Introduction
Pulmonary arterial hypertension (PAH) is a complex,
multi-factorial disorder in which pulmonary arterial obstruction
increases pulmonary vascular resistance, resulting in an overload
of the right ventricle and progressive right-sided heart
dysfunction (1,2). Excessive proliferation of pulmonary
artery smooth muscle cells (PASMCs) is a hallmark feature of
pulmonary vascular remodeling, which is a critical factor in the
pathogenesis and development of PAH (3,4).
Various stimuli are able to induce the proliferation of PASMCs,
resulting in arterial wall remodeling and PAH (5). As a potential mitogen and
chemoattractant, platelet-derived growth factor (PDGF) has
previously been demonstrated to have a crucial role in the
pathogenesis of PAH. In addition, the upregulated expression levels
of PDGF and PDGF receptor have been correlated with PAH in various
experimental animal models (6,7) and
in humans (8,9). There are numerous members of the PDGF
protein family: A, B, C and D. PDGF-BB is a potent inducer of
vascular smooth muscle cell (VSMC) proliferation, and has been
proposed to have important roles in the development of
atherosclerosis, lung fibrosis and PAH (10–12).
Hesperetin is a member of the flavanone subclass of
flavonoids, which exists naturally in its glycoside form,
hesperedin. Hesperetin has previously been reported to exert
various biological activities, including anti-cancer,
anti-oxidative, anti-inflammatory and neuroprotective effects
(13–19). A previous study demonstrated that
hesperetin was able to alleviate the inhibitory effects of
high-level glucose on the osteoblastic differentiation of
periodontal ligament stem cells by regulating the levels of
reactive oxygen species, and the phosphoinositide 3-kinase/AKT and
β-catenin signaling pathways (20). In addition, hesperetin may induce
G1 phase cell cycle arrest in MCF-7 human breast cancer
cells by regulating cyclin-dependent kinase (CDK)4 and p21
expression (15), and may inhibit
the proliferation of breast cancer cells by downregulating the
expression of glucose transporter 1, and the phosphorylation of the
insulin receptor-β subunit and AKT (13). However, the effects of hesperetin
on the proliferation of PASMCs remain largely unknown.
The present study aimed to investigate whether
hesperetin could inhibit the proliferation of PDGF-BB-induced
PASMCs, and to examine the underlying molecular mechanisms.
Materials and methods
Materials
Hesperetin (98% purity, as determined by
high-performance liquid chromatography analysis) was purchased from
Shanghai Winherb Medical S&T Development Co., Ltd. (Shanghai,
China). Recombinant human PDGF-BB was obtained from Peprotech, Inc.
(Rocky Hill, NJ, USA). Cell Counting kit (CCK)-8 was obtained from
Dojindo Molecular Technologies, Inc. (Kumamoto, Japan). Cell
proliferation enzyme-linked immunosorbent assay (ELISA) and
bromodeoxyuridine (BrdU) proliferation assay kits were obtained
from Roche Diagnostics GmbH (Mannheim, Germany). TRIzol®
was purchased from Invitrogen (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). Antibodies targeting total (T) and
phosphorylated (P) forms of extracellular signal-regulated kinases
(ERK)1/2 (cat. nos. 4695 and 4370, respectively), T-p38 (cat. no.
92120), P-p38 (cat. no. 45110), T-c-Jun N-terminal kinases (T-JNK)
(cat. no. 9258), P-JNK (cat. no. 4668), P-AKT (cat. no. 4691),
T-AKT (cat. no. 4060), T-glycogen synthase kinase (T-GSK)3β (cat.
no. 9315), P-GSK3β (cat. no. 9322) and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH; cat. no. 2118) were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Adult male Sprague
Dawley rats (weight, 150–200 g) were obtained from Wuhan University
Center for Animal Experiment (Wuhan, China). Animals were kept
under a 12-h light/dark cycle and maintained at a constant
temperature of 22°C with free access to food and water. All of the
animal experiments were approved by the Institutional Animal Care
and Use Committee of the Renmin Hospital of Wuhan University
(Wuhan, China).
PASMC isolation and cell culture
Sprague-Dawley rats (weight, 150–200 g) were
intraperitoneally administered 1% sodium pentobarbital (50 mg/kg;
Sinopharm Chemical Reagent Co., Shanghai, China). Subsequently, the
pleura of each rat were rapidly sectioned, and the heart and lungs
were removed and transferred to a petri dish filled with
phosphate-buffered saline, in order to wash off the residual blood.
To separate the pulmonary artery, the outer fibrous arterial
connective tissue was stripped under a microscope (CKX41; Olympus,
Tokyo, Japan) using tweezers, and was rinsed several times in
Dulbecco's modified Eagle's medium/F12 (DMEM/F12) supplemented with
1% penicillin-streptomycin (Gibco; Thermo Fisher Scientific). The
artery was subsequently cut into 1 mm sections and was digested in
a centrifuge tube pre-filled with 0.2% collagenase I. Every 20 to
30 min, the mixture was observed and gently agitated. The total
duration of the digestion was 2–4 h. When the arterial fragments
had been digested, the samples were centrifuged at 195 × g for 5
min, and the supernatant was discarded. The pellet was rinsed with
DMEM/F12 supplemented with 20% fetal bovine serum (FBS) (both from
Gibco) and was cultured at 37°C in an incubator containing 5%
CO2. After 4–5 days, the cells were passaged and grown
in DMEM/F12 supplemented with 10% FBS. The purity of the PASMC
cultures was assessed by immunocytochemical localization of
α-smooth muscle actin according to a previously described procedure
(21). The cells from passages 4
and 10 were used in the present study. The PASMCs were grown to
70–80% confluence in 96-well microplates and were then subjected to
serum starvation for 24 h prior to experimentation.
Drug administration
After 24 h serum starvation, the cells were
pretreated with various concentrations of hesperetin for 1 h. They
were subsequently treated with PDGF-BB (20 ng/ml) for 12, 24 and 48
h in the presence of hesperetin.
Measurement of cell proliferation and DNA
synthesis
Cell proliferation was determined using the CCK-8
assay, according to the manufacturer's protocol. The cells were
treated with PDGF-BB (20 ng/ml) for 12, 24 and 48 h in the presence
of hesperetin, and incubated with CCK-8 for the final 3 h. Cell
proliferation was determined by measuring the absorbance at 450 nm.
DNA synthesis was determined using a BrdU assay. Briefly, BrdU
labeling mixture was added to each well and the cells were
incubated for 2 h. Finally, DNA synthesis was determined by
measuring the incorporation of BrdU, using a cell proliferation
ELISA kit as previously described (21) and a Synergy HT plate reader (BioTek
Instruments, Inc., Winooski, VT, USA).
Evaluation of cell viability
The toxicity of hesperetin on PASMCs was determined
using the trypan blue exclusion method (Sinopharm Chemical Reagent
Co.). After 12-, 24- and 48-h incubations with hesperetin (12.5–100
µM), the PASMCs were removed from the culture and the cells
that excluded 0.4% trypan blue were counted in an automated cell
counter (Invitrogen; Thermo Fisher Scientific, Inc.) (21).
Cell cycle analysis
The cell cycle distribution was determined using a
Cell Cycle and Apoptosis Analysis kit (Beyotime Institute of
Biotechnology, Haimen, China) according to the manufacturer's
protocol. Propidium iodide (PI) staining and fluorescence-activated
cell sorting (FACS) were performed. Once the PASMCs had reached
70–80% confluence in six-well plates, they were subjected to serum
starvation for 24 h. The cells were subsequently pre-incubated with
hesperetin (100 µM) for 1 h and subsequently treated with
PDGF-BB (20 ng/ml) for 24 h prior to analysis. The cells were then
trypsinized (Hyclone Laboratories, Inc., Logan, UT, USA) and fixed
with 70% ethanol overnight. The fixed cells were collected by
centrifugation at 438.75 × g for 5 min, washed once in
phosphate-buffered saline, incubated with 0.5 ml PI staining buffer
and analyzed using FACS. Cell cycle distributions were analyzed
using Multicycle AV software, version 2.50 (Phoenix Flow Systems,
San Diego, CA, USA).
Reverse transcription-quantitative
polymerase chain reaction (qPCR)
After 24 h serum starvation and preincubation with
100 µM hesperetin for 1 h, the cells were treated with
PDGF-BB for 24 h in the absence or presence of hesperetin. Total
RNA was extracted from the PASMCs using TRIzol®
(Invitrogen) and its yield and purity was spectrophotometrically
estimated via the A260/A280 and A230/260 ratios using a SmartSpec
Plus Spectrophotometer (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The RNA (2 µg) was reverse-transcribed into cDNA using
oligo (dT18) primers from the Transcriptor First Strand cDNA
Synthesis kit (cat. no. 04896866001; Roche Diagnostics GmbH) using
oligo(dT) primers. qPCR was conducted using the LightCycler 480
SYBR Green 1 Master mix (Roche Diagnostics GmbH) and the
LightCycler 480 qPCR system (Roche Diagnostics GmbH). The PCR
cycling conditions were as follows: Initial denaturation at 95°C
for 10 min, followed by 40 cycles of 95°C for 15 sec and 60°C for 1
min. Target gene mRNA expression was normalized to the GAPDH
internal control and was expressed relative to the control group
using the ΔΔCT method (22). The
primer sequences (Sangon Biotech, Shanghai, China) used were as
follows: Cyclin D1 forward, 5′-GAGACCATCCCCCTGACGGC-3′ and reverse,
5′-TCTTCCTCCTCCTCGGCGGC-3′; Cyclin E forward,
5′-GTCCTGGCTGAATGTATACATGC-3′, and reverse,
5′-CCCTATTTTGTTCAGACAACATGGC-3′; CDK2 forward,
5′-GCTTTCTGCCATTCTCATCG-3′ and reverse, 5′-GTCCCCAGAGTCCGAAAGAT-3′;
CDK4 forward, 5′-ATGTTGTCCGGCTGATGG-3′ and reverse,
5′-CACCAGGGTTACCTTGATCTCC-3′; p27 forward,
5′-TGCAACCGACGATTCTTCTACTCAA-3′ and reverse,
5′-CAAGCAGTGATGTATCTGATAAACAAGGA-3′; GAPDH forward,
5′-ATTCCATGGCACCGTCAAGG-3′ and reverse,
5′-AATTCGTTGTCATACCAGGA-3′.
Western blot analysis
Confluent serum-starved PASMCs were treated with
hesperetin (100 µM) for 1 h following exposure to 20 ng/ml
PDGF-BB for 5, 10 or 15 min. The cells were lysed in
radioimmunoprecipitation assay buffer supplemented with a protease
and phosphatase inhibitor cocktail (Thermo Fisher Scientific) and
were centrifuged at 3,362 × g for 30 min at 4°C. The protein
concentration was assessed using a bicinchoninic acid assay (Thermo
Fisher Scientific). The protein extracts (20 µg) were
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and blotted onto Immobilon-FL transfer membranes
(Millipore, Billerica, MA, USA). The blotted membranes were blocked
with 5% skimmed milk in Tris-buffered saline containing 0.1% Tween
20 for 2 h and were subsequently incubated with primary antibodies
against T-ERK1/2, P-ERK1/2, T-p38, P-p38, T-JNK, P-JNK, T-GSK3β,
P-GSK3β, T-AKT, P-AKT and GAPDH overnight at 4°C. After three
washes in Tris-buffered saline containing 0.1% Tween 20, the
membranes were incubated with the secondary antibody, IRdye 800
anti-rabbit (cat. no. 926-32211; LI-COR Biosciences, Lincoln, NE,
USA) at a dilution of 1:2,500 for 1 h. The bands were quantified
using the Odyssey infrared imaging system (LI-COR Biosciences). The
protein expression levels were normalized to the expression levels
of the GAPDH internal control.
Statistical analysis
The results are expressed as the mean ± standard
deviation. Differences among the groups were assessed using one-way
analysis of variance or unpaired two-tailed t-tests. All
statistical analyses were performed using SPSS 16.0 software (SPSS,
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Effects of hesperetin on the
proliferation of PDGF-BB-induced PASMCs
As compared with the control cells, PDGF-BB
significantly increased the optical density (OD) of the cells in a
time-dependent manner, which represented the time-dependent
proliferation of PASMCs. In addition, pretreatment with hesperetin
significantly suppressed the PDGF-BB-increased proliferation in a
concentration-dependent manner, and the greatest suppression of
proliferation was observed when the cells were treated with 100
µM hesperetin (Fig. 1A).
Similarly, treatment with PDGF-BB increased DNA synthesis in PASMCs
in a time-dependent manner, whereas treatment with hesperetin
significantly suppressed the increase in DNA synthesis in a
dose-and time-dependent manner (Fig.
1B).
Toxicity of hesperetin on PASMCs
Treatment with hesperetin at concentrations between
12.5 and 100 µM did not induce significant levels of cell
necrosis in PASMCs after 12, 24 or 48 h, as compared with the
untreated cells (Fig. 2).
Regardless of whether the cells were treated with hesperetin or
not, cell viability remained at ~95%. This result suggests that
hesperetin is not cytotoxic at the tested concentrations.
Effects of hesperetin on cell cycle
progression
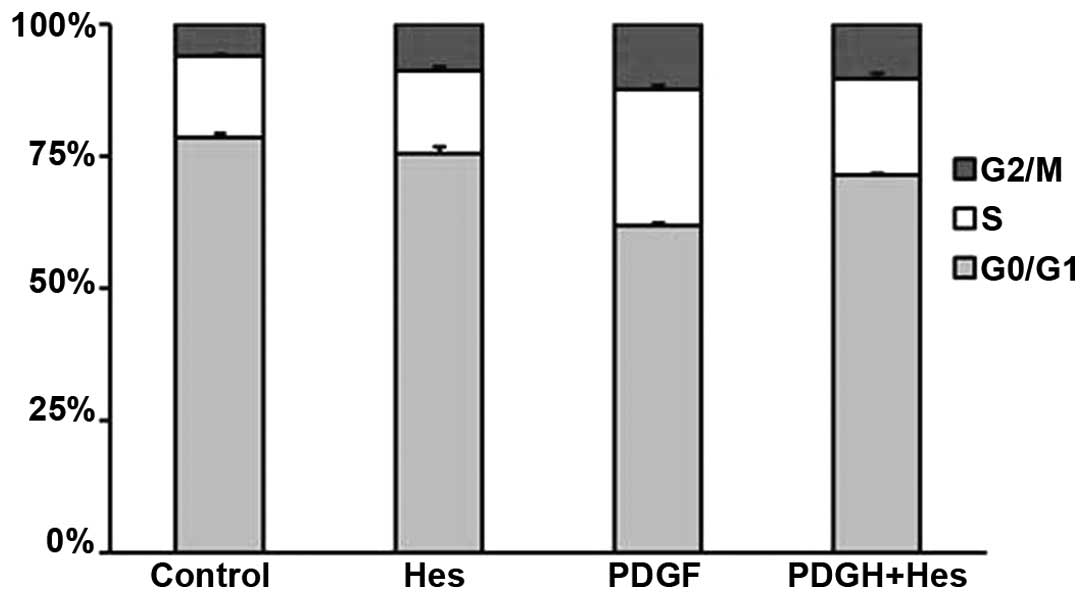
As shown in Fig. 3,
the percentage of cells in S phase increased from 15.3 to 25.7%
following PDGF-BB stimulation. In addition, the percentage of cells
in G0/G1 phase decreased from 78.7 to 62.0%.
These effects were markedly reversed by pretreatment with 100
µM hesperetin. The percentage of cells in S phase decreased
to 17.7%, whereas the percentage of cells in
G0/G1 phase increased to 69.7%. These results
indicate that hesperetin affects G0/G1 to S
phase transition; however, no effect was detected on the transition
between S and G2/M phase.
Effects of hesperetin on associated gene
expression
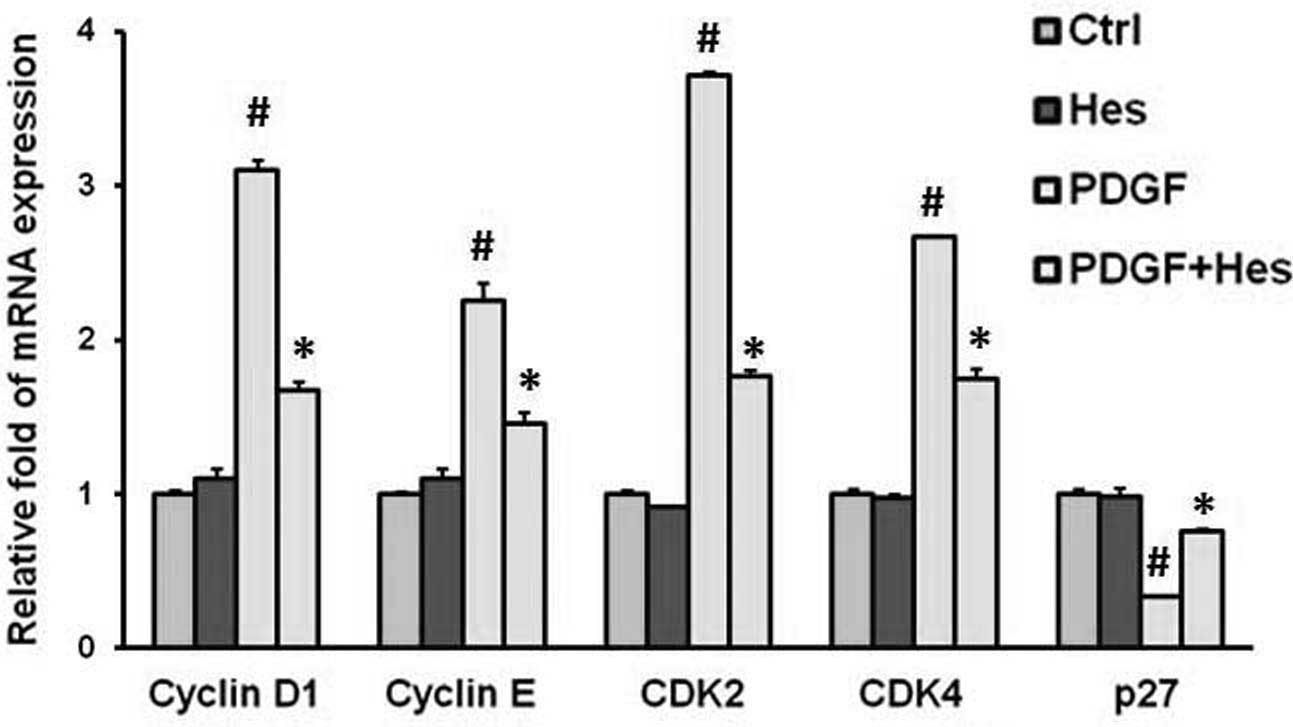
Treatment with PDGF-BB significantly increased the
mRNA expression levels of cyclin D1, cyclin E, CDK2 and CDK4.
Conversely, pretreatment with hesperetin significantly suppressed
the PDGF-BB-induced upregulation of the studied genes (Fig. 4). Cyclin-CDK complexes, which are
formed during cell cycle progression, are regulated by CDK
inhibitors (CKIs), including p27, which lead to cell cycle arrest
at the G1 and G1/S boundary (23). As shown in Fig. 4, hesperetin upregulated the
expression levels of p27.
Involvement of mitogen-activated protein
kinases (MAPK) and AKT/GSK3β signaling in the hesperetin-induced
inhibition of PASMCs proliferation
Significant activation of ERK1/2, p38, JNK and
AKT/GSK3β was observed 5, 10 and 15 min after PDGF treatment;
however, the total levels of these molecules were not affected, as
determined by western blotting (Fig.
5). In addition, hesperetin significantly reduced the
phosphorylation of AKT/GSK3β and p38, but did not exhibit any
inhibitory effects on the phosphorylation of ERK1/2 and JNK
(Fig. 5).
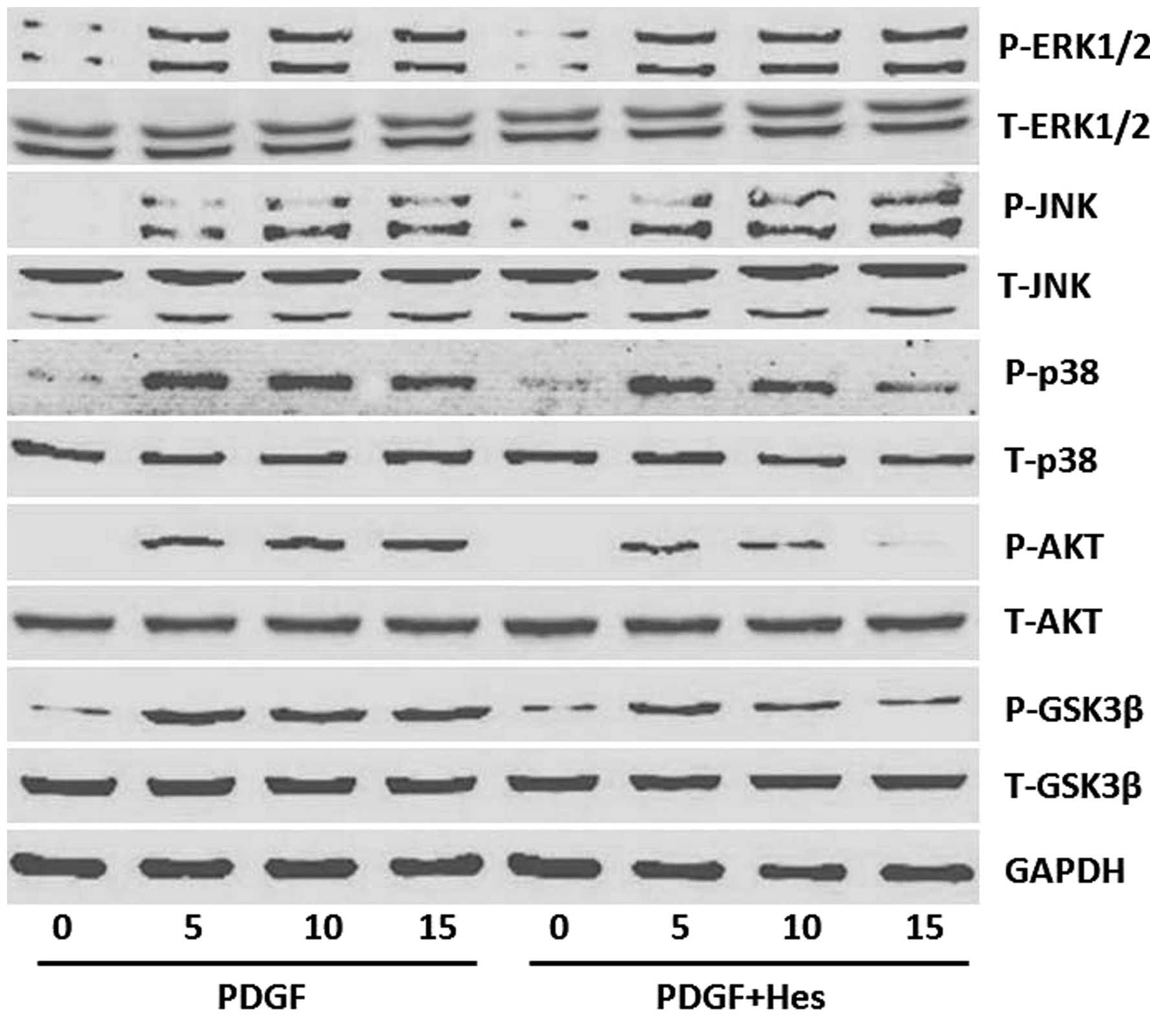 | Figure 5Effects of Hes on the activation of
signaling pathways in PDGF-BB-stimulated PASMCs. PASMCs were
pretreated with Hes (100 µM) for 1 h prior to treatment with
20 ng/ml PDGF-BB. The protein expression levels of P-ERK1/2,
T-ERK1/2, P-JNK, T-JNK, P-p38, T-p38, P-AKT, T-AKT, P-GSK3β and
T-GSK3β were determined after 5, 10 and 15 min by western blot
analysis. One representative image out of three independently
performed experiments is shown. T, total; P, phosphorylated; ERK,
extracellular signal-regulated kinases; JNK, c-Jun N-terminal
kinases; GSK, glycogen synthase kinase; PDGF, platelet-derived
growth factor; PASMCs, pulmonary artery smooth muscle cells; Hes,
hesperetin. |
Discussion
PAH, which is characterized by a sustained increase
in pulmonary artery pressure and pulmonary vascular remodeling, is
a progressive disease that is associated with a poor prognosis
(24,25). PAH is associated with the excessive
proliferation of PASMCs; therefore, inhibition of PASMC
proliferation may be considered an effective therapeutic strategy.
The present study demonstrated that hesperetin inhibited
PDGF-induced PASMC proliferation and DNA synthesis in a
concentration- and time-dependent manner, without exerting cell
cytotoxic effects.
Cell proliferation is tightly regulated by the cell
cycle, and hesperetin has been reported to induce cell cycle arrest
in cancer cells (15). In the
present study, flow cytometric analysis demonstrated that treatment
with 100 µM hesperetin for 24 h led to a significant
increase in the number of cells in G0/G1
phase and a decrease in the number of cells in S phase, without any
significant effect on the number of cells in G2/M phase.
According to these results, hesperetin suppressed the
PDGF-BB-induced progression of PASMCs from
G0/G1 to S phase. Progression through the
cell cycle is regulated by the coordinated activities of cyclin-CDK
complexes. Regulation of these cyclin-CDK complexes may be provided
by their binding to CKIs (26).
CDK2 and CDK4 form complexes with cyclin E and cyclin D1, which are
essential for the cell cycle progression from
G0/G1 phase to S phase (27,28).
In addition, as a CKI, p27 phosphorylation has been reported to
inhibit CDK2-containing complexes and cyclin D-CDK4 complexes
(29,30).
In the present study, the expression levels of cell
cycle regulatory genes in PASMCs were detected in response to
PDGF-BB. Hesperetin decreased the mRNA expression levels of cyclin
D1, cyclin E, CDK2 and CDK4, and increased the mRNA expression
levels of p27. These results suggested that the anti-proliferative
activity of hesperetin has multifaceted effects on numerous target
molecules involved in growth inhibition.
MAPK families (including ERKs, p38 and JNK) and the
AKT pathway have an important role in the regulation of cell
proliferation. It has previously been reported that PDGF-BB
significantly stimulates the phosphorylation of ERK1/2, JNK, p38
and AKT/GSK3β in PASMCs (31). AKT
is involved in the regulation of various downstream signaling
pathways, including those associated with metabolism, cell
proliferation, survival, growth and angiogenesis; GSK3β is one of
these critical downstream molecules. The results of the present
study demonstrated that hesperetin was able to inhibit the
phosphorylation of AKT/GSK3β after PDGF-BB induction. These results
are consistent with previous studies, which have reported that
hesperetin may inhibit the AKT pathway (13,17).
However, other studies have reported conflicting results, in which
hesperetin has been shown to stimulate the AKT pathway (20,32).
In rat aortic vascular smooth muscle cells, hesperetin exerted
anti-proliferative activities; however, it had no effect on
PDGF-BB-stimulated ERK1/2, AKT, JNK and p38 phosphorylation
(33). Whether these differences
are associated with the various types of cells used, different
biological effects or different concentrations remains to be
elucidated.
Cyclin D1 is regulated by GSK3β (34,35).
Activation of GSK3β has been reported to regulate cyclin D1 export
from the nucleus to the cytoplasm for proteolysis, thus
downregulating the expression of cyclin D1 (36). Furthermore, inhibition of the
phosphorylation of the AKT/GSK3β signaling pathway has been
reported to decrease the expression levels of cyclin D1 in cultured
VSMCs (37). Inhibition of GSK3β
has also been shown to decrease the expression levels of the CKI,
p27 (38). These results suggested
that hesperetin may have an anti-proliferative effect on PASMCs via
the AKT/GSK3β signaling pathway.
The present study also examined the effects of
hesperetin on the activation of MAPKs, including ERK1/2, p38 and
JNK, which are important factors implicated in the proliferation of
PASMCs. p38 has an important role in the proliferation of PASMCs,
whereas hesperetin may inhibit the activation of p38 MAPK.
Therefore, it was hypothesized that the anti-proliferative effects
of hesperetin may be associated with the inhibition of p38, thus
suggesting that hesperetin may inhibit the PGDF-BB-induced
proliferation of PASMCs via regulating p38 MAPK signal
transduction.
In conclusion, the present study demonstrated that
hesperetin was able to inhibit the PDGF-BB-induced proliferation of
PASMCs. Notably, this process appears to be associated with an
inhibition of cyclin D1 expression and an increased expression of
p27, via suppression of the AKT/GSK3β and p38 signaling pathway.
These results suggest that hesperetin may be a potential
therapeutic strategy for the treatment of pulmonary vascular
remodeling diseases.
Acknowledgments
The present study was supported by the National
Nature Science Foundation of China (grant nos. 81270303 and
81300104), the Specialized Research Fund for the Doctoral Program
of Higher Education of China (grant no. 20130141120042), Hubei
Province's Outstanding Medical Academic Leader Program, the Natural
Science Foundation of Hubei Province, China (grant no. 2013CFB303)
and the Fundamental Research Funds for the Central Universities
(grant no. 2042014kf0130).
References
|
1
|
Archer SL, Weir EK and Wilkins MR: Basic
science of pulmonary arterial hypertension for clinicians: New
concepts and experimental therapies. Circulation. 121:2045–2066.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tuder RM, Abman SH, Braun T, Capron F,
Stevens T, Thistlethwaite PA and Haworth SG: Development and
pathology of pulmonary hypertension. J Am Coll Cardiol. 54(Suppl
1): S3–S9. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pietra GG, Capron F, Stewart S, Leone O,
Humbert M, Robbins IM, Reid LM and Tuder RM: Pathologic assessment
of vasculopathies in pulmonary hypertension. J Am Coll Cardiol.
43(12 Suppl S): 25S–32S. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Crosswhite P and Sun Z: Molecular
mechanisms of pulmonary arterial remodeling. Mol Med. 20:191–201.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yildiz P: Molecular mechanisms of
pulmonary hypertension. Clin Chim Acta. 403:9–16. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Balasubramaniam V, Le Cras TD, Ivy DD,
Grover TR, Kinsella AP and Abman SH: Role of platelet-derived
growth factor in vascular remodeling during pulmonary hypertension
in the ovine fetus. Am J Physiol Lung Cell Mol Physiol.
284:L826–L833. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schermuly RT, Dony E, Ghofrani HA,
Pullamsetti S, Savai R, Roth M, Sydykov A, Lai YJ, Weissmann N,
Seeger W and Grimminger F: Reversal of experimental pulmonary
hypertension by PDGF inhibition. J Clin Invest. 115:2811–2821.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Perros F, Montani D, Dorfmüller P,
Durand-Gasselin I, Tcherakian C, Le Pavec J, Mazmanian M, Fadel E,
Mussot S, Mercier O, et al: Platelet-derived growth factor
expression and function in idiopathic pulmonary arterial
hypertension. Am J Respir Crit Care Med. 178:81–88. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Humbert M, Monti G, Fartoukh M, Magnan A,
Brenot F, Rain B, Capron F, Galanaud P, Duroux P, Simmoneau G and
Emilie D: Platelet-derived growth factor expression in primary
pulmonary hypertension: Comparison of HIV seropositive and HIV
seronegative patients. Eur Respir J. 11:554–559. 1998.PubMed/NCBI
|
|
10
|
Sachinidis A, Locher R, Hoppe J and Vetter
W: The platelet-derived growth factor isomers, PDGF-AA, PDGF-AB and
PDGF-BB, induce contraction of vascular smooth muscle cells by
different intracellular mechanisms. FEBS Lett. 275:95–98. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sakao S, Tatsumi K and Voelkel NF:
Reversible or irreversible remodeling in pulmonary arterial
hypertension. Am J Respir Cell Mol Biol. 43:629–634. 2010.
View Article : Google Scholar :
|
|
12
|
Jawien A, Bowen-Pope DF, Lindner V,
Schwartz SM and Clowes AW: Platelet-derived growth factor promotes
smooth muscle migration and intimal thickening in a rat model of
balloon angioplasty. J Clin Invest. 89:507–511. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang Y, Wolfram J, Boom K, Fang X, Shen H
and Ferrari M: Hesperetin impairs glucose uptake and inhibits
proliferation of breast cancer cells. Cell Biochem Funct.
31:374–379. 2013. View
Article : Google Scholar
|
|
14
|
Zarebczan B, Pinchot SN, Kunnimalaiyaan M
and Chen H: Hesperetin, a potential therapy for carcinoid cancer.
Am J Surg. 201:329–333. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Choi EJ: Hesperetin induced G1-phase cell
cycle arrest in human breast cancer MCF-7 cells: Involvement of
CDK4 and p21. Nutr Cancer. 59:115–119. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J, Zhu H, Yang Z and Liu Z:
Antioxidative effects of hesperetin against lead acetate-induced
oxidative stress in rats. Indian J Pharmacol. 45:395–398. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Deng W, Jiang D, Fang Y, Zhou H, Cheng Z,
Lin Y, Zhang R, Zhang J, Pu P, Liu Y, Bian Z and Tang Q: Hesperetin
protects against cardiac remodelling induced by pressure overload
in mice. J Mol Histol. 44:575–585. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cho J: Antioxidant and neuroprotective
effects of hesperidin and its aglycone hesperetin. Arch Pharm Res.
29:699–706. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang HL, Chen SC, Senthil Kumar KJ, Yu KN,
Lee Chao PD, Tsai SY, Hou YC and Hseu YC: Antioxidant and
anti-inflammatory potential of hesperetin metabolites obtained from
hesperetin-administered rat serum: An ex vivo approach. J Agric
Food Chem. 60:522–532. 2012. View Article : Google Scholar
|
|
20
|
Kim SY, Lee JY, Park YD, Kang KL, Lee JC
and Heo JS: Hesperetin alleviates the inhibitory effects of high
glucose on the osteoblastic differentiation of periodontal ligament
stem cells. PLoS One. 8:e675042013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen C, Tang Y, Deng W, Huang C and Wu T:
Salidroside blocks the proliferation of pulmonary artery smooth
muscle cells induced by platelet-derived growth factor-BB. Mol Med
Rep. 10:917–922. 2014.PubMed/NCBI
|
|
22
|
Deng W, Jiang D, Fang Y, Zhou H, Cheng Z,
Lin Y, Zhang R, Pu P, Liu Y, et al: Hesperetin protects against
cardiac remodelling induced by pressure overload in mice. J Mol
Histol. 44:575–585. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Abukhdeir AM and Park BH: P21 and p27:
Roles in carcinogenesis and drug resistance. Expert Rev Mol Med.
10:e192008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cogolludo A, Moreno L and Villamor E:
Mechanisms controlling vascular tone in pulmonary arterial
hypertension: Implications for vasodilator therapy. Pharmacology.
79:65–75. 2007. View Article : Google Scholar
|
|
25
|
Paulin R, Meloche J and Bonnet S: STAT3
signaling in pulmonary arterial hypertension. JAKSTAT. 1:223–233.
2012.
|
|
26
|
Besson A, Dowdy SF and Roberts JM: CDK
inhibitors: Cell cycle regulators and beyond. Dev Cell. 14:159–169.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jirawatnotai S, Aziyu A, Osmundson EC,
Moons DS, Zou X, Kineman RD and Kiyokawa H: Cdk4 is indispensable
for postnatal proliferation of the anterior pituitary. J Biol Chem.
279:51100–51106. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Martín A, Odajima J, Hunt SL, Dubus P,
Ortega S, Malumbres M and Barbacid M: Cdk2 is dispensable for cell
cycle inhibition and tumor suppression mediated by p27(Kip1) and
p21(Cip1). Cancer Cell. 7:591–598. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chu I, Sun J, Arnaout A, Kahn H, Narod S,
Sun P, Tan CK, Hengst L and Slingerland J: p27 phosphorylation by
Src regulates inhibition of cyclin E-Cdk2. Cell. 128:281–294. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
James MK, Ray A, Leznova D and Blain SW:
Differential modification of p27Kip1 controls its cyclin D-cdk4
inhibitory activity. Mol Cell Biol. 28:498–510. 2008. View Article : Google Scholar :
|
|
31
|
Guan H, Chen C, Zhu L, Cui C, Guo Y, Fu M,
Wang L and Tang Q: Indole-3-carbinol blocks platelet-derived growth
factor-stimulated vascular smooth muscle cell function and reduces
neointima formation in vivo. J Nutr Biochem. 24:62–69. 2013.
View Article : Google Scholar
|
|
32
|
Hwang SL and Yen GC: Modulation of Akt,
JNK, and p38 activation is involved in citrus flavonoid-mediated
cytoprotection of PC12 cells challenged by hydrogen peroxide. J
Agric Food Chem. 57:2576–2582. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jin YR, Han XH, Zhang YH, Lee JJ, Lim Y,
Kim TJ, Yoo HS and Yun YP: Hesperetin, a bioflavonoid, inhibits rat
aortic vascular smooth muscle cells proliferation by arresting cell
cycle. J Cell Biochem. 104:1–14. 2008. View Article : Google Scholar
|
|
34
|
Diehl JA, Cheng M, Roussel MF and Sherr
CJ: Glycogen synthase kinase-3beta regulates cyclin D1 proteolysis
and subcellular localization. Genes Dev. 12:3499–3511. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pontano LL and Diehl JA: DNA
damage-dependent cyclin D1 proteolysis: GSK3beta holds the smoking
gun. Cell Cycle. 8:824–827. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Malumbres M and Barbacid M: To cycle or
not to cycle: A critical decision in cancer. Nat Rev Cancer.
1:222–231. 2001. View
Article : Google Scholar
|
|
37
|
Yin M, Tian S, Huang X, Huang Y and Jiang
M: Role and mechanism of tissue plasminogen activator in venous
wall fibrosis remodeling after deep venous thrombosis via the
glycogen synthase kinase-3 beta signaling pathway. J Surg Res.
184:1182–1195. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tseng AS, Engel FB and Keating MT: The
GSK-3 inhibitor BIO promotes proliferation in mammalian
cardiomyocytes. Chem Biol. 13:957–963. 2006. View Article : Google Scholar : PubMed/NCBI
|