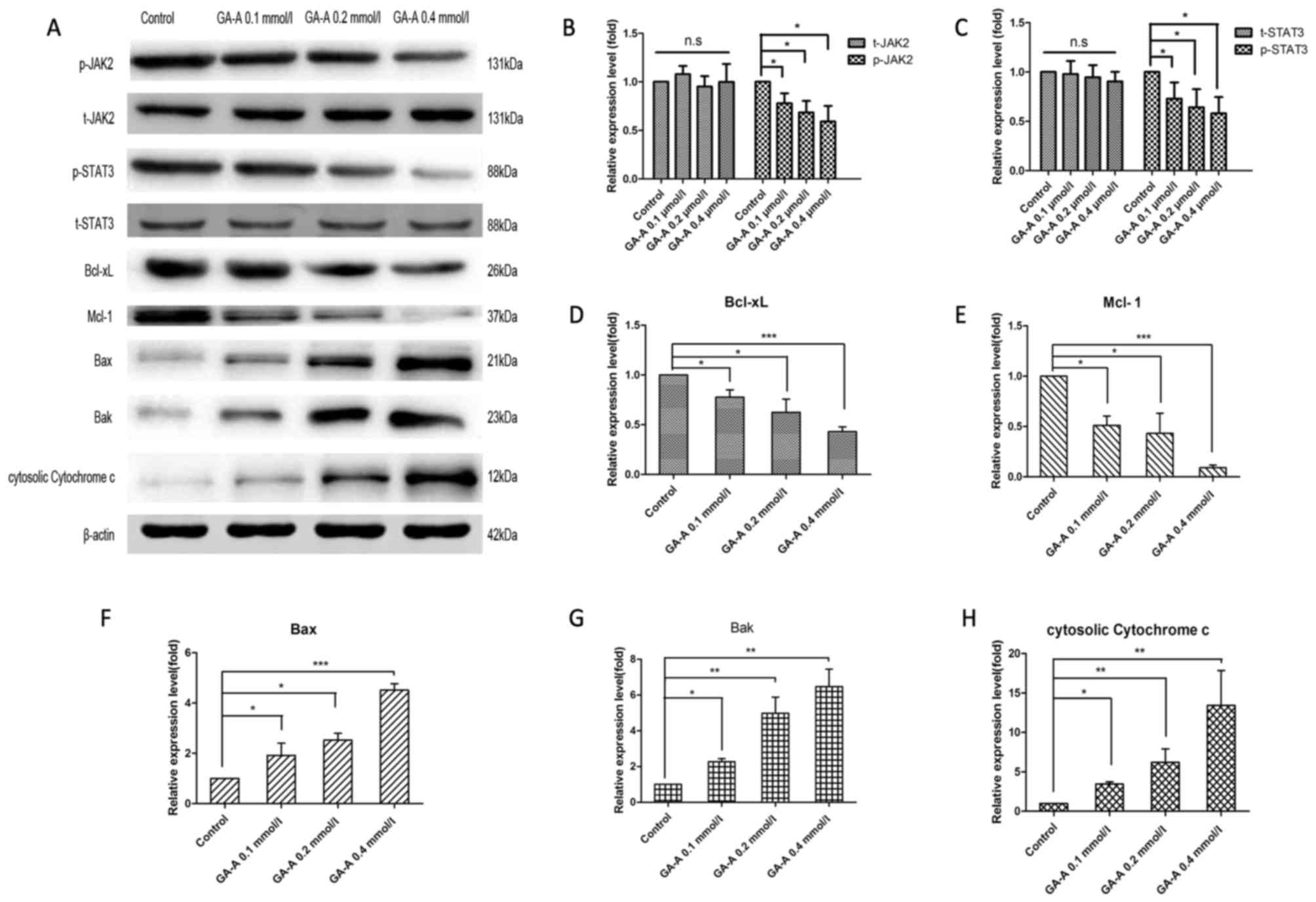 | Figure 6.GA-A affected JAK2/STAT3 signaling
pathway and mitochondrial apoptotic pathway associated proteins in
MDA-MB-231 cells. (A) The bands of western blot analysis. β-actin
was used as an internal control. (B) Statistical charts of the
relative expression levels of t-JAK2 and p-JAK2. (C) Statistical
charts of the relative expression levels of t-STAT3 and p-STAT3.
(D) Statistical charts of the relative expression levels of Bcl-xL.
(E) Statistical charts of the relative expression levels of Mcl-1.
(F) Statistical charts of the relative expression levels of Bax.
(G) Statistical charts of the relative expression levels of Bak.
(H) Statistical charts of the relative expression levels of
cytosolic Cytochrome c. ***P<0.001, **P<0.01, *P<0.05 vs.
GA-A-untreated control. GA-A, Ganoderic acid A; STAT, signal
transducer and activator of transcription; JAK, Janus kinase;
Bcl-xL, B cell lymphoma-extra-large; Mcl-1, myeloid cell leukemia
1; Bax, Bcl2 associated X protein; Bak, Bcl2 antagonist/killer;
n.s., not significant; p-, phosphorylated; t-, total. |
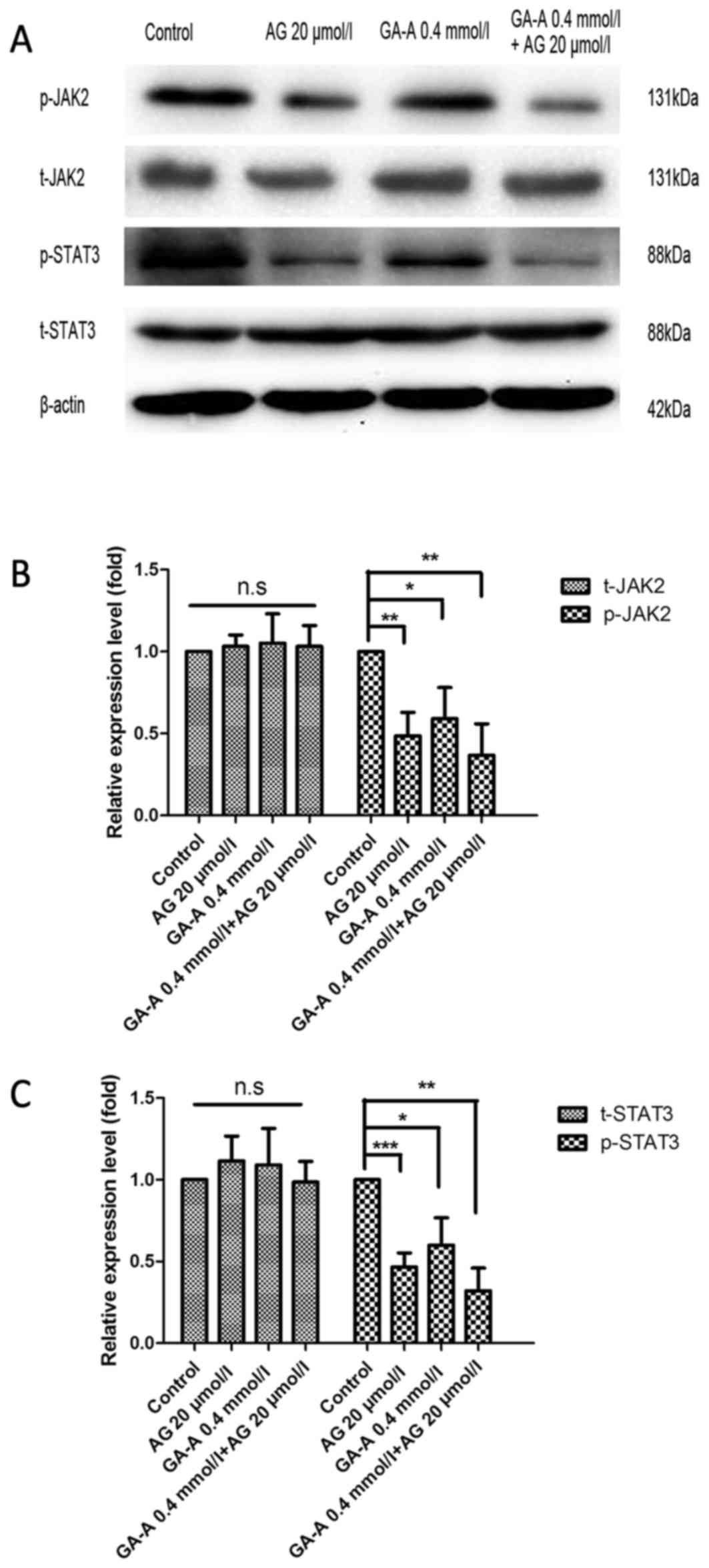
Effects of GA-A/AG490 combined
treatment on the JAK2/STAT3 signaling pathway
The combined treatment of GA-A and AG490 further
decreased JAK2/STAT3 expression levels compared with the control
(P<0.01; Fig. 7). This result
suggests that the anti-breast cancer effects of GA-A may involve
other mechanisms, including JAK2/STAT3. Thus, the result supports
the hypothesis that the antitumor effects of GA-A are associated
with certain molecules, including Bcl-xL, Bak, Mcl-1 and Bax, and
the JAK2/STAT3 signaling pathways.
Discussion
In the present study, it was demonstrated that GA-A
decreases cell viability, induces apoptosis and suppresses the
invasion of human MDA-MB-231 cells in vitro. Previously, the
effects of GA-A have been demonstrated in lymphoma, hepatocellular
carcinoma and osteosarcoma cells (12–14), in
which cellular proliferation, colony formation and invasive
behaviors were revealed to be suppressed by GA-A in osteosarcoma,
which is similar to the results of the present study in breast
cancer.
JAK/STAT transduces a series of signals that are
critical for physiological development or homeostasis. JAK1, JAK2,
JAK3 and thymidine kinase 2 constitute the JAK family, which is
involved in proliferation, migration and apoptosis (17). Constitutively activated JAKs
phosphorylate a number of substrates, including STAT3; and these
substrates are commonly associated with oncogenic signaling
pathways (18,19). STAT3 activation in regulating tumor
cell metastasis, proliferation, apoptosis, invasion, migration and
angiogenesis has been demonstrated in previous studies, revealing
its critical function (20–22). Distinct human cancer cell lines and
tumor tissues were previously studies to detect the persistent
activation of STAT3 through the use of various experiments
(23–29). Based on a number of previous studies,
the proliferation and survival of various types of cancer were
suppressed by the inhibition of STAT3 with a dominant negative form
of STAT3 or other inhibitors (21,30). The
results of the present study revealed that the anti-MDA-MB-231
function of GA-A worked through the JAK2/STAT3 signaling pathway,
and indicated that the activities of JAK2 and STAT3 were directly
prohibited by GA-A treatment, which also inhibited the expression
of Bcl-xL and Mcl-1. Additionally, cell viability was also further
decreased by GA-A when combined with AG490. In addition to
JAK2/STAT3, it was demonstrated that other mechanisms were involved
in the anti-MDA-MB-231 effect of GA-A. These results suggest that a
considerable number of molecules and pathways are associated with
the anticancer properties of GA-A.
There are two apoptotic signaling pathways in
mitochondria: One is the intrinsic, and the other is the extrinsic.
The former is more commonly observed in chemotherapy (31), including the important members
cytochrome c, caspase3, Bak and Bax (32). Suppression of the JAK2/STAT3 signaling
pathway induces apoptosis through the mitochondrial apoptotic
pathway (33,34). Additionally, one of the sources for
ROS is from the mitochondria during the process of apoptosis
(35). The results of the present
study demonstrate that GA-A significantly increased expression of
proteins associated with ROS generation and mitochondrial
apoptosis. This supported previous studies that suggested that for
human cancer cells, mitochondrial apoptosis is associated with
inhibiting JAK2/STAT3 (33,34).
As a cell cycle regulator, cyclin D1 regulates G1
cells entering the S-phase and serves a function as a cofactor for
a number of transcription factors (36). Cyclin D1 overexpression may contribute
to cancer incidence (37). p21 and
p27, as CDK inhibitors (CDKIs), attenuate cancer cell proliferation
with their upregulation (38). It has
previous been demonstrated that JAK2/STAT3 phosphorylation is
suppressed by WP1193 inhibiting glioma stem-like cell proliferation
and sphere formation; the reason for this effect partially lies in
decreased cyclin D1 and increased p21 expression (39). Research on colon cancer cells also
suggests that STAT3 controls the balance between CDK/cyclin and
CDKIs (34). The present study
demonstrated that GA-A treatment may result in delays in the cell
cycle in G0-G1 phase. Furthermore, a decrease was observed in
cyclin D1 expression but conversely an increase in p21 and p27
expression in MDA-MB-231 cells following GA-A treatment, compared
with the control. Collectively, the results of the present study
provide supporting evidence for JAK2/STAT3 in the counterpoise.
According to the result of the present study, GA-A
treatment inhibits MDA-MB-231 cell viability by inhibiting the
JAK2/STAT3 signaling pathway, and GA-A regulates various targets
that jointly generate the anti-MDA-MB-231 effects. These effects
include affecting the pathways involved in the mitochondrial
apoptosis and cell cycle control.
Acknowledgements
The present study was completed in the Oncology Key
Lab of the Heilongjiang Province Institution of Higher
Education.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YY performed the experiments, drafted the manuscript
and analyzed the data. HZ assisted with the data analysis. JCW
participated in the cell culture, reagents and viability
experiments. LQ participated in the cell apoptosis detection and
analysis of ROS. HL and LG participated in the western blot
analysis. XY, WL and JW assisted with the experiments. ZW and XL
participated in the study design and coordination and helped to
revised the manuscript. QZ was responsible for the study design and
final approval of the manuscript. All authors read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
References
|
1
|
Foulkes WD, Smith IE and Reis-Filho JS:
Triple-negative breast cancer. N Engl J Med. 363:1938–1948. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sun S, Zhao Y and Xu K: Post-adjuvant
chemotherapy for triple-negative breast cancer. Med Hypotheses.
90:74–75. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang X, Crowe PJ, Goldstein D and Yang JL:
STAT3 inhibition, a novel approach to enhancing targeted therapy in
human cancers (review). Int J Oncol. 41:1181–1191. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim MS, Lee WS, Jeong J, Kim SJ and Jin W:
Induction of metastatic potential by TrkB via activation of
IL6/JAK2/STAT3 and PI3K/AKT signaling in the breast cancer.
Oncotarget. 6:40158–40171. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu L, Guo L, Liang Y, Liu X, Jiang L and
Wang L: Curcumin suppresses stem-like traits of lung cancer cells
via inhibiting the JAK2/STAT3 signaling pathway. Oncol Rep.
34:3311–3317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao H, Guo Y, Li S, Han R, Ying J, Zhu H,
Wang Y, Yin L, Han Y, Sun L, et al: A novel anti-cancer agent
Icaritin suppresses hepatocellular carcinoma initiation and
malignant growth through the IL-6/Jak2/Stat3 pathway. Oncotarget.
6:31927–31943. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu X, Wang J, Wang H, Yin G, Liu Y, Lei X
and Xiang M: REG3A accelerates pancreatic cancer cell growth under
IL-6-associated inflammatory condition: Involvement of a
REG3A-JAK2/STAT3 positive feedback loop. Cancer Lett. 362:45–60.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang SW, Hu J, Guo QH, Zhao Y, Cheng JJ,
Zhang DS, Fei Q, Li J and Sun YM: AZD1480, a JAK inhibitor,
inhibits cell growth and survival of colorectal cancer via
modulating the JAK2/STAT3 signaling pathway. Oncol Rep.
32:1991–1998. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jorvig JE and Chakraborty A: Zerumbone
inhibits growth of hormone refractory prostate cancer cells by
inhibiting JAK2/STAT3 pathway and increases paclitaxel sensitivity.
Anticancer Drugs. 26:160–166. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gritsina G, Xiao F, O'Brien SW, Gabbasov
R, Maglaty MA, Xu RH, Thapa RJ, Zhou Y, Nicolas E, Litwin S, et al:
Targeted blockade of JAK/STAT3 signaling inhibits ovarian carcinoma
Growth. Mol Cancer Ther. 14:1035–1047. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Judd LM, Menheniott TR, Ling H, Jackson
CB, Howlett M, Kalantzis A, Priebe W and Giraud AS: Inhibition of
the JAK2/STAT3 pathway reduce gastric cancer growth in vitro and in
vivo. PLoS One. 9:e959932014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu Y, Wang L, Wu Y, Lv C, Li X, Cao X,
Yang M, Feng D and Luo Z: Pterostilbene exerts antitumor activity
against human osteosarcoma cells by inhibiting the JAK2/STAT3
signaling pathway. Toxicology. 304:120–131. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ruan W, Wei Y and Popovich DG: Distinct
responses of cytotoxic Ganoderma lucidum trierpenoids in human
carcinoma cells. Phytother Res. 29:1744–1752. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Radwan FF, Hossain A, God JM, Leaphart N,
Elvington M, Nagarkatti M, Tomlinson S and Haque A: Reduction of
myeloid-derived suppressor cells and lymphoma growth by a natural
triterpenoid. J Cell Biochem. 116:102–114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yao X, Li G, Xu H and Lü C: Inhibition of
the JAK-STAT3 signaling pathway by ganoderic acid A enhances
chemosensitivity of HepG2 cells to cisplatin. Planta Med.
78:1740–1748. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shao J, Li Z, Jiao G, Sun G and Zhou Z:
Ganoderic acid A suppresses proliferation and invasion and induces
apoptosis in human osteosarcoma cells. Nan Fang Yi Ke Da Xue Xue
Bao. 35:619–624. 2015.(In Chinese). PubMed/NCBI
|
|
17
|
Quintás-Cardama A and Verstovsek S:
Molecular pathways: JAK/STAT pathway: Mutations, inhibitors, and
resistance. Clin Cancer Res. 19:1933–1940. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Buchert M, Burns CJ and Ernst M: Targeting
JAK kinase in solid tumors: Emerging opportunities and challenges.
Oncogene. 35:939–951. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Khanna P, Chua PJ, Bay BH and Baeg GH: The
JAK/STAT signaling cascade in gastric carcinoma (Review). Int J
Oncol. 47:1617–1626. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
O'Shea JJ, Holland SM and Staudt LM: JAKs
and STATs in immunity, immunodeficiency, and cancer. N Engl J Med.
368:161–170. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bowman T, Garcia R, Turkson J and Jove R:
STATs in oncogenesis. Oncogene. 19:2474–2488. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu H, Pardoll D and Jove R: STATs in
cancer inflammation and immunity: A leading role for STAT3. Nat Rev
Cancer. 9:798–809. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Walker SR, Xiang M and Frank DA: STAT3
activity and function in cancer: Modulation by STAT5 and miR-146b.
Cancers (Basel). 6:958–968. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
He G and Karin M: NF-κB and STAT3-key
players in liver inflammation and cancer. Cell Res. 21:159–168.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Frank DA: STAT3 as a central mediator of
neoplastic cellular transformation. Cancer Lett. 251:199–210. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schroeder A, Herrmann A, Cherryholmes G,
Kowolik C, Buettner R, Pal S, Yu H, Müller-Newen G and Jove R: Loss
of androgen receptor expression promotes a stem-like cell phenotype
in prostate cancer through STAT3 signaling. Cancer Res.
74:1227–1237. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lin L, Liu A, Peng Z, Lin HJ, Li PK, Li C
and Lin J: STAT3 is necessary for proliferation and survival in
colon cancer-initiating cells. Cancer Res. 71:7226–7237. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim DY, Cha ST, Ahn DH, Kang HY, Kwon CI,
Ko KH, Hwang SG, Park PW, Rim KS and Hong SP: STAT3 expression in
gastric cancer indicates a poor prognosis. J Gastroenterol Hepatol.
24:646–651. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wei D, Le X, Zheng L, Wang L, Frey JA, Gao
AC, Peng Z, Huang S, Xiong HQ, Abbruzzese JL and Xie K: Stat3
activation regulates the expression of vascular endothelial growth
factor and human pancreatic cancer angiogenesis and metastasis.
Oncogene. 22:319–329. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kamran MZ, Patil P and Gude RP: Role of
STAT3 in cancer metastasis and translational advance. Biomed Res
Int. 2013:4218212013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Y, Yang F, Zheng W, Hu M, Wang J, Ma S,
Deng Y, Luo Y, Ye T and Yin W: Punica granatum (pomegranate) leaves
extract induces apoptosis through mitochondrial intrinsic pathway
and inhibits migration and invasion in non-small cell lung cancer
in vitro. Biomed Pharmacother. 80:227–235. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li X, Zhang Q, Cai L, Wang Y, Wang Q,
Huang X, Fu S, Bai J, Liu J, Zhang G and Qi J: Inhibitor of growth
4 induces apoptosis in human lung adenocarcinoma cell line A549 via
Bcl-2 family proteins and mitochondria apoptosis pathway. J Cancer
Res Clin Oncol. 135:829–835. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Su JC, Lin KL, Chien CM, Chuang PW, Chang
LS and Lin SR: Concomitant inactivation of the epidermal growth
factor receptor, phosphatidylinositol 3-kinase/Akt and Janus
tyrosine kinase 2/signal transducer and activator of transcription
3 signalling pathways in cardiotoxin III-treated A549 cells. Clin
Exp Pharmacol Physiol. 37:833–840. 2010.PubMed/NCBI
|
|
34
|
Du W, Hong J, Wang YC, Zhang YJ, Wang P,
Su WY, Lin YW, Lu R, Zou WP, Xiong H and Fang JY: Inhibition of
JAK2/STAT3 signalling induces colorectal cancer cell apoptosis via
mitochondrial pathway. J Cell Mol Med. 16:1878–1888. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bhardwaj M, Kim NH, Paul S, Jakhar R, Han
J and Kang SC: 5-Hydroxy-7-methoxyflavone triggers
mitochondrial-associated cell death via reactive oxygen species
signaling in human colon carcinoma cells. PLoS One.
11:e01545252016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang F, Wang Z, Yuan J, Wei X, Tian R and
Niu R: RNAi-mediated silencing of Anxa2 inhibits breast cancer cell
proliferation by downregulating cyclin D1 in STAT3-dependent
pathway. Breast Cancer Res Treat. 153:263–275. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cai Q, Lin J, Wei L, Zhang L, Wang L, Zhan
Y, Zeng J, Xu W, Shen A, Hong Z and Peng J: Hedyotis diffusa Willd
inhibits colorectal cancer growth in vivo via inhibition of STAT3
signaling pathway. Int J Mol Sci. 13:6117–6128. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhou Y, Zeng Z, Zhang W, Xiong W, Wu M,
Tan Y, Yi W, Xiao L, Li X, Huang C, et al: Lactotransferrin: A
candidate tumor suppressor-deficient expression in human
nasopharyngeal carcinoma and inhibition of NPC cell proliferation
by modulating the mitogen-activated protein kinase pathway. Int J
Cancer. 123:2065–2072. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sai K, Wang S, Balasubramaniyan V, Conrad
C, Lang FF, Aldape K, Szymanski S, Fokt I, Dasgupta A, Madden T, et
al: Induction of cell-cycle arrest and apoptosis in glioblastoma
stem-like cells by WP1193, a novel small molecule inhibitor of the
JAK2/STAT3 pathway. J Neurooncol. 107:487–501. 2012. View Article : Google Scholar : PubMed/NCBI
|