Liver cancer is the sixth most common cancer and the
second most common cause of cancer-associated mortality worldwide
(1). Approximately 75% of all
primary liver cancer types are hepatocellular carcinoma (HCC) that
formed from liver cells. Liver cancer can be formed from other
structures in the liver such as bile duct, blood vessels and immune
cells. Secondary liver cancer is a result of metastasis of cancer
from other body sites into the liver. The major cause of primary
liver cancer is viral infection with either hepatitis C virus (HCV)
or hepatitis B virus (HBV), which leads to massive inflammation,
fibrosis and eventual cirrhosis in the liver. Many genetic and
epigenetic alterations have been identified in hepatocytes during
HCV and HBV infection (2). Other
causes, such as alcohol, aflatoxin, high-grade dysplastic nodules,
obesity, diabetes and smoking may increase the risk of liver
cancer. Surgical resection is an option for liver cancer treatment
(3), whereas liver transplantation
can be used in cases of liver cancer where surgical resection can
be tolerated and the tumor fits specific criteria, i.e., Milan
criteria (4).
Signaling pathways are complex processes of signal
transduction involving the mutual activation of a protein cascade
transmitting signals from the cell surface to the cytoplasm and the
nucleus (5). In recent decades,
emerging studies have greatly improved our understanding of liver
tumorigenesis through investigation of a series of signaling
pathways including PI3K/AKT. Cell signaling receptors,
intracellular secondary messengers/molecules and transcription
factors are essential components for signaling pathways, and the
protein-protein interactions (PPIs) among these components act as
connectors that mediate signal transduction from one step to the
following within a single signaling pathway, and act as
transmitters that play an important role in the crosstalk of
several signaling pathways (6).
PPIs refer to the intentional physical contacts established between
two or more proteins as a result of biochemical events and/or
electrostatic forces (7). Proteins
rarely act alone at both cellular and systemic levels. A number of
essential molecular processes are performed by molecular machines
that are constructed from a large number of protein components
organized by their PPIs. PPIs have been largely investigated in
signal transduction and aberrant PPIs in these signaling pathways
are considered the basic events of liver cancer. Targeting these
important PPIs may be useful for the treatment of liver cancer.
In the present review, we focus on PPIs in the
PI3K/AKT and other important signaling pathways in liver cancer.
The potential antitumor therapies targeting these pivotal PPIs and
the strategies of how to investigate and analyze PPIs are also
assessed.
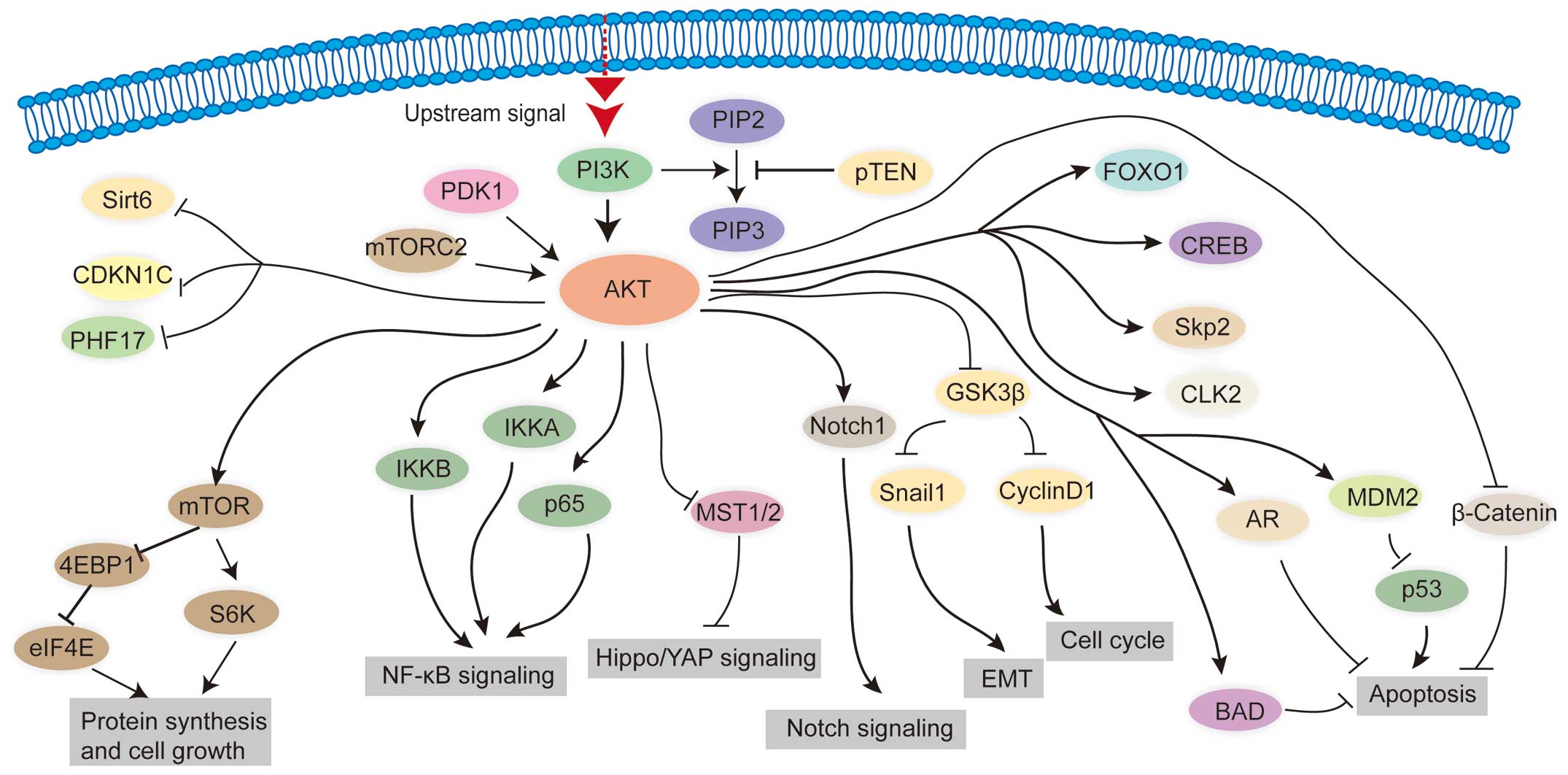
The PI3K/AKT pathway is an intracellular pathway
that is involved in cell cycle, growth, survival, proliferation and
migration. Enhanced PI3K/AKT activities have been reported in many
human cancer types, including cancers of colon, breast, brain,
liver, stomach and lung (8). The
PI3K/AKT signaling pathway can be activated by four main types of
sensors: the receptor tyrosine kinases (RTKs), cytokines, G
protein-coupled receptors and the integrins (9–11).
These four types of sensors bind with their cofactors and activate
downstream kinases in the PI3K families. PI3K, by transferring a
phosphoryl group, converts phosphatidylinositol 4,5-diphosphate
(PIP2) to phosphatidylinositol 3,4,5-triphosphate (PIP3) (12). PIP3 can interact with AKT which
contains pleckstrin homology (PH) domain on the inner surface of
the plasma membrane, resulting in conformational changes of these
proteins (13). Following binding
to PIP3 at the membrane, AKT can then be phosphorylated by
phosphoinositide-dependent kinase 1 (PDK1) at threonine 308 or be
phosphorylated by the mammalian target of rapamycin complex (mTORC)
at serine 473 (14,15). Fully phosphorylated AKT can directly
interact with >100 proteins including the mTORC,
Bcl-2-associated death promoter (BAD), caspase-9, various forkhead
box protein O (FOXO) proteins, glycogen synthase kinase 3 β
(GSK3β), mouse double minute 2 homolog (MDM2) and tuberous
sclerosis 1 (TSC1) (16).
AKT, the core protein in the PI3K/AKT signaling
pathway, can physically interact with proteins in other signaling
pathway. Thus, activity of the PI3K/AKT signaling pathway in liver
cancer can directly affect the activities of other signaling
pathways, such as Hippo/YAP, NF-κB, Wnt/β-catenin, Notch, p53,
JAK/STAT and MAPK/ERK signaling pathways.
AKT is shown to physically bind to the proteins,
IKKA and IKKB in the NF-κB signaling pathway. AKT phosphorylates
IKKA on threonine (Thr) 23, thereby activating the NF-κB signaling
pathway and subsequently inducing key immune and inflammatory
responses (17,18). In addition, IKKB is a direct target
of AKT, and activation of the AKT/IKKB signal is closely correlated
with the anti-apoptotic and pro-cell survival function of NF-κB
signaling in breast cancer cells (19). Similarly, the activation of AKT can
directly induce activation of the NF-κB signaling pathway and
eventually suppress apoptosis in liver cancer cells (20,21).
By contrast, the inactivation of AKT strongly prevents NF-κB
transcription factor p65 from entering the nucleus, the site at
which p65 exerts its effects, and subsequently induces apoptosis in
the HepG2 liver cancer cell line (22).
AKT is also capable of interacting and
phosphorylating MST1/2 kinases, key components of the Hippo/YAP
signaling pathway, on Thr 120. Such effects reduce the inhibitory
impact of MST1/2 on the activity of YAP, the terminal effector of
the Hippo/YAP signaling pathway, and thereby enhance AKT-maintained
cell survival signaling (23).
AKT was found to have the ability to activate the
Wnt/β-catenin signaling pathway by directly interacting with GSK3β,
a natural inhibitor of β-catenin. AKT represses GSK3β by initiating
its phosphorylation at serine 9 and vice versa (24). The phosphorylation of β-catenin via
GSK3β is then repressed, which facilitates β-catenin translocation
from the cytoplasm into the nucleus and Wnt/β-catenin signaling is
eventually activated (25,26). Notably, the close relationship
between the activation of PI3K/AKT and the upregulation of
Wnt/β-catenin activity was also observed in liver cancer (27). Additionally, promoted liver cancer
cell growth and proliferation was maintained by the PI3K/AKT and
Wnt/β-catenin signaling pathways (28). The interaction between PI3K/AKT and
Wnt/β-catenin is also critical for regulation of the cell cycle and
epithelial-mesenchymal transition (EMT) during tumor formation, as
following the respression of GSK3β by AKT, Wnt/β-catenin signaling
down-stream effectors, such as cyclin D1, Snail and Mucin 1, are
affected (29–31).
The MAPK/ERK is another important signaling pathway
involved in a variety of cell processes including proliferation,
differentiation, migration and survival. The MAPK/ERK signaling
pathway is frequently activated in liver cancer often due to
activating mutations or amplification of several components such as
Ras, Raf and MEK (32,33). Notably, emerging evidence suggests
that AKT is also capable of activating the MAPK/ERK signaling
pathway via interaction with MAPK/ERK signaling components.
Firstly, Raf, MAP3K5, MAP3K8 and MAP3K11 are activated by AKT via
phosphorylation (34–37). Secondly, MAPK2K4 is phosphorylated
by AKT on serine 78 to suppress apoptosis (38). Additionally, MAPK14/p38 is directly
bound to and activated by AKT, thus establishing a crosstalk
between the MAPK/ERK and PI3K/AKT signaling pathways (39). In addition to the direct binding to
the MAPK/ERK components, AKT can physically interact with certain
proteins, which can indirectly influence MAPK/ERK signaling
activity. For example, AKT is known to interact with MAPK8IP1/JIP1,
a regulator of MAPK8/JNK, to control inflammatory responses, cell
proliferation and apoptosis (40).
Furthermore, the downstream effectors of AKT, such as mTOR and
GSK3β bind to the core regulator in the MAPK/ERK signaling pathway.
For example, GSK3β physically interacts with and activates MAP3K1
in vitro and in vivo to regulate cell differentiation
and apoptosis (41).
PPIs are also important for the crosstalk between
PI3K/AKT and tumor suppressor p53 signaling pathways. AKT can
directly interact with and regulate MDM2, one of the most
well-characterized oncogenic ubiquitin E3 ligases that negatively
regulates p53 transcription activity. Activation of AKT leads to
phosphorylation of MDM2 on Ser 166 and 186, whereas inhibition of
AKT decreases such phosphorylated levels (42,43).
Phosphorylation of MDM2 decreases the protein levels of p53,
thereby suppressing apoptosis in liver cancer cells (44). When the cells were pretreated with
Wortmannin, a well-known PI3K/AKT inhibitor, to suppress AKT
activation, both the upregulation of phosphorylation of MDM2 and
downregulation of p53 were reversed in the HepG2 liver cancer cell
line (45).
As for the JNK/STAT signaling pathway, the
downstream effector of AKT, mTOR, has been shown to physically
interact with STAT1 and STAT3 and regulates the transcription
activity of these two transcription factors (50,51).
In general, according to the results of the
aforementioned studies, AKT can physically bind to a series of core
protein in different signaling pathways and activation of the AKT
signaling pathway can directly or indirectly lead to the activation
of several other signaling pathways. Thus, we suggest that PI3K/AKT
is central to the complex signaling network involved in liver as
well as in other organ tumorigenesis.
In liver cancer, deregulated PI3K/AKT signaling
pathway often leads to uncontrolled cell growth, metabolism,
survival, metastasis and tumor formation. The PPI between AKT and
mTOR and the mechanism associated with this interaction has been
largely investigated in liver cancer. mTOR exists in two different
complexes, mTOR complex 1 (mTORC1) and mTORC2. The mTORC2 complex
directly phosphorylates AKT on Ser473 and AKT conversely
phosphorylates mTORC1 at two COOH-terminal sites (Thr2446 and
Ser2448) (14,53). p70S6 kinase and translational
repressor protein 4E-binding protein 1 (4EBP1), the downstream
effector of mTOR, are then phosphorylated by mTOR and regulate the
translation of several important proliferative and angiogenic
factors, such as c-Myc, cyclin D1, hypoxia-inducible factor (HIF)
1α and vascular endothelial growth factor (VEGF) (53–55),
which are associated with tumor progression in liver cancer. The
deregulated expression of mTOR signaling effectors ia present in
40–50% of HCC and activation of mTOR is correlated with poor
prognosis and recurrence in HCC (56,57).
FOXO1 is also regarded as an AKT downstream
effector. AKT has been proven to interact with FOXO1, which is a
transcription factor involved in the regulation of gluconeogenesis
and glycogenolysis via insulin signaling, and FOXO1 is also central
to the decision for a preadipocyte to commit to adipogenesis
(58). When FOXO1 is phosphorylated
by AKT on Thr24, Ser256 and Ser319, it is spatially excluded from
the nucleus and is then readily ubiquitinated and degraded
(59). The phosphorylation of FOXO1
by AKT also impairs FOXO1-induced hepatic glucose production
through a reduction in the transcription of glucose 6-phosphatase
(G6PC) gene (60).
Additionally, BAD has been shown to physically
interact with AKT. BAD protein is a pro-apoptotic member of the
Bcl-2 gene family, which is involved in initiating apoptosis.
Dephosphorylated BAD forms a heterodimer with Bcl-2 or Bcl-xL,
represses them and thus initiates Bax/Bak-triggered apoptosis. When
BAD is phosphorylated by AKT, it forms the BAD-14-3-3 protein
heterodimer, allowing Bcl-2 to inhibit Bax-induced apoptosis
(61). Inactivation of AKT removes
its inhibitory effect to BAD, which may also decrease the levels of
anti-apoptotic Bcl-2 and Bcl-XL proteins, and eventually lead to
mitochondria-induced apoptosis in tumor cells (44). In liver cancer cells, AKT-mediated
inhibitory effects on BAD-induced mitochondrial apoptotic signals
were also observed (62).
AKT interacts with and activates S-phase
kinase-associated protein 2 (Skp2) through phosphorylation of this
protein on Ser72 (63,64). Skp2 behaves as an oncogene, and
overexpression of this protein is frequently observed in human
cancer progression and metastasis (wenxian). In human liver cancer
cell lines and a murine liver cancer model, overexpression of AKT
also led to the overexpression of Skp2 (65), indicating Skp2 may act as a
downstream oncogenic effector of AKT during liver
tumorigenesis.
Androgen receptor (AR) is activated by teh binding
of androgenic hormones testosterone or dihydrotestosterone in the
cytoplasm, and exerting its nuclear receptor function in the
nucleus (66). AKT is capable of
preventing AR from activation by androgen via the phosphorylation
of AR on Ser210 and Ser790 allowing AKT to suppress
androgen-induced apoptosis (67).
AR has been shown to promote the initiation and development of
liver cancer during the early stage of the disease but to suppress
liver cancer cell invasion during the later stages of the disease
(68). Evidence from Nie et
al (69) indicates that the
activation of AKT directly impacts AR to inhibit apoptosis in HCC
cells, suggesting the function of downstream AR responds to the
function of upstream AKT.
In addition to the PPIs between AKT and the proteins
described above, AKT can also interact with other proteins that are
found to play significant roles in liver cancer. These proteins
include T-cell leukemia/lymphoma protein 1 (TCL1) (70,71),
breast cancer type 1 susceptibility protein (BRCA1) (72), vimentin (73), integrin-linked kinase (ILK)
(74,75), and heat shock protein 27 (HSP27)
(76). The proteins that AKT can
bind to liver cancer are provided in Table I.
A number of novel AKT-involved PPIs have been
identified in other cancer types. For example, Nam et al
(77) found that Cdc-2-like kinase
2 (CLK2) is phosphorylated by AKT at Ser34 and Thr127 in
vitro and in vivo. This type of phosphorylation
significantly increased cell growth whereas it inhibits cell
apoptosis in HeLa cells. Snail1, a transcriptional factor essential
for triggering EMT, can directly interact and thus enhance
AKT-induced open chromatin around the Snail1-binding site within
the E-cadherin promoter in different cancer cells (78). Sirtuin-6 (Sirt6), a tumor suppressor
that plays negative roles on DNA repair, telomere maintenance,
glycolysis and inflammation, is directly inhibited by AKT through
phosphorylation and subsequent degradation by MDM2, and this type
of PPI between Sirt6 and AKT promotes tumorigenesis in breast
cancer (79). Cyclin-dependent
kinase inhibitor 1C (CDKN1C), an inhibitor of cyclin-dependent
kinases, is pivotal in regulating cell cycle progression. AKT
phosphorylates and inhibits CDKN1C on Ser 282 or Thr310, and then
promotes cell proliferation, transformational activity and
tumorigenicity in breast cancer cells (80). Although studies have mainly focused
on how AKT regulates activities of other proteins, few have
discussed how other proteins regulate AKT. Zeng et al
(81) reported Jade-1 is a novel
tumor suppressor that is bound to the catalytic domain and the
C-terminal regulatory tail of AKT. This PPI inhibits AKT kinase
activity and reduced Jade-1 expression in clear-cell renal cell
carcinoma and is regarded as a poor prognostic factor.
Cylindromatosis (CYLD) is a directly deubiquitinating enzyme that
triggers deubiquitination of K63-linked ubiquitination and
inactivation of AKT. CYLD deficiency releases its inhibition to AKT
and thereby promotes cell proliferation, glucose uptake and growth
of prostate tumors (82). These
AKT-involved PPIs were important in tumor initiation and
progression in other cancer types. However, whether and how these
PPIs are critical during liver tumorigenesis remains largely
unclear. Nevertheless, the role of these PPIs as novel therapeutic
targets in clinical treatment remains to be investigated.
Since the PI3K/AKT signaling pathway is a crucial
pathway in liver cancer formation and progression, targeting
PI3K/AKT pathway, these PI3K/AKT-involved physical PPIs in
particular are novel aspects in the clinical treatment of liver
cancer. mTOR inhibitors can abolish the interaction between AKT and
mTOR by inhibiting the phosphorylation of AKT on Ser 473 (83–85).
As PPI between AKT and mTOR are important in liver cancer, the use
of mTOR inhibitors, such as sirolimus, can significantly reduce the
recurrence of liver cancer in the post-liver transplantation
patient population (86). In a
recent meta-analysis including 474 patients, the 1-, 3- and 5-year
recurrence-free survival (RFS) and overall survival (OS) was
considerably improved for the sirolimus group in comparison with
the calcineurin inhibitors (CNIs) group. Lower recurrence, lower
recurrence-associated mortality and lower overall mortality were
observed in the sirolimus group compared to the CNIs group
(87). Other second-generation mTOR
inhibitors, such as everolimus, Pp242, OSI027, CC-223 and AZD8055,
have similar antitumor efficacy in liver cancer cell lines and
xenograft models (88). A phase 1/2
study including 28 patients revealed that everolimus is well
tolerated in patients with advanced liver cancer, and 10 mg/day was
defined as the phase 2 dose (89).
In another cohort of 36 patients, everolimus was observed to
repress cancer progression in patients with advanced liver cancer
when used at a maximum tolerated dose of 70 mg weekly (88,90).
In addition to the therapy targeting mTOR, which may
interrupt the PPI between mTOR and AKT, some drugs have been
identified to simultaneous inhibit more than one signaling pathway.
For example, hydroxytyrosol is capable of inhibiting cell
proliferation and inducing G2/M cell cycle arrest and apoptosis in
HCC cells by suppressing the PI3K/AKT and NF-κB signaling pathways
(91). OSU-A9, a potent
indole-3-carbinol-derived PI3K/AKT/NF-κB signaling pathway
inhibitor, can induce apoptosis by inactivating PI3K/AKT/NF-κB
signaling and killing HCC cells (92). NS398, a selective cyclooxygenase-2
(Cox2) inhibitor and simvastatin, a 3-hydroxy-3-methylglutaryl
coenzyme A reductase inhibitor, were previously simultaneously used
and this co-administration significantly reduced the activity of
the PI3K/AKT and NF-κB signaling pathways, leading to inhibited
liver cancer cell proliferation and induction of apoptosis
(93). Baicalein is another drug
that plays a negative role against liver cancer by targeting AKT
and β-catenin in clinical treatment (94). An in vitro study showed that
Baicalein has significant cytotoxicity against liver cancer cells
but moderate cytotoxicity against immortalized human hepatocytes,
suggesting Baicalein is an ideal drug that is less harmful to
normal cells as compared to cancer cells. Baicalein has the ability
to induce G0/G1-phase arrest in liver cancer cells by inhibiting
PI3K/AKT signaling and promoting degradation of β-catenin, the key
factor of Wnt/β-catenin signaling pathway. An in vivo study
also demonstrated that Baicalein impairs tumor growth in a
xenograft mouse model by inhibiting PI3K/AKT and β-catenin. Through
a similar mechanism against PI3K/AKT and β-catenin, BCT-100, a new
recombinant human arginase, has been revealed to inhibit cell
proliferation and enhance caspase-dependent cell apoptosis
(95). PI3K/AKT, mTORC1 and MEK
signaling can be simultaneously inhibited in vitro and in
vivo by a novel 2-pyrimidyl-5-amidothiazole compound, DC120.
Thus, this drug is able to suppress proliferation but also induce
apoptosis in liver cancer cells (96).
The treatment of ANISpm, a novel
3-amino-naphthalimide-spermine conjugate, results in the
inactivation of PI3K/AKT signaling followed by dissociation of BAD
from BAD-Bcl complexes and the induction of Bcl-mediated apoptosis
in liver cancer cells (99). The
treatment of 3-nitro-naphthalimide and nitrogen mustard conjugate
is another potential therapy that can induce apoptosis in liver
cancer cells through the inhibition of PI3K/AKT signaling (100).
Macromolecules (macrodrugs) can be developed that
interfere with PPIs by binding with high affinity and specificity
to contact surfaces. Although macrodrugs have inherent problems of
bio-distribution and delivery to target cells in patients, their
efficacies on the inhibition of cancers suggest that efforts to
achieve the goal of clinical use should be pursued (101). Targeting physical PPIs especially
PI3K/AKT-involved PPIs may become a new aspect in the clinical
treatment of liver cancer (Fig.
1).
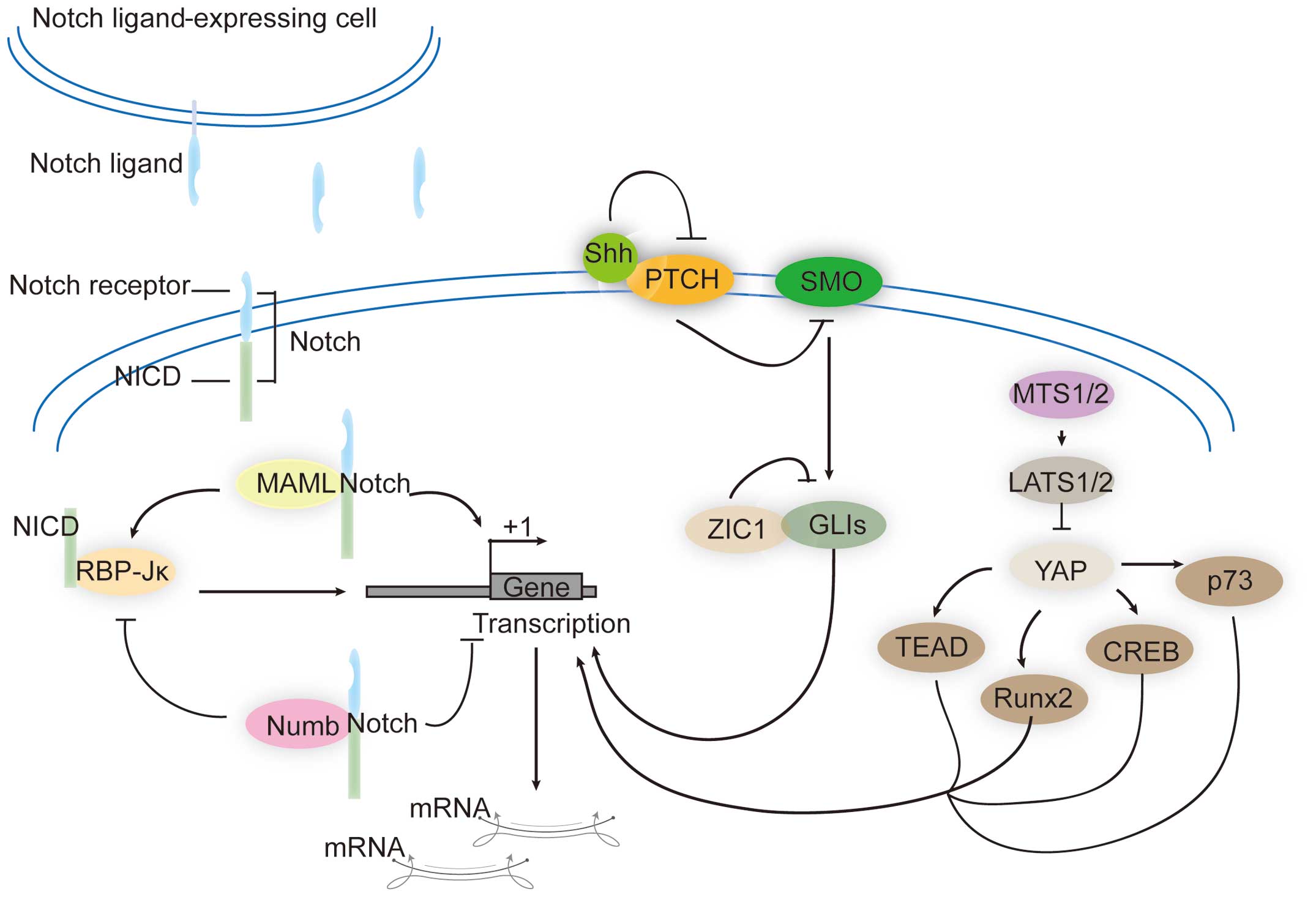
There are a number of signaling pathways that can
play oncogenic roles in liver cancer. For example, the Notch
signaling pathway is a highly conserved pathway in most
multicellular organisms (102).
The Notch system comprises four types of transmembrane Notch
receptors (Notch-1, -2, -3 and -4), and two types of ligands,
Serrate/Jagged (Jag-1 and -2) and Delta-like (Dll-1, -3 and -4)
(103). Extracellular epidermal
growth factor (EGF)-like repeats in Notch receptors can interact
with the delta serrate LAG-2 (DSL) domain in the ligands. Following
ligand binding to the receptors, the extracellular domain of Notch
receptors is cleaved by the γ-secretase complex, and the Notch
intracellular domain (NICD) is released. NICD then shuttles into
the nucleus and interacts with CBF1/Drosophila Su (H)/C.
elegans LAG-1 (CSL)-binding proteins, which are also known as
recombination signal binding protein immunoglobulin Jκ (RBP-Jκ).
The co-repressors of RBP-Jκ are then replaced and the expression of
a set of Notch target genes is activated (104). Notch proteins have been shown to
interact with a family of mastermind-like transcriptional
coactivators (MAML1, MAML2 and MAML3). MAML1 binds to the ankyrin
repeat domain of the four mammalian Notch receptors, forms a
DNA-binding complex with NICD and RBP-Jκ, and amplifies the
Notch-induced transcription of HES1 (105). A dominant negative form of MAML
can significantly reduce the proliferation of liver cancer cells
(106). By contrast, there are
also some PPIs that can inhibit Notch signaling. Protein Numb can
interact with Notch receptors and antagonize Notch signaling.
Mechanically, Numb recruits components of the ubiquitination
machinery to the Notch receptor and thereby facilitating
ubiquitination of Notch1 at the membrane and promoting the
degradation of NICD. Numb acts as a tumor suppressor, and its
function of inhibiting tumor cell proliferation occurs largely
through the suppression of Notch signaling. In liver cancer cells,
the downregulation of Numb is positively associated with activation
of Notch signaling-induced cell proliferation and growth (107).
Hedgehog (Hh) is another important protumorigenic
signaling pathway that was first identified by the Nobel laureates
Nüsslein-Volhard and Wieschaus through mutagenesis screening assays
in Drosophila (108). In
mammals, hedgehog homologues include the Desert hedgehog (DHh),
Indian hedgehog (IHh) and Sonic hedgehog (SHh). Hh proteins are
synthesized as ~45-kDa precursors, followed by modifications at the
amino-terminus with palmitic acids and carboxy-terminus with
cholesterol groups (109,110). Hh proteins can bind to the Protein
patched homolog (PTCH) receptor, which is a 12-span transmembrane
protein (111,112). PTCH is also a negative regulator
in Hh signaling because it can inhibit the activity of the 7-pass
transmembrane receptor-like protein smoothened (SMO) (113). Binding of Hh proteins to PTCH
leads to loss of the inhibitory activity of PTCH on SMO, which
initiates an intracellular signaling cascade by releasing GLI
proteins, terminal effectors of Hh signaling. These GLI proteins
then enter into the cell nucleus to activate the transcription of
Hh signaling target genes. The GLI proteins found in mammals
include GLI1, GLI2 and GLI3. Numerous genes have been found to be
regulated by these three GLI proteins. Overactivation of Hh
signaling is responsible for proliferative diseases, including
cancer. In 2006, Hh signaling was firstly studied in HCC and
investigators identified that SMO and GLI1 proteins are
overexpressed in established liver cancer cell lines and liver
cancer tissue samples. Furthermore, an increase in the
stoichiometric ratio of SMO to PTCH mRNA levels in liver cancer was
revealed to correlate with tumor size and be treated as a
prognostic marker of liver cancer (114,115). GLI1 expression in HCC tissues was
observed to be negatively associated with disease-free and overall
survival. Overexpression of GLI1 promotes the proliferation,
viability, colony formation, migration and invasion of liver cancer
cells, while silencing GLI1 expression in liver cancer cells leads
to the opposite output (116). The
protein zinc finger of the cerebellum 1 (ZIC1) interacts with GLI1
and repress the activity of GLI1 (117), thus ZIC1 is regarded as a tumor
suppressor. In liver cancers, methylation frequencies of ZIC1
promoter are significantly higher than those in the corresponding
non-cancerous tissues. Moreover, patients whose ZIC1 promoters are
methylated have poorer survival rates than those without such
methylation (118).
The Hippo/YAP signaling pathway has become a hot
research topic. This pathway was first identified in
Drosophila and controls organ size through the regulation of
cell proliferation and apoptosis. All of the core components of the
Hippo/YAP signaling pathway are conserved in mammalians. By
phosphorylating the terminal transcriptional regulator of the
Yes-associated protein (YAP) signaling pathway, large tumor
suppressor kinase (LATS) promotes the PPI between YAP and 14-3-3
proteins, which helps to anchor YAP in the cytoplasm and prevents
its transportation into the nucleus. When the upstream Hippo
signaling is inactivated, YAP can translocate into the nucleus and
bind to several transcription factors including p73, runt-related
transcription factor 2 (Runx2) and TEA-domain family member (TEAD)
protein families (119). A series
of recent studies have demonstrated that the core components of the
Hippo/YAP signaling pathway are important for liver tumorigenesis.
Approximately 50% of human HCCs show aberrant expression and
nuclear localization of YAP (120), with 30% HCCs showing low
phosphorylation of YAP on Ser127, a hallmark of the inactivation of
YAP (121). PPIs between YAP and
other proteins have gradually been identified, including SMADs
(122), p73 (123), ErbB4 (124), TEADs (119), RUNX (125), angiomotins (AMOTs) (126–128), zona occludens 1/2 (ZO1/2)
(129) and LATS1/2 (130). In liver cancer, these PPIs play
pivotal roles in promoting or inhibiting tumor formation. LATS
kinases inhibit YAP function by promoting the cytoplasmic retention
of YAP by phosphorylating YAP on Ser127 (131). A significant decrease in the
expression and activity of LATS kinases is evident in HCC and CCC
(132). YAP has been shown to
interact with the TEAD family of transcription factors and
upregulate genes that promote cell growth and inhibit apoptosis
(133,134). The YAP-TEAD complex is important
in YAP-overexpressing cancers and disruption of the YAP-TEAD
interaction may provide an important approach for the treatment of
liver cancer. The treatment of various types of cancer with
verteporfin (VP) to disrupt the PPI between YAP and TEAD was
suggested to become a new approach in clinical treatment (135). In addition to TEAD, YAP has been
found to interact with tight junction proteins angiomotin (AMOT)
and zona occludens-2 (ZO-2). AMOT acts as a YAP cofactor,
preventing YAP phosphorylation and increasing its activity towards
a specific set of genes that facilitate tumorigenesis in liver
cancer. However, the functional role of PPI between YAP and ZO-2 in
liver cancer remains to be investigated. These direct PPIs in the
Notch, hedgehog, and Hippo/YAP pathways that are associated with
liver cancer are shown in Fig.
2.
Massive identification of PPIs generates numerous
interactions, which are collected together in PPI databases that
are continuously updated to provide complete interactions. The
database of interacting proteins (DIP) is the first PPI database
globally (154). The number of
public PPI databases has increased rapidly. For example, the
Biological General Repository for Interaction Datasets (BioGRID:
http://thebi-ogrid.org) is an open access
database that houses genetic and protein interactions curated from
the primary biomedical literature for all major model organisms and
human. BioGRID contains 749,912 interactions as drawn from 43,149
publications describing studies in >30 model organisms (155). Another example is the Protein
Interaction Network Analysis (PINA) platform, which is for protein
interaction network construction, filtering, analysis,
visualization and management. It integrates PPI data from six
public curated databases and constructs a complete, non-redundant
protein interaction dataset for six model organisms. PINA also
provides a variety of built-in tools to filter and analyze the
network to gain insight into the network (156).
In summary, signal transduction plays a fundamental
role in many biological processes as well as in many diseases. In
liver cancer, many signaling pathways including PI3K/AKT,
Ras/Mek/ERK, IKK/NF-κB, Wnt/β-catenin, TGF-β, Notch, hedgehog and
Hippo/YAP are shown to be dysregulated. The majority of these
signaling pathways have PPIs with PI3K/AKT signaling pathways.
Identification of PPI is the crucial step involved in identifying
the signal transduction pathways. Signals propagation to inside
and/or along the interior of cells depends on PPIs between the
various signaling molecules. Numerous properties of PPI such as
allosteric sites and hotspots, have been incorporated into
drug-design strategies. The relevance of PPI as putative
therapeutic targets for the development of new treatments is
particularly evident in liver cancer. The investigation of PPIs in
signaling pathways may provide knowledge on biochemical cascades
and disease pathogenesis, and new therapeutic targets in liver
cancer.
The present study was supported by the Yangfan
Project of the Shanghai Committee of Science and Technology
(14YF1412300), the Outstanding Youth Training Program of Tongji
University (1501219080) and the Shanghai Tenth People's Hospital
Climbing Training Program (04.01.13024).
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arzumanyan A, Reis HM and Feitelson MA:
Pathogenic mechanisms in HBV- and HCV-associated hepatocellular
carcinoma. Nat Rev Cancer. 13:123–135. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Squires RH, Ng V, Romero R, Ekong U,
Hardikar W, Emre S and Mazariegos GV: Evaluation of the pediatric
patient for liver transplantation: 2014 practice guideline by the
American Association for the Study of Liver Diseases, American
Society of Transplantation and the North American Society for
Pediatric Gastroenterology, Hepatology, and Nutrition. J Pediatr
Gastroenterol Nutr. 59:112–131. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lo CM: Liver transplantation in 2012:
Transplantation for liver cancer - more with better results. Nat
Rev Gastroenterol Hepatol. 10:74–76. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lie PP, Cheng CY and Mruk DD: Signalling
pathways regulating the blood-testis barrier. Int J Biochem Cell
Biol. 45:621–625. 2013. View Article : Google Scholar :
|
|
6
|
Kandori H, Sudo Y and Furutani Y:
Protein-protein interaction changes in an archaeal light-signal
transduction. J Biomed Biotechnol. 2010(424760)2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chakraborty C, Doss CGP, Chen L and Zhu H:
Evaluating protein-protein interaction (PPI) networks for diseases
pathway, target discovery, and drug-design using 'in silico
pharmacology'. Curr Protein Pept Sci. 15:561–571. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pal I and Mandal M: PI3K and Akt as
molecular targets for cancer therapy: Current clinical outcomes.
Acta Pharmacol Sin. 33:1441–1458. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Danielsen SA, Eide PW, Nesbakken A, Guren
T, Leithe E and Lothe RA: Portrait of the PI3K/AKT pathway in
colorectal cancer. Biochim Biophys Acta. 1855:104–121. 2015.
|
|
10
|
Chang F, Lee JT, Navolanic PM, Steelman
LS, Shelton JG, Blalock WL, Franklin RA and McCubrey JA:
Involvement of PI3K/Akt pathway in cell cycle progression,
apoptosis, and neoplastic transformation: A target for cancer
chemotherapy. Leukemia. 17:590–603. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sabine VS, Crozier C, Brookes CL, Drake C,
Piper T, van de Velde CJ, Hasenburg A, Kieback DG, Markopoulos C,
Dirix L, et al: Mutational analysis of PI3K/AKT signaling pathway
in tamoxifen exemestane adjuvant multinational pathology study. J
Clin Oncol. 32:2951–2958. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Safdari Y, Khalili M, Ebrahimzadeh MA,
Yazdani Y and Farajnia S: Natural inhibitors of PI3K/AKT signaling
in breast cancer: Emphasis on newly-discovered molecular mechanisms
of action. Pharmacol Res. 93:1–10. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Matsuoka T and Yashiro M: The role of
PI3K/Akt/mTOR signaling in gastric carcinoma. Cancers (Basel).
6:1441–1463. 2014. View Article : Google Scholar
|
|
14
|
Sarbassov DD, Guertin DA, Ali SM and
Sabatini DM: Phosphorylation and regulation of Akt/PKB by the
rictor-mTOR complex. Science. 307:1098–1101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jacinto E, Facchinetti V, Liu D, Soto N,
Wei S, Jung SY, Huang Q, Qin J and Su B: SIN1/MIP1 maintains
rictor-mTOR complex integrity and regulates Akt phosphorylation and
substrate specificity. Cell. 127:125–137. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Manning BD and Cantley LC: AKT/PKB
signaling: Navigating downstream. Cell. 129:1261–1274. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ozes ON, Mayo LD, Gustin JA, Pfeffer SR,
Pfeffer LM and Donner DB: NF-kappaB activation by tumour necrosis
factor requires the Akt serine-threonine kinase. Nature. 401:82–85.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu Y and Wahl LM: Production of matrix
metalloproteinase-9 by activated human monocytes involves a
phosphatidylinositol-3 kinase/Akt/IKKalpha/NF-kappaB pathway. J
Leukoc Biol. 78:259–265. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vandermoere F, El Yazidi-Belkoura I,
Adriaenssens E, Lemoine J and Hondermarck H: The antiapoptotic
effect of fibroblast growth factor-2 is mediated through nuclear
factor-kappaB activation induced via interaction between Akt and
IkappaB kinase-beta in breast cancer cells. Oncogene. 24:5482–5491.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tu CC, Cheng LH, Hsu HH, Chen LM, Lin YM,
Chen MC, Lee NH, Tsai FJ, Huang CY and Wu WJ: Activation of snail
and EMT-like signaling via the IKKαβ/NF-κB pathway in
Apicidin-resistant HA22T hepatocellular carcinoma cells. Chin J
Physiol. 56:326–333. 2013. View Article : Google Scholar
|
|
21
|
Noh JH, Bae HJ, Eun JW, Shen Q, Park SJ,
Kim HS, Nam B, Shin WC, Lee EK, Lee K, et al: HDAC2 provides a
critical support to malignant progression of hepatocellular
carcinoma through feedback control of mTORC1 and AKT. Cancer Res.
74:1728–1738. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang R, Cao X, Wang C, Hou L, Nie J, Zhou
M and Feng Y: An antitumor peptide from Musca domestica pupae
(MATP) induces apoptosis in HepG2 cells through a JNK-mediated and
Akt-mediated NF-κB pathway. Anticancer Drugs. 23:827–835. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yuan Z, Kim D, Shu S, Wu J, Guo J, Xiao L,
Kaneko S, Coppola D and Cheng JQ: Phosphoinositide 3-kinase/Akt
inhibits MST1-mediated pro-apoptotic signaling through
phosphorylation of threonine 120. J Biol Chem. 285:3815–3824. 2010.
View Article : Google Scholar
|
|
24
|
Wang CY, Tsai AC, Peng CY, Chang YL, Lee
KH, Teng CM and Pan SL: Dehydrocostuslactone suppresses
angiogenesis in vitro and in vivo through inhibition of Akt/GSK-3β
and mTOR signaling pathways. PLoS One. 7:e311952012. View Article : Google Scholar
|
|
25
|
Su Y, Fu C, Ishikawa S, Stella A, Kojima
M, Shitoh K, Schreiber EM, Day BW and Liu B: APC is essential for
targeting phosphorylated beta-catenin to the SCFbeta-TrCP ubiquitin
ligase. Mol Cell. 32:652–661. 2008. View Article : Google Scholar
|
|
26
|
Bauer L, Langer R, Becker K, Hapfelmeier
A, Ott K, Novotny A, Höfler H and Keller G: Expression profiling of
stem cell-related genes in neoadjuvant-treated gastric cancer: A
NOTCH2, GSK3B and β-catenin gene signature predicts survival. PLoS
One. 7:e445662012. View Article : Google Scholar
|
|
27
|
Hsieh CH, Cheng LH, Hsu HH, Ho TJ, Tu CC,
Lin YM, Chen MC, Tsai FJ, Hsieh YL and Huang CY: Apicidin-resistant
HA22T hepatocellular carcinoma cells strongly activated the
Wnt/β-catenin signaling pathway and MMP-2 expression via the
IGF-IR/PI3K/Akt signaling pathway enhancing cell metastatic effect.
Biosci Biotechnol Biochem. 77:2397–2404. 2013. View Article : Google Scholar
|
|
28
|
Mavila N, James D, Utley S, Cu N, Coblens
O, Mak K, Rountree CB, Kahn M and Wang KS: Fibroblast growth factor
receptor-mediated activation of AKT-β-catenin-CBP pathway regulates
survival and proliferation of murine hepatoblasts and hepatic tumor
initiating stem cells. PLoS One. 7:e504012012. View Article : Google Scholar
|
|
29
|
Liu L, Dai Y, Chen J, Zeng T, Li Y, Chen
L, Zhu YH, Li J, Li Y, Ma S, et al: Maelstrom promotes
hepatocellular carcinoma metastasis by inducing
epithelial-mesenchymal transition by way of Akt/GSK-3β/Snail
signaling. Hepatology. 59:531–543. 2014. View Article : Google Scholar
|
|
30
|
Gotoh J, Obata M, Yoshie M, Kasai S and
Ogawa K: Cyclin D1 over-expression correlates with beta-catenin
activation, but not with H-ras mutations, and phosphorylation of
Akt, GSK3 beta and ERK1/2 in mouse hepatic carcinogenesis.
Carcinogenesis. 24:435–442. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Woo JK, Choi Y, Oh SH, Jeong JH, Choi DH,
Seo HS and Kim CW: Mucin 1 enhances the tumor angiogenic response
by activation of the AKT signaling pathway. Oncogene. 31:2187–2198.
2012. View Article : Google Scholar
|
|
32
|
Niault TS and Baccarini M: Targets of Raf
in tumorigenesis. Carcinogenesis. 31:1165–1174. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nikolaev SI, Rimoldi D, Iseli C, Valsesia
A, Robyr D, Gehrig C, Harshman K, Guipponi M, Bukach O, Zoete V, et
al: Exome sequencing identifies recurrent somatic MAP2K1 and MAP2K2
mutations in melanoma. Nat Genet. 44:133–139. 2012. View Article : Google Scholar
|
|
34
|
Barthwal MK, Sathyanarayana P, Kundu CN,
Rana B, Pradeep A, Sharma C, Woodgett JR and Rana A: Negative
regulation of mixed lineage kinase 3 by protein kinase B/AKT leads
to cell survival. J Biol Chem. 278:3897–3902. 2003. View Article : Google Scholar
|
|
35
|
Kim AH, Khursigara G, Sun X, Franke TF and
Chao MV: Akt phosphorylates and negatively regulates apoptosis
signal-regulating kinase 1. Mol Cell Biol. 21:893–901. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zimmermann S and Moelling K:
Phosphorylation and regulation of Raf by Akt (protein kinase B).
Science. 286:1741–1744. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kane LP, Mollenauer MN, Xu Z, Turck CW and
Weiss A: Akt-dependent phosphorylation specifically regulates Cot
induction of NF-kappa B-dependent transcription. Mol Cell Biol.
22:5962–5974. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Park HS, Kim MS, Huh SH, Park J, Chung J,
Kang SS and Choi EJ: Akt (protein kinase B) negatively regulates
SEK1 by means of protein phosphorylation. J Biol Chem.
277:2573–2578. 2002. View Article : Google Scholar
|
|
39
|
Rane MJ, Coxon PY, Powell DW, Webster R,
Klein JB, Pierce W, Ping P and McLeish KR: p38 Kinase-dependent
MAPKAPK-2 activation functions as 3-phosphoinositide-dependent
kinase-2 for Akt in human neutrophils. J Biol Chem. 276:3517–3523.
2001. View Article : Google Scholar
|
|
40
|
Nishitani Y and Matsumoto H: Ethanol
rapidly causes activation of JNK associated with ER stress under
inhibition of ADH. FEBS Lett. 580:9–14. 2006. View Article : Google Scholar
|
|
41
|
Kim JW, Lee JE, Kim MJ, Cho EG, Cho SG and
Choi EJ: Glycogen synthase kinase 3 beta is a natural activator of
mitogen-activated protein kinase/extracellular signal-regulated
kinase kinase kinase 1 (MEKK1). J Biol Chem. 278:13995–14001. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhou BP, Liao Y, Xia W, Zou Y, Spohn B and
Hung MC: HER-2/neu induces p53 ubiquitination via Akt-mediated MDM2
phosphorylation. Nat Cell Biol. 3:973–982. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ashcroft M, Ludwig RL, Woods DB, Copeland
TD, Weber HO, MacRae EJ and Vousden KH: Phosphorylation of HDM2 by
Akt. Oncogene. 21:1955–1962. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fu Z, Ren L, Wei H, Lv J, Che X, Zhu Z,
Jia J, Wang L, Lin G, Lu R, et al: Effects of Tyroserleutide on
phosphatidylinositol 3′-kinase/AKT pathway in human hepatocellular
carcinoma cell. J Drug Target. 22:146–155. 2014. View Article : Google Scholar
|
|
45
|
Wang C, Qi R, Li N, Wang Z, An H, Zhang Q,
Yu Y and Cao X: Notch1 signaling sensitizes tumor necrosis
factor-related apoptosis-inducing ligand-induced apoptosis in human
hepato-cellular carcinoma cells by inhibiting Akt/Hdm2-mediated p53
degradation and up-regulating p53-dependent DR5 expression. J Biol
Chem. 284:16183–16190. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Song J, Park S, Kim M and Shin I:
Down-regulation of Notch-dependent transcription by Akt in vitro.
FEBS Lett. 582:1693–1699. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xu L, Zhu Y, Xu J, Wu K, Li J, Xu W, Liu
H, Wang S, Yin H, Chen L, et al: Notch1 activation promotes renal
cell carcinoma growth via PI3K/Akt signaling. Cancer Sci.
103:1253–1258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huntzicker EG, Hötzel K, Choy L, Che L,
Ross J, Pau G, Sharma N, Siebel CW, Chen X and French DM:
Differential effects of targeting Notch receptors in a mouse model
of liver cancer. Hepatology. 61:942–952. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Brana I, Berger R, Golan T, Haluska P,
Edenfield J, Fiorica J, Stephenson J, Martin LP, Westin S, Hanjani
P, et al: A parallel-arm phase I trial of the humanised anti-IGF-1R
antibody dalotuzumab in combination with the AKT inhibitor MK-2206,
the mTOR inhibitor ridaforolimus, or the NOTCH inhibitor MK-0752,
in patients with advanced solid tumours. Br J Cancer.
111:1932–1944. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fielhaber JA, Han YS, Tan J, Xing S, Biggs
CM, Joung KB and Kristof AS: Inactivation of mammalian target of
rapamycin increases STAT1 nuclear content and transcriptional
activity in alpha4- and protein phosphatase 2A-dependent fashion. J
Biol Chem. 284:24341–24353. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang F, Zhang W, Li D and Zhan Q: Gadd45a
suppresses tumor angiogenesis via inhibition of the mTOR/STAT3
protein pathway. J Biol Chem. 288:6552–6560. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sekulić A, Hudson CC, Homme JL, Yin P,
Otterness DM, Karnitz LM and Abraham RT: A direct linkage between
the phosphoinositide 3-kinase-AKT signaling pathway and the
mammalian target of rapamycin in mitogen-stimulated and transformed
cells. Cancer Res. 60:3504–3513. 2000.
|
|
53
|
Dunlop EA and Tee AR: Mammalian target of
rapamycin complex 1: Signalling inputs, substrates and feedback
mechanisms. Cell Signal. 21:827–835. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Mamane Y, Petroulakis E, LeBacquer O and
Sonenberg N: mTOR, translation initiation and cancer. Oncogene.
25:6416–6422. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhou Q, Lui VW and Yeo W: Targeting the
PI3K/Akt/mTOR pathway in hepatocellular carcinoma. Future Oncol.
7:1149–1167. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhou L, Huang Y, Li J and Wang Z: The mTOR
pathway is associated with the poor prognosis of human
hepatocellular carcinoma. Med Oncol. 27:255–261. 2010. View Article : Google Scholar
|
|
57
|
Nissen NN1, Menon V, Bresee C, Tran TT,
Annamalai A, Poordad F, Fair JH, Klein AS, Boland B and Colquhoun
SD: Recurrent hepatocellular carcinoma after liver transplant:
identifying the high-risk patient. HPB. 13:626–632. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Nakae J, Kitamura T, Kitamura Y, Biggs WH
III, Arden KC and Accili D: The forkhead transcription factor Foxo1
regulates adipocyte differentiation. Dev Cell. 4:119–129. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Hay N: Interplay between FOXO, TOR, and
Akt. Biochim Biophys Acta. 1813:1965–1970. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Nakae J, Kitamura T, Silver DL and Accili
D: The forkhead transcription factor Foxo1 (Fkhr) confers insulin
sensitivity onto glucose-6-phosphatase expression. J Clin Invest.
108:1359–1367. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Khor TO, Gul YA, Ithnin H and Seow HF:
Positive correlation between overexpression of phospho-BAD with
phosphorylated Akt at serine 473 but not threonine 308 in
colorectal carcinoma. Cancer Lett. 210:139–150. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Huang CS, Lee YR, Chen CS, Tu SH, Wang YJ,
Lee CH, Chen LC, Chang HW, Chang CH, Chih-Ming S, et al: Long-term
ethanol exposure causes human liver cancer cells to become
resistant to mitomycin C treatment through the inactivation of
bad-mediated apoptosis. Mol Carcinog. 49:728–738. 2010.PubMed/NCBI
|
|
63
|
Carrano AC and Pagano M: Role of the F-box
protein Skp2 in adhesion-dependent cell cycle progression. J Cell
Biol. 153:1381–1390. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lin HK, Wang G, Chen Z, Teruya-Feldstein
J, Liu Y, Chan CH, Yang WL, Erdjument-Bromage H, Nakayama KI, Nimer
S, et al: Phosphorylation-dependent regulation of cytosolic
localization and oncogenic function of Skp2 by Akt/PKB. Nat Cell
Biol. 11:420–432. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ho C, Wang C, Mattu S, Destefanis G, Ladu
S, Delogu S, Armbruster J, Fan L, Lee SA, Jiang L, et al: AKT
(v-akt murine thymoma viral oncogene homolog 1) and N-Ras
(neuroblastoma ras viral oncogene homolog) coactivation in the
mouse liver promotes rapid carcinogenesis by way of mTOR (mammalian
target of rapamycin complex 1), FOXM1 (forkhead box M1)/SKP2, and
c-Myc pathways. Hepatology. 55:833–845. 2012. View Article : Google Scholar :
|
|
66
|
Lu NZ, Wardell SE, Burnstein KL, Defranco
D, Fuller PJ, Giguere V, Hochberg RB, McKay L, Renoir JM, Weigel
NL, et al: International Union of Pharmacology. LXV. The
pharmacology and classification of the nuclear receptor
superfamily: Glucocorticoid, mineralocorticoid, progesterone, and
androgen receptors. Pharmacol Rev. 58:782–797. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Lin HK, Yeh S, Kang HY and Chang C: Akt
suppresses androgen-induced apoptosis by phosphorylating and
inhibiting androgen receptor. Proc Natl Acad Sci USA. 98:7200–7205.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ma WL, Jeng LB, Lai HC, Liao PY and Chang
C: Androgen receptor enhances cell adhesion and decreases cell
migration via modulating β1-integrin-AKT signaling in
hepatocellular carcinoma cells. Cancer Lett. 351:64–71. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Nie H, Cao Q, Zhu L, Gong Y, Gu J and He
Z: Acetylcholine acts on androgen receptor to promote the migration
and invasion but inhibit the apoptosis of human hepatocarcinoma.
PLoS One. 8:e616782013. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Hong X, Song R, Song H, Zheng T, Wang J,
Liang Y, Qi S, Lu Z, Song X, Jiang H, et al: PTEN antagonises
Tcl1/hnRNPK-mediated G6PD pre-mRNA splicing which contributes to
hepatocarcinogenesis. Gut. 63:1635–1647. 2014. View Article : Google Scholar
|
|
71
|
Wang XQ, Ongkeko WM, Chen L, Yang ZF, Lu
P, Chen KK, Lopez JP, Poon RT and Fan ST: Octamer 4 (Oct4) mediates
chemotherapeutic drug resistance in liver cancer cells through a
potential Oct4-AKT-ATP-binding cassette G2 pathway. Hepatology.
52:528–539. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Vasan N, Yelensky R, Wang K, Moulder S,
Dzimitrowicz H, Avritscher R, Wang B, Wu Y, Cronin MT, Palmer G, et
al: A targeted next-generation sequencing assay detects a high
frequency of therapeutically targetable alterations in primary and
metastatic breast cancers: Implications for clinical practice.
Oncologist. 19:453–458. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Kittaka N, Takemasa I, Takeda Y, Marubashi
S, Nagano H, Umeshita K, Dono K, Matsubara K, Matsuura N and Monden
M: Molecular mapping of human hepatocellular carcinoma provides
deeper biological insight from genomic data. Eur J Cancer.
44:885–897. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Chan J, Ko FC, Yeung YS, Ng IO and Yam JW:
Integrin-linked kinase overexpression and its oncogenic role in
promoting tumorigenicity of hepatocellular carcinoma. PLoS One.
6:e169842011. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Peroukides S, Bravou V, Varakis J,
Alexopoulos A, Kalofonos H and Papadaki H: ILK overexpression in
human hepatocellular carcinoma and liver cirrhosis correlates with
activation of Akt. Oncol Rep. 20:1337–1344. 2008.PubMed/NCBI
|
|
76
|
Cui Y, Wu W, Zhou Y, Xie Q, Liu T, Jin J
and Liu K: HSP27 expression levels are associated with the
sensitivity of hepatocellular carcinoma cells to
17-allylamino-17-demethoxygeldanamycin. Future Oncol. 9:411–418.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Nam SY, Seo HH, Park HS, An S, Kim JY,
Yang KH, Kim CS, Jeong M and Jin YW: Phosphorylation of CLK2 at
serine 34 and threonine 127 by AKT controls cell survival after
ionizing radiation. J Biol Chem. 285:31157–31163. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Villagrasa P, Díaz VM, Viñas-Castells R,
Peiró S, Del Valle-Pérez B, Dave N, Rodríguez-Asiain A, Casal JI,
Lizcano JM, Duñach M, et al: Akt2 interacts with Snail1 in the
E-cadherin promoter. Oncogene. 31:4022–4033. 2012. View Article : Google Scholar
|
|
79
|
Thirumurthi U, Shen J, Xia W, LaBaff AM,
Wei Y, Li CW, Chang WC, Chen CH, Lin HK, Yu D, et al: MDM2-mediated
degradation of SIRT6 phosphorylated by AKT1 promotes tumorigenesis
and trastuzumab resistance in breast cancer. Sci Signal.
7:ra712014. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zhao R, Yang HY, Shin J, Phan L, Fang L,
Che TF, Su CH, Yeung SC and Lee MH: CDK inhibitor
p57Kip2 is downregulated by Akt during HER2-mediated
tumorigenicity. Cell Cycle. 12:935–943. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zeng L, Bai M, Mittal AK, El-Jouni W, Zhou
J, Cohen DM, Zhou MI and Cohen HT: Candidate tumor suppressor and
pVHL partner Jade-1 binds and inhibits AKT in renal cell carcinoma.
Cancer Res. 73:5371–5380. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Yang WL, Jin G, Li CF, Jeong YS, Moten A,
Xu D, Feng Z, Chen W, Cai Z, Darnay B, et al: Cycles of
ubiquitination and deubiquitination critically regulate growth
factor-mediated activation of Akt signaling. Sci Signal. 6:ra32013.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Thoreen CC, Kang SA, Chang JW, Liu Q,
Zhang J, Gao Y, Reichling LJ, Sim T, Sabatini DM and Gray NS: An
ATP-competitive mammalian target of rapamycin inhibitor reveals
rapamycin-resistant functions of mTORC1. J Biol Chem.
284:8023–8032. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Yu K, Toral-Barza L, Shi C, Zhang WG,
Lucas J, Shor B, Kim J, Verheijen J, Curran K, Malwitz DJ, et al:
Biochemical, cellular, and in vivo activity of novel
ATP-competitive and selective inhibitors of the mammalian target of
rapamycin. Cancer Res. 69:6232–6240. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
García-Martínez JM, Moran J, Clarke RG,
Gray A, Cosulich SC, Chresta CM and Alessi DR: Ku-0063794 is a
specific inhibitor of the mammalian target of rapamycin (mTOR).
Biochem J. 421:29–42. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Monaco AP: The role of mTOR inhibitors in
the management of posttransplant malignancy. Transplantation.
87:157–163. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Menon KV, Hakeem AR and Heaton ND:
Meta-analysis: Recurrence and survival following the use of
sirolimus in liver transplantation for hepatocellular carcinoma.
Aliment Pharmacol Ther. 37:411–419. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Ashworth RE and Wu J: Mammalian target of
rapamycin inhibition in hepatocellular carcinoma. World J Hepatol.
6:776–782. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Zhu AX, Abrams TA, Miksad R, Blaszkowsky
LS, Meyerhardt JA, Zheng H, Muzikansky A, Clark JW, Kwak EL, Schrag
D, et al: Phase 1/2 study of everolimus in advanced hepatocellular
carcinoma. Cancer. 117:5094–5102. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Chen L, Shiah HS, Chen CY, Lin YJ, Lin PW,
Su WC and Chang JY: Randomized, phase I, and pharmacokinetic (PK)
study of RAD001, and mTOR inhibitor, in patients (pts) with
advanced hepatocellular carcinoma (HCC). J Clin Oncol.
27(4587)2009.
|
|
91
|
Zhao B, Ma Y, Xu Z, Wang J, Wang F, Wang
D, Pan S, Wu Y, Pan H, Xu D, et al: Hydroxytyrosol, a natural
molecule from olive oil, suppresses the growth of human
hepatocellular carcinoma cells via inactivating AKT and nuclear
factor-kappa B pathways. Cancer Lett. 347:79–87. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Omar HA: Arafa el SA, Maghrabi IA and Weng
JR: Sensitization of hepatocellular carcinoma cells to Apo2L/TRAIL
by a novel Akt/NF-kappaB signalling inhibitor. Basic Clin Pharmacol
Toxicol. 114:464–471. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Lee SJ, Hwang JW, Yim H, Yim HJ, Woo SU,
Suh SJ, Hyun JJ, Jung SW, Koo JS, Kim JH, et al: Synergistic effect
of simvastatin plus NS398 on inhibition of proliferation and
survival in hepatocellular carcinoma cell line. J Gastroenterol
Hepatol. 29:1299–1307. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zheng YH, Yin LH, Grahn TH, Ye AF, Zhao YR
and Zhang QY: Anticancer effects of baicalein on hepatocellular
carcinoma cells. Phytother Res. 28:1342–1348. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Chow AK, Ng L, Sing Li H, Cheng CW, Lam
CS, Yau TC, Cheng PN, Fan ST, Poon RT and Pang RW: Anti-tumor
efficacy of a recombinant human arginase in human hepatocellular
carcinoma. Curr Cancer Drug Targets. 12:1233–1243. 2012.PubMed/NCBI
|
|
96
|
Yang F, Deng R, Qian XJ, Chang SH, Wu XQ,
Qin J, Feng GK, Ding K and Zhu XF: Feedback loops blockade
potentiates apoptosis induction and antitumor activity of a novel
AKT inhibitor DC120 in human liver cancer. Cell Death Dis.
5:e11142014. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Vassilev LT, Vu BT, Graves B, Carvajal D,
Podlaski F, Filipovic Z, Kong N, Kammlott U, Lukacs C, Klein C, et
al: In vivo activation of the p53 pathway by small-molecule
antagonists of MDM2. Science. 303:844–848. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Li M, Zhang Z, Hill DL, Wang H and Zhang
R: Curcumin, a dietary component, has anticancer,
chemosensitization, and radiosensitization effects by
down-regulating the MDM2 oncogene through the PI3K/mTOR/ETS2
pathway. Cancer Res. 67:1988–1996. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Li M, Li Q, Zhang YH, Tian ZY, Ma HX, Zhao
J, Xie SQ and Wang CJ: Antitumor effects and preliminary systemic
toxicity of ANISpm in vivo and in vitro. Anticancer Drugs.
24:32–42. 2013. View Article : Google Scholar
|
|
100
|
Xie SQ, Zhang YH, Li Q, Xu FH, Miao JW,
Zhao J and Wang CJ: 3-Nitro-naphthalimide and nitrogen mustard
conjugate NNM-25 induces hepatocellular carcinoma apoptosis via
PARP-1/p53 pathway. Apoptosis. 17:725–734. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Tanaka T and Rabbitts TH: Interfering with
protein-protein interactions: Potential for cancer therapy. Cell
Cycle. 7:1569–1574. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Artavanis-Tsakonas S, Rand MD and Lake RJ:
Notch signaling: Cell fate control and signal integration in
development. Science. 284:770–776. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Brou C, Logeat F, Gupta N, Bessia C,
LeBail O, Doedens JR, Cumano A, Roux P, Black RA and Israël A: A
novel proteolytic cleavage involved in Notch signaling: The role of
the disintegrin-metalloprotease TACE. Mol Cell. 5:207–216. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Ayaz F and Osborne BA: Non-canonical notch
signaling in cancer and immunity. Front Oncol. 4(345)2014.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Wu L, Aster JC, Blacklow SC, Lake R,
Artavanis-Tsakonas S and Griffin JD: MAML1, a human homologue of
Drosophila mastermind, is a transcriptional co-activator for NOTCH
receptors. Nat Genet. 26:484–489. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Villanueva A, Alsinet C, Yanger K, Hoshida
Y, Zong Y, Toffanin S, Rodriguez-Carunchio L, Solé M, Thung S,
Stanger BZ, et al: Notch signaling is activated in human
hepatocellular carcinoma and induces tumor formation in mice.
Gastroenterology. 143:1660–1669.e7. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Liu M, Lee DF, Chen CT, Yen CJ, Li LY, Lee
HJ, Chang CJ, Chang WC, Hsu JM, Kuo HP, et al: IKKα activation of
NOTCH links tumorigenesis via FOXA2 suppression. Mol Cell.
45:171–184. 2012. View Article : Google Scholar :
|
|
108
|
Nüsslein-Volhard C and Wieschaus E:
Mutations affecting segment number and polarity in Drosophila.
Nature. 287:795–801. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Pepinsky RB, Zeng C, Wen D, Rayhorn P,
Baker DP, Williams KP, Bixler SA, Ambrose CM, Garber EA, Miatkowski
K, et al: Identification of a palmitic acid-modified form of human
Sonic hedgehog. J Biol Chem. 273:14037–14045. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Porter JA, Young KE and Beachy PA:
Cholesterol modification of hedgehog signaling proteins in animal
development. Science. 274:255–259. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Marigo V, Davey RA, Zuo Y, Cunningham JM
and Tabin CJ: Biochemical evidence that patched is the Hedgehog
receptor. Nature. 384:176–179. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Stone DM, Hynes M, Armanini M, Swanson TA,
Gu Q, Johnson RL, Scott MP, Pennica D, Goddard A, Phillips H, et
al: The tumour-suppressor gene patched encodes a candidate receptor
for Sonic hedgehog. Nature. 384:129–134. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Taipale J, Cooper MK, Maiti T and Beachy
PA: Patched acts catalytically to suppress the activity of
Smoothened. Nature. 418:892–897. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Sicklick JK, Li YX, Jayaraman A, Kannangai
R, Qi Y, Vivekanandan P, Ludlow JW, Owzar K, Chen W, Torbenson MS,
et al: Dysregulation of the Hedgehog pathway in human
hepato-carcinogenesis. Carcinogenesis. 27:748–757. 2006. View Article : Google Scholar
|
|
115
|
Patil MA, Zhang J, Ho C, Cheung ST, Fan ST
and Chen X: Hedgehog signaling in human hepatocellular carcinoma.
Cancer Biol Ther. 5:111–117. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Zheng X, Zeng W, Gai X, Xu Q, Li C, Liang
Z, Tuo H and Liu Q: Role of the Hedgehog pathway in hepatocellular
carcinoma (Review). Oncol Rep. 30:2020–2026. 2013.PubMed/NCBI
|
|
117
|
Koyabu Y, Nakata K, Mizugishi K, Aruga J
and Mikoshiba K: Physical and functional interactions between Zic
and Gli proteins. J Biol Chem. 276:6889–6892. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Wang YY, Jiang JX, Ma H, Han J, Sun ZY,
Liu ZM and Xu ZG: Role of ZIC1 methylation in hepatocellular
carcinoma and its clinical significance. Tumour Biol. 35:7429–7433.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Badouel C, Garg A and McNeill H: Herding
Hippos: Regulating growth in flies and man. Curr Opin Cell Biol.
21:837–843. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Dong J, Feldmann G, Huang J, Wu S, Zhang
N, Comerford SA, Gayyed MF, Anders RA, Maitra A and Pan D:
Elucidation of a universal size-control mechanism in Drosophila and
mammals. Cell. 130:1120–1133. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Zhou D, Conrad C, Xia F, Park JS, Payer B,
Yin Y, Lauwers GY, Thasler W, Lee JT, Avruch J, et al: Mst1 and
Mst2 maintain hepatocyte quiescence and suppress hepatocellular
carcinoma development through inactivation of the Yap1 oncogene.
Cancer Cell. 16:425–438. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Aragón E, Goerner N, Xi Q, Gomes T, Gao S,
Massagué J and Macias MJ: Structural basis for the versatile
interactions of Smad7 with regulator WW domains in TGF-β pathways.
Structure. 20:1726–1736. 2012. View Article : Google Scholar
|
|
123
|
Strano S, Munarriz E, Rossi M, Castagnoli
L, Shaul Y, Sacchi A, Oren M, Sudol M, Cesareni G and Blandino G:
Physical interaction with Yes-associated protein enhances p73
transcriptional activity. J Biol Chem. 276:15164–15173. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Komuro A, Nagai M, Navin NE and Sudol M:
WW domain-containing protein YAP associates with ErbB-4 and acts as
a co-transcriptional activator for the carboxyl-terminal fragment
of ErbB-4 that translocates to the nucleus. J Biol Chem.
278:33334–33341. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Yagi R, Chen LF, Shigesada K, Murakami Y
and Ito Y: A WW domain-containing yes-associated protein (YAP) is a
novel transcriptional co-activator. EMBO J. 18:2551–2562. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Wang W, Huang J and Chen J:
Angiomotin-like proteins associate with and negatively regulate
YAP1. J Biol Chem. 286:4364–4370. 2011. View Article : Google Scholar :
|
|
127
|
Chan SW, Lim CJ, Chong YF, Pobbati AV,
Huang C and Hong W: Hippo pathway-independent restriction of TAZ
and YAP by angiomotin. J Biol Chem. 286:7018–7026. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Zhao B, Li L, Lu Q, Wang LH, Liu CY, Lei Q
and Guan KL: Angiomotin is a novel Hippo pathway component that
inhibits YAP oncoprotein. Genes Dev. 25:51–63. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Oka T, Remue E, Meerschaert K, Vanloo B,
Boucherie C, Gfeller D, Bader GD, Sidhu SS, Vandekerckhove J,
Gettemans J, et al: Functional complexes between YAP2 and ZO-2 are
PDZ domain-dependent, and regulate YAP2 nuclear localization and
signalling. Biochem J. 432:461–472. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Oka T, Mazack V and Sudol M: Mst2 and Lats
kinases regulate apoptotic function of Yes kinase-associated
protein (YAP). J Biol Chem. 283:27534–27546. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Zhao B, Wei X, Li W, Udan RS, Yang Q, Kim
J, Xie J, Ikenoue T, Yu J, Li L, et al: Inactivation of YAP
oncoprotein by the Hippo pathway is involved in cell contact
inhibition and tissue growth control. Genes Dev. 21:2747–2761.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Li H, Wolfe A, Septer S, Edwards G, Zhong
X, Abdulkarim AB, Ranganathan S and Apte U: Deregulation of Hippo
kinase signalling in human hepatic malignancies. Liver Int.
32:38–47. 2012. View Article : Google Scholar
|
|
133
|
Vassilev A, Kaneko KJ, Shu H, Zhao Y and
DePamphilis ML: TEAD/TEF transcription factors utilize the
activation domain of YAP65, a Src/Yes-associated protein localized
in the cytoplasm. Genes Dev. 15:1229–1241. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Zhao B, Kim J, Ye X, Lai ZC and Guan KL:
Both TEAD-binding and WW domains are required for the growth
stimulation and oncogenic transformation activity of yes-associated
protein. Cancer Res. 69:1089–1098. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Pobbati AV and Hong W: Emerging roles of
TEAD transcription factors and its coactivators in cancers. Cancer
Biol Ther. 14:390–398. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Rao VS, Srinivas K, Sujini GN and Kumar
GN: Protein-protein interaction detection: Methods and analysis.
Int J Proteomics. 2014(147648)2014. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
London AS, Patel K, Quinn L and Lemmerer
M: Application of coupled affinity-sizing chromatography for the
detection of proteolyzed HSA-tagged proteins. Protein Expr Purif.
180:80–84. 2014.
|
|
138
|
Tong AH, Evangelista M, Parsons AB, Xu H,
Bader GD, Pagé N, Robinson M, Raghibizadeh S, Hogue CW, Bussey H,
et al: Systematic genetic analysis with ordered arrays of yeast
deletion mutants. Science. 294:2364–2368. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Song Z, Dong C, Wang L, Chen DE, Bi G, Dai
M and Liu J: A novel method for purifying bluetongue virus with
high purity by co-immunoprecipitation with agarose protein A. Virol
J. 7(126)2010.
|
|
140
|
Rigaut G, Shevchenko A, Rutz B, Wilm M,
Mann M and Séraphin B: A generic protein purification method for
protein complex characterization and proteome exploration. Nat
Biotechnol. 17:1030–1032. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
141
|
MacBeath G and Schreiber SL: Printing
proteins as microarrays for high-throughput function determination.
Science. 289:1760–1763. 2000.PubMed/NCBI
|
|
142
|
Westwick JK and Michnick SW:
Protein-fragment complementation assays (PCA) in small GTPase
research and drug discovery. Methods Enzymol. 407:388–401. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Palmer AG III: Enzyme dynamics from NMR
spectroscopy. Acc Chem Res. 48:457–465. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Vidal M and Fields S: The yeast two-hybrid
assay: Still finding connections after 25 years. Nat Methods.
11:1203–1206. 2014. View Article : Google Scholar
|
|
145
|
Güell O, Sagués F and Serrano MA:
Essential plasticity and redundancy of metabolism unveiled by
synthetic lethality analysis. PLOS Comput Biol. 10:e10036372014.
View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Zhang Y, Jin Q, Wang S and Ren R: Modeling
and prediction of peptide drift times in ion mobility spectrometry
using sequence-based and structure-based approaches. Comput Biol
Med. 41:272–277. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Vyas VK, Goel A, Ghate M and Patel P:
Ligand and structure-based approaches for the identification of
SIRT1 activators. Chem Biol Interact. 228:9–17. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Enright AJ, Iliopoulos I, Kyrpides NC and
Ouzounis CA: Protein interaction maps for complete genomes based on
gene fusion events. Nature. 402:86–90. 1999.PubMed/NCBI
|
|
149
|
Foster HA, Estrada-Girona G, Themis M,
Garimberti E, Hill MA, Bridger JM and Anderson RM: Relative
proximity of chromosome territories influences chromosome exchange
partners in radiation-induced chromosome rearrangements in primary
human bronchial epithelial cells. Mutat Res. 756:66–77. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Pazos F and Valencia A: In silico
two-hybrid system for the selection of physically interacting
protein pairs. Proteins. 47:219–227. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Whidden C and Matsen FA IV: Quantifying
MCMC exploration of phylogenetic tree space. Syst Biol. 64:472–491.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Altman J, Hédl R, Szabó P, Mazůrek P,
Riedl V, Müllerová J, Kopecký M and Doležal J: Tree-rings mirror
management legacy: Dramatic response of standard oaks to past
coppicing in Central Europe. PLoS One. 8:e557702013. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Gene Ontology, C; Gene and Ontology
Consortium: Gene Ontology Consortium: Going forward. Nucleic Acids
Res. 43(D1): D1049–D1056. 2015. View Article : Google Scholar
|
|
154
|
Xenarios I, Salwínski L, Duan XJ, Higney
P, Kim SM and Eisenberg D: DIP, the Database of Interacting
Proteins: A research tool for studying cellular networks of protein
interactions. Nucleic Acids Res. 30:303–305. 2002. View Article : Google Scholar :
|
|
155
|
Chatr-Aryamontri A, Breitkreutz BJ,
Oughtred R, Boucher L, Heinicke S, Chen D, Stark C, Breitkreutz A,
Kolas N, O'Donnell L, et al: The BioGRID interaction database: 2015
update. Nucleic Acids Res. 43(D1): D470–D478. 2015. View Article : Google Scholar
|
|
156
|
Cowley MJ, Pinese M, Kassahn KS, Waddell
N, Pearson JV, Grimmond SM, Biankin AV, Hautaniemi S and Wu J: PINA
v2.0: Mining interactome modules. Nucleic Acids Res. 40(D1):
D862–D865. 2012. View Article : Google Scholar :
|
|
157
|
Patil A, Nakai K and Nakamura H:
HitPredict: A database of quality assessed protein-protein
interactions in nine species. Nucleic Acids Res. 39(Database):
D744–D749. 2011. View Article : Google Scholar
|
|
158
|
Hermjakob H, Montecchi-Palazzi L,
Lewington C, Mudali S, Kerrien S, Orchard S, Vingron M, Roechert B,
Roepstorff P, Valencia A, et al: IntAct: An open source molecular
interaction database. Nucleic Acids Res. 32:D452–D455. 2004.
View Article : Google Scholar :
|
|
159
|
Prieto C and De Las Rivas J: APID: Agile
Protein Interaction DataAnalyzer. Nucleic Acids Res. 34(Web
Server): W298–W302. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Licata L, Briganti L, Peluso D, Perfetto
L, Iannuccelli M, Galeota E, Sacco F, Palma A, Nardozza AP,
Santonico E, et al: MINT, the molecular interaction database: 2012
update. Nucleic Acids Res. 40(D1): D857–D861. 2012. View Article : Google Scholar :
|
|
161
|
Song JJ and Lee YJ: Dissociation of Akt1
from its negative regulator JIP1 is mediated through the
ASK1-MEK-JNK signal transduction pathway during metabolic oxidative
stress: A negative feedback loop. J Cell Biol. 170:61–72. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Kim J, Kang D, Sun BK, Kim JH and Song JJ:
TRAIL/MEKK4/p38/HSP27/Akt survival network is biphasically
modulated by the Src/CIN85/c-Cbl complex. Cell Signal. 25:372–379.
2013. View Article : Google Scholar
|
|
163
|
Deregibus MC, Cantaluppi V, Doublier S,
Brizzi MF, Deambrosis I, Albini A and Camussi G: HIV-1-Tat protein
activates phosphatidylinositol 3-kinase/AKT-dependent survival
pathways in Kaposi's sarcoma cells. J Biol Chem. 277:25195–25202.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Polzien L, Baljuls A, Rennefahrt UE,
Fischer A, Schmitz W, Zahedi RP, Sickmann A, Metz R, Albert S, Benz
R, et al: Identification of novel in vivo phosphorylation sites of
the human proapoptotic protein BAD: Pore-forming activity of BAD is
regulated by phosphorylation. J Biol Chem. 284:28004–28020. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Xiang T, Ohashi A, Huang Y, Pandita TK,
Ludwig T, Powell SN and Yang Q: Negative regulation of AKT
activation by BRCA1. Cancer Res. 68:10040–10044. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Xiang T, Jia Y, Sherris D, Li S, Wang H,
Lu D and Yang Q: Targeting the Akt/mTOR pathway in Brca1-deficient
cancers. Oncogene. 30:2443–2450. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Du K and Montminy M: CREB is a regulatory
target for the protein kinase Akt/PKB. J Biol Chem.
273:32377–32379. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Matsuzaki H, Daitoku H, Hatta M, Aoyama H,
Yoshimochi K and Fukamizu A: Acetylation of Foxo1 alters its
DNA-binding ability and sensitivity to phosphorylation. Proc Natl
Acad Sci USA. 102:11278–11283. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Brent MM, Anand R and Marmorstein R:
Structural basis for DNA recognition by FoxO1 and its regulation by
posttranslational modification. Structure. 16:1407–1416. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Biggs WH III, Meisenhelder J, Hunter T,
Cavenee WK and Arden KC: Protein kinase B/Akt-mediated
phosphorylation promotes nuclear exclusion of the winged helix
transcription factor FKHR1. Proc Natl Acad Sci USA. 96:7421–7426.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Yang H, Zhao R, Yang HY and Lee MH:
Constitutively active FOXO4 inhibits Akt activity, regulates p27
Kip1 stability, and suppresses HER2-mediated tumorigenicity.
Oncogene. 24:1924–1935. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Matsuzaki H, Ichino A, Hayashi T, Yamamoto
T and Kikkawa U: Regulation of intracellular localization and
transcriptional activity of FOXO4 by protein kinase B through
phosphorylation at the motif sites conserved among the FOXO family.
J Biochem. 138:485–491. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Zhu QS, Rosenblatt K, Huang KL, Lahat G,
Brobey R, Bolshakov S, Nguyen T, Ding Z, Belousov R, Bill K, et al:
Vimentin is a novel AKT1 target mediating motility and invasion.
Oncogene. 30:457–470. 2011. View Article : Google Scholar
|
|
174
|
Drendall CI, Pham QH and Dietze EC:
Purification and characterization of recombinant CH3 domain
fragment of the CREB-binding protein. Protein Expr Purif.
70:196–205. 2010. View Article : Google Scholar :
|
|
175
|
Connor MK, Azmi PB, Subramaniam V, Li H
and Seth A: Molecular characterization of ring finger protein 11.
Mol Cancer Res. 3:453–461. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Yang WL, Wang J, Chan CH, Lee SW, Campos
AD, Lamothe B, Hur L, Grabiner BC, Lin X, Darnay BG, et al: The E3
ligase TRAF6 regulates Akt ubiquitination and activation. Science.
325:1134–1138. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Hartman AD, Wilson-Weekes A, Suvannasankha
A, Burgess GS, Phillips CA, Hincher KJ, Cripe LD and Boswell HS:
Constitutive c-jun N-terminal kinase activity in acute myeloid
leukemia derives from Flt3 and affects survival and proliferation.
Exp Hematol. 34:1360–1376. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Kim CK, Lee SB, Nguyen TL, Lee KH, Um SH,
Kim J and Ahn JY: Long isoform of ErbB3 binding protein, p48,
mediates protein kinase B/Akt-dependent HDM2 stabilization and
nuclear localization. Exp Cell Res. 318:136–143. 2012. View Article : Google Scholar
|
|
179
|
Deep G, Oberlies NH, Kroll DJ and Agarwal
R: Isosilybin B causes androgen receptor degradation in human
prostate carcinoma cells via PI3K-Akt-Mdm2-mediated pathway.
Oncogene. 27:3986–3998. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Milne D, Kampanis P, Nicol S, Dias S,
Campbell DG, Fuller-Pace F and Meek D: A novel site of AKT-mediated
phosphorylation in the human MDM2 oncoprotein. FEBS Lett.
577:270–276. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Facchinetti V, Ouyang W, Wei H, Soto N,
Lazorchak A, Gould C, Lowry C, Newton AC, Mao Y, Miao RQ, et al:
The mammalian target of rapamycin complex 2 controls folding and
stability of Akt and protein kinase C. EMBO J. 27:1932–1943. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Oh WJ, Wu CC, Kim SJ, Facchinetti V,
Julien LA, Finlan M, Roux PP, Su B and Jacinto E: mTORC2 can
associate with ribosomes to promote cotranslational phosphorylation
and stability of nascent Akt polypeptide. EMBO J. 29:3939–3951.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Glidden EJ, Gray LG, Vemuru S, Li D,
Harris TE and Mayo MW: Multiple site acetylation of Rictor
stimulates mammalian target of rapamycin complex 2
(mTORC2)-dependent phosphorylation of Akt protein. J Biol Chem.
287:581–588. 2012. View Article : Google Scholar :
|
|
184
|
Fan CD, Lum MA, Xu C, Black JD and Wang X:
Ubiquitin-dependent regulation of phospho-AKT dynamics by the
ubiquitin E3 ligase, NEDD4-1, in the insulin-like growth factor-1
response. J Biol Chem. 288:1674–1684. 2013. View Article : Google Scholar :
|
|
185
|
Persaud A, Alberts P, Amsen EM, Xiong X,
Wasmuth J, Saadon Z, Fladd C, Parkinson J and Rotin D: Comparison
of substrate specificity of the ubiquitin ligases Nedd4 and Nedd4-2
using proteome arrays. Mol Syst Biol. 5(333)2009. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Mistafa O, Ghalali A, Kadekar S, Högberg J
and Stenius U: Purinergic receptor-mediated rapid depletion of
nuclear phosphorylated Akt depends on pleckstrin homology domain
leucine-rich repeat phosphatase, calcineurin, protein phosphatase
2A, and PTEN phosphatases. J Biol Chem. 285:27900–27910. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Woods NT, Mesquita RD, Sweet M, Carvalho
MA, Li X, Liu Y, Nguyen H, Thomas CE, Iversen ES Jr, Marsillac S,
et al: Charting the landscape of tandem BRCT domain-mediated
protein interactions. Sci Signal. 5:rs62012. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Sarbassov DD, Ali SM, Sengupta S, Sheen
JH, Hsu PP, Bagley AF, Markhard AL and Sabatini DM: Prolonged
rapamycin treatment inhibits mTORC2 assembly and Akt/PKB. Mol Cell.
22:159–168. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Ikenoue T, Inoki K, Yang Q, Zhou X and
Guan KL: Essential function of TORC2 in PKC and Akt turn motif
phosphorylation, maturation and signalling. EMBO J. 27:1919–1931.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Chen CH, Shaikenov T, Peterson TR,
Aimbetov R, Bissenbaev AK, Lee SW, Wu J, Lin HK and Sarbassov D: ER
stress inhibits mTORC2 and Akt signaling through GSK-3β-mediated
phosphorylation of rictor. Sci Signal. 4:ra102011. View Article : Google Scholar
|
|
191
|
Sundaresan NR, Pillai VB, Wolfgeher D,
Samant S, Vasudevan P, Parekh V, Raghuraman H, Cunningham JM, Gupta
M and Gupta MP: The deacetylase SIRT1 promotes membrane
localization and activation of Akt and PDK1 during tumorigenesis
and cardiac hypertrophy. Sci Signal. 4:ra462011. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Gao D, Inuzuka H, Tseng A, Chin RY, Toker
A and Wei W: Phosphorylation by Akt1 promotes cytoplasmic
localization of Skp2 and impairs APCCdh1-mediated Skp2 destruction.
Nat Cell Biol. 11:397–408. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Romano D, Matallanas D, Weitsman G,
Preisinger C, Ng T and Kolch W: Proapoptotic kinase MST2
coordinates signaling crosstalk between RASSF1A, Raf-1, and Akt.
Cancer Res. 70:1195–1203. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Kim D, Shu S, Coppola MD, Kaneko S, Yuan
ZQ and Cheng JQ: Regulation of proapoptotic mammalian ste20-like
kinase MST2 by the IGF1-Akt pathway. PLoS One. 5:e96162010.
View Article : Google Scholar : PubMed/NCBI
|