Introduction
For many years, cancer was predominantly considered
to be a cell-autonomous disease. However, studies on the tumor
microenvironment have given rise to the emerging concept that tumor
progression depends on a dense network of interactions among cancer
cells and the surrounding stroma, including different types of
immune cells (1). For glioblastomas
(GBMs), high malignant brain tumors with a median survival of
<15 months despite multimodal therapy (2) tumor-associated macrophages and
microglia (TAMs) are the predominant infiltrating immune cell
population. This population is characterized by
CD11b+/CD45dim (microglia) and
CD11b+/CD45high (macrophages) phenotypes and
accounts for 13–34% (microglia) and 4.2–12% (macrophages) of the
tumor cell mass in experimental gliomas (3). There is compelling evidence that TAMs
are involved in creating a microenvironment that favors glioma
growth (4–6). They are regarded as M2-macrophages,
which exhibit anti-inflammatory properties as they show elevated
expression of the mannose (CD204/206) and scavenger receptor
(CD163) as well as an enhanced expression and secretion of
immunosuppressive cytokines, such as transforming growth factor
(TGF)-β1 and IL-10 (7). In
contrast, the classically activated or M1-polarized macrophages are
characterized by a pro-inflammatory phenotype, including enhanced
expression and release of IL-1β, tumor necrosis factor (TNF)-α,
interleukin (IL)-6 or IL-12 along with the ability to induce
Th1-mediated immune responses (7).
However, the precise molecular mechanisms underlying the phenomena
of tumor-progression facilitated by TAMs, remain unclear. Since it
is known that TAMs could be recruited to sites of gliomas by an
interplay of CCL2/MCP-1 (constitutively produced by glioma cells)
and its receptor CCR2 (expressed on TAMs) (8,9) and by
a CCL7/MCP-3- CCR1/CCR2/CCR3- crosstalk (10), it seems that small chemotactic
cytokines, known as chemokines, are important key players in GBM
progression by facilitating the infiltration of TAMs into glioma
tissues. In accordance with this, we have previously shown that
CX3CL1 promotes recruitment of TAMs into GBM and enhances
expression of the matrix metalloproteases (MMPs) 2, 9 and 14 in
these tumor cells (11). To date,
there are no data available regarding the expression of the
transmembrane chemokine CXCL16 and its receptor CXCR6 and regarding
the expression of CXCL12 with both its receptors CXCR4 and CXCR7 on
TAMs freshly isolated from GBM. Since we and others showed that
both CXCL16 and CXCL12 and their respective receptors are involved
in GBM progression (12–17), the elucidation of these
chemokine/-receptors on TAMs will help to unravel the role of TAMs
in glioma and may promote the design of more effective treatment
options for these tumors.
Therefore, the present study demonstrated the
expression profile of CX3CL1/CX3CR1, CXCL16/CXCR6 and
CXCL12/CXCR4/CXCR7 in both freshly isolated human GBM-associated
macrophages/microglia and in in vitro generated M1- and
M2-macrophages and related these results to the M1- and M2-marker
expression profile of the different macrophage populations.
Materials and methods
CD11b MACS separation procedure for TAM
isolation
Twenty different freshly obtained human GBM samples
were used for CD11b MACS separation technology as previously
described (11). Therefore,
single-cell suspensions of 400 mg tumor tissue were generated using
the Neural Dissociation Kit (T), and up to 2×107 cells
were immediately labeled with CD11b MicroBeads (Miltenyi Biotec
GmbH, Bergisch Gladbach, Germany) and separated using MACS LS
columns according to the manufacturer’s instructions. Then, the
identity of freshly isolated CD11b positive TAMs was confirmed by
immunofluorescence using a monoclonal mouse anti-human CD11b
FITC-labeled antibody (Miltenyi Biotec GmbH) according to the
manufacturer’s instructions (10 μl undiluted CD11b-FITC for up to
107 separated cells). Additionally, mRNA expression
levels of glial fibrillary acidic protein (GFAP; glioma-cell
specific) and ionized calcium-binding adapter molecule (Iba-1;
monocyte/microglia-specific) of isolated human TAM-enriched
fractions and non-TAM fractions were evaluated by qPCR (as
described below). Only TAM preparations which were CD11b positive
compared to corresponding non-TAM fractions and exhibited high
Iba-1 mRNA expression were used for further analysis. Materials
were obtained in accordance with the Helsinki Declaration of 1975
and with approval of the Ethics Committee of the University of
Kiel.
Generation of M1- and M2-macrophages
Macrophages were generated from monocytes from
healthy adult blood donors obtained from the Research Center
Borstel, Germany. Informed consent was obtained from all donors.
Monocytes (15×106; ~95% purity) generated from human
PBMCs by counterflow centrifugation (elutriation) (18) were cultured in teflon-coated bags
(Süd-Laborbedarf, Gauting, Germany) in RPMI-1640 medium containing
1% L-glutamine (both from PAA Laboratories, Cölbe, Germany), 10%
FCS and 100 μg/ml penicillin/streptomycin (both from Biochrom,
Berlin, Germany) in the presence of either 50 ng/ml M-CSF or GM-CSF
(BioLegend, Fell, Germany) for 7 days. To detach macrophages, bags
were placed on ice for 1 h and cells were resuspended in PBS (PAA
Laboratories). The phenotype of macrophages was characterized
following established protocols (19,20)
and as recently described (21).
Quantitative real-time RT-PCR (qPCR)
Twenty different TAM-enriched and non-TAM fractions
as well as five individual in vitro generated M1- and
M2-macrophage preparations were used. Total RNA from these cells
(>1×103) was purified with the PicoPure RNA Isolation
kit (MDS Analytical Technologies, Sunnyvale, CA, USA) according to
the manufacturer’s instructions. The RNA samples were treated with
RNase-free DNase (1 U/μl) (Promega, Madison, WI, USA), and reverse
transcribed by RevertAid™ H Minus M-MuLV Reverse Transcriptase (200
U/μl) (Fermentas, Vilnius, Lithuania) as previously described
(11,15). Quantitative PCR was performed in
triplicate using a total reaction volume of 20 μl, containing 1 μl
of 20X Assays-on-Demand™ Gene Expression Assay Mix (Iba-1,
Hs_00610419_g1; GFAP, Hs_00157674_m1; CX3CL1, Hs_00171086_m1;
CX3CR1, Hs_00365842_m1; CXCL12, Hs_00171022_m1; CXCR4,
Hs_00237052_m1; CXCR7, Hs_00664172_m1; CXCL16, Hs_00222859_m1;
CXCR6, Hs_00174843_m1; CD163, Hs_00195682_m1; IL-10,
Hs_00961619_m1; TGF-β1, Hs_00171257_m1; TNFα, Hs_00174128_m1;
IL-1β, Hs_01555410_m1 and IL-6, Hs_00985639_m1; Applied Biosystems,
Foster City, CA, USA), 10 μl of 2X TaqMan Universal PCR Master Mix
and 100 or 10 ng of cDNA template (diluted in RNase-free water to 9
μl). After 2 min at 50°C and 10 min at 95°C, 40 cycles of 15 sec at
95°C and 1 min at 60°C were performed. Glyceraldehyde-3-phosphate
dehydrogenase (GAPDH, Hs99999905_m1; Applied Biosystems) mRNA was
amplified in each sample as internal positive control. Each plate
included at least three ‘no template controls (NTC)’. The reaction
was carried out with the MyiQ™ Single-Color Real-time PCR Detection
System (Bio-Rad, Munich, Germany) and fluorescence data were
converted into CT measurements. ΔCT values of
each sample were calculated as: CTgene of interest −
CTGAPDH. Relative gene expression was calculated with
2(normalized CT non-stimulated − normalized CT
stimulated) = n-fold of control. ΔCT = 3.33
corresponds to one order of magnitude. Low ΔCT values
indicate high expression.
Results
TAM enrichment and characterization
To obtain further insight into the expression of M1-
and M2-marker as well as chemokines and their receptors in human
TAMs, we isolated this cell type from 20 different fresh solid
human GBM samples using well-established CD11b MACS separation
technology (11). As fresh human
GBM samples were mostly of small size (max. 400 mg tumor weight),
CD11b separation technology had to be performed in micro format and
analytical methods were essentially limited to qPCR.
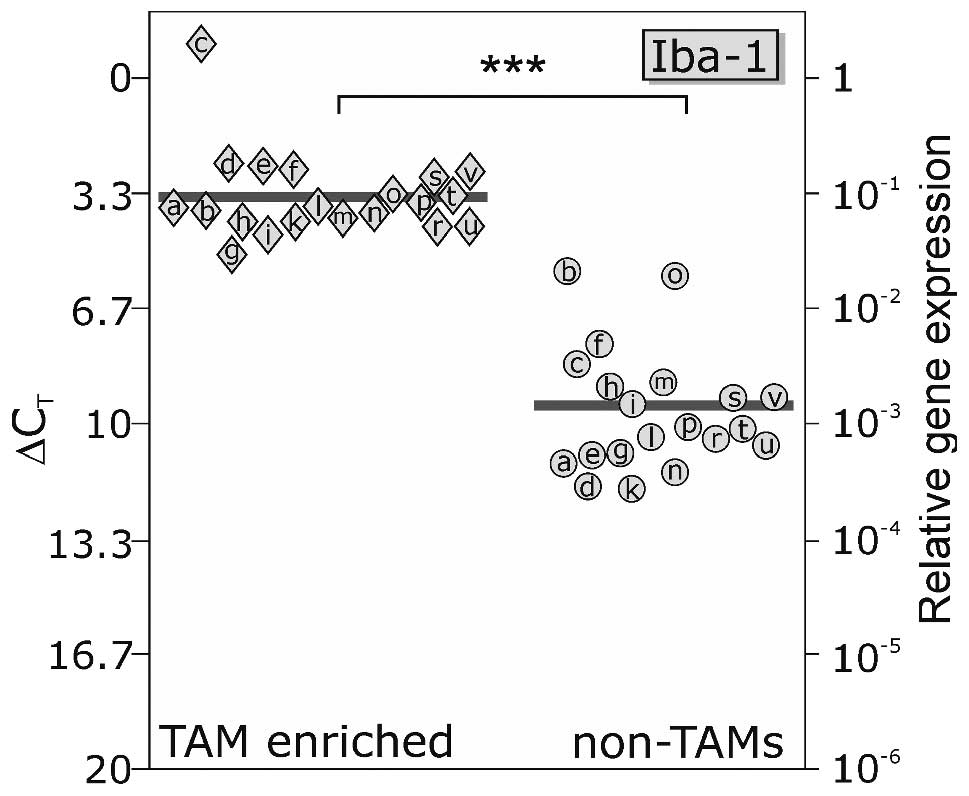
Fig. 1 shows qPCR
results of enriched human TAM and non-TAM fractions measured for
expression of the microglial/macrophage-specific molecule Iba-1.
Iba-1 mRNA expression differed only slightly between individual
TAM-enriched fractions (mean ΔCT value, 3.40), and was
clearly detected at higher levels in TAM-enriched vs. non-TAM
fractions (mean ΔCT value, 9.60). Since a ΔCT
difference of 3.33 corresponds to one order of magnitude, for Iba-1
a 75.0-fold higher expression in TAM-enriched vs. non-TAM fractions
was measured (p<0.001), indicating an accurate isolation
procedure.
We investigated the M1- and M2-marker expression
profile of freshly isolated TAM-enriched fractions and observed
that both M1- and M2-markers were detectable in TAMs but with
different expression intensities (Fig.
2). Regarding M1-specific markers, IL-1β and TNFα were found at
clearly lower levels in TAM-enriched fractions in relation to in
vitro polarized M1-macrophages, whereas IL-6 mRNA was expressed
in approximately similar amounts (Fig.
2, top). In detail, mean ΔCT value for IL-1β was
0.056 in TAMs (vs. −2.09 in M1-polarized macrophages), mean
ΔCT value for TNFα was 6.44 in TAMs (vs. 2.11 in
M1-polarized macrophages), and mean ΔCT value for IL-6
was 4.78 in TAMs (vs. 4.14 in M1-polarized macrophages). Thus, the
mean TAM-enriched fractions were characterized by a 4- and 20-fold
lower expression of IL-1β and TNFα, respectively, but exhibited
somewhat similar IL-6 expression amounts compared to M1-polarized
macrophages.
For M2-specific markers, CD163 and TGFβ were
transcribed in lower amounts in TAM-enriched fractions in
comparison to in vitro M2-polarized macrophages, whereas
IL-10 showed somewhat similar transcription amounts in TAM-enriched
fractions and M2-polarized macrophages (Fig. 2, bottom). In detail, mean
ΔCT value for CD163 was 1.01 in TAMs (vs. −2.60 in
M2-polarized macrophages), mean ΔCT value for TGFβ was
3.08 in TAMs (vs. −0.52 in M2-polarized macrophages), and mean
ΔCT value for IL-10 was 5.64 in TAMs (vs. 5.09 in
M2-polarized macrophages). Thus, TAMs were characterized by a
12-fold lower mean expression of both CD163 and TGFβ, but nearly
equal expression of IL-10 compared to M2-polarized macrophages.
Chemokine expression profile of
TAM-enriched fractions
The expression patterns of the two transmembrane
chemokines CXCL16 and CX3CL1 with their corresponding receptors
CXCR6 and CX3CR1 as well as the expression of CXCL12 with both its
known receptors CXCR4 and CXCR7 were determined in TAM-enriched
fractions as well as M1- and M2-polarized macrophages (Fig. 3).
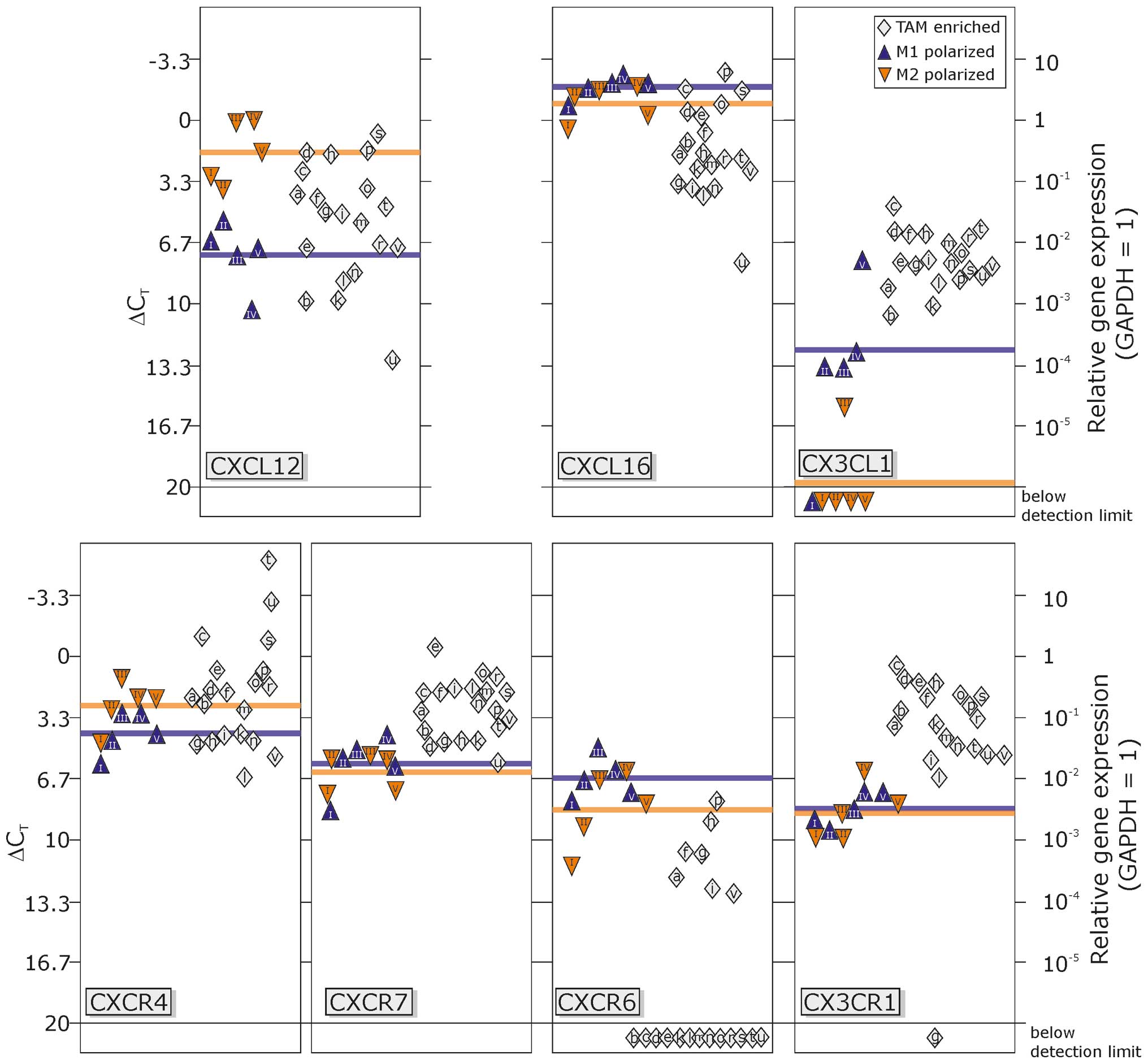 | Figure 3Transcription levels of chemokines
and their receptors in TAM-enriched fractions in comparison to M1-
or M2-polarized macrophages. The chemokine CXCL12 was expressed at
moderate levels in M1-polarized macrophages, but was ~10-fold
higher in M2-polarized macrophages. The transcription in
TAM-enriched fractions showed considerable variation, between high
mRNA levels comparable to M2-macrophages and moderate levels as in
M1-macrophages. The expression of the two CXCL12 receptors, CXCR4
and CXCR7, was very similar in M1- and M2-polarized macrophages; in
TAM-enriched samples, transcription of CXCR4 was comparable to both
polarized macrophage subsets, but slightly elevated for CXCR7. In
both in vitro polarized macrophages, expression of the
chemokine CXCL16 was very high with only slight variations, also
between M1- and M2-macrophages. In contrast, in enriched TAMs,
CXCL16 expression was clearly lower, but still at high expression
levels. The only known receptor for CXCL16, CXCR6 was expressed at
moderate levels in polarized macrophages, but only rarely found in
TAMs (7/20 samples). Expression of CX3CL1 (fractalkine) was low in
M1-polarized macrophages, nearly absent in M2-polarized
macrophages, but detectable in TAM-enriched fractions. In contrast,
the fractalkine receptor CX3CR1 was moderately found in M1- and
M2-polarized macrophages and even ~10-fold higher in TAM-enriched
cell fractions. TAM, tumor-associated macrophages and
microglia. |
In general, all investigated chemokines and
receptors were transcribed at moderate to high levels in
TAM-enriched fractions (except for CXCR6) and M1-/M2-polarized
macrophages (except for CX3CL1). Analysis of CXCL12 and its
receptors CXCR4 and CXCR7 (left part of Fig. 3) revealed that CXCL12 transcription
was ~48-fold higher in M2-polarized macrophages compared to
M1-polarized macrophages (mean ΔCT value, 1.75 vs.
7.34), and the mean transcription level of TAM-enriched fractions
ranged between these two pure macrophage populations (mean
ΔCT 5.58) with variations of the single preparations
ranging from M2- to M1-macrophage levels. The receptors CXCR4 and
CXCR7 were almost equally expressed in M1- and M2-macrophages [mean
ΔCT values, 4.19 and 2.68 (CXCR4), 5.86 and 6.31
(CXCR7)], and in TAM-enriched fractions CXCR4 was despite some
outliers to higher values in this range (mean ΔCT value,
2.03), whereas mean CXCR7 expression (mean ΔCT value,
2.84) was ~10-fold higher compared to both M1- and
M2-macrophages.
Regarding the expression of CXCL16 and CXCR6, M1-
and M2-macrophages both exhibited a very high expression of CXCL16
(mean ΔCT values, −1.82 and −0.90), while TMA-enriched
fractions showed ~5–10-fold lower levels (mean ΔCT
value, 1.64). Markedly, this expression amount was even higher than
expression levels in total GBM samples previously reported by
Hattermann et al (13). In
contrast, the corresponding receptor CXCR6 was rarely detectable in
TAM-enriched fractions although it was present in all M1- and
M2-polarized macrophage preparations (mean ΔCT values,
6.64 and 8.36).
Although expression of CX3CL1, the ligand of CX3CR1,
was quite low in M1-macrophages (mean ΔCT value, 11.80),
and mostly absent in M2-macrophages, it was elevated (~17-fold
compared to M1) in TAM-enriched fractions (mean ΔCT
value, 7.70) but not as prominent as expression of CXCL12 and
CXCL16 (as glioma cells highly produce both chemokines, this could,
however, be caused by a certain contamination). The receptor CX3CR1
was almost similarly expressed in M1- and M2-polarized macrophages
(mean ΔCT values, 9.19 and 8.74), and markedly higher
(55- and 40-fold) in TAM-enriched fractions (mean ΔCT
value, 3.42).
In summary, as shown in Fig. 3, TAM-enriched fractions represent
quite homogenous populations without considerable variations for
the expression of CXCR7, CXCL16 and CXCR6, CX3CL1 and CX3CR1. In
the case of CXCR7, CX3CL1 and CX3CR1, the mean expression levels
were higher than those of M1- and M2-macrophages, while the
expression of CXCL16 and CXCR6 was lower. For both CXCL12 and
CXCR4, the transcription levels varied between expression amounts
of M1- and M2-polarized macrophages (with some outliers for CXCR4
with very high expression levels) and displayed a very
heterogeneous expression pattern with greater differences between
single TAM-enriched fractions.
Discussion
Since tumor-associated macrophages and microglia
(TAMs) create a microenvironment that favors glioma growth, an
understanding of the molecular pathways and signaling molecules
affecting gene expression which lead to the altered phenotype of
glioma TAMs is of considerable importance. Thus, several
investigations focused on the role of TAMs in gliomas and sought to
elucidate exactly how the interplay between TAMs and tumor cells is
regulated (4,22,23).
It becomes clear that pro-tumor functions of TAMs are derived from
their ability e.g. to regulate angiogenic programming and to
provide soluble factors to malignant cells favoring proliferation,
survival and invasion (23).
Nevertheless, these diverse activities depend on the polarization
state of TAMs that seems to be, beside others, regulated by
cytokines and chemokines (23).
Chemokines are a family of peptide mediators that
play essential roles in cellular migration and intercellular
communication and were originally discovered as inducible cytokines
facilitating the recruitment of specific leukocyte subsets.
However, it is now well accepted that their role is not limited to
inflammatory reactions, but extends to various other physiological
and pathophysiological responses (24,25).
Chemokine and chemokine receptors are upregulated in a variety of
human cancers suggesting that they play multifaceted roles in
malignancy (26–29) and TAMs invasion and gene expression
pattern is known to be clearly regulated by different
chemokine/-receptors (11,30). Thus, here, we analyzed the
expression of CXCL16, CXCR6, CX3CL1, CX3CR1, CXCL12, CXCR4 and
CXCR7 in freshly isolated human GBM-associated
macrophages/microglia and in in vitro generated M1- and
M2-macrophages and related these results to the M1- and M2-marker
expression profile of these different cell types.
Initially, we showed that freshly isolated TAMs of
human GBMs were characterized by both M1- and M2-specific markers.
In detail, in comparison to M1-polarized macrophages, TAMs showed
reduced IL-1β and TNF-α expression, whereas IL-6 mRNA level was
somewhat similar. Additionally, in relation to M2-polarized
macrophages, CD163 and TGFβ were found in lower amounts, whereas
IL-10 was expressed at the same levels. Therefore, it seems that
GBM-associated macrophages/microglia share properties of both
‘end-differentiation-types’ of M1- and M2-polarized macrophages.
These results are in accordance with previous findings
demonstrating that TAMs summarize a number of functions rendered by
M2-polarized macrophages such as tuning of inflammatory responses
and adaptive angiogenesis, tissue remodeling and repair and
promoting of angiogenesis (31).
Nevertheless, it was reported that TAMs also express IFN-inducible
chemokines (32). As described by
the authors themselves, a tight distinction between M1- and
M2-macrophages does not fully represent the continuum of functional
states that macrophages can express and is rather a simplified view
(31). In this context, TAMs of GBM
tissues seem to exhibit more functions of M2-polarized macrophages
concomitantly preserving properties of M1-characteristics as has
recently been shown by us for TAMs from pancreatic cancer patients
(33).
Compared to M1- and M2-polarized macrophages,
GBM-associated macrophages/microglia were characterized by a unique
chemokine-specific profile. In detail, for CXCR7, CXCL16 and CXCR6
expression amounts of these chemokines were below those of M1- and
M2-polarized macrophages; for CX3CL1 and CX3CR1 expression amounts
found in TAMs were higher than those of different polarized
macrophages; and for CXCL12 and CXCR4 expression amounts
corresponded to those observed for M1- and M2-polarized
macrophages. Collectively, CXCL16, CXCL12, CXCR4 and CXCR7 seem to
be highly expressed, whereas CX3CL1, CX3CR1 and CXCR6 were lower
transcribed in all investigated macrophage populations.
Since chemokines play a pivotal role in tumor
progression, not only in GBMs but also in many other tumor types,
these results are notable. For example, it has been shown that TAMs
could be recruited to tumor cells by the expression of CCL2/MCP-1,
CCL57RANTES and CXCL1 as products of tumor or stromal cells
(34–36). Especially for CCL2, the level of
tumor-derived CCL2 correlates with amounts of TAMs in several types
of adenocarcinoma (37,38). Additionally, it is well accepted
that the chemokine ligand-receptor pair CXCL12/CXCR4 is implicated
in neoangiogenesis (31,39), and CXCR4 is selectively upregulated
in TAMs tumor and stromal cells (40). Moreover, TAMs can release chemokines
that preferentially attract T cell subsets devoid of cytotoxic
functions by release of, among others, CCL18, CCL17 and CCL22
(31,38,41).
However, high levels of Th1 chemokines such as CXCL9 and CXCL10 are
also expressed by TAMs which suggested deviation from the typical
M2 features of TAMs (42,43).
For GBMs, we and others have shown that the
chemokine/-receptor pairs CXCL12/CXCR4/CXCR7, CXCL16/CXCR6 and
CX3CL1/CX3CR1 are clearly involved in tumor progression (11–15).
Here, we showed that GBM-associated macrophages/microglia are also
characterized by expression of these chemokine/-receptor pairs
indicating a pivotal role of this expression profile in TAM biology
in gliomas. This aspect becomes more important since a clear
tumor-promoting role of TAMs in glioma progression has been
demonstrated in a recent report by Pyonteck et al (44). Her group showed that the inhibition
of the colony-stimulating factor-1 in a GBM model significantly
increased survival and suppressed growth of established tumors.
Moreover, under these conditions, expression of M2-markers
decreased in TAMs, which is consistent with their impaired
tumor-promoting functions. In line with these results, Pong et
al (45) demonstrated that F11R
expression, a microglia surface marker, correlates positively with
glioma malignancy grade and negatively with patient survival
independent of the GBM molecular subtype. Additionally, Ye et
al (46) stated that
tumor-associated microglia/macrophages enhance the invasion of
glioma stem-like cells via the TGF-β1 signaling pathway.
In summary, TAMs are important mediators in glioma
progression and one aspect of this progression is the chemokine
profile of these high malignant brain tumors. Here, we showed for
the first time that freshly isolated human GBM-associated TAMs
share properties of pro- and anti-inflammatory macrophages and are
characterized by an abundant expression of the
chemokines/-receptors CXCL12/CXCR4/CXCR7, CXCL16/CXCR6 and
CX3CL1/CX3CR1 underscoring the potential of these
chemokines/-receptor pairs as possible therapeutic targets.
Acknowledgements
The authors thank B. Rehmke, F. Ebrahim and J.
Krause for their technical assistance. This study was supported by
grants from the University of Kiel, the ‘Deutsche
Forschungsgemeinschaft’ (HE3400/5-1 and ME758/10-1) and the BMBF
(The PopGen 2.0 Network (P2N); support code: 01EY1103).
Abbreviations:
|
GAPDH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
|
GBM
|
glioblastoma
|
|
GFAP
|
glial fibrillary acidic protein
|
|
TAM
|
tumor-associated macrophages and
microglia
|
|
Iba-1
|
ionized calcium-binding adapter
molecule
|
References
|
1
|
Elinav E, Nowarski R, Thaiss CA, Hu B, Jin
C and Flavell RA: Inflammation-induced cancer: crosstalk between
tumours, immune cells and microorganisms. Nat Rev Cancer.
13:759–771. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ohgaki H and Kleihues P: Epidemiology and
ethiology of gliomas. Acta Neuropathol. 109:93–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Badie B and Schartner JM: Flow cytometric
characterization of tumor-associated macrophages in experimental
gliomas. Neurosurgery. 46:957–961. 2000.PubMed/NCBI
|
|
4
|
Li W and Graber MB: The molecular profile
of microglia under the influence of glioma. Neuro Oncol.
14:958–978. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Markovic DS, Glass R, Synowitz M, Rooijen
Nv and Kettenmann H: Microglia stimulate the invasiveness of glioma
cells by increasing the activity of metalloprotease-2. J
Neuropathol Exp Neurol. 64:754–762. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhai H, Heppner FL and Tsirka SE:
Microglia/macrophages promote glioma progression. Glia. 59:472–485.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mantovani A, Germano G, Marchesi F,
Locatelli M and Biswas SK: Cancer-promoting tumor-associated
macrophages: new vistas and open questions. Eur J Immunol.
41:2522–2525. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kielian T, van Rooijen N and Hickey WF:
MCP-1 expression in CNS-1 astrocytoma cells: implications for
macrophage infiltration into tumors in vivo. J Neurooncol. 56:1–12.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Galasso JM, Stegman LD, Blaivas M,
Harrison JK, Ross BD and Silverstein FS: Experimental gliosarcoma
induces chemokine receptor expression in rat brain. Exp Neurol.
161:85–95. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Okada M, Saio M, Kito Y, Ohe N, Yano H,
Yoshimura S, Iwama T and Takami T: Tumor-associated
macrophage/microglia infiltration in human gliomas is correlated
with MCP-3, but not MCP-1. Int J Oncol. 34:1621–1627.
2009.PubMed/NCBI
|
|
11
|
Held-Feindt J, Hattermann K, Müerköster
SS, Wedderkopp H, Knerlich-Lukoschus F, Ungefroren H, Mehdorn HM
and Mentlein R: CX3CR1 promotes recruitment of human
glioma-infiltrating microglia/macrophages (GIMs). Exp Cell Res.
316:1553–1566. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hattermann K and Mentlein R: An infernal
trio: the chemokine CXCL12 and its receptors CXCR4 and CXCR7 in
tumor biology. Ann Anat. 195:103–110. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hattermann K, Held-Feindt J, Ludwig A and
Mentlein R: The CXCL16-CXCR6 axis in glial tumors. J Neuroimmunol.
260:47–54. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hattermann K1, Held-Feindt J, Lucius R,
Müerköster SS, Penfold ME, Schall TJ and Mentlein R: The chemokine
receptor CXCR7 is highly expressed in human glioma cells and
mediates antiapoptotic effects. Cancer Res. 70:3299–3308. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hattermann K, Ludwig A, Gieselmann V,
Held-Feindt J and Mentlein R: The chemokine CXCL16 induces
migration and invasion of glial precursor cells via its receptor
CXCR6. Mol Cell Neurosci. 39:133–141. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Domanska UM, Kruizinga RC, den Dunnen WF,
Timmer-Bosscha H, de Vries EG and Walenkamp AM: The chemokine
network, a newly discovered target in high grade gliomas. Crit Rev
Oncol Hematol. 79:154–163. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sciumè G, Santoni A and Bernardini G:
Chemokines and glioma: invasion and more. J Neuroimmunol. 224:8–12.
2010.PubMed/NCBI
|
|
18
|
Grage-Griebenow E, Zwatzky R, Kahlert H,
Brade L, Flad H and Ernst E: Identification of a novel dendritic
cell-like subset of CD64+/CD16+ blood
monocytes. Eur J Immunol. 31:48–56. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Krausgruber T, Blazek K, Smallie T,
Alzabin S, Lockstone H, Sahgal N, Hussell T, Feldmann M and Udalova
IA: IRF5 promotes inflammatory macrophages polarization and
TH1–TH17 responses. Nat Immunol. 12:231–238.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Verreck FA1, de Boer T, Langenberg DM, van
der Zanden L and Ottenhoff TH: Phenotypic and functional profiling
of human proinflammatory type-1 and anti-inflammatory type-2
macrophages in response to microbial antigens and IFN-γ- and
CD40L-mediated costimulation. J Leukoc Biol. 79:285–292.
2006.PubMed/NCBI
|
|
21
|
Schäfer H, Struck B, Feldmann EM, Bergmann
F, Grage-Griebenow E, Geismann C, Ehlers S, Altevogt P and Sebens
S: TGF-β1-dependent L1CAM expression has an essential role in
macrophage-induced apoptosis resistance and cell migration of human
intestinal epithelial cells. Oncogene. 32:180–189. 2013.
|
|
22
|
Kennedy BC, Showers CR, Anderson DE,
Anderson L, Canoll P, Bruce JN and Anderson RCE: Tumor-associated
macrophages in glioma: friend or foe? J Oncol. 2013:4869122013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ruffel B, Affara NI and Coussens LM:
Differential macrophage programming in the tumor microenvironment.
Trends Immunol. 33:119–126. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Miller RJ, Rostene W, Apartis E, Banisadr
G, Biber K, Milligan ED, White FA and Zhang J: Chemokine action in
the nervous system. J Neurosci. 28:11792–11795. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vandercappellen J, Van Damme J and Struyf
S: The role of CXC chemokines and their receptors in cancer. Cancer
Lett. 267:226–244. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kakinuma T and Hwang ST: Chemokines,
chemokine receptors, and cancer metastasis. J Leukoc Biol.
79:639–651. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zlotnik A: Chemokines and cancer. Int J
Cancer. 119:2026–2029. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu H, Ouyang W and Huang C: Inflammation,
a key event in cancer development. Mol Cancer Res. 4:221–233. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ben-Baruch A: The multifaceted roles of
chemokines in malignancy. Cancer Metastasis Rev. 25:357–371. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Platten M, Kretz A, Naumann U, Aulwurm S,
Egashira K, Isenmann S and Weller M: Monocyte chemoattractant
protein-1 increases microglial infiltration and aggressiveness of
gliomas. Ann Neurol. 54:388–392. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Allavena P, Sica A, Solinas G, Porta C and
Mantovani A: The inflammatory micro-environment in tumor
progression: the role of tumor-associated macrophages. Crit Rev
Oncol Hematol. 66:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sica A, Schioppa T, Mantovani A and
Allavena P: Tumour-associated macrophages are a distinct M2
polarised population promoting tumour progression: potential
targets of anti-cancer therapy. Eur J Cancer. 42:717–727. 2006.
View Article : Google Scholar
|
|
33
|
Helm O, Held-Feindt J, Grage-Griebenow E,
Reiling N, Ungefroren H, Vogel I, Krüger U, Becker T, Ebsen M,
Röcken C, Kabelitz D, Schäfer H and Sebens S: Tumor-associated
macrophages exhibit pro- and anti-inflammatory properties by which
they impact on pancreatic tumorigenesis. Int J Cancer. Jan
23–2014.(Epub ahead of print). View Article : Google Scholar
|
|
34
|
Arenberg DA, Keane MP, DiGiovine B, Kunkel
SL, Strom SR, Burdick MD, Iannettoni MD and Strieter RM: Macrophage
infiltration in human non-small-cell lung cancer: the role of CC
chemokines. Cancer Immunol Immunother. 49:63–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Balkwill F: Cancer and the chemokine
network. Nat Rev Cancer. 4:540–550. 2004. View Article : Google Scholar
|
|
36
|
Mantovani A, Allavena P, Sozzani S, Vecchi
A, Locati M and Sica A: Chemokines in the recruitment and shaping
of the leukocyte infiltrate of tumors. Semin Cancer Biol.
14:155–160. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ueno T, Toi M, Saji H, Muta M, Bando H,
Kuroi K, Koike M, Inadera H and Matsushima K: Significance of
macrophage chemoattractant protein-1 in macrophage recruitment,
angiogenesis, and survival in human breast cancer. Clin Cancer Res.
6:3282–3289. 2000.PubMed/NCBI
|
|
38
|
Monti P, Leone BE, Marchesi F, Balzano G,
Zerbi A, Scaltrini F, Pasquali C, Calori G, Pessi F, Sperti C, Di
Carlo V, Allavena P and Piemonti L: The CC chemokine MCP-1/CCL2 in
pancreatic cancer progression: regulation of expression and
potential mechanisms of antimalignant activity. Cancer Res.
63:7451–7461. 2003.PubMed/NCBI
|
|
39
|
Bachleder RE, Wendt MA and Mercurio AM:
Vascular endothelial growth factor promotes breast carcinoma
invasion in an autocrine manner by regulating the chemokine
receptor CXCR4. Cancer Res. 62:7203–7206. 2002.PubMed/NCBI
|
|
40
|
Schioppa T, Uranchimeg B, Saccani A,
Biswas SK, Doni A, Rapisarda A, Bernasconi S, Saccani S, Nebuloni
M, Vago L, Mantovani A, Melillo G and Sica A: Regulation of the
chemokine receptor CXCR4 by hypoxia. J Exp Med. 198:1391–1402.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Schutyser E, Struyf S, Proost P,
Opdenakker G, Laureys G, Verhasselt B, Peperstraete L, Van de Putte
I, Saccani A, Allavena P, Mantovani A and Van Damme J:
Identification of biologically active chemokine isoforms from
ascetic fluid and elevated levels of CCL18/pulmonary and
activation-regulated chemokine in ovarian carcinoma. J Biol Chem.
277:24584–24593. 2002. View Article : Google Scholar
|
|
42
|
Sica A, Larghi P, Mancino A, Rubino L,
Porta C, Totaro MG, Rimoldi M, Biswas SK, Allavena P and Mantovani
A: Macrophage polarization in tumor progression. Semin Cancer Biol.
18:349–355. 2008. View Article : Google Scholar
|
|
43
|
Biswas SK, Gangi L, Paul S, Schioppa T,
Saccani A, Sironi M, Bottazzi B, Doni A, Vincenzo B, Pasqualini F,
Vago L, Nebuloni M, Mantovani A and Sica A: A distinct and unique
transcriptional program expressed by tumor-associated macrophages
(defective NF-κB and enhanced IRF-3/STAT1 activation). Blood.
107:2112–2122. 2006.PubMed/NCBI
|
|
44
|
Pyonteck SM, Akkari L, Schuhmacher AJ,
Bowman RL, Sevenich L, Quail DF, Olson OC, Quick ML, Huse JT,
Teijeiro V, Setty M, Leslie CS, Oei Y, Pedraza A, Zhang J, Brennan
CW, Sutton JC, Holland EC, Daniel D and Joyce JA: CSF-1R inhibition
alters macrophage polarization and blocks glioma progression. Nat
Med. 19:1264–1272. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pong WW, Walker J, Wylie T, Magrini V, Luo
J, Emnett RJ, Choi J, Cooper ML, Griffith M, Griffith OL, Rubin JB,
Fuller GN, Piwnica-Worms D, Feng X, Hambardzumyan D, DiPersio JF,
Mardis ER and Gutmann DH: F11R is a novel monocyte prognostic
biomarker for malignant glioma. PLoS One. e775712013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ye XZ, Xu SL, Xin YH, Yu SC, Ping YF, Chen
L, Xiao HL, Wang B, Yi L, Wang QL, Jiang XF, Yang L, Zhang P, Qian
C, Cui YH, Zhang X and Bian XW: Tumor-associated
microglia/macrophages enhance the invasion of glioma stem-like
cells via TGF-β1 signaling pathway. J Immunol. 189:444–453.
2012.PubMed/NCBI
|