Introduction
Despite the overall decreasing rates of incidence
and mortality, gastric cancer remains the second most common cause
of cancer-related mortality worldwide (1). Currently, prognosis of patients with
gastric cancer remains dismal, as patients are often at advanced
stages of the disease at diagnosis. This emphasizes the need for
novel biomarkers for early diagnosis and more accurate prognosis of
gastric cancer. Hypermethylation is an important mechanism
underlying inactivation of tumor suppressor genes (TSGs), and has
been recognized as one of the most important markers for the
identification of novel TSGs and detection of cancer and prediction
of prognosis (2–4). Several TSGs including hMLH1, P16,
MGMT, APC and Ras association domain family 1 isoform A (RASSF1A)
have frequently been reported in gastric cancer (5–9). The
identification of novel TSGs that are silenced by tumor-specific
methylation in gastric cancer will considerably improve the
molecular diagnosis for gastric cancer.
RASSF family members including RASSF1–6 contain
C-terminal RA (of the RalGDS/AF-6 variety) and Sav/RASSF/Hippo
(SARAH) interaction domains. Several RASSF genes that encode
prominent tumor suppressors are frequently epigenetically silenced
in various types of cancer. The silenced genes are involved in cell
death regulation, cell cycle control and microtubule stability
(10).
Recently, four other proteins were added to the
RASSF family and renamed RASSF7–10; they are divergent and
structurally distinct from RASSF1–6, as they contain an RA domain
in the N-terminal but lack the SARAH domain (11). These N-terminal RASSF proteins
represent a new group of potential Ras effectors, which have
important biological functions (12). RASSF7 plays an essential role in the
regulation of microtube organization and mitotic progression
(11,13), and is upregulated in several types
of cancer (14–16). Expression of RASSF7 could also be
upregulated by hypoxia (17,18).
Upon stress, RASSF7/N-Ras could promote cell survival through the
inhibition of mitogen-activated protein kinase kinase 7
(MKK7)/c-Jun N-terminal kinase (JNK) mediated signaling pathway.
However, with prolonged stress, the RASSF7 protein is degraded so
as to allow cell death signaling pathways to be activated (19). Although there is currently no direct
evidence to support that expression of RASSF7 promotes tumor
formation, the evidence indicates that RASSF7 may act as an
oncogene in tumorigenesis. RASSF8, another NT-RASSF member
identified as a tumor suppressor in lung adenocarcinoma and male
germ cell tumor, plays an important role in suppressing tumor
metastasis by regulating the Wnt and NF-κB signaling pathways
(20–22). Notably, RASSF7 and RASSF8 have been
shown to be required for necroptosis, a regulated form of necrosis
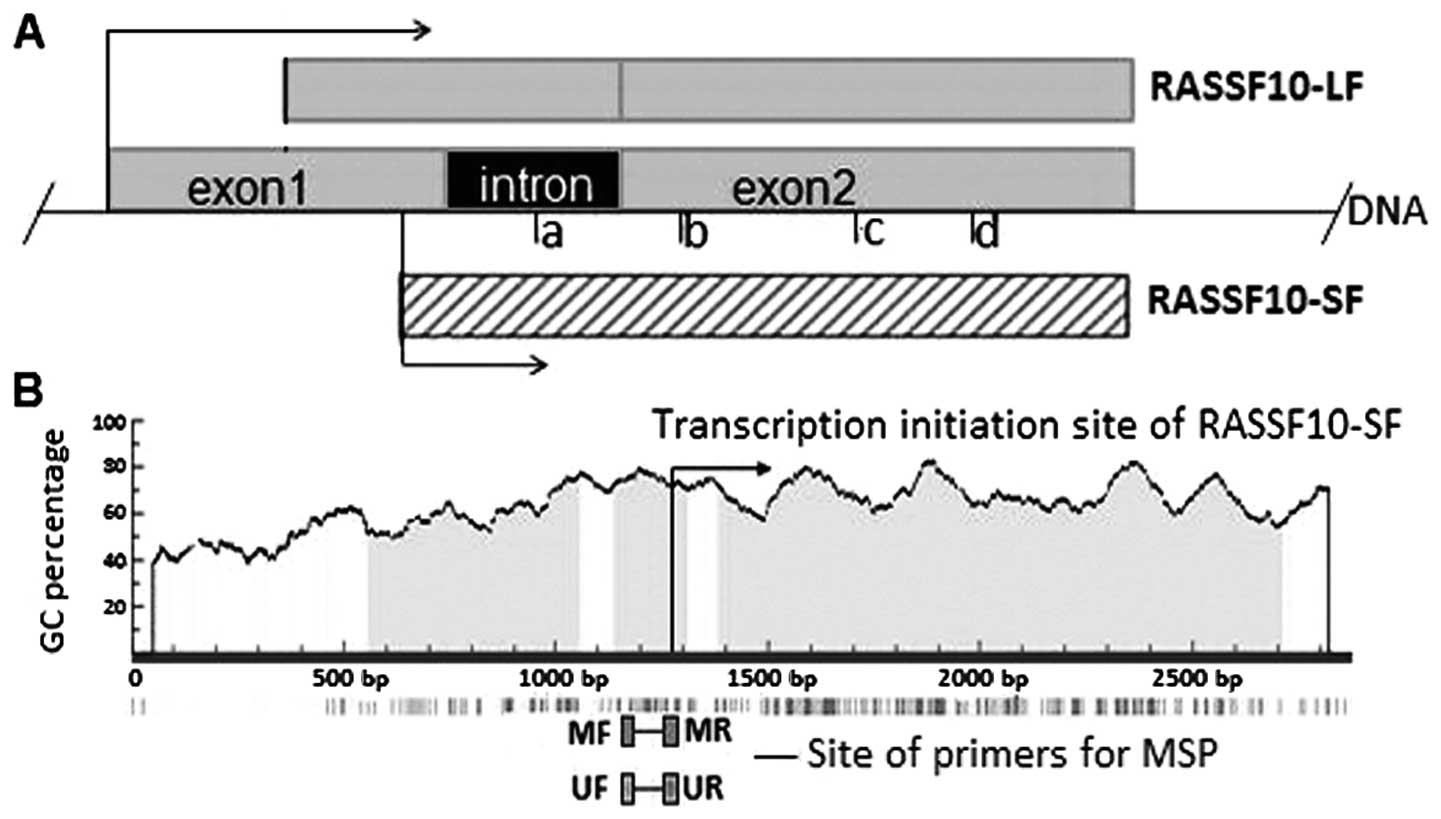
distinct from apoptosis, in mice (23). RASSF10 contains two isoforms, known
as long form (RASSF10-LF) and short form (RASSF10-SF), according to
different transcription initiation sites (TIS) as described in
Fig. 1A. RASSF10 has also been
identified as a candidate TSG and is frequently downregulated by
DNA hypermethylation in human cancers (24–27).
Direct evidence obtained from a glioma study confirmed the role of
RASSF10 in the suppression of tumor growth and cell proliferation
(26). It was shown that frequent
hypermethylation of RASSF10 promoter was associated with tumor
metastasis (25,27), and RASSF10 methylation is an
independent prognostic factor associated with poor progression-free
survival and overall survival. RASSF10 methylation occurred in
early stages of secondary glioblastoma development (26), suggesting that tumor-specific
methylation of RASSF10 might act as valuable biomarkers for the
detection of cancer and the prediction of prognosis.
To date, whether RASSF10 is downregulated through
DNA methylation in gastric cancer remains unknown. Therefore, in
the present study, we examined the expression of both RASSF7 and
RASSF10 in gastric cancer and performed a comparative analysis of
RASSF7 and RASSF10 expression. We also evaluated the status of
RASSF10 methylation in gastric cancer, using both
methylation-specific PCR (MSP) and sequencing, and analyzed the
correlation between RASSF10 methylation with clinicopathological
changes.
Materials and methods
Tissues and cell lines
Tumor specimens including 86 of gastric cancer, 1 of
pancreatic cancer and 1 of rectal cancer were obtained from
patients with gastric carcinoma who underwent surgery at the Cancer
Institute, China Medical University. All samples used for
methylation analysis included carcinoma and adjacent non-carcinoma
tissues. An additional 54 pairs of primary gastric carcinoma and
adjacent non-carcinoma tissues were used for expression analysis.
Six gastric cancer cell lines, SGC7901, BGC823, MGC803, AGS, MKN45
and HGC27, and one normal gastric mucosa cell line, GES-1, were
obtained from the Institute of Biochemistry and Cell Biology, China
Academy of Science, Shanghai, China. Cells were maintained in
RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with
10% fetal bovine serum (FBS) at 37°C with 5% CO2 and 95%
humidity, as recommended.
RT-PCR and quantitative real-time
PCR
Total RNA was isolated using TRIzol reagent
(Invitrogen) from the cultured cells and 54 pairs of gastric
carcinoma and adjacent non-carcinoma tissues for RT-PCR. To
determine the methylation status of RASSF10 in gastric cancer, the
MKN45 cell line was selected as a model, and was treated with DNA
demethylating agent 5-aza-2′-deoxycytidine (5-Aza-dC) (Sigma, St.
Louis, MO, USA) and histone deacetylase inhibitor trichostatin A
(TSA) (Sigma). The treatments were divided into 5 groups: i)
control, ii) 5-Aza-dC alone (5 μmol/l), iii) 5-Aza-dC alone (10
μmol/l), iv) TSA alone (0.3 μmol/l), and v) combination of 5-Aza-dC
(5 μmol/l) and TSA (0.3 μmol/l). Cells were treated for 72 h and
the medium containing 5-Aza-dC in groups ii, iii and v was changed
every 24 h. In group v, 5-Aza-dC was used for 48 h, followed by TSA
treatment for an additional 24 h, while in group iv, cells were
treated with TSA only for 24 h. After treatment, cells were
harvested for RNA extraction. To eliminate DNA contamination, 1 μg
of total RNA was incubated in a volume of total 10 μl with 1U DNase
I (Fermentas, Burlington, ON, Canada) in 1 μl 10× DNase I buffer.
After a 30-min incubation at 37°C, DNase I was inactivated by
adding 1 μl of 25 mmol/l EDTA and incubated at 65°C for an
additional 15 min. Reverse transcription reaction was performed
with PrimeScript™ RT reagent kit (Takara, Beijing, China) using 500
ng RNA from the above prepared RNA sample. The mRNA expression of
RASSF10 in a total of seven cell lines was determined by
quantitative real-time RT-PCR with SYBR® Premix Ex Taq™
II (Takara). GAPDH was used as an internal control of RNA integrity
and relative mRNA levels were assessed using the 2−ΔΔCT
method as previously described (28). To compare the expression of RASSF7
and RASSF10, the cDNA of 7 cell lines and 54 pairs of carcinoma and
adjacent non-carcinoma tissues was used for conventional RT-PCR,
and PCR products were separated by 2% agarose gel. The PCR reaction
was performed with appropriate primers for RASSF10 (primer c-d)
(24), RASSF7 and GAPDH as shown in
Table I. All samples were submitted
to PCR in a total 50 μl of reaction mixture, containing 5 μl of 10×
buffer, 5 μl of dNTP, 0.25 μl of Dream Taq DNA polymerase (5 U/μl)
(Fermentas), 1.0 μl each of the sense and antisense primers, 2.0 μl
of cDNA and 35.75 μl of double-distilled water. Reaction conditions
were: pre-denaturation at 95°C for 5 min, followed by 35 cycles of
95°C for 30 sec, 58°C for 30 sec, 72°C for 30 sec and final
extension at 72°C for 10 min.
 | Table IPrimers for RT-PCR and
methylation-specific PCR. |
Table I
Primers for RT-PCR and
methylation-specific PCR.
| Sequence | Product length
(bp) |
|---|
| RASSF10 (primer
c-d) | F:
5′-CCATGACCCAGGAGAAACAG-3′ | |
| R:
5′-TGCTGGCGAATTGTGTGGTC-3′ | 226 |
| RASSF10 (primer
a-b) | F:
5′-GGGAACAGGGCTAGTGCAG-3′ | |
| R:
5′-TCTCTTCCTGGCAGATCCAC-3′ | 183 |
| RASSF7 | F:
5′-AAGTGGTCATCGCACTAGCC-3′ | |
| R:
5′-TCAGGACAAACTGGACATCG-3′ | 159 |
| RASSF10-M | F:
5′-GGGTATTTTGGGTAGAGTTAGAGC-3′ | |
| R:
5′-AAACAAACTAAAAAACGACTACGAC-3′ | 126 |
| RASSF10-U | F:
5′-GGGTATTTTGGGTAGAGTTAGAGTG-3′ | |
| R:
5′-AAACAAACTAAAAAACAACTACAAC-3′ | 126 |
| GAPDH F: |
5′-CATGAGAAGTATGACAACAGCCT-3′ | |
| R:
5′-AGTCCTTCCACGATACCAAAGT-3′ | 113 |
RT-PCR with primer a-b
To determine the isoform of RASSF10 expression, we
designed one pair of primers in the consensus region of subtypes
(primer c-d), according to the difference between RASSF-LF and
RASSF-SF in mRNA, for which detailed information is shown in
Fig. 1A. To confirm the expression
of RASSF10, we used another pair of primers, in which the forward
primer was at the intron and the reverse primer was at the
consensus region (primer a-b; Fig.
1A, Table I). The primers were
verified using the cDNA from GES-1 cells as PCR template.
DNA extraction and sodium bisulfite
modification
Genomic DNA was extracted from frozen tissues and
cultured cells by the standard phenol/chloroform procedure.
Bisulphate modification was performed as previously described.
Briefly, 2 μg of genomic DNA in a volume of 50 μl was denatured by
NaOH (final concentration, 0.2 mol/l) for 30 min at 42°C. The
denatured DNA samples were treated with freshly prepared 30 μl of
hydroquinone (10 mmol/l) (Sigma) and 520 μl of sodium bisulphate
(3.9 mol/l; pH 5.0) (Sigma) at 55°C for 16 h, followed by a stop
reaction using NaOH (final concentration, 0.3 mol/l) at room
temperature for 5 min. Finally, modified DNA was recovered using
Wizard DNA Clean-Up System (Promega, Madison, WI, USA), according
to the manufacturer’s instructions and precipitated using a
combination of glycogen, ammonium acetate and ethanol, followed by
a wash using 70% ethanol. The pellet of recovered DNA was
resuspended in 15 μl TE buffer.
MSP
The methylation status of RASSF10 was determined by
MSP using bisulfite-converted DNA as template. Total reaction
mixture volume was 50 μl, containing 5 μl of 10× buffer, 5 μl of
dNTP, 0.25 μl of Dream Taq DNA polymerase (5 U/μl) (Fermentas), 1
μl each of the sense and antisense primers, 2.0 μl of DNA and 35.75
μl of double-distilled water. Reaction conditions were:
pre-denaturation at 95°C for 5 min, followed by 40 cycles of 95°C
for 30 sec, 56°C for 30 sec, 72°C for 30 sec, followed by a final
extension at 72°C for 10 min. The annealing temperature of
methylated and unmethylated reaction was at 56°C. The
methyltransferase SssI (New England Biolabs, Ipswich, MA,
USA)-treated and untreated cord blood cells from newborn were used
as a positive control for methylation or unmethylation,
respectively. Double-distilled water was used as the blank control.
Methylation-specific (RASSF10-M) and unmethylation-specific
(RASSF10-U) primers are listed in Table
I. The size of methylation and unmethylation PCR products was
126 bp, from the region −120 to +6 relative to the TIS of
RASSF10-SF. MSP products were verified by 2% agarose gel. The
accuracy of the verified MSP products was confirmed by
sequencing.
Statistical analysis
Statistical analysis was carried out using SPSS 13.0
software. Correlation of DNA methylation and clinicopathological
parameters was assessed using the Chi-square test. All reported
P-values were two-sided and a P-value <0.05 was considered to
indicate a statistically significant difference.
Results
Expression of RASSF7 and RASSF10 in
gastric cancer
To evaluate the expression of N-terminal RASSF genes
in gastric cancer, the mRNA level of RASSF10 in gastric cancer cell
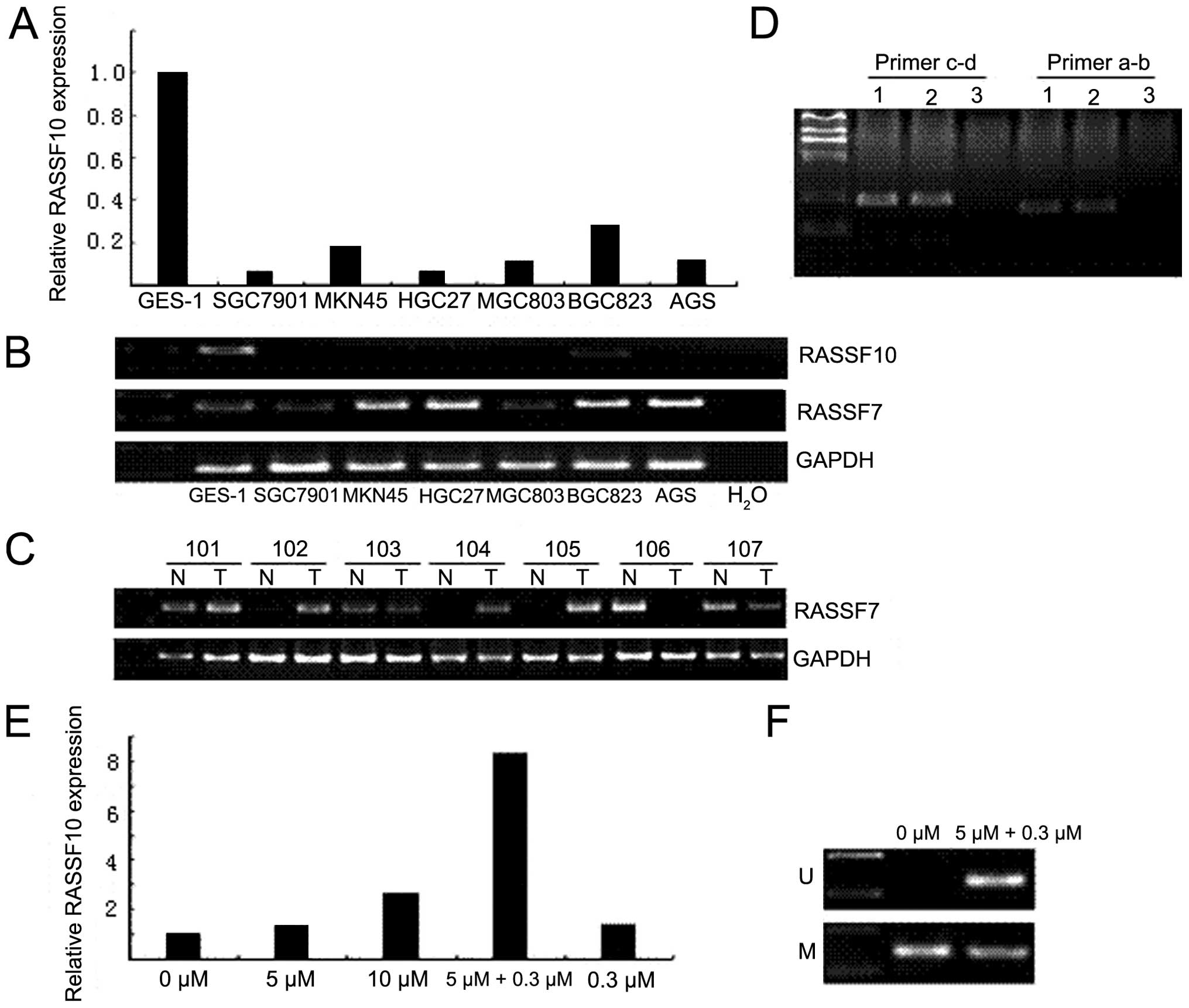
lines was examined by quantitative real-time RT-PCR. The results
showed that RASSF10 was strongly expressed in GES-1 cells, while
its expression was downregulated in 6 gastric cancer cell lines
(Fig. 2A). By contrast, the RASSF7
expression was strong in MKN45, HGC27, BGC823 and AGS, but moderate
in GES-1, SGC7901 and MGC803 (Fig.
2B). In addition, 87% (47/54) of primary gastric cancer tissues
exhibited positive RASSF7 expression, whereas 66.7% (36/54) of
corresponding non-cancerous tissues had positive RASSF7 expression
(P<0.05) (Fig. 2C; Table II). However, the RASSF10 expression
in both carcinoma and non-carcinoma tissues was below detectable
levels (data not shown). Next, to determine the isoform of RASSF10,
we used primer a-b to identify the isoform of the expressed RASSF10
in GES-1 cells. The results indicated that the positive samples
detected using the primer c-d were the same as those detected with
the primer a-b in GES-1 cells (Fig.
2D).
| Table IIStatistical analysis of RASSF7
expression between carcinoma tissues and adjacent non-carcinoma
tissues. |
Table II
Statistical analysis of RASSF7
expression between carcinoma tissues and adjacent non-carcinoma
tissues.
| | RASSF7
expression | |
|---|
| |
| |
|---|
| Patient (n) | Positive (%) | Negative (%) | P-value |
|---|
| Non-carcinoma
tissue | 54 | 36 (66.7) | 18 (33.3) | <0.05 |
| Carcinoma tissue | 54 | 47 (87.0) | 7 (13.0) | |
Upregulation of RASSF10 expression after
5-Aza-dC and TSA treatment
To confirm promoter hypermethylation-mediated
RASSF10 silencing in gastric cancer, MKN45, a cell line negative
for RASSF10 was selected as a model for the re-activation study.
After treatment with DNA demethylating agent and histone
deacetylase inhibitor, re-expression of RASSF10 was analyzed using
quantitative real-time PCR. The result confirmed that the
hypermethylation of RASSF10 promoter occurred in MKN45 cells.
Re-expression of RASSF10 was induced after treatment of 5-Aza-dC
and TSA (Fig. 2E). Furthermore, the
re-expression level of RASSF10 induced by the combination of
5-Aza-dC and TSA was much higher than that induced by 5-Aza-dC
alone or TSA alone, even at the same concentration. Meanwhile, the
MSP results showed that both unmethylated and methylated bands
existed in the combined treated cells, but unmethylated band did
not appear in the control cells (untreated), suggesting that the
methylation status of RASSF10 promoter was partially changed after
the treatment in MKN45 (Fig.
2F).
Promoter hypermethylation of RASSF10 in
gastric cancer
A typical CpG island (CGI) was found in the region
of the RASSF10 gene as shown in Fig.
1B, according to the following criteria: GC content >55%,
ObsCpG/ExpCpG>0.65, and length >500 bp (http://www.urogene.org/methprimer/index1.html). In the
present study, we examined the methylation status of this CGI in
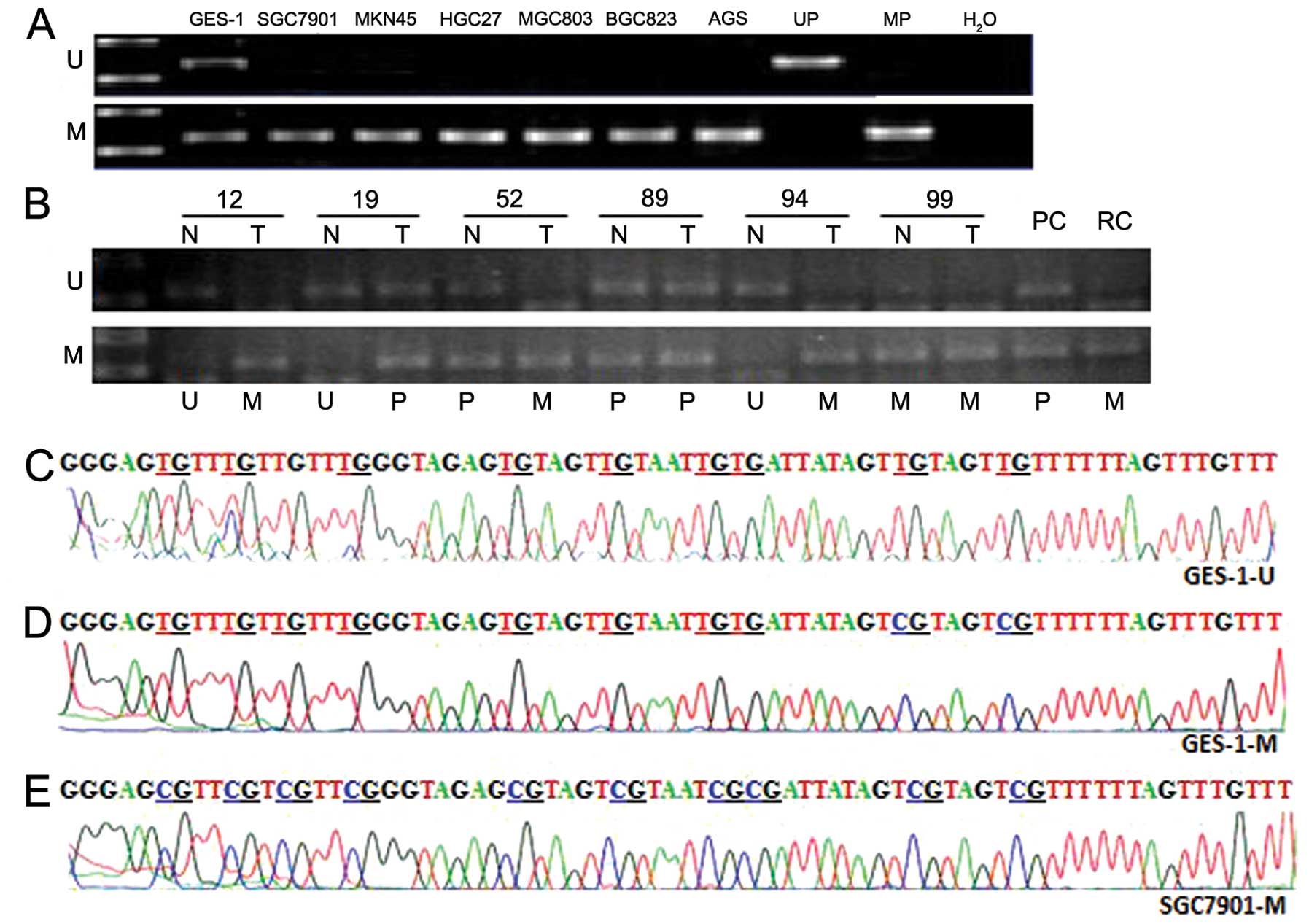
gastric cancer by MSP. As shown in Fig.
3A, RASSF10 hypermethylation was detected in all 6 gastric
cancer cell lines, but in the GES-1 cell line, only partial
methylation was found. To verify our finding from the cell lines,
we analyzed the methylation status of RASSF10 promoter in 86 paired
samples of primary tissues of gastric carcinoma and adjacent
non-carcinoma, 1 sample of pancreatic cancer and 1 sample of rectal
cancer (Fig. 3B). To confirm the
sufficiency of sodium bisulfite modification, both unmethylated and
methylated PCR products of GES-1 (GES-1-U and GES-1-M,
respectively) and methylated product of SGC7901 (SGC7901-M) were
sent for sequencing and the results are shown in Fig. 3C–E. The present study showed that
hypermethylation of RASSF10 promoter was detected in 61.6% (53/86)
of gastric tumor tissues (Table
III), and the rest were partial or unmethylated. By contrast,
the ratio of hypermethylation was only 38.4% (33/86) in the
adjacent non-carcinoma tissues. Statistical analysis revealed a
significant increase of the ratio of RASSF10 hypermethylation in
gastric carcinoma tissues, compared with that in the adjacent
non-carcinoma tissues (P<0.01). The foci with size >10 cm had
a much higher ratio of hypermethylation, compared to those in
smaller size (91.7 vs. 56.8%; P<0.05) (Table IV). Furthermore, compared with
other types (EGC, Borrmann I, Borrmann II and Borrmann III),
Borrmann IV was more frequently methylated (85.7 vs. 56.9%;
P<0.05). Correlation analysis of RASSF10 methylation and TNM
stage (UICC, version 7, 2009) revealed that the tumor with invasion
depth at T3 and T4 had much higher methylation frequency than that
with invasion depth at T1 and T2 (67.1 vs. 37.5%; P<0.05). In
patients with lymph node metastasis (N1, N2 and N3), their tumors
had a higher frequency of RASSF10 hypermethylation than that in the
patients with unaffected lymph nodes (N0), i.e. 68.8% (44/64) of
N1+N2+N3 vs. 40.9% (9/22) of N0; P<0.05. For TNM stage, only 20%
(2/10) of tumors showed hypermethylation of RASSF10 in stage I.
However, the total frequency of RASSF10 hypermethylation in stage
II and stage III was 67.1% (51/76), which was significantly higher
than the frequency in stage I (67.1 vs. 20%; P<0.05). No
significant differences were found between hypermethylation status
and histological type, gender or age.
| Table IIIStatistical analysis of RASSF10
methylation status between adjacent non-carcinoma tissues and
carcinoma tissues. |
Table III
Statistical analysis of RASSF10
methylation status between adjacent non-carcinoma tissues and
carcinoma tissues.
| | RASSF10 methylation
status | |
|---|
| |
| |
|---|
| Patient (n) | U+P (%) | M (%) | P-value |
|---|
| Non-carcinoma
tissue | 86 | 53 (61.6) | 33 (38.4) | <0.01 |
| Carcinoma
tissue | 86 | 33 (38.4) | 53 (61.6) | |
| Table IVCorrelation of clinicopathological
parameters and RASSF10 methylation status in gastric cancer
tissues. |
Table IV
Correlation of clinicopathological
parameters and RASSF10 methylation status in gastric cancer
tissues.
| | RASSF10 methylation
status | |
|---|
| |
| |
|---|
| Variables | Patients
(n=86) | U+P (%) | M (%) | P-value |
|---|
| Gender | | | | >0.05 |
| Male | 60 | 25 (41.7) | 35 (58.3) | |
| Female | 26 | 8 (30.8) | 18 (69.2) | |
| Age (years) | | | | >0.05 |
| <50 | 12 | 7 (58.3) | 5 (41.7) | |
| ≥50 | 74 | 46 (62.2) | 28 (37.8) | |
| Tumor size
(cm) | | | | <0.05 |
| <10 | 74 | 32 (43.2) | 42 (56.8) | |
| ≥10 | 12 | 1 (8.3) | 11 (91.7) | |
| Histological
type | | | | >0.05 |
| Mass+nest | 34 | 15 (44.1) | 19 (55.9) | |
| Diffuse | 52 | 18 (34.6) | 34 (65.4) | |
| Gross type | | | | <0.05a |
| EGC | 4 | 2 (50.0) | 2 (50.0) | |
| Borrmann I | 5 | 3 (60.0) | 2 (40.0) | |
| Borrmann II | 2 | 1 (50.0) | 1 (50.0) | |
| Borrmann III | 61 | 25 (41.0) | 36 (59.0) | |
| Borrmann IV | 14 | 2 (14.3) | 12 (85.7) | |
| Invasion depth | | | | <0.05b |
| T1 | 4 | 2 (50.0) | 2 (50.0) | |
| T2 | 12 | 8 (66.7) | 4 (33.3) | |
| T3 | 15 | 5 (33.3) | 10 (66.7) | |
| T4 | 55 | 18 (32.7) | 37 (67.3) | |
| Lymph node
metastasis | | | | <0.05c |
| N0 | 22 | 13 (59.1) | 9 (40.9) | |
| N1 | 21 | 8 (38.1) | 13 (61.9) | |
| N2 | 14 | 6 (42.9) | 8 (57.1) | |
| N3 | 29 | 6 (20.7) | 23 (79.3) | |
| TNM stage | | | | <0.05d |
| I | 10 | 8 (80.0) | 2 (20.0) | |
| II | 22 | 7 (31.8) | 15 (68.2) | |
| III | 54 | 18 (33.3) | 36 (66.7) | |
| IV | - | - | - | |
Discussion
In recent years, many TSGs have been found to be
epigenetically inactivated in gastric cancer, indicating that
epigenetic silencing of TSGs is one of the major molecular
alterations in the process of gastric carcinogenesis (29). Previously, RASSF10 was identified as
a candidate TSG and its expression is frequently silenced by DNA
methylation in human cancer (24–27).
Our studies also demonstrated that RASSF10 was frequently silenced
in gastric cancer through promoter hypermethylation. However,
RASSF10 expression in both primary carcinoma tissues and adjacent
non-carcinoma tissues was negative, whereas most of the
non-carcinoma tissues were chronic gastritis and intestinal
metaplasia, which are considered as precancerous lesions.
Therefore, our findings indicate that the epigenetic silencing of
RASSF10 is a wide and frequent event, which might be a significant
aberrance related to RASSF10 function during carcinogenesis. In
addition, in the present study, the combination treatment of
5-Aza-dC and TSA in MKN45 cells induced higher RASSF10 expression
compared to single agent treatment with either 5-Aza-dC or TSA,
suggesting that epigenetic alterations including both DNA
hypermethylation and histone deacetylation are involved in the
downregulation of RASSF10 expression in gastric cancer.
Emerging evidence has shown that the
hypermethylation of TSG promoter is one of the major molecular
alterations in cancer development. Methylation of TSGs, such as
COX-2, hMLH1 and p16 are rare in the non-neoplastic gastric mucosa
including chronic gastritis and intestinal metaplasia, but appear
frequently in carcinoma (30).
Therefore, the methylation status of p16 may be a useful biomarker
for the prediction of the malignant potential of dysplasia, in
particular gastric biopsies (31).
However, in the present study, 38.4% (33/86) of adjacent
non-carcinoma tissues were hypermethylated. These results may not
correctly reflect the actual scenario of RASSF10 methylation in
normal gastric mucosa, since most were tissues of chronic gastritis
and intestinal metaplasia, which are considered as precancerous
lesions. Therefore, we propose that the methylation status of
RASSF10 might be used as a biomarker to predict the malignant
potential of the gastric precancerous lesions. In fact, the RASSF10
methylation was rarely found in early tumor cases, but in advanced
tumors, especially in cases which showed extremely malignant
clinicopathological characteristics, such as Borrmann IV, high
degree of invasion, cancerous lymph node and large size. The
increase in RASSF10 hypermethylation suggests that RASSF10 may be
involved in the suppression of tumor invasion and metastasis in
gastric cancer. Overall, epigenetic silencing of RASSF10 is
associated with the pathogenic development of gastric cancer, and
the methylation status of RASSF10 may be used as a biomarker to
evaluate the malignant potential of the precancerous lesions and
malignant degree of tumor in gastric cancer. Thus, we propose that
RASSF10 acts as a novel TSG that is frequently inactivated in
gastric cancer by the epigenetic alterations including DNA
hypermethylation and histone deacetylation.
Previous studies found that RASSF7 expression was
upregulated by hypoxia (17–18).
In particular, RASSF7/N-Ras could promote cell survival through the
inhibition of the mitogen-activated protein kinase kinase 7
(MKK7)/c-Jun N-terminal kinase (JNK) pathway during initial stress.
However, in order to activate cell death signaling pathway with
prolonged stress, the RASSF7 protein was degraded (19). These findings suggest that RASSF7
plays a role in oxidative stress. Several studies have reported
that RASSF7 is upregulated in cancer (14–16).
We also observed a similar phenomenon in gastric cancer. Depletion
of RASSF7 in HeLa cells inhibited cell growth, although knockdown
of RASSF7 did not significantly induce apoptosis (13). Therefore, we speculate that RASSF7
has multiple functions in tumorigenesis, such as promoting mitosis
or survival function to protect cancer cells from damage caused by
stress. Further investigations are required to clarify the
functions of RASSF7 in the development of cancer.
RASSF10 or RASSF10-LF is a 615 amino acid protein
encoded by a two exon gene separated by a short intron of 104 bp. A
new variant of RASSF10 (RASSF10-SF) was reported by Hesson et
al (24), that was found by 5′
rapid amplification of cDNA ends (RACE). RASSF10-SF is a single
exon gene that has a 5′ untranslated region (5′UTR) of 431 bp, a
3′UTR of 478 bp and an open reading frame that encodes a protein of
507 amino acids. To distinguish between the two isoforms by RT-PCR,
we designed a pair of primers where the forward primer binds to the
so-called intron and reverse primer binds to the consensus region
of these two isoform. In the present study, the cDNA of GES-1 was
used as a template for PCR amplification. Results showed that the
primers were able to amplify the cDNA from GES-1 cells, suggesting
that the RASSF gene does not contain the so-called intron and
RASSF10-SF is the true isoform in gastric mucosa.
Our results indicated that RASSF10 is a
characteristically epigenetically silenced tumor suppressor
correlated with invasion and metastasis in gastric cancer and its
epigenetic alteration may contribute to the pathogenic development
of gastric cancer. RASSF-SF may represent the biologically relevant
isoform in human tissues. Exploring the roles of RASSF10 in cancer
development will not only increase our understanding of the biology
of gastric cancer, but may also enable the methylation status of
RASSF10 to be used as a molecular target for diagnosis and therapy.
The expression of RASSF7 was negatively correlated with that of
RASSF10, suggesting a potential oncogenic role of RASSF7 in gastric
cancer. Collectively, the present study revealed a novel tumor
suppressor that is silenced by epigenetic mechanisms in gastric
cancer and provides insight for further studies on the roles of
RASSF7 and RASSF10 in gastric cancer development.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (no. 30572162), the Higher
Specialized Research Fund for Doctoral Program of the Ministry of
Education of China (no. 20102104110001) and the Liaoning Province
Science and Technology Plan Project (no. 2013225021). The authors
are grateful to the Institute of General Surgery of China Medical
University.
Abbreviations:
|
RASSF
|
Ras association domain family
|
|
NT
|
N-terminal
|
|
TSG
|
tumor suppressor gene
|
|
MSP
|
methylation-specific PCR
|
|
5-Aza-dC
|
5-aza-2′-deoxycytidine
|
|
TIS
|
transcription initiation site
|
References
|
1
|
Jemal A, Bray F, Center MM, Farley J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Jones PA and Baylin SB: The fundamental
role of epigenetic events in cancer. Nat Rev Genet. 3:415–428.
2002.PubMed/NCBI
|
|
3
|
Esteller M: Epigenetics in cancer. N Engl
J Med. 358:1148–1159. 2008. View Article : Google Scholar
|
|
4
|
Jones PA and Baylin SB: The epigenomics of
cancer. Cell. 128:683–692. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fleisher AS, Esteller M, Wang S, et al:
Hypermethylation of the hMLH1 gene promoter in human gastric
cancers with microsatellite instability. Cancer Res. 59:1090–1095.
1999.
|
|
6
|
Lee YY, Kang SH, Seo JY, et al:
Alterations of p16INK4A and p15INK4B genes in gastric carcinomas.
Cancer. 80:1889–1896. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Oue N, Shigeishi H, Kuniyasu H, et al:
promoter hypermethylation of MGMT is associated with protein loss
in gastric carcinoma. Int J Cancer. 93:805–809. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tsuchiya T, Tamura G, Sato K, et al:
Distinct methylation patterns of two APC gene promoters in
normal and cancerous gastric epithelia. Oncogene. 19:3642–3646.
2000.PubMed/NCBI
|
|
9
|
Byun DS, Lee MG, Chae KS, Ryu BG and Chi
SG: Frequent epigenetic inactivation of RASSF1A by aberrant
promoter hypermethylation in human gastric adenocarcinoma. Cancer
Res. 61:7034–7038. 2001.PubMed/NCBI
|
|
10
|
Richter AM, Pfeifer GP and Dammann RH: The
RASSF proteins in cancer; from epigenetic silencing to functional
characterization. Biochim Biophys Acta. 1796:114–128.
2009.PubMed/NCBI
|
|
11
|
Sherwood V, Manbodh R, Sheppard C and
Chalmers AD: RASSF7 is a member of a new family of RAS association
domain-containing proteins and is required for completing mitosis.
Mol Biol Cell. 19:1772–1782. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sherwood V, Recino A, Jeffries A, Ward A
and Chalmers AD: The N-terminal RASSF family: a new group of
Ras-association-domain-containing proteins, with emerging links to
cancer formation. Biochem J. 425:303–311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Recino A, Sherwood V, Flaxman A, et al:
Human RASSF7 regulates the microtubule cytoskeleton and is required
for spindle formation, Aurora B activation and chromosomal
congression during mitosis. Biochem J. 430:207–213. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mutter GL, Baak JP, Fitzgerald JT, et al:
Global expression changes of constitutive and hormonally regulated
genes during endometrial neoplastic transformation. Gynecol Oncol.
83:177–185. 2001. View Article : Google Scholar
|
|
15
|
Logsdon CD, Simeone DM, Binkley C, et al:
Molecular profiling of pancreatic adenocancer and chronic
pancreatitis identifies multiple genes differentially regulated in
pancreatic cancer. Cancer Res. 63:2649–2657. 2003.
|
|
16
|
Lowe AW, Olsen M, Hao Y, et al: Gene
expression patterns in pancreatic tumors, cells and tissues. PLoS
One. 2:e3232007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Camps C, Buffa FM, Colella S, et al:
hsa-miR-210 is induced by hypoxia and is an independent prognostic
factor in breast cancer. Clin Cancer Res. 14:1340–1348. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liang GP, Su YY, Chen J, Yang ZC, Liu YS
and Luo XD: Analysis of the early adaptive response of endothelial
cells to hypoxia via a long serial analysis of gene expression.
Biochem Biophys Res Commun. 384:415–419. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Takahashi S, Ebihara A, Kajiho H, Kontani
K, Nishina H and Katada T: RASSF7 negatively regulates
pro-apoptotic JNK signaling by inhibiting the activity of
phosphorylated-MKK7. Cell Death Differ. 18:645–655. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Falvella FS, Manenti G, Spinola M, et al:
Identification of RASSF8 as a candidate lung tumor suppressor gene.
Oncogene. 25:3934–3938. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Korkola JE, Houldsworth J, Chadalavada RS,
et al: Down-regulation of stem cell genes, including those in a
200-kb gene cluster at 12p13.31, is associated with in vivo
differentiation of human male germ cell tumors. Cancer Res.
66:820–827. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lock FE, Underhill-Day N, Dunwell T, et
al: The RASSF8 candidate tumor suppressor inhibits cell growth and
regulates the Wnt and NF-κB signaling pathways. Oncogene.
29:4307–4316. 2010.PubMed/NCBI
|
|
23
|
Hitomi J, Christofferson DE, Ng A, et al:
Identification of a molecular signaling network that regulates a
cellular necrotic cell death pathway. Cell. 135:1311–1323. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hesson LB, Dunwell TL, Cooper WN, et al:
The novel RASSF6 and RASSF10 candidate tumour suppressor genes are
frequently epigenetically inactivated in childhood leukaemias. Mol
Cancer. 8:422009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schagdarsurengin U, Richter AM, Wöhler C
and Dammann RH: Frequent epigenetic inactivation of RASSF10 in
thyroid cancer. Epigenetics. 4:571–576. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hill VK, Underhill-Day N, Krex D, et al:
Epigenetic inactivation of the RASSF10 candidate tumor suppressor
gene is a frequent and an early event in gliomagenesis. Oncogene.
30:978–989. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Helmbold P, Richter AM, Walesch S, et al:
RASSF10 promoter hypermethylation is frequent in malignant melanoma
of the skin but uncommon in nevus cell nevi. J Invest Dermatol.
132:687–694. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
|
|
29
|
Choi IS and Wu TT: Epigenetic alterations
in gastric carcinogenesis. Cell Res. 15:247–254. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kang GH, Lee S, Kim JS and Jung HY:
Profile of aberrant CpG island methylation along multistep gastric
carcinogenesis. Lab Invest. 83:519–526. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun Y, Deng D, You WC, et al: Methylation
of p16 CpG islands associated with malignant transformation of
gastric dysplasia in a population-based study. Clin Cancer Res.
10:5087–5093. 2004. View Article : Google Scholar : PubMed/NCBI
|