Introduction
Types of primary liver cancer include hepatocellular
carcinoma (HCC), intrahepatic cholangiocarcinoma (ICC) and hepatic
angiosarcoma. HCC accounts for between 85 and 90% of all cases of
primary liver cancer and is the third leading cause of mortality
from cancer worldwide and the fifth most common type of malignancy
(1–4). It has been reported that ~21,000
cases of HCC are diagnosed and ~700,000 patients succumb to the
disease worldwide annually (5).
The established risk factors of HCC include viral hepatitis,
alcohol abuse and non-alcoholic fatty liver disease (6). For the majority of patients with HCC,
surgical resection and liver transplantation are the only effective
treatment options; however, only between 10 and 20% of patients are
eligible for surgical intervention, due to the difficulty in
effectively diagnosing HCC in the early stages of the disease
(7). Frequent tumor metastasis and
recurrence following surgical intervention lead to poor prognoses
of patients with HCC with a five-year survival rate of ~5%
(8). Advances in functional
genomics have provided a deeper understanding of
hepatocarcinogenesis; however, the molecular pathogenesis of HCC
remains to be fully elucidated (9,10).
In addition, the clinical heterogeneity of HCC and insufficient
effective diagnostic markers and therapeutic approaches for the
treatment of HCC have rendered this disease a major challenge
(11). Therefore, in order to
improve the prognosis of patients with HCC, novel diagnostic
markers and treatments are required.
MicroRNAs (miRNAs) are a type of small, endogenous,
single-stranded, non-coding RNA, which consist of between 20 and 25
bases (12). miRNA was first
detected during a developmental timing experiment, which was
performed in the nematode Caenorhabditis elegans in 1993
(13). To date, the family of
human miRNAs consists of >2,000 mature miRNAs and, using in
silico techniques, it was predicted that ~60% of human
messenger (m)RNA may be targets of miRNAs (14). In addition, miRNAs abberrantly
expressed in several types of human cancer may function as
oncogenes and tumor suppressors (15). Based upon a large number of
experimental studies performed over the past two decades, it has
been confirmed that miRNAs are important in the regulation of gene
expression, which primarily occurs through post-transcriptional
destabilization, translational repression of target mRNAs which
bear complementary sites or a combination of these two mechanisms
(16–19).
Previous studies have demonstrated that miRNAs are
essential in the biology of various types of human cancer,
including cell differentiation, proliferation, apoptosis, invasion
and angiogenesis (20,21). It was reported that ≥17 miRNAs are
downregulated in HCC, while six miRNAs are upregulated(22). miRNAs which are upregulated in
cancer may function as oncogenes through the negative regulation of
tumor suppressors. By contrast, miRNAs which are downregulated in
cancer may normally function as tumor suppressor genes, which
inhibit cancer through the regulation of oncogenes (23). Therefore, the identification of
miRNA targets is critical in order to fully elucidate the function
of miRNAs in the development and progression of cancer. In
addition, it has been suggested that miRNAs may be potential target
for the treatment of cancer (24).
It has been reported that miRNA (miR)-133a is among
the most frequently downregulated miRNAs in various types of human
malignancy, including HCC (25),
renal cell carcinoma (26),
esophageal squamous cell carcinoma (27), bladder cancer (28), ileal carcinoid (29) and rhabdomyosarcoma (30). The aim of the present study was to
determine the effects of miR-133a on cell proliferation, colony
formation, migration and invasion in HCC HepG2 and SMMC-7721 cell
lines, as well as to investigate whether matrix metallopeptidase 9
(MMP-9) may be a target of miR-133a.
Materials and methods
Cells and culture conditions
The human HCC cell lines, HepG2 and SMMC-7721, were
obtained from the Shanghai Institute Of Biochemistry And Cell
Biology (Shanghai, China). The HepG2 and SMMC-7721 cells were
cultured in RPMI 1640 medium (Gibco, Grand Island, NY, USA)
supplemented with 10% heat-inactivated fetal bovine serum (FBS;
Gibco) under a humidified atmosphere of 5% CO2 at 37°C.
To propagate sphere formation in vitro, spheres were
collected through gentle centrifugation at 200 × g for 5 min,
dissociated to single cells by blowing gently and cultured in
RPMI-1640 to produce the next generation of spheres.
Transfection
Mature miR-133a mimics and negative control (NC)
miRNA mimics were designed and synthesized by GenePharma (Shanghai,
China). The sequences were as follows: miR-133a mimic,
5′-UUUGGUCCCCUUCAACCAGCUG-3′ and NC mimic,
5′-UUCUCCGAACGUGUCACGUTT-3′. The cells were transfected using
Lipofectamine 2000 (Invitrogen Life Technologies, Carlsbad, CA,
USA), according to manufacturer’s instructions.
Quantitative detection of miR-133a
Total RNA was extracted from the cells using TRIzol
reagent (Invitrogen Life Technologies) in a one-step extraction
procedure, as previously described (31). RNA was stored in
diethylpyrocarbonate-treated water at −80°C and the quantity and
quality of samples were evaluated using a ND-1000 NanoDrop
spectrophotometer (NanoDrop, Wilmington, DE, USA). Reverse
transcription quantitative polymerase chain reaction (RT-qPCR) for
miR-133a was performed using a SYBR Green miRNA assay (GenePharma),
according to the manufacturer’s instructions. qPCR was performed on
an AB7300 thermo-recycler (Applied Biosystems, Waltham, MA, USA)
using miR-133 primer set (Tiangen Biotech, Co., Ltd, Beijing,
China) and double strand binding dye SYBR Green. GAPDH was used as
an internal control. Each sample was replicated three times, with
no RT or template control included. Data were analyzed by comparing
Ct values (32).
3-(4, 5-dimethylthiazol-2-yl)-2,5-
diphenyltetrazolium bromide (MTT) assay
Cell proliferation was determined using a MTT Cell
Proliferation and Cytotoxicity Assay kit (Beyotime Institute of
Biotechnology, Shanghai, China). The transfected cells (miR-133a
mimics and NC) were seeded into 96-well flat-bottomed plates
(Becton-Dickinson, Heidelberg, Germany) at a density of 3,000 cells
per well. Every 24 h for 5 days, the viable cells were assayed for
their ability to transform MTT into purple formazan, and their
optical density was measured at 490 nm (OD)490. The
suppression rate was calculated using the following formula:
Suppression rate = (1−ODmiR-145/ODmiR-NC) ×
100%. Proliferation curves were drawn on the basis of the mean
absorbance at each time-point. All experiments were performed in
triplicate.
Colony formation assay
The colony formation ability of the
miR-133a-transfected HepG2 and SMMC-7721 cells was assessed using a
colony formation assay. In brief, the transfected cells (miR-133a
mimics and NC) growing in the logarithmic phase were trypsinized
with 0.05% Trypsin-EDTA (Gibco) and seeded into six-well plates at
a density of 2,000 cells per well. The cells were maintained in an
incubator at 37°C for 7 days. On day 8, the colonies were washed
with phosphate-buffered saline (PBS), fixed with formalin (10%),
and stained with methyl violet, which were all purchased from
Beyotime Institute of Biotechnology. The methyl violet dye was then
washed with PBS and the number of colonies were counted under a
microscope (IX53; Olympus Corp., Tokyo, Japan). The following
calculations were then performed: Colony-inhibition rate = [(1 −
number of colonies in experimental groups)/control group] × 100%;
and colony-forming efficiency = 1 − colony-inhibition rate.
Cell migration and invasion assay
The cell migration and invasion were assayed using
Transwell® chambers (8 μm; Corning Costar, Cambridge,
MA, USA). For the Transwell® migration assay,
1×105 transfected cells (miR 133a mimics and NC) were
placed into the upper chamber, which was cultured in medium with 2%
FBS, while 500 μl RPMI-1640 medium containing 20% FBS was added to
the lower chamber. For the Transwell® invasion assay, a
Transwell® chamber coated with Matrigel® (BD
Biosciences, San Jose, CA, USA) and a total of 1×105
cells were seeded into the upper chamber, while the lower chamber
was incubated with 500 μl RPMI-1640 medium containing 20% FBS. The
cells were incubated under a humidified atmosphere of 5%
CO2 at 37°C for 12 h for the migration assay and 24 h
for the invasion assay. Subsequently, cells remaining in the upper
chambers or on the upper membrane of the inserts were carefully
removed with cotton swabs. Following fixation and staining in a dye
solution containing 0.5% crystal violet (Beyotime Institute of
Biotechnology) and 20% methanol (Macklin Biochemical Co., Ltd,
Shanghai, China), the cells adhering to the lower membrane of the
inserts were counted and imaged with microscopy (magnification,
×200). Five five fields of vision for each insert were randomly
selected and counted under a light microscope (IX53; Olympus
Corp.). Each condition was assayed in triplicate and each
experiment was repeated a minimum of three times.
Target of miR-133b
In order to determine whether miR-133a targets the
MMP-9 3′-UTR, TARGETSCAN 5.2 (http://www.targetscan.org/) and PICTAR (http://pictar.mdc-berlin.de/) were used.
Western blot analysis
The transfected cells (miR-133a mimics and NC) were
washed with ice-cold PBS and lysed with 1% radioimmunoprecipitation
assay lysis buffer (Beyotime Institute of Biotechnology) 72 h after
transfection. The supernatants were collected and the protein
concentrations were determined using a Bicinchoninic Acid Assay kit
(Beyotime Institute of Biotechnology). Equal quantities of the
proteins were separated and analyzed using 10% SDS-PAGE (Beyotime
Institute of Biotechnology) and then transferred onto
polyvinylidene difluoride membranes (Beyotime Institute of
Biotechnology). The membranes were then blocked with 5% skimmed
milk (Shyuanmu, Shanghai, China), followed by incubation overnight
at 4°C with a primary rabbit anti-human polyclonal MMP-9 antibody
(1:1,000; Bioworld Technology, Inc., St. Louis Park, MN, USA),
according to the manufacturer’s instructions. The membranes were
then washed three times with Tris-buffered saline with 1% Tween 20
(TBST; Beyotime) and then incubated at room temperature with the
corresponding horseradish peroxidase-conjugated goat anti-rabbit
secondary antibody (1:1,000) in TBST. Western blots were developed
using enhanced chemilluminescence solution (Pierce Biotechnology,
Inc., Rockford, IL, USA) and images were captured using a FluorChem
imaging system (Alpha Innotech Corp., San Leandro, CA, USA). The
data were normalized to β-actin.
Luciferase assay
The HepG2 and SMMC-7721 cells were transfected with
0.5 μg reporter plasmid, 40 nmol miR-133a mimics or NC in a 12-well
plate using Lipofectamine 2000, according to manufacturer’s
instructions. The assays were performed using the Dual-Luciferase
Reporter Assay system (Promega Corporation, Manheim, Germany) 48 h
after transfection. The activities of Firefly and Renilla
luciferase were measured using a luminometer (Tecan Group, Ltd,
Maennedorf, Switzerland). The firefly luciferase activity was
normalized to that of the Renilla luciferase for each transfected
well. Each reporter plasmid was transfected a minimum of three
times (on different days) and each sample was assayed in
triplicate.
Statistical analysis
Data are presented as the mean ± standard deviation
and were compared using the Student’s t-test using Stata 10.0
software (StataCorp LP, College Station, TX, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of miR-133a prior to and
following transfection of miR-133a mimics in HepG2 and SMMC-7721
cells
The endogenous levels of miR-133a in the HepG2 and
SMMC-7721 cells, and its expression following transfection with
miR-133a, was determined every 24 h. As expected, the basal
expression of miR-133a was too low to be shown in Fig. 1. However, following transfection
with miR-133a, the expression levels of miR-133a were markedly
increased compared with those of the untransfected cells at 24 h,
until 144 h. Of note, the expression of miR-133a decreased in a
time-dependent manner from 24 h.
miR-133a reduces the proliferation and
colony formation abilities of HepG2 and SMMC-7721 cells
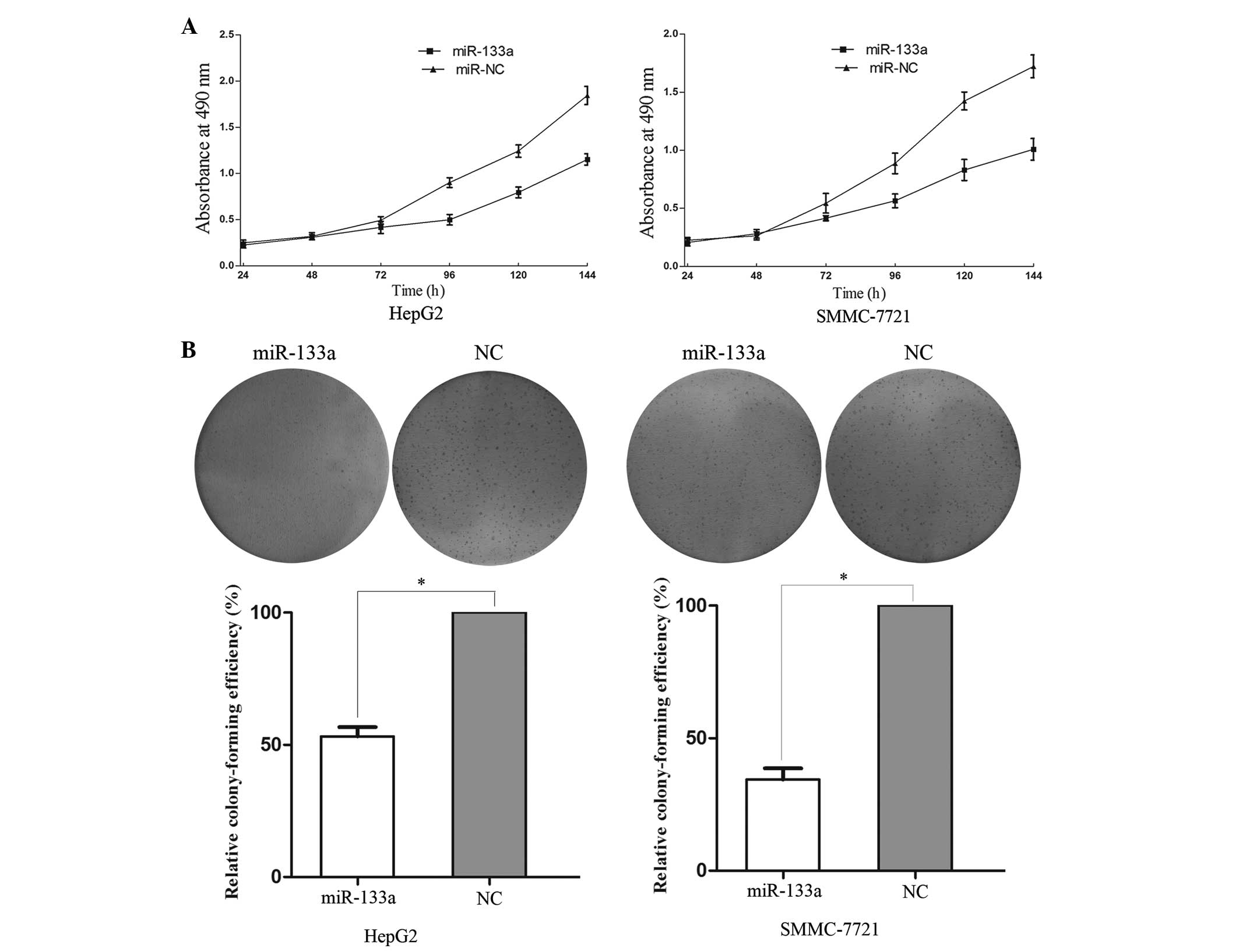
In order to investigate the effect of miR-133a on
cell proliferation, an MTT assay was performed. The results
demonstrated that upregulation of miR-133a significantly inhibited
cell proliferation compared with the NC-transfected cells (Fig. 2A). In addition, the MTT assays
revealed that after 144 h of treatment, the suppression rate of
miR-133a reached 35.80±3.2% in the HepG2 cells and 41.15±3.6% in
the SMMC-7721 cells.
In order to investigate the effect of miR-133a on
colony formation in theHCC cells, a colony formation assay was
performed. As shown in Fig. 2B,
the relative colony-formation efficiency of the
miR-133a-transfected cells was 53.2±4.5% in the HepG2 cells and
34.4±5.9% in the SMMC-7721 cells, compared with the cells
transfected with the NC (P<0.05). Overall, these results
indicated that miR-133a may be important in the proliferation and
colony formating ability of HCC HepG2 and SMMC-7721 cells.
miR-133a suppresses cell migration and
invasion in HCC HepG2 and SMMC-7721 cells
In order to measure the effect of miR-133a on tumor
cell migration and invasion, a Transwell® apparatus
assay was performed. The results of the migration assay
demonstrated that migration was significantly decreased in the
miR-133a-transfected groups to 48.25±5.39% in the HepG2 cells and
58.46±6.21% in the SMMC-7721 cells compared with those of the
NC-transfected groups (P<0.05; Fig.
3A). In the invasion assay (Fig.
3B), miR-133a-transfection induced a 58.35±7.89% decrease in
the number of invasive HepG2 cells and a 63.12±6.52% decrease in
the number of invasive SMMC-7721 cells compared with the
NC-transfected cells (P<0.05). These results indicated that the
overexpression of miR-133a reduced the migration and invasion
abilities of the HCC cell lines.
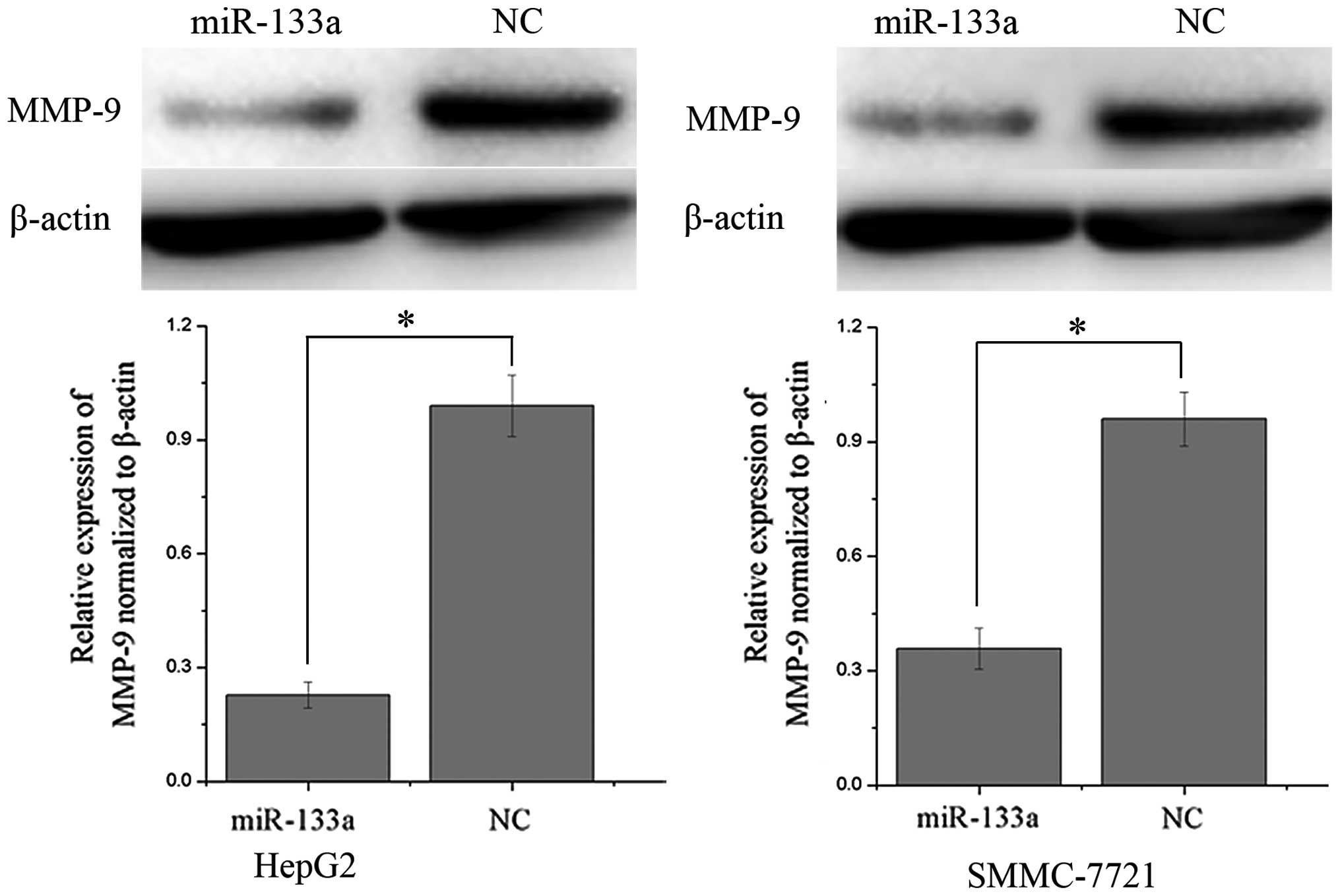
MMP-9 is downregulated following
overexpression of miR-133a in HCC cells
Western blot analysis was performed in order to
determine whether the protein expression of MMP-9 was altered
following transfection with miR-133a mimics in the HCC HepG2 and
SMMC-7721 cell lines. As shown in Fig.
4, MMP-9 was significantly downregulated in the HCC HepG2 and
SMMC-7721 cell lines following overexpression of miR-133a compared
with the NC-transfected cells (P<0.05). These results indicated
that miR-133a may reduce the protein level of MMP-9 in HCC
cells.
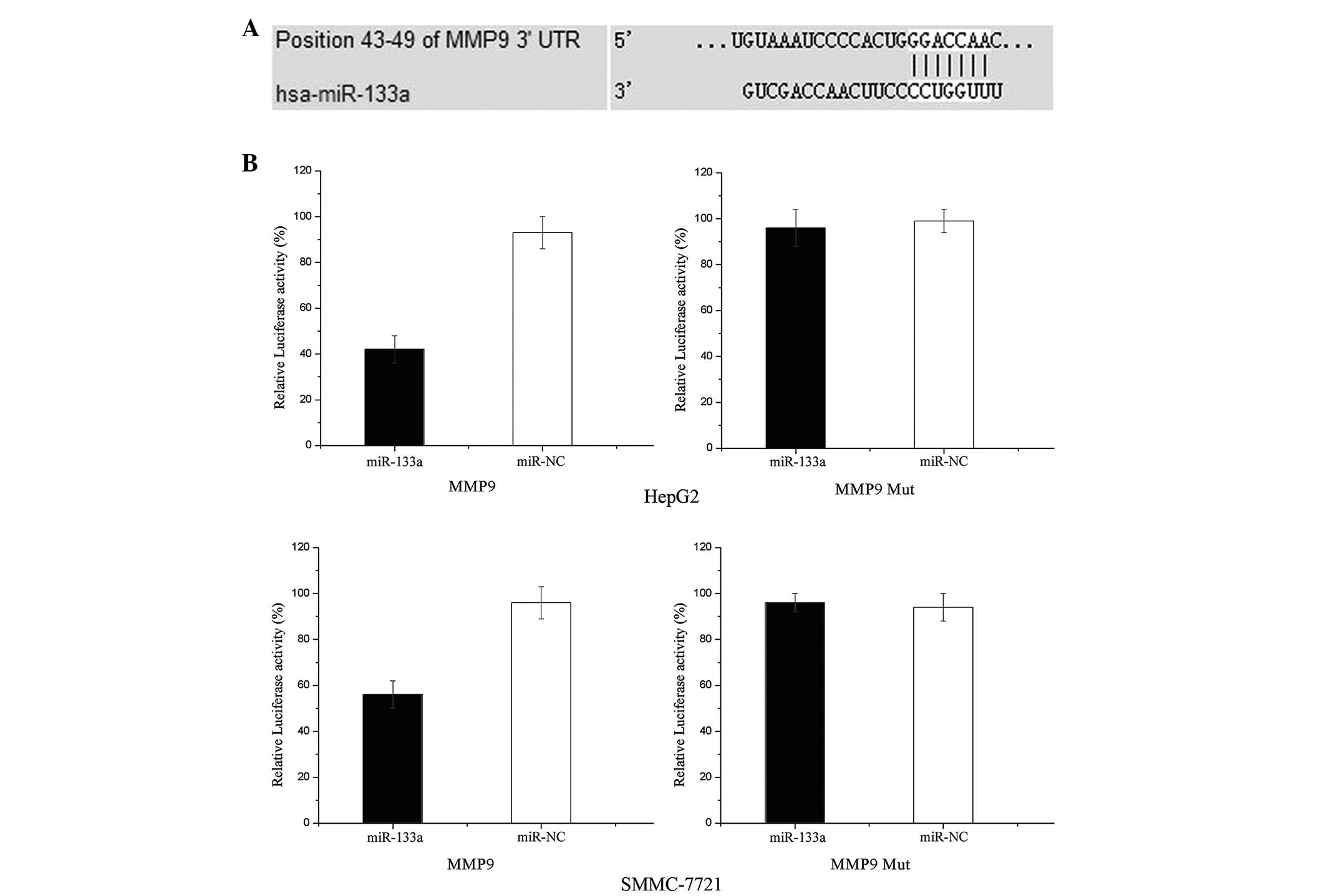
MMP-9 is a direct target gene of miR-133a
in HCC
To determine whether miR-133a targets the MMP-9
3′-untranslated region (UTR), TARGETSCAN 5.2 and PICTAR were used
to assess the complementarity of miR-133a to the MMP-9 3′-UTR. It
was demonstrated that MMP-9 mRNA contained an miR-133a
seven-nucleotide seed match at position 43–49 of the MMP-9 3′-UTR
(Fig. 5A).
Luciferase reporter assays were performed to
evaluate whether MMP-9 was a target of miR-133a in HCC cells. As
shown in Fig. 5B, overexpression
of miR-133a suppressed the activity of MMP-9 3′-UTR-luciferase by
55% in the HepG2 cells and 42% in the SMMC-7721 cells compared with
the NC-transfected cells (P<0.05). Overall, these results
indicated that MMP-9 may be a direct target of miR-133a in
vitro.
Discussion
miR-133 is an miRNA family containing miR-133a and
miR-133b; which differ by only one base in the terminal 3′ position
(33). miR-133a is a multicopy
gene, with two copies in chromosomes 18 and 20, which are located
next to another muscle-enriched miRNA, miR-1, while miR-133b is
located in chromosome 6 (34).
miR-133a has been recognized as a muscle-specific miRNA, which may
regulate myoblast differentiation and be involved in myogenic and
heart diseases (30,35,36).
Furthermore, miR-133a has been commonly identified as being
downregulated in various human malignancies, including
hepatocellular carcinoma (25),
renal cell carcinoma (26),
esophageal squamous cell carcinoma (27), bladder cancer (28), ileal carcinoid (29) and rhabdomyosarcoma (30).
Identification of miR-133a target genes is essential
for understanding its role in tumorigenesis and for defining novel
therapeutic targets. Studies have found that miR-133a regulates
oncogenic transcripts in human cells, including FSCN1, EGFR, LASP1,
GSTP1 and TAGLN2 (34,37,38).
Therefore, upregulating miR-133a or providing exogenous analogous
pharmaceutical compounds may provide effective cancer therapies for
HCC, which resulted from the overexpression of these oncogenic
transcripts. The results of the present study revealed multiple
inhibitory effects of miR-133a in the HepG2 and SMMC-7721 HCC cell
lines, including growth arrest, reduced cell colony formation
ability and suppression of migration and invasion, by
downregulating the expression of MMP-9. These findings suggested
that miR-133a may be used for the development of novel molecular
markers and therapeutic approaches for the inhibition of metastasis
in HCC.
The incidence of HCC is increasing in several
countries and is becoming one of the most prevalent types of
terminal cancer worldwide (39).
In addition, HCC is characterized by rapid progression, early
metastasis and frequent recurrence (40). Despite improvements in the
diagnosis and treatment of HCC, however, it remains an aggressive
type of cancer with a poor prognosis. Tissue invasion and
metastasis are the primary cause of mortality in patients with HCC
(40). Defining the mechanisms
regulating HCC invasion may identify novel elements, which may be
exploited therapeutically to reduce metastasis and improve patient
survival. The process of metastasis involves several steps,
including the detachment of cancer cells from the primary tumour,
followed by the migration, adhesion and invasion of cancer cells
into blood or lymphatic vessels; cancer cells then undergo
extravasation and subsequently interact with target tissues, where
they form metastastic foci in distant organs (41). The destruction of the extracellular
matrix (ECM) by enzymes is an essential initial step in the
processes of tumor cell invasion and metastasis, and several
studies have reported that, among the enzymes responsible for ECM
degradation, MMPs are have a critical role (42,43).
MMPs are a family of zinc-dependent endopeptidases,
which have share several structural and functional properties, but
with different substrate specificities (44). MMPs are important in various
physiological processes, which include tissue remodeling, organ
development, angiogenesis, inflammatory processes, vascular and
autoimmune disorders and cancer (45). In addition, MMPs are upregulated in
various types of human malignant tumor. The majority of clinical
data had revealed correlations between the expression of MMPs and
advanced tumor stages, invasion, metastasis and decreased survival
rates (46). Previous studies have
demonstrated that several MMPs are overexpressed in HCC and that
high expression levels of MMPs are associated with cancer
progression and metastasis (47–49).
It was therefore suggested that inhibitors of MMP activity may be
investigated for the prevention or reduction of tumor
metastasis.
A total of 24 soluble and membrane-anchored members
of the MMP family have been identified, which are subdivided into
four families based on structure and substrate specificity as
follows: Collagenases, gelatinases, stromelysins and
membrane-associated MMPs (50).
Among these MMPs, the activities of MMP-2 and -9 have been
associated with the progression of HCC (48,51).
In addition, these studies revealed an association between the
overexpression of MMP-2 or -9 and the invasion and metastasis of
HCC. Increased levels of MMP-9 in HCC also correlate with increased
tumour recurrence and metastasis following resection (48,49,52).
Hayasaka et al (53)
reported that MMP-9 plasma levels were upregulated in HCC patients,
particularly in patients who presented with macroscopic portal vein
invasion, which suggested that MMP-9 may serve as a marker for
transformation and invasion in HCC or as a therapeutic target for
the inhibition of metastasis in HCC. The results of the present
study suggested that miR-133a suppressed HCC cell migration and
invasion via the downregulation of MMP-9, therefore indicating the
predictive value of MMP-9 for early detection of tumor metastasis
and as a target for preventative therapies to inhibit HCCs becoming
invasive.
In conclusion, to the best of our knowledge, the
present study was the first to demonstrate that miR-133a inhibited
HCC cell proliferation, colony formation, migration and invasion by
downregulating the expression of MMP-9. These findings have
therapeutic implications and may be exploited for further treatment
of HCC. Further studies are required to determine whether the
potential of miR-133a may be fully realized in cancer
treatment.
References
|
1
|
Kudo M: Hepatocellular carcinoma in 2011
and beyond: from the pathogenesis to molecular targeted therapy.
Oncology. 81(Suppl 1): 1–10. 2011. View Article : Google Scholar
|
|
2
|
Meguro M, Mizuguchi T, Kawamoto M and
Hirata K: The molecular pathogenesis and clinical implications of
hepatocellular carcinoma. Int J Hepatol. 2011:8186722011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yamazaki K, Masugi Y and Sakamoto M:
Molecular pathogenesis of hepatocellular carcinoma: altering
transforming growth factor-β signaling in hepatocarcinogenesis. Dig
Dis. 29:284–288. 2011. View Article : Google Scholar
|
|
4
|
Rong M, Chen G and Dang Y: Increased
miR-221 expression in hepatocellular carcinoma tissues and its role
in enhancing cell growth and inhibiting apoptosis in vitro. BMC
Cancer. 13:212013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zou C, Li Y, Cao Y, Zhang J, Jiang J,
Sheng Y, Wang S, Huang A and Tang H: Up-regulated MicroRNA-181a
induces carcinogenesis in Hepatitis B virus-related hepatocellular
carcinoma by targeting E2F5. BMC Cancer. 14:972014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Su ZX, Zhao J, Rong ZH, Geng WM, Wu YG and
Qin CK: Upregulation of microRNA-25 associates with prognosis in
hepatocellular carcinoma. Diagn Pathol. 9:472014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang L, Yao J, Shi X, Hu L, Li Z, Song T
and Huang C: MicroRNA-302b suppresses cell proliferation by
targeting EGFR in human hepatocellular carcinoma SMMC-7721 cells.
BMC Cancer. 13:4482013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aravalli RN, Steer CJ and Cressman EN:
Molecular mechanisms of hepatocellular carcinoma. Hepatology.
48:2047–2063. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Thorgeirsson SS, Lee JS and Grisham JW:
Functional genomics of hepatocellular carcinoma. Hepatology. 43(2
Suppl 1): S145–S150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Villanueva A, Newell P, Chiang DY,
Friedman SL and Llovet JM: Genomics and signaling pathways in
hepatocellular carcinoma. Semin Liver Dis. 27:55–76. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Roessler S, Budhu A and Wang XW:
Deciphering cancer heterogeneity: the biological space. Front Cell
Dev Biol. 2:122014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shirasaki T, Honda M, Shimakami T, Horii
R, Yamashita T, Sakai Y, Sakai A, Okada H, Watanabe R, Murakami S,
et al: MicroRNA-27a regulates lipid metabolism and inhibits
hepatitis C virus replication in human hepatoma cells. J Virol.
87:5270–5286. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar :
|
|
15
|
Wu D, Ding J, Wang L, Pan H, Zhou Z, Zhou
J and Qu P: microRNA-125b inhibits cell migration and invasion by
targeting matrix metallopeptidase 13 in bladder cancer. Oncol Lett.
5:829–834. 2013.PubMed/NCBI
|
|
16
|
Bartel DP: MicroRNAs: target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ghildiyal M and Zamore PD: Small silencing
RNAs: an expanding universe. Nat Rev Genet. 10:94–108. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Takahashi RU, Makiko O and Ochiya T: Role
of microRNA in cancer development: biology and clinical
applications. Nihon Geka Gakkai Zasshi. 113:197–203. 2012.(In
Japanese). PubMed/NCBI
|
|
21
|
Farazi TA, Hoell JI, Morozov P and Tuschl
T: MicroRNAs in human cancer. Adv Exp Med Biol. 774:1–20. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ura S, Honda M, Yamashita T, Ueda T,
Takatori H, Nishino R, Sunakozaka H, Sakai Y, Horimoto K and Kaneko
S: Differential microRNA expression between hepatitis B and
hepatitis C leading disease progression to hepatocellular
carcinoma. Hepatology. 49:1098–1112. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ventura A and Jacks T: MicroRNAs and
cancer: short RNAs go a long way. Cell. 136:586–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu D, Zhou Y, Pan H, Zhou J, Fan J and Qu
P: microRNA-99a inhibiting cell proliferation, migration and
invasion by targeting fibroblast growth factor receptor 3 in
bladder cancer. Oncol Lett. 7:1219–1224. 2014.PubMed/NCBI
|
|
25
|
Zhou Y, Wu D, Tao J, Qu P, Zhou Z and Hou
J: MicroRNA-133 inhibits cell proliferation, migration and invasion
by targeting epidermal growth factor receptor and its downstream
effector proteins in bladder cancer. Scand J Urol. 47:423–432.
2013. View Article : Google Scholar
|
|
26
|
Kawakami K, Enokida H, Chiyomaru T,
Tatarano S, Yoshino H, Kagara I, Gotanda T, Tachiwada T, Nishiyama
K, Nohata N, et al: The functional significance of miR-1 and
miR-133a in renal cell carcinoma. Eur J Cancer. 48:827–836. 2012.
View Article : Google Scholar
|
|
27
|
Kano M, Seki N, Kikkawa N, Fujimura L,
Hoshino I, Akutsu Y, Chiyomaru T, Enokida H, Nakagawa M and
Matsubara H: miR-145, miR-133a and miR-133b: Tumor-suppressive
miRNAs target FSCN1 in esophageal squamous cell carcinoma. Int J
Cancer. 127:2804–2814. 2010. View Article : Google Scholar
|
|
28
|
Chiyomaru T, Enokida H, Tatarano S,
Kawahara K, Uchida Y, Nishiyama K, Fujimura L, Kikkawa N, Seki N
and Nakagawa M: miR-145 and miR-133a function as tumour suppressors
and directly regulate FSCN1 expression in bladder cancer. Br J
Cancer. 102:883–891. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ruebel K, Leontovich AA, Stilling GA,
Zhang S, Righi A, Jin L and Lloyd RV: MicroRNA expression in ileal
carcinoid tumors: downregulation of microRNA-133a with tumor
progression. Mod Pathol. 23:367–375. 2010. View Article : Google Scholar :
|
|
30
|
Rao PK, Missiaglia E, Shields L, Hyde G,
Yuan B, Shepherd CJ, Shipley J and Lodish HF: Distinct roles for
miR-1 and miR-133a in the proliferation and differentiation of
rhabdomyosarcoma cells. FASEB J. 24:3427–3437. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chomczynski P and Sacchi N: The
single-step method of RNA isolation by acid guanidinium
thiocyanate-phenol-chloroform extraction: twenty-something years
on. Nature Protoc. 1:581–585. 2006. View Article : Google Scholar
|
|
32
|
Tao J, Lu Q, Wu D, Li P, Xu B, Qing W,
Wang M, Zhang Z and Zhang W: microRNA-21 modulates cell
proliferation and sensitivity to doxorubicin in bladder cancer
cells. Oncol Rep. 25:1721–1729. 2011.PubMed/NCBI
|
|
33
|
Dong DL, Chen C, Huo R, Wang N, Li Z, Tu
YJ, Hu JT, Chu X, Huang W and Yang BF: Reciprocal repression
between microRNA-133 and calcineurin regulates cardiac hypertrophy:
a novel mechanism for progressive cardiac hypertrophy.
Hypertension. 55:946–952. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tao J, Wu D, Xu B, Qian W, Li P, Lu Q, Yin
C and Zhang W: microRNA-133 inhibits cell proliferation, migration
and invasion in prostate cancer cells by targeting the epidermal
growth factor receptor. Oncol Rep. 27:1967–1975. 2012.PubMed/NCBI
|
|
35
|
Rao PK, Kumar RM, Farkhondeh M,
Baskerville S and Lodish HF: Myogenic factors that regulate
expression of muscle-specific microRNAs. Proc Nat Acad Sci USA.
103:8721–8726. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bostjancic E, Zidar N, Stajer D and Glavac
D: MicroRNAs miR-1, miR-133a, miR-133b and miR-208 are dysregulated
in human myocardial infarction. Cardiology. 115:163–169. 2010.
View Article : Google Scholar
|
|
37
|
Uchida Y, Chiyomaru T, Enokida H, Kawakami
K, Tatarano S, Kawahara K, Nishiyama K, Seki N and Nakagawa M:
MiR-133a induces apoptosis through direct regulation of GSTP1 in
bladder cancer cell lines. Urol Oncol. 31:115–123. 2013. View Article : Google Scholar
|
|
38
|
Chiyomaru T, Enokida H, Kawakami K,
Tatarano S, Uchida Y, Kawahara K, Nishiyama K, Seki N and Nakagawa
M: Functional role of LASP1 in cell viability and its regulation by
microRNAs in bladder cancer. Urol Oncol. 30:434–443. 2012.
View Article : Google Scholar
|
|
39
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
40
|
Han X, Yan DM, Zhao XF, Matsuura H, Ding
WG, Li P, Jiang S, Du BR, Du PG and Zhu X: GHGKHKNK octapeptide
(P-5m) inhibits metastasis of HCCLM3 cell lines via regulation of
MMP-2 expression in in vitro and in vivo studies. Molecules.
17:1357–1372. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lou L, Chen YX, Jin L, Li X, Tao X, Zhu J,
Chen X, Wu S, Ye W, He J, et al: Enhancement of invasion of
hepatocellular carcinoma cells through lysophosphatidic acid
receptor. J Int Med Res. 41:55–63. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sun MH, Han XC, Jia MK, Jiang WD, Wang M,
Zhang H, Han G and Jiang Y: Expressions of inducible nitric oxide
synthase and matrix metalloproteinase-9 and their effects on
angiogenesis and progression of hepatocellular carcinoma. W J
Gastroenterol. 11:5931–5937. 2005.
|
|
43
|
Verma S, Kesh K, Ganguly N, Jana S and
Swarnakar S: Matrix metalloproteinases and gastrointestinal
cancers: Impacts of dietary antioxidants. World J Biol Chem.
5:355–376. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kallakury BV, Karikehalli S, Haholu A,
Sheehan CE, Azumi N and Ross JS: Increased expression of matrix
metalloproteinases 2 and 9 and tissue inhibitors of
metalloproteinases 1 and 2 correlate with poor prognostic variables
in renal cell carcinoma. Clin Cancer Res. 7:3113–3119.
2001.PubMed/NCBI
|
|
45
|
Morgia G, Falsaperla M, Malaponte G,
Madonia M, Indelicato M, Travali S and Mazzarino MC: Matrix
metalloproteinases as diagnostic (MMP-13) and prognostic (MMP-2,
MMP-9) markers of prostate cancer. Urol Res. 33:44–50. 2005.
View Article : Google Scholar
|
|
46
|
Gao ZH, Tretiakova MS, Liu WH, Gong C,
Farris PD and Hart J: Association of E-cadherin, matrix
metalloproteinases, and tissue inhibitors of metalloproteinases
with the progression and metastasis of hepatocellular carcinoma.
Mod Pathol. 19:533–540. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jang JW, Park ST, Kwon JH, You CR, Choi
JY, Jung CK, Bae SH and Yoon SK: Suppression of hepatic tumor
growth and metastasis by metronomic therapy in a rat model of
hepatocellular carcinoma. Exp Mol Med. 43:305–312. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xiang ZL, Zeng ZC, Fan J, Tang ZY, Zeng HY
and Gao DM: Gene expression profiling of fixed tissues identified
hypoxia-inducible factor-1alpha, VEGF, and matrix
metalloproteinase-2 as biomarkers of lymph node metastasis in
hepatocellular carcinoma. Clin Cancer Res. 17:5463–5472. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chen R, Cui J, Xu C, Xue T, Guo K, Gao D,
Liu Y, Ye S and Ren Z: The significance of MMP-9 over MMP-2 in HCC
invasiveness and recurrence of hepatocellular carcinoma after
curative resection. Ann Surg Oncol. 19(Suppl 3): S375–S384. 2012.
View Article : Google Scholar
|
|
50
|
Hong S, Park KK, Magae J, Ando K, Lee TS,
Kwon TK, Kwak JY, Kim CH and Chang YC: Ascochlorin inhibits matrix
metalloproteinase-9 expression by suppressing activator
protein-1-mediated gene expression through the ERK1/2 signaling
pathway: inhibitory effects of ascochlorin on the invasion of renal
carcinoma cells. J Biol Chem. 280:25202–25209. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Santhekadur PK, Gredler R, Chen D, Siddiq
A, Shen XN, Das SK, Emdad L, Fisher PB and Sarkar D: Late SV40
factor (LSF) enhances angiogenesis by transcriptionally
up-regulating matrix metalloproteinase-9 (MMP-9). J Biol Chem.
287:3425–3432. 2012. View Article : Google Scholar :
|
|
52
|
Jiang YF, Yang ZH and Hu JQ: Recurrence or
metastasis of HCC: predictors, early detection and experimental
antiangiogenic therapy. World J Gastroenterol. 6:61–65. 2000.
|
|
53
|
Hayasaka A, Suzuki N, Fujimoto N, Iwama S,
Fukuyama E, Kanda Y and Saisho H: Elevated plasma levels of matrix
metalloproteinase-9 (92-kd type IV collagenase/gelatinase B) in
hepatocellular carcinoma. Hepatology. 24:1058–1062. 1996.
View Article : Google Scholar : PubMed/NCBI
|