Introduction
Langerhans cell (LC) histiocytosis (LCH) is a
disease characterized by the proliferation of CD1a-positive
abnormal LC-like cells (LCH cells). The Writing Group of the
Histiocyte Society has defined LCH as a single system disease or a
multisystem disease (1). Although
it has not yet determined whether LCH is a reactive or neoplastic
disease, recent data suggest a reactive disorder with an underlying
oncogenic potential. In this context, both LCH and pulmonary LCH
harbor the BRAF V600E mutation (2,3) and
appear to be related to stimuli such as viral infection (4–6) and
cigarette smoking (7,8). In addition, it has been reported that
the extinction of stimuli may cause spontaneous healing of the LCH
disease (9–11).
In terms of LCH as a reactive disease, it has been
suggested that viruses might act as causal candidates (6,12–15).
Scappaticci et al (16)
found that peripheral blood lymphocytes from LCH patients contained
chromatid and/or chromosomal breaks, as well as structural
chromosomal rearrangements, and concluded that LCH is
pathogenetically related to an inherent genetic instability or is
caused by environmental viral agents. We recently described the
possibility of a causal relationship between Merkel cell
polyomavirus (MCPyV) and LCH (4).
Establishment of a cell line derived from an LCH
lesion may provide significant information regarding the cell
origin and pathogenesis of the disease. In a previous report, we
described the establishment of a bone LCH lesion-derived cell line
named DOR-1, which showed CD10-positive bone marrow stromal cell
characteristics (17). In the
present study, we report a second cell line named PRU-1,
established also from bone LCH, which shows stromal dermal
dendritic cell (DDC) characteristics.
Materials and methods
Patient
A lytic lesion developed in the skull of a
7-year-old boy who was biopsied at surgery. Histological analysis
confirmed an LCH lesion, which was composed of CD1a+ and
S100+ cells that intermingled with inflammatory cells
and were surrounded by dense mesenchymal tissue.
Cell culture
A written consent to use the biopsy material for
laboratory purposes was obtained from the patient’s parents. The
cells were then allowed to attach to the culture flask and
incubated at 37°C in Dulbecco’s modified Eagle’s medium (DMEM)
supplemented with 10% fetal calf serum, L-glutamine and antibiotics
(Invitrogen-Life Technologies, Cergy-Pontoise, France). After
several days, spindle-shaped cells appeared, which were admixed
with macrophage-like cells and lymphocytes. After 5–6 weeks of
culture, a mixed cell population, including polygonal and
spindle-shaped cells, developed. The culture, which was then named
PRU-1, continued growth and proliferation and was passaged at a
density of 2×105 cells/ml.
Flow cytometry
For two-color flow cytometry, PRU-1 cells
(2×105) were detached from the culture flask and washed
with PBS containing 0.5% bovine serum albumin, followed by
incubation with specific antibodies. Non-specific immunolabeling
was blocked by treating the cells with heat-inactivated rabbit
serum (Sigma-Aldrich). Most of the monoclonal antibodies (mAbs)
used were directly conjugated with phycoerythrin or fluorescein
isothiocyanate. For the control, isotype-matched irrelevant mAbs at
the same dilution as the specific antibodies were used. The stained
cells were analyzed on a fluorescence activated cell sorter Calibur
flow cytometer (BD Biosciences, San Jose, CA, USA), and data
evaluation was performed using the CellQuest software (BD
Biosciences).
Immunocytochemistry and special staining
of PRU-1 cells
PRU-1 cells were analyzed by immunocytochemistry
using the mAbs and polyclonal antibodies listed in Table I. Cytocentrifuge smears, which were
prepared after the detachment of adherent PRU-1 cells growing on a
culture chamber slide (Falcon; BD Labware, Franklin Lakes, NJ,
USA), were fixed in cold acetone for 10 min, rinsed in PBS and
incubated with a primary antibody. The specific antibodies were
revealed using a polymer-based immunoperoxidase technique (EnVision
Plus; DakoCytomation, Glostrup, Denmark). PAS reaction and diastase
PAS reaction were conducted.
 | Table IImmunohistochemical analyses of
PRU-1, a new cell line derived from a bone LCH lesion. |
Table I
Immunohistochemical analyses of
PRU-1, a new cell line derived from a bone LCH lesion.
| Antibodies
against | Clones | Sources | Dilution |
Immunoreactivity |
|---|
| CD1a | SK9 | BD | 1:20 | − |
| CD10 | 56C6 | Novocastra | 1:50 | − |
| S100 | - | Dako Japan | 1:1,000 | − |
| CD11c | 3.9 | YLEM | 1:20 | + |
| CD14 | MφP9 | BD Biosciences | 1:20 | − |
| CD34 | QBEND-10 | Dako Japan | Diluted | − |
| CD43 | MT1 | Euro-Diagnostica
AB | 1:20 | + |
| CD54 (ICAM-1) | BBIG-I1 | Seikagakukogyo | 1:1,000 | + |
| CD56 (NCAM) | 123C3 | Monosan | 1:20 | + |
| CD68 | KP-1 | Dako Japan | 1:50 | + |
| CD99 (MIC2) | 19 | Sigma | 1:40 | + |
| CD106
(αVCAM-1) | BBIG-V1 | British Bio
Technology | 1:1,000 | + |
| CD141
(thrombomodulin) | 1009 | Dako Japan | 1:25 | + |
| αSMA | 1A4 | Dako Japan | 1:50 | + |
| Muscle actin | HHF35 | Enzo
Diagnostics | 1:50 | + |
| FXIIIa | - | Lab Vision
Corporation | 10 μg/ml | + |
| HLA-DR | DK22 | Dako Japan | 1:20 | − |
| Keratin
(AE1/3) | AE1/3 | Chemicon | 1:500 | + |
| Vimentin | Vim3B4 | Dako Japan | 1:50 | + |
DNA extraction and analysis of TCRγ
rearrangements in the primary LCH lesion and PRU-1 cells
Extraction of DNA from PRU-1 was performed using the
proteinase K and phenol/chloroform/isoamyl alcohol (25:24:1 v/v/v)
standard procedure. The presence of TCRγ rearrangement in the cells
was investigated using PCR-amplified DNA from a paraffin-embedded,
formalin-fixed original LCH lesion and the proliferating PRU-1
cells following the BIOMED-2 collaboration study protocol (18). PCR products were analyzed using
GeneMapper™ software v.3.5 (Applied Biosystems, Foster City, CA,
USA).
Electron microscopy (EM)
Ultrastructural examination of PRU-1 was performed
at different passages after fixation in glutaraldehyde following
classical EM protocols of the Okayama University Central Laboratory
(Okayama, Japan).
Cytogenetics
PRU-1 metaphases were analyzed according to standard
methods of SRL, Inc. (Tachikawa, Tokyo, Japan).
Effects of a conditioned medium from
PRU-1 cell culture supernatant on the biology and phenotype of
lymphocytes and monocytes from a healthy donor
A conditioned medium was prepared using the
supernatant of 1-week cultured PRU-1 cells and diluted with DMEM at
a volume ratio of 1:1; this was then used to treat lymphocytes
and/or monocytes isolated using a pore filter [0.4-μm pore filter
(30-mm Millicell; Nihon Millipore, Tokyo, Japan)]. The conditioned
medium from HeLa cell culture was used as the control.
Xenografting into SCID mice
Animal studies were approved by Okayama University
Animal Research Laboratory. PRU-1 cells [5×106 cells in
0.2 ml PBS (−)] were injected subcutaneously into five female SCID
mice (CB-17 SCID; Okayama University Animal Research Laboratory,
Okayama, Japan). The progress of the xenografts was monitored two
times a week for 3 months.
Multiplex quantitative real-time PCR
(Q-PCR) for MCPyV detection
Multiplex Q-PCR was performed as previously
described (4,19).
Results
Histology and immunohistochemistry of the
original LCH lesion
Histological examination of hematoxylin and eosin
(H&E)-stained sections (Fig.
1A–D) and immunohistochemistry of the initial LCH bone lesion
showed the presence of CD1a-positive (Fig. 1E and F) and S100-positive cells
(Fig. 1J and L). Cytokeratin
(AE1/3)-positive cells with kidney-like nuclei were distributed
across the LCH lesion (Fig. 1H, I and
K). Double staining for S100 and AE1/3 showed the presence of
four types of immunoreactive cells within the lesion:
S100−/AE1/3−;
S100+/AE1/3− (green in Fig. 1J and L);
S100−/AE1/3+ (red in Fig. 1K and L); and
S100+/AE1/3+ (orange in Fig. 1L). Within the peripheral stromal
component of the lesion, scattered spindle- or stellate-shaped
FXIIIa+ cells (Fig. 1G)
and a low number of CD1a+ cells harboring foci
(arrowhead) were observed (Fig.
1E).
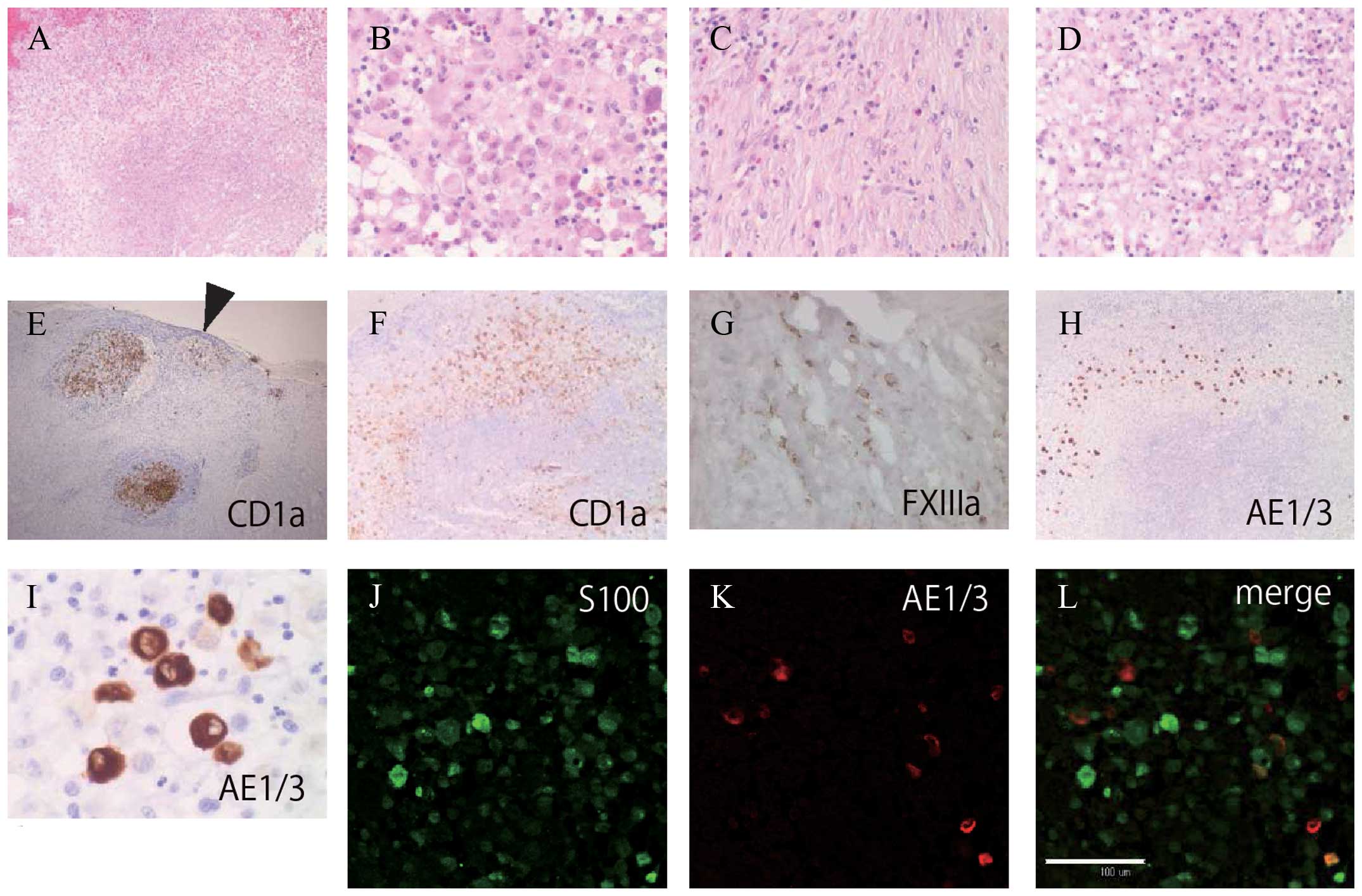 | Figure 1Histological and immunohistochemical
analysis of the original LCH lesion including CD1a-positive
abnormal Langerhans cell-like cells (LCH cells) with a zoning
phenomenon. (A) Granulomatous lesion with central necrosis,
hematoxylin and eosin (H&E) stain (magnification, ×4). (B) LCH
cells with kidney-like nuclei and pale cytoplasm admixed with
lymphocytes and eosinophils (H&E, magnification, ×20). (C)
Peripheral part of the lesion showing a fibrosis-like pattern
(H&E, magnification, ×20). (D) The central necrotic region of
the nodular lesion (H&E, magnification, ×20). (E)
Immunohistochemical analysis showed two large nodular clusters and
a peripheral small cluster (arrowhead) of CD1a-positive LCH cells
(magnification, ×4). (F) CD1a-positive LCH cell-proliferated lesion
contained a CD1a-negative central necrotic area (magnification,
×4). (G) Factor XIIIa (FXIIIa)-positive cells scattered in the
peripheral fibrous area (magnification, ×40). (H and I)
AE1/3-positive cells scattered in the middle area. H: same field of
(A) (H, magnification, ×4; I, magnification, ×40). (J–L) Double
staining for S100 protein and AE1/3 in the same lesion. (J) S100
(magnification, ×20). (K) AE1/3 (magnification, ×20). (L) Merge
(magnification, ×20). S100+/AE1/3+ cells
(orange), S100+/AE1/3− cells (green), and
S100−/AE1/3+ cells (red) intermingled. White
scale bar, 100 μm. |
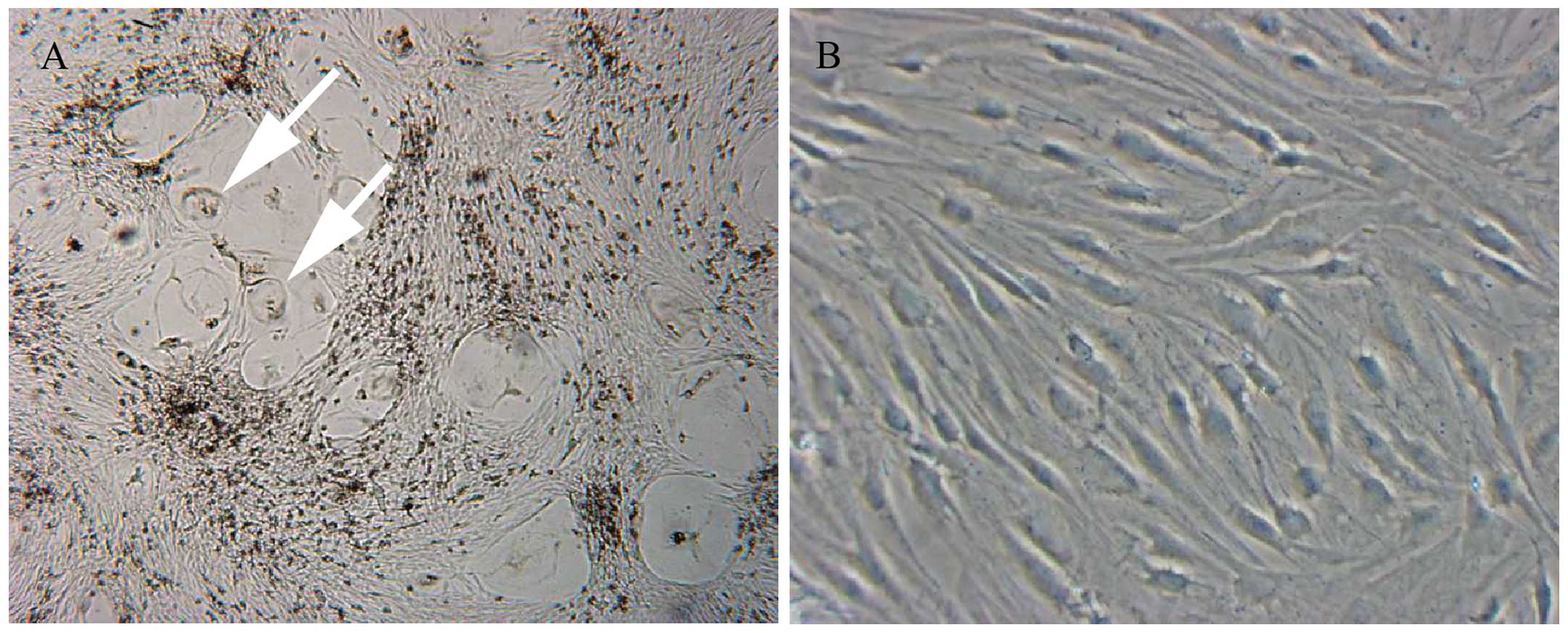
Establishment of the PRU-1 cell line
Primary cultures obtained after seeding the isolated
cells from the LCH infiltrate contained mixed adherent cell
populations of variable size and morphology (Fig. 2A). During the second in vitro
passage, adherent spindle-shaped cells grew predominantly. One
month after initial plating, the proliferating cells could be
maintained in DMEM supplemented with 10% fetal calf serum,
L-glutamine and antibiotics. After 6 weeks of continuous culture,
densely packed adherent cells were further passaged at a density of
2×105 cells/ml. The cells continued to grow stably for
at least 50 passages. This new cell line, was named PRU-1 (Fig. 2B) and was extensively
characterized.
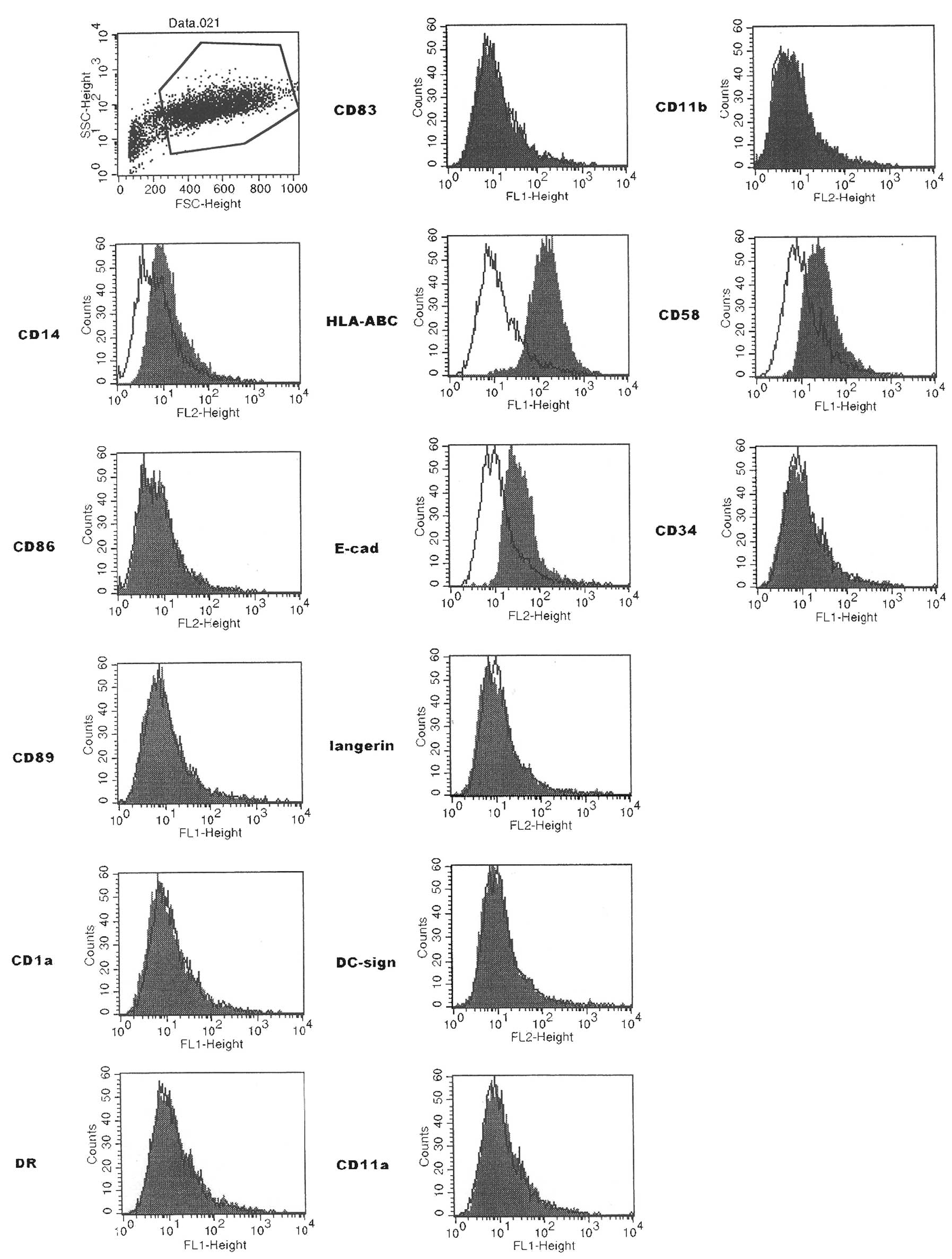
Flow cytometry
The analysis of PRU-1 from the early seventh passage
showed that the predominantly expressed molecules were CD14,
HLA-ABC, CD58 (leukocyte-function associated molecule-3) and
E-cadherin (Fig. 3). LC markers
such as CD1a, CD207 (langerin) and CD209 (DC sign; immature DC
marker) were negative. CD83 (B cell activation protein; DC marker),
CD11b, CD86 [CD152 (CTLA4) ligand; interdigitating cell marker] and
CD34 were negative.
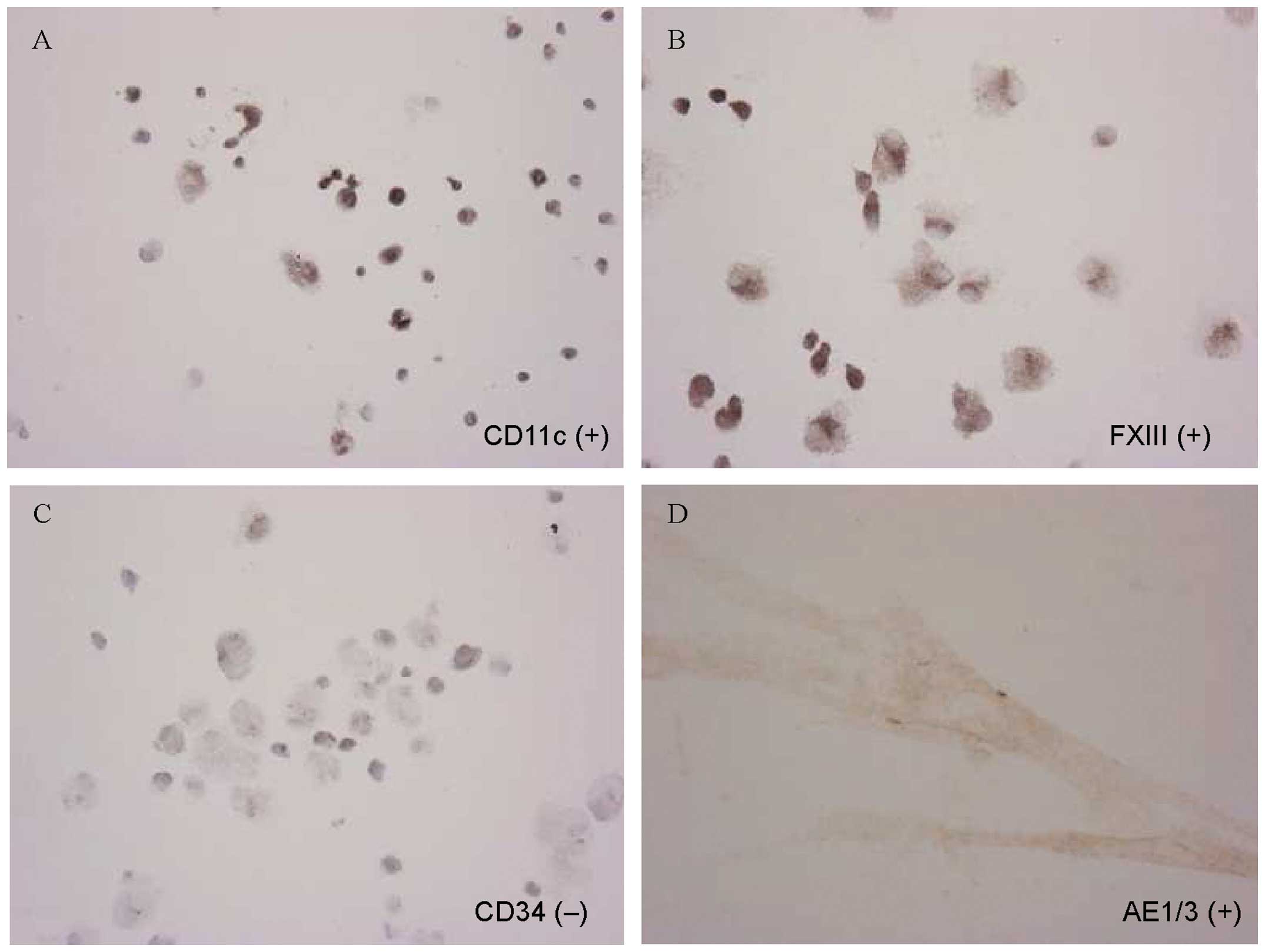
Immunophenotype of PRU-1 and a special
reaction for glycogen
The immunocytochemical profiles of PRU-1 cells are
summarized in Table I. Application
of a large panel of antibodies demonstrated that the PRU-1 cells
were immunoreactive to CD11c (Fig.
4A), FXIIIa (Fig. 4B), CD43,
CD54 (ICAM-1), CD56 (NCAM), CD99 (MIC2), CD106 (αVCAM-1), and CD141
(thrombomodulin), but not to CD1a, CD10, S100, CD14 and CD34
(Fig. 4C). In addition, AE1/3
keratin (Fig. 4D), vimentin,
α-smooth muscle actin and muscle actin were expressed.
Cytocentrifuge smears of the PRU-1 cells showed the presence of
glycogen, as indicated by the positive PAS reaction and confirmed
by the diastase PAS reaction.
Analysis of TCRγ in the LCH lesion and
PRU-1 cells by PCR
Since TCRγ expansion has been reported in LCH
lesions occurring in patients treated for T-lymphoblastic
leukemia/lymphoma (20), we
determined the TCRγ gene status in both the LCH original tissue and
the PRU-1 cells by PCR. GeneMapper™ retrieved only polyclonal bands
using the amplified DNA. However, histological analysis of the
lesion showed that numerous T cells intermingled with LC cells. No
rearrangement of the TCRγ band was detected in the PRU-1 cells.
EM
Various organelles such as the mitochondria, rough
ER, Golgi apparatus, and lysosomes were observed in the PRU-1 cells
by EM. No Birbeck granules were detected.
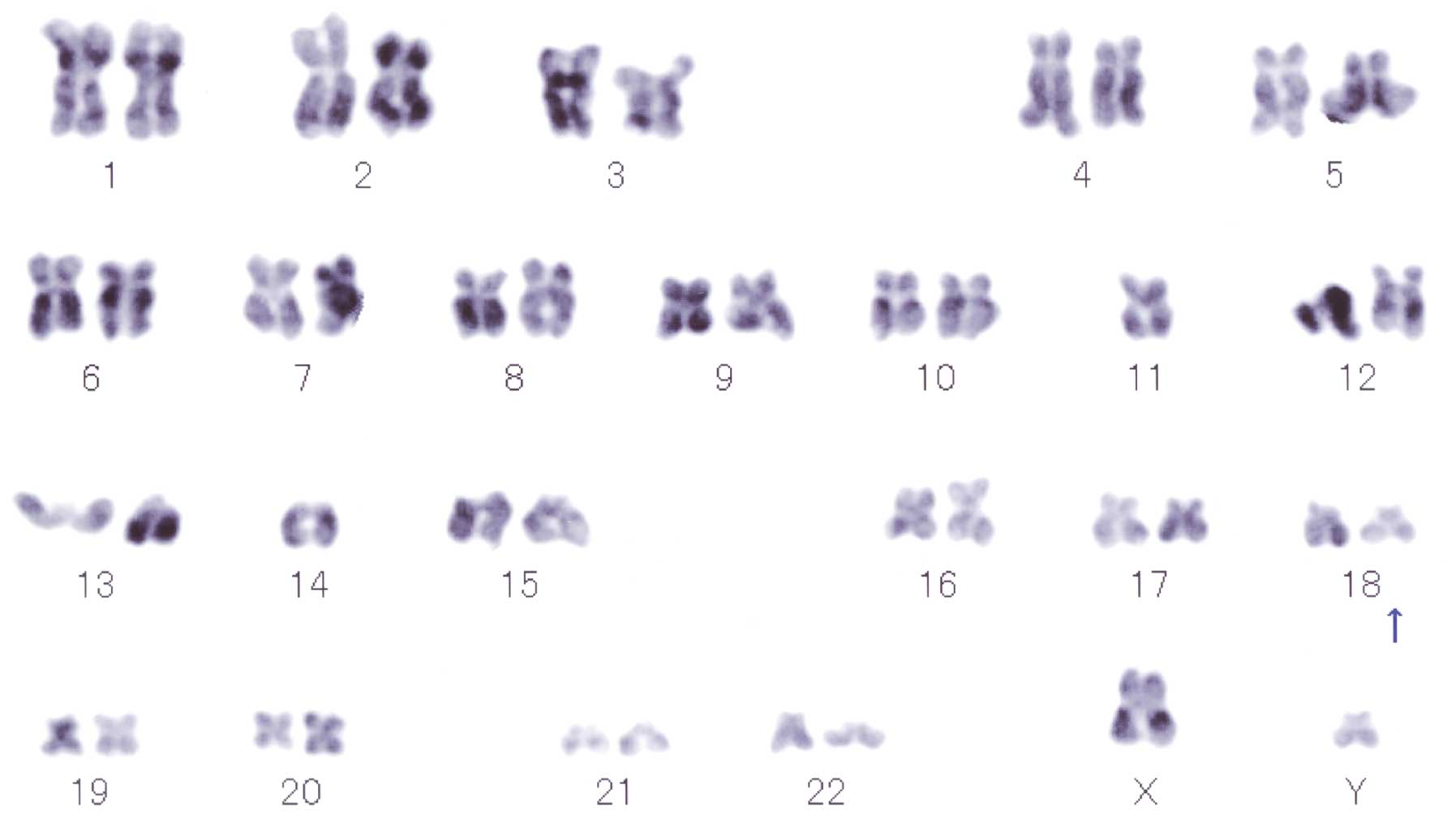
Cytogenetic analysis
Repeated karyotype analysis of seven metaphases was
performed, which showed male karyotypes consisting of the following
constitutions: one metaphase spread showing 39, XY, −2, −4, −8,
−12, −12, −14, add(18)(q21), 20,
+mar, one metaphase spread showing 44, XY, −11, −14, add(18)(q21) (Fig.
5), one metaphase spread showing 44, XY, −10, −14, and four
metaphase spreads showing 46, XY. The 18q21 abnormality was
detected two times.
Effects of the PRU-1 conditioned medium
on the biology and phenotype of lymphocytes and monocytes from a
healthy donor
Lymphocytes and monocytes were co-cultured in the
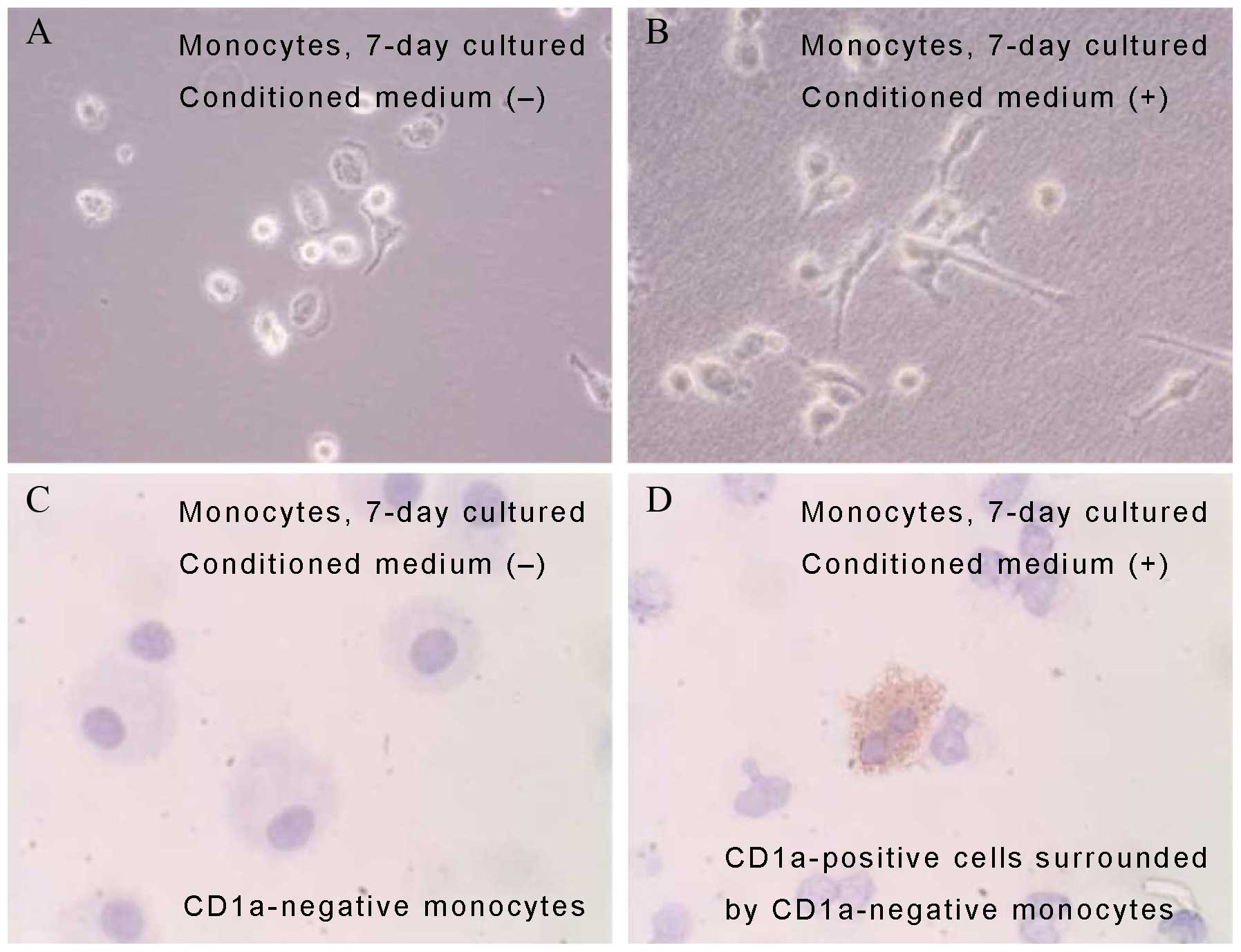
PRU-1 conditioned medium. After 7 days of culture in the
conditioned medium, the monocytes underwent a change in morphology,
resulting in spindle-shaped cells (Fig.
6A and B), whereas some monocytes acquired CD1a positivity
(Fig. 6C and D). Lymphocytes
cultured in the PRU-1 supernatant showed a higher proliferation
rate than the control lymphocytes cultured in a conditioned medium
from HeLa cell culture.
Growth in SCID mice
To evaluate whether PRU-1 possessed normal or
neoplastic characteristics, we examined the tumorigenicity of the
PRU-1 cell line. No tumor growth was detected in the five female
SCID mice that were monitored for 3 months.
Q-PCR for MCPyV
No MCPyV DNA sequences were detected in the PRU-1
cell line.
Discussion
Although several studies have established cell lines
from LCH lesions, it is generally difficult to retain its full LC
characteristics (17,21–23).
Previous studies have shown success in short-term culture of LCH
tissues; however, these only allowed analyses of cell morphology
(24), cytokine production (IL-1
and PGE2) (22), and cell
differentiation capacities (23). A
few subsequent passages have been achieved, whereas most studies
have suggested that LCH cells might have a limited life span in
vitro. In the present study, a long-term growing cell line was
obtained, which in turn gave rise to the question of whether the
PRU-1 cell line was related to the LC lineage or represented an LC
progenitor. LC progenitors are components of multipotent bone
marrow-derived stromal cellular compartments that have been
implicated in self-perpetuating granulomatous lesions. Thus, the
immunocytochemical profile, namely the FXIIIa positivity and the
absence of CD34 expression and distinct cytoplasmic Birbeck
granules, were a priori consistent with a DDC origin for the
PRU-1 cells. It was previously reported that CD1a-positive DCs from
peripheral blood mononuclear cells could differentiate into any of
the three cell types: histiocyte DC-like cells
(FXIIIa+++), fibroblast-like cells
(FXIIIa++), and giant cell-like cells
(FXIIIa+) (25). In
fact, fibroblast-like FXIIIa+ DDC is the cell type that
typically proliferates in dermatofibromas. In this regard, PRU-1
exhibited phenotypic characteristics of fibroblast-like DDCs in
terms of FXIIIa positivity and its spindle-shaped cellular
morphology. In addition, EM analysis showed that PRU-1 harbored
numerous organelles. Similarly, dermatofibroma tumoral cells show
well-developed rough ER, conspicuous Golgi apparatus, and a
variable number of mitochondria (26). These features thus suggested that
PRU-1 might be a clonal expansion derived from FXIIIa+
stellate cells present in the fibrous area of the original
lesion.
The PRU-1 cells also expressed cytokeratin (AE1/3=
cytokeratin 1–8/10/14/15/16/19) (Table
I). The original LCH lesions systematically showed scattered
keratin-positive cells (Fig. 1H–L).
Double staining indicated that the LCH lesion contained a mixture
of S100+/AE1/3+,
S100+/AE1/3−,
S100−/AE1/3+ and
S100−/AE1/3− cells. Bone LCH has a
self-limiting tendency and develops into scar-like granuloma with
the accumulation of a mixture of foamy cells and macrophages
(27). One of the important
questions that needs to be answered is the origin of LCH cells. The
presence of S100+/AE1/3+ cells raises the
question of whether the AE1/3+ cells originate from
typical LCH cells or from other cell types that comprise the LCH
lesion. Keratin is present in the cellular cytoskeleton, and
cytokeratin+ cells could produce cytokine (28) with admixed cells in the LCH lesion
(29). PRU-1 phenotypically
consisted of S100−, AE1/3+ and
FXIIIa+ cells. Moreover, scattered FXIIIa+
cells were detected in the peripheral fibrous area of the original
LCH lesion (Fig. 1G). In general,
it is accepted that LC is derived from an LC precursor through an
immature cell type and that LC can differentiate into
interdigitating cells through veiled cells (30). Similarly, DDCs seem to have been
derived from a cellular precursor that has differentiated into DCs
or LCs. There are data indicating that LCs are incapable of
differentiating into the DDC type (31). We think PRU-1 was derived from
stromal cell types that comprised the LCH lesion.
It has been suggested that some LCH cells are
derived from the T cell lineage with TCRγ monoclonality (20), although a diverse collection of data
exists. Our PCR results allowed the characterization of the
polyclonal TCRγ genomic structure of the PRU-1 original tissue, as
well as indicated the absence of TCRγ monoclonality in the PRU-1
cells. These data showed the presence of polyclonal T cells in the
LCH original tissue and that PRU-1 elements did not originate from
the T cell lineage.
Cytogenetic analyses of PRU-1 showed the presence of
an 18q21 chromosomal abnormality. We recently described the
possibility of a causal relationship between MCPyV and LCH
(4) and Scappaticci et al
(16) found that peripheral blood
lymphocytes from LCH patients contained chromosomal abnormality and
pointed out that LCH was caused by environmental viral agents. The
18q21 abnormality which was detected two times may also indicate a
clonal proliferation of PRU-1 cells and the effect of viral
infection as shown in blood lymphocytes by Scappaticci et al
(16). In this context, chromosomal
rearrangements have been detected in a few LCH cases (32,33),
as well as in pulmonary LCH which is thought to be a reactive and
neoplastic disorder caused by cigarette smoking (2,7).
Familial clustering suggesting a genetic event in LCH has also been
reported (6,34–36).
To assess cytokine production, the conditioned
medium was added to the monocyte and lymphocyte cultures (37) as well as to lymphocytes or monocytes
from a healthy donor. Observed phenotypic changes included some
CD1a− monocytes transforming into CD1a+ cells
and a high proliferation rate compared with that of the control.
These findings suggest that the PRU-1 cultured cells produced
stimulatory growth or differentiation factors that have yet to be
fully characterized.
Analysis of specific chromosomal aberrations using
the PRU-1 in vitro model and conditioned medium might also
provide new insights on LCH pathogenesis, i.e. LCH microenvironment
contributed by stromal cells.
We recently described the relationship between MCPyV
and LCH (4) and hypothesized that
LCH was a reactive disorder with an underlying oncogenic potential.
Extinction of stimuli such as MCPyV infection could cause
spontaneous healing of LCH, although MCPyV has been considered as a
potential neoplastic agent for Merkel cell carcinoma (38). However, in the PRU-1 cells, MCPyV
DNA was not detected and was not likely to contribute to the cell
line establishment, although the genomic instability of PRU-1 cells
might be induced by MCPyV infection (16).
Finally, the LCH lesion is a mixture of various
cells, including LC-like cells, DDC-like cells, macrophages,
lymphocytes, neutrophils, lymphocytes and eosinophils. We were
successful in establishing a CK+/S100− cell
line, PRU-1, from a bone LCH lesion that retains DDC-like
characteristics such as CD11c, CD54 and CD141 immunoreactivity, in
addition to FXIIIa expression. The PRU-1 conditioned medium may
contain yet unknown specific molecules that may contribute to the
formation of LCH lesions, as well as facilitate in induction
studies using CD1a+ or LCH cells.
Acknowledgements
The authors are indebted to Ms. Masumi Furutani
(Central Laboratory, Okayama University) for her help in performing
EM analysis.
References
|
1
|
Writing Group of the Histiocyte Society.
Histiocytosis syndromes in children. Writing Group of the
Histiocyte Society. Lancet. 1:208–209. 1987.PubMed/NCBI
|
|
2
|
Yousem SA, Dacic S, Nikiforov YE and
Nikiforova M: Pulmonary Langerhans cell histiocytosis: profiling of
multifocal tumors using next-generation sequencing identifies
concordant occurrence of BRAF V600E mutations. Chest.
143:1679–1684. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Badalian-Very G, Vergilio JA, Degar BA, et
al: Recurrent BRAF mutations in Langerhans cell histiocytosis.
Blood. 116:1919–1923. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Murakami I, Matsushita M, Iwasaki T, et
al: Merkel cell polyomavirus DNA sequences in peripheral blood and
tissues from patients with Langerhans cell histiocytosis. Hum
Pathol. 45:119–126. 2014. View Article : Google Scholar
|
|
5
|
Sakata N, Toguchi N, Kimura M, Nakayama M,
Kawa K and Takemura T: Development of Langerhans cell histiocytosis
associated with chronic active Epstein-Barr virus infection.
Pediatr Blood Cancer. 50:924–927. 2008. View Article : Google Scholar
|
|
6
|
Chen CJ, Ho TY, Lu JJ, et al: Identical
twin brothers concordant for Langerhans’ cell histiocytosis and
discordant for Epstein-Barr virus-associated haemophagocytic
syndrome. Eur J Pediatr. 163:536–539. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yousem SA, Colby TV, Chen YY, Chen WG and
Weiss LM: Pulmonary Langerhans’ cell histiocytosis: molecular
analysis of clonality. Am J Surg Pathol. 25:630–636. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tazi A, Hiltermann JN and Vassallo R:
Adult lung histiocytosis. Histiocytic Disorders of Children and
Adults. Weitzman S and Egeler RM: Cambridge University Press;
Cambridge: pp. 187–207. 2005
|
|
9
|
Corbeel L, Eggermont E, Desmyter J, et al:
Spontaneous healing of Langerhans cell histiocytosis (histiocytosis
X). Eur J Pediatr. 148:32–33. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mogulkoc N, Veral A, Bishop PW, Bayindir
U, Pickering CA and Egan JJ: Pulmonary Langerhans’ cell
histiocytosis: radiologic resolution following smoking cessation.
Chest. 115:1452–1455. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Von Essen S, West W, Sitorius M and
Rennard SI: Complete resolution of roentgenographic changes in a
patient with pulmonary histiocytosis X. Chest. 98:765–767. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kawakubo Y, Kishimoto H, Sato Y, et al:
Human cytomegalo-virus infection in foci of Langerhans cell
histiocytosis. Virchows Arch. 434:109–115. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Leahy MA, Krejci SM, Friednash M, et al:
Human herpesvirus 6 is present in lesions of Langerhans cell
histiocytosis. J Invest Dermatol. 101:642–645. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Glotzbecker MP, Carpentieri DF and Dormans
JP: Langerhans cell histiocytosis: a primary viral infection of
bone? Human herpes virus 6 latent protein detected in lymphocytes
from tissue of children. J Pediatr Orthop. 24:123–129. 2004.
View Article : Google Scholar
|
|
15
|
Jenson HB, McClain KL, Leach CT, Deng JH
and Gao SJ: Evaluation of human herpesvirus type 8 infection in
childhood langerhans cell histiocytosis. Am J Hematol. 64:237–241.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Scappaticci S, Danesino C, Rossi E, et al:
Cytogenetic abnormalities in PHA-stimulated lymphocytes from
patients with Langerhans cell histocytosis. AIEOP-Istiocitosi
Group. Br J Haematol. 111:258–262. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gogusev J, Telvi L, Murakami I, et al:
DOR-1, A novel CD10+ stromal cell line derived from
progressive Langerhans cell histiocytosis of bone. Pediatr Blood
Cancer. 44:128–137. 2005. View Article : Google Scholar
|
|
18
|
Van Dongen JJ, Langerak AW, Bruggemann M,
et al: Design and standardization of PCR primers and protocols for
detection of clonal immunoglobulin and T-cell receptor gene
recombinations in suspect lymphoproliferations: report of the
BIOMED-2 Concerted Action BMH4-CT98-3936. Leukemia. 17:2257–2317.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kuwamoto S, Higaki H, Kanai K, et al:
Association of Merkel cell polyomavirus infection with morphologic
differences in Merkel cell carcinoma. Hum Pathol. 42:632–640. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Feldman AL, Berthold F, Arceci RJ, et al:
Clonal relationship between precursor T-lymphoblastic
leukaemia/lymphoma and Langerhans-cell histiocytosis. Lancet Oncol.
6:435–437. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nezelof C and Basset F: An hypothesis
Langerhans cell histiocytosis: the failure of the immune system to
switch from an innate to an adaptive mode. Pediatr Blood Cancer.
42:398–400. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Arenzana-Seisdedos F, Barbey S, Virelizier
JL, Kornprobst M and Nezelof C: Histiocytosis X. Purified
(T6+) cells from bone granuloma produce interleukin 1
and prostaglandin E2 in culture. J Clin Invest. 77:326–329. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Geissmann F, Lepelletier Y, Fraitag S, et
al: Differentiation of Langerhans cells in Langerhans cell
histiocytosis. Blood. 97:1241–1248. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nezelof C and Basset F: Langerhans cell
histiocytosis research. Past, present, and future. Hematol Oncol
Clin North Am. 12:385–406. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aiba S and Tagami H: Phorbol 12-myristate
13-acetate can transform monocyte-derived dendritic cells to
different cell types similar to those found in dermatofibroma. A
possible in vitro model of the histogenesis of dermatofibroma. J
Cutan Pathol. 25:65–71. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Erlandson RA: Ultrastructural features of
specific human neoplasms with clinicopathologic,
immunohistochemical, and cytogenetic correlations. Diagnostic
Transmission Electron Microscopy of Tumors. Raven Press; New York,
NY: pp. 243–832. 1994
|
|
27
|
Weitzman S and Egeler RM: Langerhans cell
histiocytosis of bone. Histiocytic Disorders of Children and
Adults. Cambridge University Press; Cambridge: pp. 154–173. 2005,
View Article : Google Scholar
|
|
28
|
Lu H, Chen J, Planko L, Zigrino P,
Klein-Hitpass L and Magin TM: Induction of inflammatory cytokines
by a keratin mutation and their repression by a small molecule in a
mouse model for EBS. J Invest Dermatol. 127:2781–2789.
2007.PubMed/NCBI
|
|
29
|
Egeler RM, Favara BE, van Meurs M, Laman
JD and Claassen E: Differential in situ cytokine profiles of
Langerhans-like cells and T cells in Langerhans cell histiocytosis:
abundant expression of cytokines relevant to disease and treatment.
Blood. 94:4195–4201. 1999.PubMed/NCBI
|
|
30
|
Raushenbakh MO, Ivanova VD, Shevchenko VE,
Makhonova LA and Sergeev AV: Congenital tyrosine metabolism
disorders in children with hemoblastoses. Vestn Akad Med Nauk SSSR.
19–24. 1981.(In Russian).
|
|
31
|
Larrengina AT and Falo LD Jr: Dendritic
cells in the context of skin immunity. Dendritic Cells. Lotze MT
and Thomson AW: Academic Press; San Diego, CA: pp. 301–314. 2001,
View Article : Google Scholar
|
|
32
|
Betts DR, Leibundgut KE, Feldges A, Pluss
HJ and Niggli FK: Cytogenetic abnormalities in Langerhans cell
histiocytosis. Br J Cancer. 77:552–555. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Murakami I, Gogusev J, Fournet JC, Glorion
C and Jaubert F: Detection of molecular cytogenetic aberrations in
langerhans cell histiocytosis of bone. Hum Pathol. 33:555–560.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Arico M, Nichols K, Whitlock JA, et al:
Familial clustering of Langerhans cell histiocytosis. Br J
Haematol. 107:883–888. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dufour C, Lanciotti M, Micalizzi C,
Valetto A and Haupt R: Non-identical twin sisters concordant for
Langerhans cell histiocytosis and discordant for secondary acute
promyelocytic leukemia. Med Pediatr Oncol. 37:70–72. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Enjolras O, Leibowitch M, Bonacini F,
Vacher-Lavenu MC and Escande JP: Congenital cutaneous Langerhans
histiocytosis. Apropos of 7 cases. Ann Dermatol Venereol.
119:111–117. 1992.(In French).
|
|
37
|
Miyatani K, Takahashi K, Yanai H, Yoshino
T and Akagi T: Partial purification and characterization of
dendritic cell differentiation factor. Acta Med Okayama. 48:67–72.
1994.PubMed/NCBI
|
|
38
|
Feng H, Shuda M, Chang Y and Moore PS:
Clonal integration of a polyomavirus in human Merkel cell
carcinoma. Science. 319:1096–1100. 2008. View Article : Google Scholar : PubMed/NCBI
|