Introduction
Nasopharyngeal carcinoma (NPC) is a human epithelial
malignancy with high incidence in southern China, particularly, in
the Cantonese population. The Epstein-Barr virus (EBV) was the
first human tumor virus to be identified and has yielded
significant insights into the pathogenesis of cancer. Latent EBV
infection is one of the most important factors for NPC in endemic
areas. NPC is associated with EBV latent infection type II and the
expression of EBV latent membrane protein (LMP) 1, EBV-LMP2A,
EBV-LMP2B, EBV nuclear antigen (EBNA) 1, EBV-encoded RNA (EBER) 1,
EBER2 and the BamHI A rightward transcripts (BARTs)
(1–3). Metastasis to the neck lymph nodes is
the predominant initial symptom in NPC. In addition, recurrence and
distant metastasis after radiotherapy are known to occur in NPC.
Therefore, it is imperative to identify novel diagnostic and
therapeutic approaches.
Loss of heterozygosity (LOH) on chromosomes 13q and
14q are common genetic events, and putative tumor-suppressor genes
located in these regions may be involved in the development of NPC
(4,5). Evidence from LOH studies showed that
NPC-related tumor-suppressor genes may be present at 14q24-q32
(5). LOH on chromosome 14q was
detected in 80% of NPC tumors with high-frequency LOH loci
clustered to 14q11-q13, 14q21-q24 and 14q32 (5). The GALC gene is located at
14q31, has 17 exons and 16 introns of an approximate size of 60 kb
(6). Aberrant methylation in the
promoters of tumor-related genes is closely associated with
epigenetically mediated gene silencing (7–14). DNA
hypermethylation in the promoter region has been reported in NPC
(7–10). Hypermethylation of the 9p21
chromosomal region genes (p14, p15 and p16) has been observed in
the peripheral blood of leukemic patients (11). DNA methylation in cancer-related
genes can be a potential biomarker and therapeutic target for
prostate cancer (12). It has been
reported that p16 promoter methylation is a promising potential
biomarker for the early diagnosis of gastric cancer (13). A potential biomarker panel assessing
the hypermethylation of six gene promoters holds great promise as a
diagnostic test for high-grade (type II) serous ovarian cancer
(14).
Therefore, we aimed to investigate the possible
tumor-suppressor effect of the GALC gene in EBV-associated
NPC, and identified that the GALC gene is downregulated by
promoter hypermethylation in the NPC cell line, CNE-2Z.
Materials and methods
Specimens
The World Health Organization has classified NPC
into different histological types [keratinizing squamous cell
carcinoma (type I), differentiated non-keratinizing carcinoma (type
II) and undifferentiated non-keratinizing carcinoma (type III)]
based on the tumor cell appearance on light microscopy (15). Type II and III NPC are predominantly
EBV-positive.
Specimens of type III NPC and chronic
nasopharyngitis were obtained from patients diagnosed with these
conditions between January 2008 and August 2012 at the Affiliated
Hospital of Guangdong Medical College and Gaozhou People's Hospital
(Guangdong, China). Informed consent was obtained from all the
participants. None of the NPC patients were previously treated with
radiotherapy or chemotherapy. Of the 41 NPC patients whose
specimens were used for immunohistochemical analysis, 7 were at TNM
stage II, 21 were at stage III and 13 were at stage IV. There were
35 men and 6 women in the NPC group, with ages ranging from 31 to
86 years (median age, 54 years; mean age, 52 years). Of the 15
chronic nasopharyngitis patients, 9 were male and 6 were female,
with ages ranging from 15 to 79 years (median age, 47 years; mean
age, 45.9 years).
EBER in situ hybridization
The sections of 41 NPC tumor specimens were dewaxed
in xylene and digested with proteinase K for 16 min. The sections
were then rehydrated in serial graded ethanol washes (70, 95 and
100%) and hybridized for 2 h at 37°C with digoxigenin-labeled EBER
probes (ZSGB-BIO, China). The hybridization products were detected
using anti-digoxigenin-horseradish peroxidase (ZSGB-BIO, Beijing,
China) for 30 min at 37°C, followed by 3 min of incubation in
diaminobenzidine solution for the detection of EBERs. The slides
were counterstained with Mayer's hematoxylin and positive staining
was observed as brown granules at the site of hybridization on
light microscopy. Only specimens with signals within the tumor cell
nuclei were considered to be positive.
Immunohistochemical staining
Tissue samples were fixed in 10% buffered formalin,
dehydrated, embedded in paraffin and cut into 4-μm sections.
The samples were evaluated by two experienced pathologists.
Sections with known high and no GALC expression were used as
positive and negative controls, respectively. The sections
incubated with 0.01 mol/l phosphate-buffered saline (PBS) instead
of primary antibody served as the blank control. A
streptavidin-peroxidase (SP) kit was purchased from Beijing
Zhongshan Golden Bridge Biotechnology, Co. (Beijing, China). Rabbit
anti-human GALC polyclonal antibody was purchased from the
Proteintech Group, Inc. (Chicago, IL, USA) (working dilution,
1:200). Procedures were implemented according to the manufacturer's
instructions.
Evaluation of in situ hybridization and
immunohistochemical staining
For each section, five low magnification fields
(×100) were randomly selected under a microscope, and the
proportions of positively stained cells per field were determined
under higher magnification (×400). The results were expressed as
the mean percentage of positively stained cells in the five fields.
GALC was specifically located in the cytoplasm. The staining
intensity (A) was classified as follows: negative, 0 points; weak,
1 point; moderate, 2 points; and strong, 3 points. The percentage
of positive cells (B) was classified as: 0%, 0 points; 1–25%, 1
point; 26–50%, 2 points; 51–75%, 3 points, and >75%, 4 points.
The product of A and B yielded the final score, which was
classified as follows: 0–1, negative (−) and ≥2, positive (+). The
sections were evaluated by three pathologists who were blinded to
the study design.
Cell culture and drug treatment
The NPC CNE-2Z cell line which has low
differentiation, was obtained from the Department of Pathology,
Guangdong Medical College. The CNE-2Z cells were cultured in
RPMI-1640 with 10% fetal bovine serum (FBS; HyClone, Laboratories,
Logan, UT, USA), 0.1 μM penicillin and 0.1 μM
streptomycin under 5% CO2 at 37°C. For drug treatment,
exponentially growing CNE-2Z cells were seeded at a density of
2×105 cells/well into 6-well plates. The cells were
treated with different doses (0, 5, 10, 20, 40 and 60 μM/l)
of the demethylation agent 5-Aza-2′-deoxycytidine (5-Aza-dC; Sigma,
St. Louis, MO, USA) for 48 h. The cells were collected for the
detection of GALC protein expression using western blotting and
immunofluorescent assay, GALC mRNA expression using reverse
transcriptase-polymerase chain reaction (RT-PCR) assay, GALC
gene promoter methylation using bisulfite sequencing PCR (BSP),
cell proliferation using Cell Counting kit (CCK)-8 assay, and cell
migration using scratch and Transwell migration assays.
Quantitative RT-PCR
Total RNA was extracted from the cultured CNE-2Z
cells using RNAiso Plus (Takara, Japan). Reverse transcription was
performed with the PrimeScript™ RT reagent kit (Takara). The
primers used for the PCR assay were: GALC,
5′-AGGGAACCTCACCATCATCA-3′ (forward) and
5′-GCTGTCAAGGAGCCATAGAGAA-3′ (reverse); and GAPDH,
5′-AGGTCGGAGTCAACGGATTTG-3′ (forward) and
5′-GTGATGGCATGGACTGTGGT-3′ (reverse). PCR was performed in a
LightCycler 480 fluorescent quantization PCR thermal cycler (Roche,
Switzerland). Quantitative RT-PCR was performed with
SYBR® Premix Ex Taq™ II (Takara) at 95°C for 30
sec, followed by 40 cycles of 95°C for 5 sec and 60°C for 30 sec,
after which a melt-curve analysis was performed once from 60 to
95°C, followed by 1 cycle at 50°C for 30 sec. The level of GAPDH
mRNA was used as an internal control. The reactions were performed
in triplicate. The relative amounts of GALC mRNA were
calculated in accordance with the 2−ΔΔCt method.
BSP
Bisulfite modification of genomic DNA converts
unmethylated deoxycytidine to uracil (read as thymidine), however,
methylated deoxycytidine remained unmodified. A 372-bp region
within the promoter (−158 to +214 nucleotides relative to the
transcriptional start site) containing the CpG island was
PCR-amplified from the bisulfite-treated DNA. The PCR fragments
were cloned and sequenced in 10 clones, thereby allowing
identification of the methylation status of each of the 37 specific
CpG sites in the GALC gene. The methylation primers of the
GALC gene promoter region used for the BSP assay were:
5′-TTG(T/C)GTTAAAGTGTTTTATTA GGTGA-3′ (forward) and
5′-CAAC(A/G)CACACAACAAC AAAAACA-3′ (reverse).
Western blot analysis
Total cellular proteins of the harvested CNE-2Z
cells were extracted. The standard western blotting protocol was
applied. The specific primary antibody anti-GALC (working dilution,
1:200; Proteintech) and a horseradish peroxidase-conjugated
secondary antibody (Mai Bio Co., Ltd, Zhejiang, China) were used.
Specific bands were visualized using enhanced chemiluminescence
reagents and recorded on film.
Immunofluorescent staining
CNE-2Z cells were fixed in 4% paraformaldehyde for
15 min at room temperature, blocked and incubated with the primary
antibody anti-GALC (working dilution, 1:200), the secondary
antibody Rhodamine-conjugated goat anti-rabbit IgG (both from
Proteintech) and 4′,6-diamidino-2-phenylindole (DAPI; Sigma). In
each experiment, the immunopositive cells in five randomly selected
fields were counted and 500–1,000 cells were counted in total.
Cell viability
The proliferation of CNE-2Z cells was determined
using the CCK-8 assay, according to the manufacturer's instructions
(Beyotime Institute of Biotechnology, Shanghai, China).
Exponentially growing CNE-2Z cells (4×103 cells/well) in
100 μl medium were seeded in 96-well plates. After 12 h, the
medium in each well was replaced with media containing different
concentrations of 5-Aza-dC (0, 5, 10, 20, 40 and 60 μM/l),
and the plates were incubated for 48 h. The supernatant was
removed, and 200 μl of CCK-8 and RPMI-1640 mixed medium was
added into each well, followed by incubation for 2 h at 37°C in the
dark. The absorbance of CCK-8-containing medium was measured at a
wavelength of 450 nm (reference wavelength, 630 nm) using a
microplate reader (Bio-Rad, Hercules, CA, USA). Cell proliferation
was calculated as: Viability = (ODtest group -
ODblank group)/(ODcontrol group -
ODblank group). The results were presented as mean ±
standard deviation (SD). Three independent experiments, each in
triplicate, were conducted.
Cell migration
In vitro scratch and Transwell chamber assays
were performed to assess the migration of CNE-2Z cells.
Exponentially growing CNE-2Z cells (8×105 cells/well)
were seeded in 6-well plates. When the cells were cultured to 90%
confluency, each well was manually scratched three times with a
200-μl pipette tip, washed with PBS three times and
incubated at 37°C with media containing different concentrations of
5-Aza-dC (0, 20 and 40 μM/l). Cell images were recorded at 0
and 24 h. The distance between two cell edges was analyzed using
Image-Pro software.
The Transwell system (24 wells, 8-μm pore
size with polycarbonate membrane; Corning Costar, NY, USA) not
coated with Matrigel was used for the migration assay. The CNE-2Z
cells were incubated with media containing different concentrations
of 5-Aza-dC (0, 20 and 40 μM/l) at 37°C for 48 h. A total of
4×104 cells were suspended in 100 μl serum-free
medium and were seeded in the upper chambers. Then, 600 μl
RPMI-1640 containing 10% FBS was added to the lower chamber. After
24 h, the cells remaining in the upper chambers were removed with a
cotton swab, while the cells attached to the lower surface were
fixed with methanol and stained with hematoxylin and eosin
(H&E). The cells migrating to the lower side in five randomly
selected fields were counted by light microscopy, and the total
number of migrating cells was determined and analyzed
statistically. Each experiment was performed in triplicate.
Statistical analysis
Statistical analyses were performed using SPSS 17.0.
One-way analysis of variance (ANOVA) was performed to analyze the
data from the CCK-8, scratch and Transwell chamber assays and
RT-PCR. The χ2 and the Fisher's exact tests, and the
logistic regression analysis were used to compare the rates between
the different groups and to test the correlation between two
factors. P<0.05 was considered to indicate a statistically
significant result.
Results
EBER and GALC protein expression in
chronic nasopharyngitis and NPC
A high expression of EBER was detected in the
specimens from the 41 NPC patients. EBER expression was not
correlated with different clinicopathological parameters, such as
age, gender, lymph node metastasis and TNM stage (P>0.05).
EBER and GALC protein expression in the
columnar epithelium of the nasopharyngeal mucosa and in NPC
No EBER expression was found in the columnar
epithelial of the nasopharyngeal mucosa obtained from the 15
individuals in the control group. By contrast, 92.68% of the NPC
specimens from the 41 NPC patients exhibited EBER expression. The
immunohistochemical SP method was used to measure the expression of
GALC protein. GALC protein expression was detected in 24.39% of NPC
specimens, which had a high rate of EBV latent infection. GALC
protein expression was detected in 60.00% of columnar epithelia of
chronic nasopharyngitis specimens with no EBV latent infection. The
difference between these two groups was statistically significant
(P<0.05) (Table I).
 | Table IEBER and GALC protein expression in
the columnar epithelium of the nasopharyngeal mucosa and in
NPC. |
Table I
EBER and GALC protein expression in
the columnar epithelium of the nasopharyngeal mucosa and in
NPC.
| Groups | EBERa
| GALCb
|
|---|
| n | − | +(%) | − | +(%) |
|---|
| Nasopharyngeal | 15 | 15 | 0 (0) | 6 | 9 (60.00) |
| mucosal | | | | | |
| columnar | | | | | |
| epithelium | | | | | |
| NPC cells | 41 | 3 | 38 (92.68) | 31 | 10 (24.39) |
Relationship between GALC protein
expression and clinicopathological parameters of NPC
The GALC protein expression was not
correlated with age, gender and TNM stage (P>0.05) (Table II). GALC protein expression
occurred significantly more frequently in tumors without lymph node
metastasis than in tumors with metastasis (P<0.05) (Table II). Logistic regression showed that
GALC protein expression protected against lymph node metastasis
(P<0.05) (Table III).
| Table IIRelationship between GALC protein
expression and clinicopathological parameters in NPC. |
Table II
Relationship between GALC protein
expression and clinicopathological parameters in NPC.
| Clinicopathological
parameters | GALC
| |
|---|
| n | − | +(%) | P-value |
|---|
| Age (years) | | | | |
| <40 | 5 | 5 | 0 (0) | 0.31 |
| ≥40 | 36 | 26 | 10 (27.77) | |
| Gender | | | | |
| Male | 35 | 26 | 9 (25.71) | 1 |
| Female | 6 | 5 | 1 (16.66) | |
| Lymph node
metastasis | | | | |
| No | 12 | 6 | 6 (50.00) | 0.040a |
| Yes | 29 | 25 | 4 (13.79) | |
| TNM stage | | | | |
| II | 7 | 3 | 4 (57.14) | 0.083 |
| III | 21 | 17 | 4 (19.04) | |
| IV | 13 | 11 | 2 (15.38) | |
| Table IIILogistic regression analysis of GALC
protein expression and lymph node metastasis in NPC (n=41). |
Table III
Logistic regression analysis of GALC
protein expression and lymph node metastasis in NPC (n=41).
| Selected
factor | β | SE | Wald | P-value | Exp (β) | 95% CI |
|---|
| GALC | −1.8326 | 0.7895 | 5.3877 | 0.020 | 0.160 | 0.034–0.752 |
Effect of different 5-Aza-dC
concentrations on GALC gene methylation status of CNE-2Z cells
To confirm that hypermethylation of the 37 CpG sites
in the promoter region (−158 to +214) of the GALC gene
decreased after 5-Aza-dC treatment, CNE-2Z cells were treated with
different concentrations of 5-Aza-dC (0, 5, 10, 20, 40 and 60
μM/l) for 48 h, and the methylation status of the
GALC gene promoter was analyzed using bisulfite genomic
sequencing in the control (0 μM/l) and treated samples
(Fig. 3). In the control sample,
CpG sites were 95.9% methylated, and in the treated samples the
sites were 86.5, 81.9, 50.8, 71.6 and 70.3% methylated at the
concentrations of 5, 10, 20, 40 and 60 μM/l,
respectively.
Effect of different concentrations of
5-Aza-dC on GALC mRNA expression in CNE-2Z cells
To provide evidence for a possible role of DNA
methylation in the regulation of GALC expression, we
analyzed GALC mRNA levels in CNE-2Z cells using RT-PCR in
the control and treated groups (Fig.
4). The results showed that GALC mRNA levels were
significantly higher in the treated groups than those in the
control group (P<0.05). GALC mRNA levels in the 5, 10, 40
and 60 μM/l groups were significantly lower than that in the
20 μM/l group (P<0.01), while the levels in the 5 and 10
μM/l groups were significantly lower than that in the 40
μM/l group (P<0.05). GALC mRNA levels in the 5, 10
and 40 μM/l groups did not significantly differ from that in
the 60 μM/l group (P>0.05).
 | Figure 4GALC mRNA expression detected
using RT-PCR analysis. The control sample and the CNE-2Z cells were
treated with different concentrations of 5-Aza-dC (5, 10, 20, 40
and 60 μM/l) for 48 h, and then subjected to RT-PCR to
analyze the GALC mRNA expression levels. The GALC
mRNA expression levels in the treated samples (20, 40 and 60
μM/l) significantly differed from those in the control
samples. Statistical analysis was performed using one-way ANOVA,
followed by the significant difference (Dunnett's T3) test. Data
are presented as means ± SD, n=3. *P<0.05, treated
groups vs. control. #P<0.01, 5, 10, 40 and 60
μM/l vs. 20 μM/l. &P<0.05, 5 and 10
μM/l vs. 40 μM/l. 5-Aza-dC,
5-Aza-2′-deoxycytidine. |
GALC protein expression in CNE-2Z cells
treated with different concentrations of 5-Aza-dC and detected
using western blotting and immunofluorescence
To provide evidence for a possible role of DNA
methylation in the regulation of GALC expression, we
analyzed GALC protein expression using western blotting and
immunofluorescence analyses before and after treatment with
different concentrations of 5-Aza-dC (5, 10, 20, 40 and 60
μM/l) for 48 h (Fig. 5). The
results showed that GALC protein expression was absent in
the control group but present in the cells treated with 5-Aza-dC.
GALC protein expression was highest in the 20 μM/l
group.
These results indicated a correlation between the
methylation status of the GALC gene and the expression level
of GALC mRNA or protein. Data suggest that promoter
methylation suppresses GALC gene expression, and
5-Aza-dC-induced demethylation of CpG sites upregulates GALC
gene expression in the NPC CNE-2Z cell line.
Effect of different concentrations of
5-Aza-dC on CNE-2Z cell proliferation
CCK-8 was used to examine the proliferation of
CNE-2Z cells treated with different concentrations of 5-Aza-dC for
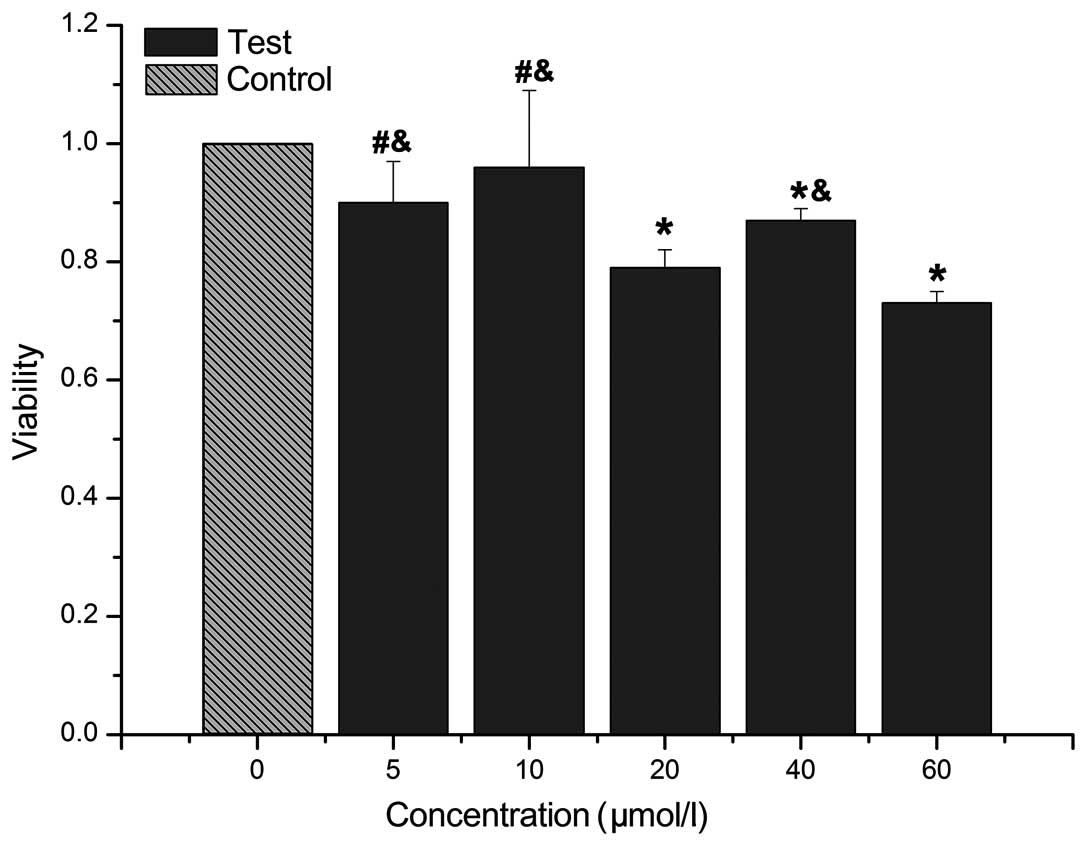
48 h (Fig. 6). The results showed
that cell viability was significantly decrased in the 20, 40 and 60
μM/l groups than in the control group (P<0.05). The cell
viability in the 5 and 10 μM/l groups significantly differed
from that in the 20 μM/l group (P<0.05), while in the 5,
10 and 40 μM/l groups cell viability significantly differed
from that in the 60 μM/l group (P<0.05). The viability in
the 40 and 60 μM/l groups did not significantly differ from
that the in 20 μM/l group (P>0.05).
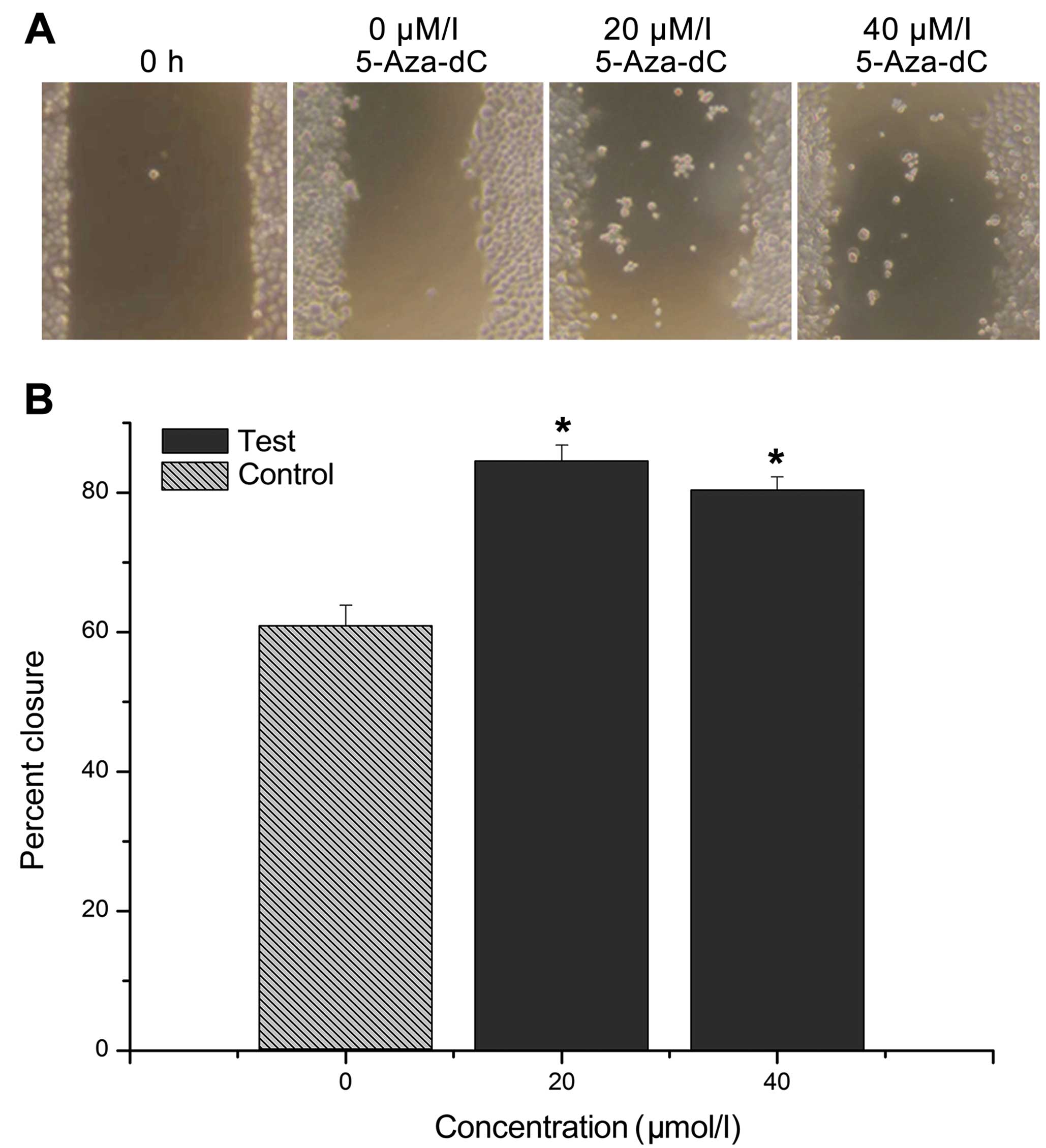
Cell migration and scratch assay
After the CNE-2Z cells were treated with 20 or 40
μM/l 5-Aza-dC for 24 h, the relative cell migration distance
was significantly higher in the treatment group than in the control
group. Moreover, the cell migration ability decreased with an
increasing concentration of 5-Aza-dC, and the differences between
the treatment and control groups were statistically significant
(P<0.01). However, no significant difference was found between
the 20 and 40 μM/l group (P>0.05; Fig. 7).
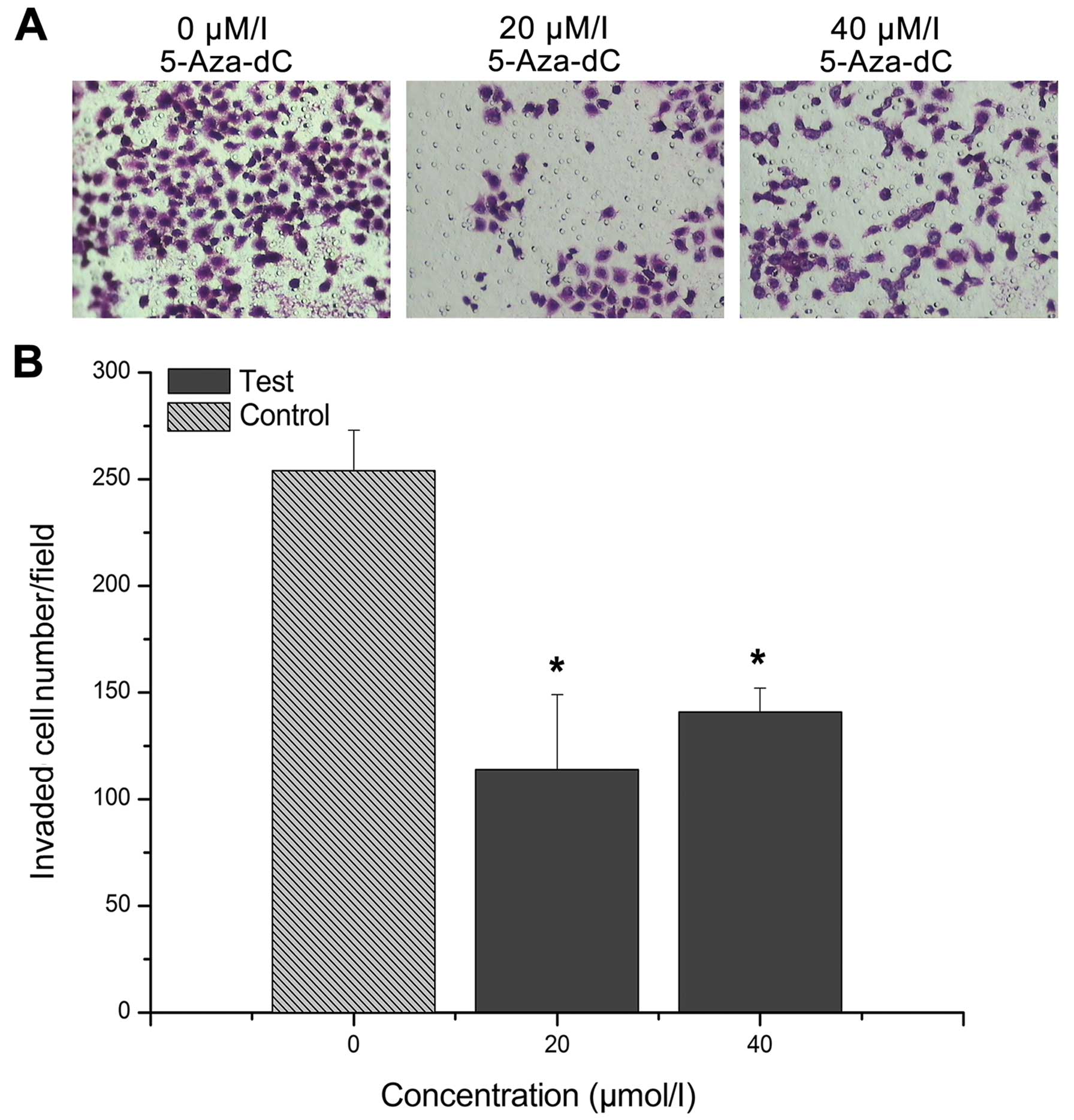
Cell migration and Transwell chamber
assay
After the CNE-2Z cells were treated with 20 or 40
μM/l 5-Aza-dC for 24 h, the number of cells that migrated to
the lower side was significantly lower in the treatment groups than
in the control group (P<0.01). In addition, the number of the
cells that migrated to the lower side was lower in the 20
μM/l group than that in the 40 μM/l group, although
the difference was not statistically significant (P>0.05;
Fig. 8).
Discussion
Latent EBV infection is one of the most important
factors for NPC in endemic areas. The transcription of EBERs is
present in most NPC tumors (1–3). In
the present study, no EBER expression was found in the columnar
epithelial of the nasopharyngeal mucosa obtained from the 15
individuals in the control group. By contrast, 92.68% of the NPC
specimens from the 41 NPC patients exhibited EBER expression. The
immunohistochemical SP method was used to measure the expression of
GALC protein. GALC protein expression was detected in 24.39% of NPC
specimens, which had a high rate of EBV latent infection. GALC
protein expression was detected in 60.00% of columnar epithelial of
chronic nasopharyngitis specimens which had no EBV latent
infection. The difference between these two groups was
statistically significant (P<0.05). Subsequently, the 41 NPC
specimens were divided into a lymph node metastasis and non-lymph
node metastasis groups according to the TNM stage. Statistical
analyses showed that the rates of the GALC protein expression were
13.79 and 50.00%, respectively, in these two groups and the
difference was statistically significant (P<0.05). Logistic
regression showed that the expression of GALC protein was a
protective factor against lymph node metastasis of NPC (P<0.05).
Compared with the normal columnar epithelial of the nasopharyngeal
mucosa, the NPC cells showed a decreased expression of GALC
protein. The results of the present study were in accordance with
the results reported by Görögh for laryngeal squamous cell
carcinoma as compared with the corresponding normal mucosa
(16). The findings of the present
study suggested that the downregulation of GALC protein expression
in the NPC cells may be associated with EBV latent infection and
the lymph node metastasis of NPC.
A study on EBV+ gastric carcinoma has
suggested that EBV-infected cells acquire extensive methylation
that silences multiple tumor-suppressor genes and rapidly
transforms into cancer cells, without going through a precancerous
stage of EBV infection or methylation accumulation (17). EBV+ gastric cancers show
distinct methylation patterns that are likely attributable to EBV
infection (18). The host defense
system induces aberrant methylation in the host cellular genome
(18). The host cells themselves
have a methylation mechanism and, besides EBV infection, also
triggers high methylation epigenotype, although this may be rare
and in most cases is triggered by EBV infection (19).
LOH on chromosome 14q is detected in 80% of NPC
tumors, with high-frequency LOH loci clustered at
14q11-q13,14q21-q24 and 14q32, while no LOH has been found at 14q31
(5). The GALC gene is
located at 14q31, has 17 exons and 16 introns of ~60 kb. The
deficiency of galactocerebrosidase (GALC) leads to the accumulation
of psychosine, which is responsible for globoid cell leukodystrophy
(Krabbe disease) in humans and certain animals. This enzyme
catalyzes the lysosomal hydrolysis of specific galactolipids,
including galactosylceramide (galactocerebroside) and
galactosylsphingosine (psychosine). The 5′-untranslated region is
GC-rich, containing no perfect CAAT or TATA sequences (6). Neither mutation nor other alterations
of the GALC gene promoter sequence have been detected
(16). Since the GALC gene
is located at 14q31, no LOH has been found at 14q31 and neither
mutation nor other alterations of the GALC gene promoter
sequence have been detected. Therefore, we speculated that the
GALC gene in NPC may be a tumor-suppressor gene, and
downregulation of GALC protein may be associated with GALC
gene promoter hypermethylation. Methylation of CpG sites is
reversible with pharmacological demethylation using epigenetic
agents such as 5-Aza-dC. Demethylation induced re-activation and
expression of the genes. The NPC CNE-2Z cell line was
EBV+ when it was established and now, after many years
of in vitro culture, the CNE-2Z cell line is
EBER− and LMP1−. In the present study, we
found that the CNE-2Z cell line exhibits a low level GALC mRNA
expression and does not express GALC protein. In addition,
hypermethylation (95.9%) was found at the CpG sites of the
GALC gene promoter in the CNE-2Z cell line. Treatment and
control groups were used in the present study. The CNE-2Z cells in
the control group were cultured for 48 h, while the cells in the
treatment groups were treated with different concentrations (5, 10,
20, 40 and 60 μM/l) of 5-Aza-dC for 48 h. Then, the GALC
mRNA and protein expression, GALC gene promoter methylation,
and cell viability in each group were measured. In addition, the
migration ability of the cells in the control group and the 20 and
40 μM/l treatment groups were also measured. The results
showed that compared with the control group, the treatment groups
showed a significantly lower rate of promotor methylation at the
CpG sites in the GALC gene, and these differences were more
pronounced in the 20, 40 and 60 μM/l treatment groups. The
expression of GALC mRNA was significantly higher in the
treated than in the control groups (P<0.05), and the expression
in the 20 μM/l treated group was the highest, which was
significantly different from that in the other groups (P<0.01).
GALC protein expression was detected in all the treatment groups
but not in the control group. Compared with the control group, the
cell viability in the 20, 40 and 60 μM/l treatment groups
was significantly lower (P<0.05), suggesting that the cell
proliferation ability decreased following treatment. The scratch
assay showed that the migration distance of the cells in the 20 and
60 μM/l treatment groups was significantly higher and the
cell-migration ability was significantly lower than the
corresponding values in the control group (P<0.01). The
difference between the 20 and 60 μM/l treatment groups was
not statistically significant (P>0.05). Transwell chamber assay
showed that significantly fewer cells migrated to the lower side in
the 20 and 60 μM/l treatment groups than in the control
group (P<0.01). The cell migration ability was also
significantly lower in the treatment groups. The difference between
the 20 and 60 μM/l treatment groups was not statistically
significant (P>0.05). These findings demonstrated that the
silencing of the GALC gene in the CNE-2Z cells was closely
associated with the hypermethylation of the CpG sites in the
promoter, while demethylation effectively inhibited the
proliferation and migration of the CNE-2Z cells. The results of the
present study were in accordance with previous findings (20,21).
Experiments on DNA methylation of cancer-related
genes have suggested that promoter hypermethylation is a potential
biomarker for the early diagnosis and a therapeutic target in
cancer (12–14). We report that GALC is
downregulated in NPC, and that promoter hypermethylation
contributes to the loss of GALC expression. To the best of
our knowledge, this is the first study demonstrating GALC
gene promoter hypermethylation in NPC. Our functional studies
support the putative tumor-suppressor effect of GALC in the
NPC CNE-2Z cell line. The present findings suggest that GALC
gene promoter hypermethylation is a potential epigenetic biomarker
for the early diagnosis and prediction of metastasis of
EBV-associated NPC.
Abbreviations:
|
NPC
|
nasopharyngeal carcinoma
|
|
EBV
|
Epstein-Barr virus
|
|
EBER
|
Epstein-Barr virus-encoded RNA
|
|
5-Aza-dC
|
5-Aza-2′-deoxycytidine
|
|
GALC
|
galactocerebrosidase
|
|
LMP
|
latent membrane protein
|
|
EBNA
|
EBV nuclear antigen
|
|
BARTs
|
BamHI A rightward transcripts
|
|
LOH
|
loss of heterozygosity
|
Acknowledgments
This study was supported by the National Basic
Research Program of China (973 Program, no. 2011CB504800), and the
2013 Scientific Research Foundation of Guangdong Medical
University, Zhanjiang, China (no. M2013033).
References
|
1
|
Young LS and Dawson CW: Epstein-Barr virus
and nasopharyngeal carcinoma. Chin J Cancer. 33:581–590.
2014.PubMed/NCBI
|
|
2
|
Busson P, Keryer C, Ooka T and Corbex M:
EBV-associated nasopharyngeal carcinomas: From epidemiology to
virustargeting strategies. Trends Microbiol. 12:356–360. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tao Q, Young LS, Woodman CB and Murray PG:
Epstein-Barr virus (EBV) and its associated human cancers -
genetics, epigenetics, pathobiology and novel therapeutics. Front
Biosci. 11:2672–2713. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lo KW, Teo PM, Hui AB, To KF, Tsang YS,
Chan SY, Mak KF, Lee JC and Huang DP: High resolution allelotype of
microdissected primary nasopharyngeal carcinoma. Cancer Res.
60:3348–3353. 2000.PubMed/NCBI
|
|
5
|
Shao J, Li Y, Wu Q, Liang X, Yu X, Huang
L, Hou J, Huang X, Ernberg I, Hu LF, et al: High frequency loss of
heterozygosity on the long arms of chromosomes 13 and 14 in
nasopharyngeal carcinoma in Southern China. Chin Med J.
115:571–575. 2002.PubMed/NCBI
|
|
6
|
Luzi P, Rafi MA and Wenger DA: Structure
and organization of the human galactocerebrosidase (GALC) gene.
Genomics. 26:407–409. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kwong J, Lo KW, To KF, Teo PM, Johnson PJ
and Huang DP: Promoter hypermethylation of multiple genes in
nasopharyngeal carcinoma. Clin Cancer Res. 8:131–137.
2002.PubMed/NCBI
|
|
8
|
Tong JH, Tsang RK, Lo KW, Woo JK, Kwong J,
Chan MW, Chang AR, van Hasselt CA, Huang DP and To KF: Quantitative
Epstein-Barr virus DNA analysis and detection of gene promoter
hypermethylation in nasopharyngeal (NP) brushing samples from
patients with NP carcinoma. Clin Cancer Res. 8:2612–2619.
2002.PubMed/NCBI
|
|
9
|
Challouf S, Ziadi S, Zaghdoudi R, Ksiaa F,
Ben Gacem R and Trimeche M: Patterns of aberrant DNA
hypermethylation in nasopharyngeal carcinoma in Tunisian patients.
Clin Chim Acta. 413:795–802. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tong JH, Ng DC, Chau SL, So KK, Leung PP,
Lee TL, Lung RW, Chan MW, Chan AW, Lo KW, et al: Putative
tumour-suppressor gene DAB2 is frequently down regulated by
promoter hypermethylation in nasopharyngeal carcinoma. BMC Cancer.
10:2532010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bodoor K, Haddad Y, Alkhateeb A, Al-Abbadi
A, Dowairi M, Magableh A, Bsoul N and Ghabkari A: DNA
hypermethylation of cell cycle (p15 and p16) and apoptotic (p14,
p53, DAPK and TMS1) genes in peripheral blood of leukemia patients.
Asian Pac J Cancer Prev. 15:75–84. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Park JY: Promoter hypermethylation as a
biomarker in prostate adenocarcinoma. Methods Mol Biol.
1238:607–625. 2015. View Article : Google Scholar
|
|
13
|
Wang HL, Zhou PY, Liu P and Zhang Y: Role
of p16 gene promoter methylation in gastric carcinogenesis: A
meta-analysis. Mol Biol Rep. 41:4481–4492. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gloss BS, Patterson KI, Barton CA,
Gonzalez M, Scurry JP, Hacker NF, Sutherland RL, O'Brien PM and
Clark SJ: Integrative genome-wide expression and promoter DNA
methylation profiling identifies a potential novel panel of ovarian
cancer epigenetic biomarkers. Cancer Lett. 318:76–85. 2012.
View Article : Google Scholar
|
|
15
|
Wei KR, Xu Y, Liu J, Zhang WJ and Liang
ZH: Histopathological classification of nasopharyngeal carcinoma.
Asian Pac J Cancer Prev. 12:1141–1147. 2011.PubMed/NCBI
|
|
16
|
Görögh T, Rudert H, Lippert BM,
Gottschlich S, Maune S, Heidorn K, Maass J, Hoffmann M, Meyer JE,
Rathcke IO, et al: Transcriptional repression of the human
galactocerebrosidase gene in squamous cell carcinomas of the
larynx. Int J Cancer. 83:750–754. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kaneda A, Matsusaka K, Aburatani H and
Fukayama M: Epstein-Barr virus infection as an epigenetic driver of
tumorigenesis. Cancer Res. 72:3445–3450. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Matsusaka K, Kaneda A, Nagae G, Ushiku T,
Kikuchi Y, Hino R, Uozaki H, Seto Y, Takada K, Aburatani H, et al:
Classification of Epstein-Barr virus-positive gastric cancers by
definition of DNA methylation epigenotypes. Cancer Res.
71:7187–7197. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fukayama M, Hino R and Uozaki H:
Epstein-Barr virus and gastric carcinoma: Virus-host interactions
leading to carcinoma. Cancer Sci. 99:1726–1733. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Owczarek TB, Suchanski J, Pula B, Kmiecik
AM, Chadalski M, Jethon A, Dziegiel P and Ugorski M:
Galactosylceramide affects tumorigenic and metastatic properties of
breast cancer cells as an anti-apoptotic molecule. PLoS One.
8:e841912013. View Article : Google Scholar
|
|
21
|
Beier UH and Görögh T: Implications of
galactocerebrosidase and galactosylcerebroside metabolism in cancer
cells. Int J Cancer. 115:6–10. 2005. View Article : Google Scholar : PubMed/NCBI
|