Introduction
Epigenetic alteration, which refers to the
regulation of gene expression via post-translational modification
of the chromatin structure without changes in the underlying DNA
sequence, is crucial in diverse physiological and pathological
cellular processes (1). Epigenetic
defects in cancer cells can be efficiently reverted by means of
pharmacological inhibitors of the enzymes that are responsible for
establishing/maintaining the epigenetic markers (2). In addition, disruption of the
epigenome is a fundamental mechanism in cancer development, and
several epigenetic drugs, including azacitidine, decitabine,
vorinostat and romidepsin, have been approved for cancer treatment
(3). Data derived from experiments
and clinical studies clearly indicate that epigenetic drugs are
effective as modifiers of cancer phenotype and positive regulators
of cancer cells in cancer therapy.
Acetylation, one of the most common modifications in
epigenetics, serves as a key regulatory mechanism for chromatin
structure and gene expression (4).
Histone deacetylases (HDACs) are a vast family of enzymes involved
in chromatin remodeling and have crucial roles in numerous
biological processes. The finding that overexpression of HDACs is
associated with tumor initiation and progression has prompted the
research and development of HDAC inhibitors (HDACIs) as anticancer
agents. HDAC overexpression alters the expression and function of
tumor-associated proteins that are predominantly involved in cell
proliferation, migration, angiogenesis and metastasis (5,6). In
addition, patients suffering from cancer with aberrant HDAC
expression show a positive correlation with poor prognosis
(7,8). HDACIs can be divided into categories
of short chain fatty acids, hydroxamic acids, cyclic tetrapeptides
and benzamides, the majority of which appear to act by blocking the
HDAC catalytic site containing a Zn2+ ion (9). HDAC inhibition provokes accumulation
of acetylated histones that become incorporated into the
nucleosomes, leading to the reversal of aberrant epigenetic
patterns observed in cancer cells. Evidently, epigenetic drugs
represent a category of promising agents in cancer treatment. Given
the potent antitumor effects of HDACIs, HDACs themselves may be
oncogenic, which is supported by the fact that HDAC function and/or
expression is perturbed in different types of cancer and is often
associated with a poor prognosis (10).
In complex biological systems, small molecules often
mediate microbe-microbe and microbe-host interactions. As reported,
there is widespread distribution of small-molecule-encoding
biosynthetic gene clusters (BGCs) in the human microbiome (11); and accordingly, a great potential
for the bacterial production of drug-like molecules in humans. Data
have suggested that there are hundreds of widely distributed BGCs
of unknown function in the human microbiome (11). Another study showed that the
chemopreventive effect of a diet rich in fiber and slowly
digestible carbohydrates has been attributed, among a variety of
factors, to enhanced butyrate formation in the colon (12). It also described phenylacetate and
phenylbutyrate as HDACIs, which were of particular interest as
these compounds could be formed from polyphenols in fruits and
vegetables during intestinal passage (13). Nutrition and the microbial flora
are considered to have a marked influence on the risk of developing
colorectal cancer, and the formation of butyrate and other
short-chain fatty acids (SCFAs) possibly exhibit a key role as
chemopreventive products of microbial fermentation in the colon.
Research has suggested that butyrate acts as an endogenous HDACI in
the colon (14).
Cinnamic acid (CA), a constituent of Cinnamomum
cassia (family, Lauraceae), exhibits a broad spectrum of
biological activity including antioxidant, anti-inflammatory and
anticancer activities (15–17).
CA was observed to be cardioprotective in a rat model of ischemic
myocardial injury and this effect was attributable to its
anti-oxidative and anti-inflammatory properties, as well as the
increased NO level (18). CA shows
protective effects against cisplatin-induced nephrotoxicity that
may be attributed to its antioxidant activities (19). CA that occurs in propolis can
modulate antigen receptors, cytokine production and the fungicidal
activity of human monocytes depending on its concentration
(20). Studies have also suggested
the possibility of using CA for preventing advanced glycation
end-product-mediated diabetic complications and for diabetes
management (21,22). CA is a naturally occurring phenolic
compound with antimicrobial activity. Cis-CA (cCA) may be a
potential antimycobacterial or synergistic agent that can be
developed against tuberculosis (23). As described, CA is one of the major
components of Brazilian green propolis, which exhibits a gastric
protective effect and anticancer activity (24). It is reported that CA may act as a
skin whitening agent via the inhibition of tyrosinase activity and
its expression in melanocytes (25). Furthermore, CA shows
antiproliferative activity against melanoma cells (26) and lung carcinoma cells (27). In addition, a variety of CA
derivatives have been reported as HDAC-binding components (28). However, the HDAC inhibitory
activity of trans-CA (tCA) itself remains to be fully
defined.
Although isolated from plants, tCA may also be found
as an endogenous substance in human body, predominantly in the
colon. It has been demonstrated that tCA is one of the intestinal
metabolites of nutritional polyphenols (13). Following analysis tCA and a great
variety of its closely related analogous compounds, such as,
caffeic acid, coumaric acid, ferulic acid and isoferulic acid, were
found to be present in feces (14). It was reported that butyrate, other
SCFAs and tCA derivatives were formed during the degradation of
polyphenolic constituents of fruits and vegetables by intestinal
microorganisms, and inhibited global HDAC activity in nuclear
extracts from HT-29 human colon carcinoma cells (14). Thus, tCA occurs as an endogenous
substance in the colon and it is worthwhile to investigate its
potential effects on various pathological processes, such as
carcinogenesis and tumor progression in the colon.
The present study aimed to assess the effect of tCA
on HDAC activity in cancer cells and the underlying molecular
mechanisms; as well as to evaluate the therapeutic efficacy of tCA
against colon carcinoma xenografts in athymic mice.
Materials and methods
Reagents
HT29 human colon carcinoma cells, H460 human lung
carcinoma cells, A549 human lung adenocarcinoma cells, and MIA
PaCa-2 human pancreatic carcinoma cells were obtained from the
American Type Culture Collection (Manassas, VA, USA). Fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific Inc., Waltham, MA,
USA), RPMI-1640 medium (Gibco, Thermo Fisher Scientific Inc.) and
Dulbecco's modified Eagle's medium (DMEM; Hyclone; Thermo Fisher
Scientific Inc.) were used in the experiments. All antibodies were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
APC Annexin V was purchased from Invitrogen (Thermo Fisher
Scientific Inc.) and SYTOX Green Nucleic Acid Stain was from BD
Biosciences (Franklin Lakes, NJ, USA). Fluorescein isothiocyanate
(FITC)-dextran and tCA were purchased from Sigma-Aldrich (St.
Louis, MO, USA). EIPA was obtained from Thermo Fisher Scientific
Inc. The HDAC-GloTM I/II Assay and Screening system were obtained
from Promega Corporation (Waltham, MA, USA).
Cell culture
HT29, H460 and A549 cells were grown in modified
RPMI-1640 supplemented with 10% heat-inactivated FBS, penicillin G
(100 U/ml) and streptomycin (100 µg/ml) (both from North
China Pharmaceutical, Co., Ltd., Shijiazhuang, China). MIA PaCa-2
cells were grown in DMEM supplemented with the same substances. All
cell lines were cultured at 37°C in a humidified 5% CO2
incubator.
Cytotoxicity assay
Cells were plated in 96-well plates at a density of
3,000 cells per well and incubated for 24 h at 37°C. Then,
different concentrations (0.09, 0.17, 034, 0.68, 1.36 and 2.72 mM)
of tCA were added. At 48 h, MTT (Amresco Inc, Solon, OH, USA) was
added to each well and further incubated for another 4 h. The
medium containing MTT solution was then discarded, and 150
µl dimethyl sulfoxide was added to each well. The absorbance
(A) at 570 nm was detected with a microplate reader (Multiskan MK3;
Thermo Fisher Scientific, Inc.). Untreated cells served as a
control. The tCA relative cell viability compared with the control
group was calculated according to the following formula: Cell
viability (%) =
[(Atreat−Ablank)/(Acontrol−Ablank)]
× 100.
HDAC activity assay in a cell-free system
and in cultured cells
HDAC activity in the cell-free system and in
cultured cells was detected with the HDAC-GloTM I/II Assay and
Screening system following the standard protocol (29). A HDAC assay was performed in a
white 96-well plate. For the HDAC activity assay in the cell-free
system, cell lysate was incubated with tCA and trichostatin A (TSA)
in HDAC assay buffer (100 ml) at 37°C for 60 min and then 100 ml
HDACTM I/II reagent were added. After 30 min, luminescence was
measured. For the HDAC assay in cultured cells, 10,000 cells per
well were treated with tCA or control agents for 6 or 12 h (for tCA
and TSA in Hela nuclear extract, the concentrations were 6.25,
12.5, 25, 50, 100 and 200 µM; for tCA in HT29 cells, 187.5,
375.5, 750, 1,500 and 3,000 µM; and for TSA in HT29 cells,
3.125, 6.25, 12.5, 25 and 50 nM). Then, cell medium was changed to
serum-free medium with HDAC buffer (50+50 ml). After 15 min
incubation, HDAC I/II reagents (100 ml) were added and luminescence
was measured using Enspire 2300 multi-label reader (Perkin Elmer,
Waltham, MA, USA) after 30 min.
Western blot analysis
After incubation at 37°C for 12 h, ice-cold
phosphate-buffered saline (PBS) solution was used twice to rinse
cells. Then cells were lysed with cell lysis buffer and the dishes
were incubated for 10–30 min at 4°C. Cells were scraped into lysis
buffer, and the lysates were clarified by centrifugation at 15,294
× g for 15 min at 4°C. Protein concentrations were determined using
a Bicinchoninic acid Protein Assay kit (Bio-Rad, Hercules, CA, USA)
and western blotting was performed. Briefly, an equal amount of
total protein extracted from cultured cells was separated by 12%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis and
transferred to polyvinylidene difluoride (PVDF; Millipore,
Billerica, MA, USA) membranes. The following primary antibodies and
horseradish peroxidase-conjugated appropriate secondary antibodies
were used to detect the designated proteins: Acetyl-histone H3
(Ac-H3), (Lys9) rabbit monoclonal antibody (1:1,000; cat. no.
#9649; Cell Signaling Technology Inc.); Ac-H4 (Lys8) rabbit
polyclonal antibody (1:1,000; cat. no. #2594; Cell Signaling
Technology Inc.); β-actin rabbit polyclonal antibody (1:5,000; cat.
no. SC-1616, Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA);
PARP polyclonal antibody (1:1,000; cat. no. #9542, Cell Signaling
Technology Inc.); Bax rabbit monoclonal antibody (1:1,000; cat. no.
#5023, Cell Signaling Technology Inc.); Bcl-2 rabbit monoclonal
antibody (1:1,000; cat. no. #2870, Cell Signaling Technology Inc.);
goat anti-mouse peroxidase-coupled antibody (cat. no. ZB-2305) and
goat anti-rabbit peroxidase-coupled antibody (cat. no. ZB-2301)
(1:5,000; Zhongshan Goldenbridge Biotechnology Co., Ltd., Beijing,
China). The bound secondary antibodies on the PVDF membrane
combined with enhanced chemiluminescence detection reagents (Pierce
Biotechnology, Waltham, MA, USA) and exposed to X-ray films (Kodak,
Tokyo, Japan), according to the manufacturer's protocol.
Cell apoptosis assay
The cell apoptosis assay was performed using APC
Annexin V and SYTOX Green Nucleic Acid staining. In brief, cultured
HT29 cells were washed with cold PBS and combined with binding
buffer. Cells were then incubated with APC Annexin V for 10 min and
SYTOX Green Nucleic Acid Stain for another 10 min at 4°C in the
dark. Then, the stained cells were observed and images were
captured with confocal microscopy LSM 710 (Zeiss, Weimar, Germany)
within 60 min.
Therapeutic experiment with HT29 colon
carcinoma xenografts
Female BALB/c nude mice (age, 4–6 weeks) were
purchased from the Institute for Experimental Animals, Chinese
Academy of Medical Sciences & Peking Union Medical College
(Beijing, China) and allowed to adapt to study environment for one
week prior to the experiment. The study protocols were in
accordance with the regulations of Good Laboratory Practice for
non-clinical laboratory studies of drugs issued by the National
Scientific and Technologic Committee of People's Republic of China.
This study was approved by the animal care committee of the
Institute of Medicinal Biotechnology, Chinese Academy of Medical
Sciences and Peking Union Medical College (Beijing, China). HT29
colon carcinoma cells (1×107) suspended in 200 µl
sterile saline were inoculated subcutaneously into the right armpit
of nude mice. After ~3 weeks, tumors in the donor animals were
aseptically dissected and cut into 2-mm3 blocks. Then,
the tumor tissue was transplanted subcutaneously by a trocar into
nude mice. When the tumor size reached ~100 mm3, mice
were divided into the following groups (n=6): Treatment groups with
different doses of tCA (1.0 and 1.5 mmol/kg respectively) and the
control group. tCA was intragastrically administered, 3 times a
week for a total of 6 doses. During the experiment, the long
diameter and the perpendicular short diameter of HT29 xenografts
were measured every 2 days for volume calculation and statistical
analysis. Tumor volume was estimated by the following formula:
V=0.5axb2, where a and b represented the long and the
perpendicular short diameters of the tumor, respectively.
Thirty-five days after inoculation, the animals were sacrificed by
overdose of isoflurane anesthesia (Shandong Keyuan Pharmaceutical
Co., Ltd., Jinan, China) and the xenograft tumors were removed and
weighed. Specimens taken from various organs including the heart,
lung, liver, pancreas, small intestine, large intestine, kidney,
spleen and bone marrow of the femur were preserved in a 4%
formaldehyde solution for further evaluation.
Immunohistochemistry
HT29 xenograft specimens were used for
immunohistochemical detection of related proteins, such as
acetyl-H3, cleaved PARP and cleaved caspase-3. Sections (4
µm) were deparaffinized and rehydrated with xylene and
graded alcohol solutions. After washing with PBS, endogenous
peroxidase activity was quenched with 3% hydrogen peroxide.
Sections were boiled in 10 mM citrate buffer (pH 6.0; Zhongshan
Goldenbridge Biotechnology Co., Ltd.) for 3 min in an autoclave
sterilizer (Zealway Instrument Inc., Xiameng, China) followed by
cooling at room temperature for >20 min. After rinsing with PBS,
sections were incubated with all primary antibodies used in the
western blot analysis (1:100 diluted in antibody diluent, Zhongshan
Goldenbridge Biotechnology Co., Ltd.) for 18 h at 4°C. Sections
were stained with the related antibodies. After rinsing with PBS,
the sections were incubated with PV6001 or PV6002 (Zhongshan
Goldbridge Biotechnology Co., Ltd.) for 30 min at 37°C and stained
with DAB (AR1022, Boster Biological Technology, Ltd., Wuhan, China)
for 1 to 2 min. The slides were counterstained with hematoxylin
(Sigma-Aldrich), dehydrated with ethanol, cleared with xylene and
mounted in neutral gum. Control sections were incubated with PBS
instead of a primary antibody. All slides were analyzed by two
independent observers.
Histopathological observation of various
organs from tCA-treated xenograft-bearing mice
At the end of experiment, specimens were obtained
from the heart, lung, liver, pancreas, small intestine, large
intestine, kidney, spleen and bone marrow of the femur, of the
xenograft-bearing nude mice. Histological sections (5 µm)
were stained with hematoxylin and eosin and observed under a light
microscope (Olympus IX81, Olympus, Tokyo, Japan).
Statistical analysis
All values are expressed as the mean ± standard
deviation. Statistical analysis was conducted using one-way
analysis of variance with SPSS statistical software 17.0 (SPSS
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Inhibition of cancer cell proliferation
by tCA
An MTT assay demonstrated that tCA inhibited the
proliferation of HT29, MIA PaCa-2, H460 and A549 cells in a
concentration-dependent manner. The IC50 values for HT29, MIA
PaCa-2, H460 and A549 cells were 1.07±0.38, 1.33±0.07, 2.10±0.43
and 3.54±0.34 mM, respectively. Among the tested cell lines, HT29
cells were the most sensitive to tCA.
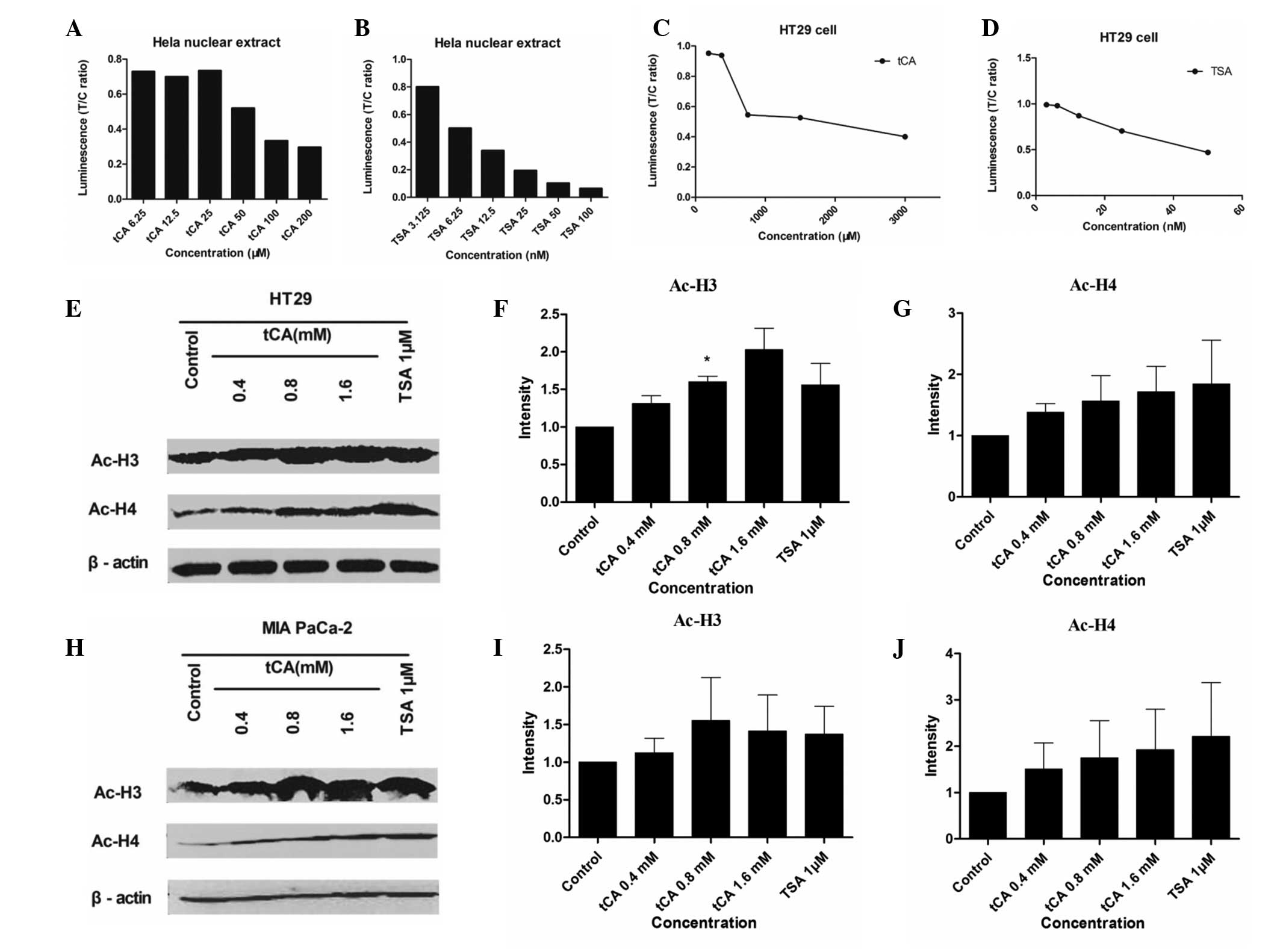
Increase of histone acetylation in
cultured cancer cells and in vivo
The present study determined whether tCA treatment
resulted in increased acetylation of histone proteins in HT29 cells
and MIA PaCa-2 cells. HDACI/II activity was detected for Hela
nuclear extract and HT29 cells treated with various doses of tCA
and TSA. As shown, tCA directly inhibited HDAC activity similar to
TSA. Western blotting of the extracted protein from HT29 cells and
MIA PaCa-2 cells treated with 0.4, 0.8 and 1.6 mM tCA showed a
marked increase in acetyl-H3 and acetyl-H4, compared with untreated
controls. The increase of acetylation observed in histone H3 and H4
after tCA exposure was similar to that observed after treatment
with TSA (Fig. 1).
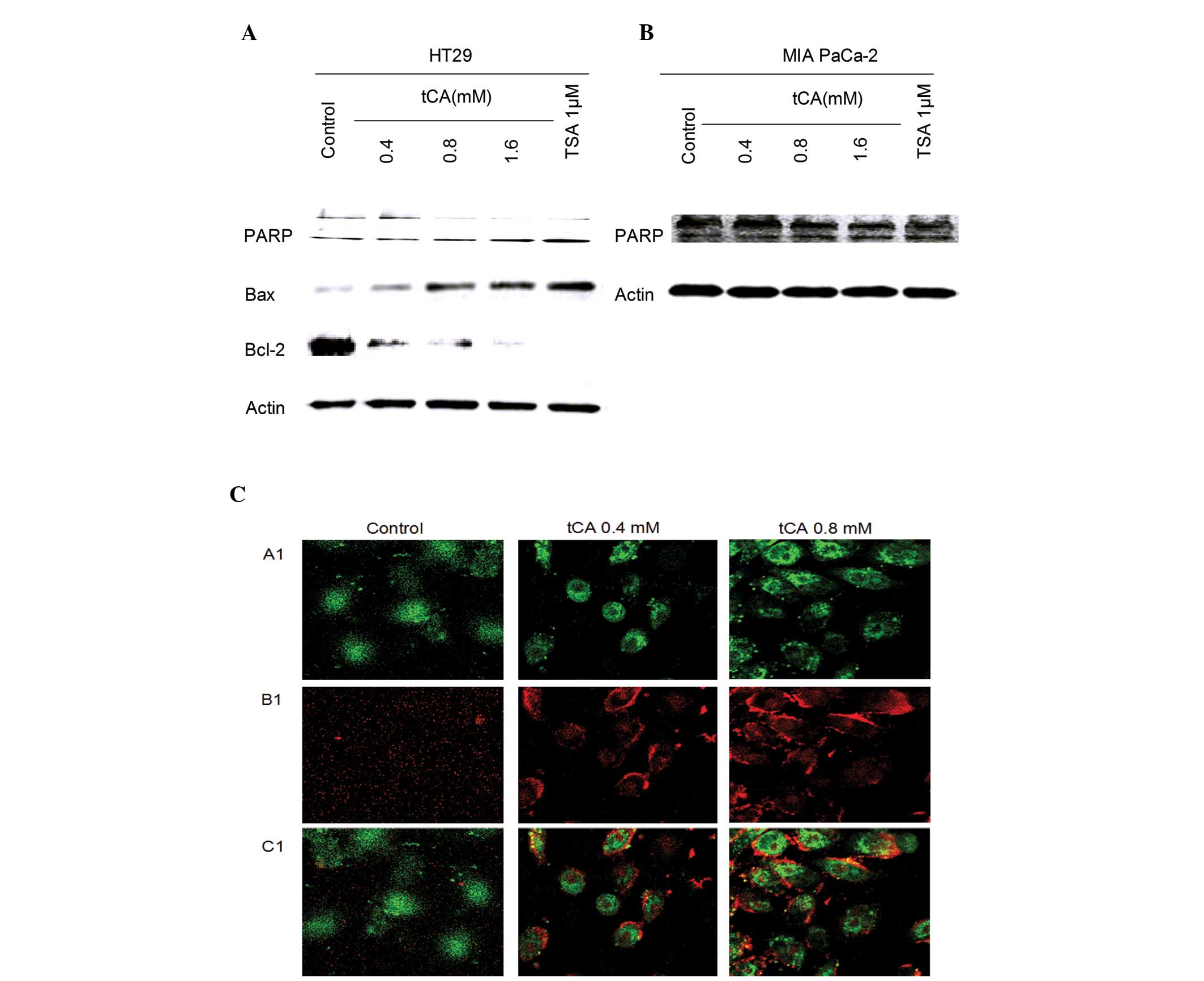
Induction of apoptosis by tCA in vivo and
in vitro
Inhibition of HDAC activity has been shown to lead
to the induction of apoptosis in cancer cells (30). The effect of tCA on apoptosis was
evaluated in HT29 cells and MIA PaCa-2 cells. The effect of tCA on
the expression of apoptosis-related proteins showed that the
expression level of Bax was upregulated; while the expression
levels of PARP and Bcl-2 were downregulated. tCA resulted in an
increase in the expression of apoptosis-related protein Bax and a
decrease in the expression levels of PARP and Bcl-2. This change in
expression levels is similar to that induced by the HDAC inhibitor
TSA (Fig. 2A and B). The cells
undergoing apoptosis were observed by APC Annexin V and SYTOX Green
Nucleic Acid staining. A significant increase in APC Annexin V and
SYTOX Green Nucleic Acid stained cells was observed after tCA
treatment in MIA PaCa2 cells (Fig.
2C).
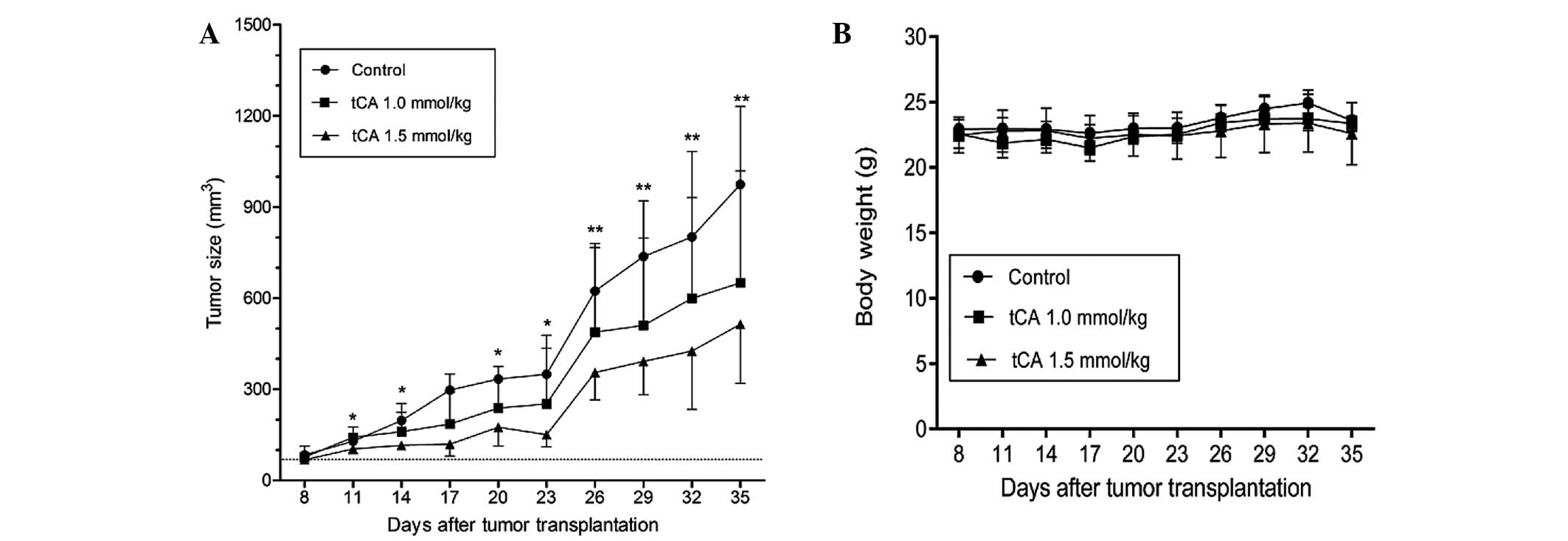
Therapeutic efficacy of tCA against HT29
colon carcinoma xenograft
Based on the above experiments which demonstrated
that tCA was active in the inhibition of HDAC, the suppression of
cell proliferation and the induction of apoptosis, the study was
extended to evaluate the therapeutic efficacy of tCA using a HT29
colon carcinoma xenograft model. As shown in Fig. 3, determined by tumor volume, tCA at
doses 1.0 and 1.5 mmol/kg inhibited xenograft growth by 39
(P<0.05) and 52.9% (P<0.01), respectively, at the termination
of the experiment. The inhibition rate of tumor growth was
calculated with the formula: Inhibition rate (%) = [1−tumor volume
(treated)/tumor volume (control)] × 100. The average daily intake
of food did not differ between the control and treated groups.
Moreover, body weights were not significantly different between the
treated and the control animals for the duration of the study
(Fig. 3B), suggesting that the
administered doses of tCA were well-tolerated.
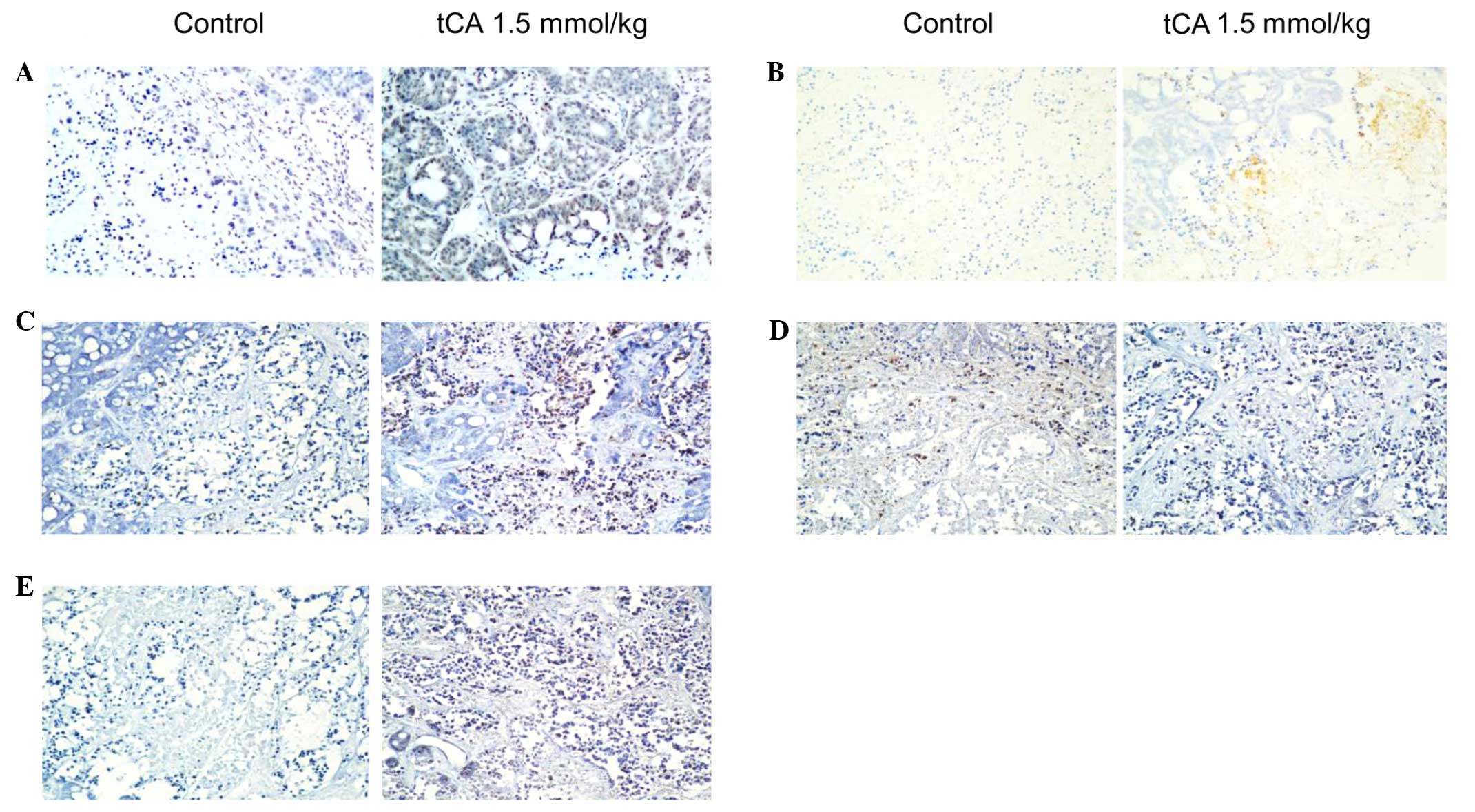
Immunohistochemical examination on HT29
tumor tissue
To determine the location of related proteins,
immunohistochemical analysis was performed using HT29 cancer
xenograft tissues from the control and tCA-treated nude mice. Using
immunohistochemical staining, it was demonstrated that the
expression of Ac-H3 in tCA-treated HT29 cancer xenograft tissues
was higher than that of control (Fig.
4A). Ac-H3 and cleaved caspase 3 were overexpressed in
tCA-treated HT29 cancer tissues compared with the control tissue.
Cleaved PARP was also overexpressed in tCA-treated HT29 cancer
tissues compared with the control tissue. In addition, the
expression of Bax increased and that of Bcl-2 decreased following
tCA treatment (Fig. 4).
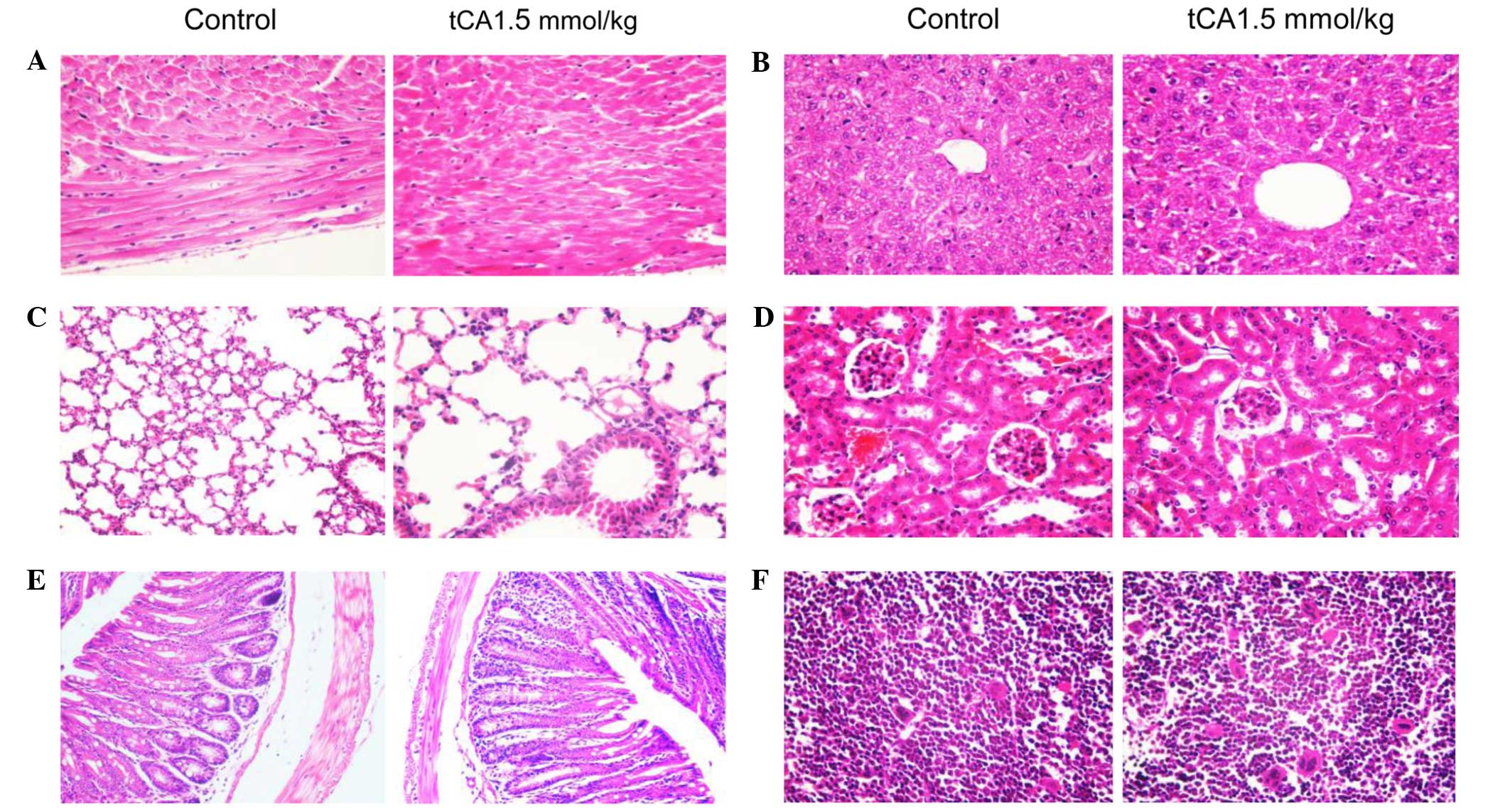
Histopathological observation of various
organs from tCA-treated xenograft-bearing mice
At the end of experiment, specimens were obtained
from the heart, lung, liver, pancreas, small intestine, large
intestine, kidney, spleen and bone marrow of the femur, of the
xenograft-bearing nude mice. No toxicopathological changes were
identified in any of the tested organs (Fig. 5).
Discussion
Epigenetic programs are now widely recognized as
being critical to the biological processes of cancer genesis. It
has been recognized in recent years that HDACs are promising
targets for therapeutic interventions intended to reverse aberrant
acetylation states. In the present study, it was shown by western
blotting that tCA upregulated the expression of acetyl-H3 and
acetyl-H4 proteins in a dose-dependent manner in treated cells and
in the tumor tissue of treated mice, which was consistent with the
well-known HDAC inhibitor TSA. Therefore, tCA could be confirmed as
an HDAC inhibitor. Furthermore, it was shown that tCA inhibited the
proliferation of HT29, H460, A549 and MIA PaCa-2 cell lines, and
the IC50 value for HT29 colon carcinoma cells was ~1 mM. According
to a previous study determined by another viability assay, the IC50
value for HT-144 cells was 2.4 mM; and activated-caspase 3 staining
demonstrated the induction of apoptosis in HT-144 cells 24 h after
exposure to 3.2 mM CA (26). As
compared with that of classical inhibitors, such as TSA, the
activity of tCA is modest. For the inhibition of cancer cell
proliferation, the concentration of tCA reaching mM levels was
required; however, the study data verified that tCA interfered in
histone acetylation via inhibiting HDAC, leading to cancer cell
cytotoxicity. In addition, the therapeutic efficacy of tCA has been
confirmed. Due to its low toxicity, tCA could suppress the growth
of colon carcinoma HT29 xenografts at well-tolerated doses. With
regards to the mechanism of action, it is possible that tCA and
HDAC were in a binding state under physiological conditions;
however, an in depth study is required to determine the precise
mechanism of action and the specificity of tCA binding to HDAC.
In recent years, the interaction between commensal
microorganisms and the host in the development of diseases has
caused much concern. The intricate relationships between microbiota
and host may benefit the host in a number of ways; however, may
also carry risks for disease development. One of the most important
topics is whether these microorganisms can produce bioactive
substances that are beneficial to health, in particular, effective
for the treatment of diseases. A recent report showed that
lactocillin, a thiopeptide antibiotic produced by a member of the
vaginal microbiota, had potent antibacterial activity against a
range of Gram-positive vaginal pathogens (11). As reported, there was a widespread
distribution of small-molecule-encoding BGCs in the human
microbiome; accordingly, a great potential for the bacterial
production of drug-like molecules in humans. Data further
highlighted the fact that there are hundreds of widely distributed
BGCs of unknown function in the human microbiome (11). In total 99% of the microbial mass
is located in the gastrointestinal tract, with the majority in the
colon. Therefore, attention should be paid to those active
substances derived from the microbes existing in the colon. A study
showed that the chemopreventive effect of a diet rich in fiber and
slowly digestible carbohydrates had been attributed, among a
variety of factors, to enhanced butyrate formation in the colon
(12). In addition, phenylacetate
and phenylbutyrate, which are active HDAC inhibitors, are formed
from polyphenols in fruits and vegetables during intestinal passage
(13). Thus, nutrition and the
microbial flora are considered to have a marked influence on the
risk of colorectal cancer, as the formation of butyrate and other
short-chain fatty acids produced by microbial fermentation possibly
exhibit a chemopreventive role. A previous study demonstrated that
butyrate exhibited a role as an endogenous HDAC inhibitor in the
colon (14). The present study
demonstrated that tCA as a HDAC inhibiting agent exhibits
therapeutic efficacy against colon carcinoma xenografts, implying
the potential of developing cancer-active compounds derived from
microorganisms that reside in the intestinal tract, particularly in
the colon.
In conclusion, this study indicates that tCA is
effective against colon cancer xenograft in nude mice. The
antitumor mechanism of tCA could be mediated, at least in part, by
inhibiting HDAC in cancer cells. Considering the fact that tCA can
be produced by the metabolic activity of the microorganisms
existing in the intestinal tract, the antitumor efficacy of tCA
warrants further investigation.
Acknowledgments
This study was supported by 'Significant New Drug
Development' Major Science and Technology Projects of China (grant
nos. 2013ZX09102064 and 2012ZX09301002-001-015).
References
|
1
|
Jaenisch R and Bird A: Epigenetic
regulation of gene expression: How the genome integrates intrinsic
and environmental signals. Nat Genet. (33 Suppl): 245–254. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yoo CB and Jones PA: Epigenetic therapy of
cancer: Past, present and future. Nat Rev Drug Discov. 5:37–50.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Knower KC, To SQ, Leung YK, Ho SM and
Clyne CD: Endocrine disruption of the epigenome: a breast cancer
link. Endocr Relat Cancer. 21:T33–T55. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sterner DE and Berger SL: Acetylation of
histones and transcription-related factors. Microbiol Mol Biol Rev.
64:435–459. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Glozak MA and Seto E: Histone deacetylases
and cancer. Oncogene. 26:5420–5432. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu WS, Parmigiani RB and Marks PA: Histone
deacetylase inhibitors: Molecular mechanisms of action. Oncogene.
26:5541–5552. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mitsiades CS, Mitsiades NS, McMullan CJ,
Poulaki V, Shringarpure R, Hideshima T, Akiyama M, Chauhan D,
Munshi N, Gu X, et al: Transcriptional signature of histone
deacetylase inhibition in multiple myeloma: Biological and clinical
implications. Proc Natl Acad Sci USA. 101:540–545. 2004. View Article : Google Scholar :
|
|
8
|
Zhang Z, Yamashita H, Toyama T, Sugiura H,
Omoto Y, Ando Y, Mita K, Hamaguchi M, Hayashi S and Iwase H: HDAC6
expression is correlated with better survival in breast cancer.
Clin Cancer Res. 10:6962–6968. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vannini A, Volpari C, Filocamo G, Casavola
EC, Brunetti M, Renzoni D, Chakravarty P, Paolini C, De Francesco
R, Gallinari P, et al: Crystal structure of a eukaryotic
zinc-dependent histone deacetylase, human HDAC8, complexed with a
hydroxamic acid inhibitor. Proc Natl Acad Sci USA. 101:15064–15069.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Falkenberg KJ and Johnstone RW: Histone
deacetylases and their inhibitors in cancer, neurological diseases
and immune disorders. Nat Rev Drug Discov. 13:673–691. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Donia MS, Cimermancic P, Schulze CJ,
Wieland Brown LC, Martin J, Mitreva M, Clardy J, Linington RG and
Fischbach MA: A systematic analysis of biosynthetic gene clusters
in the human microbiome reveals a common family of antibiotics.
Cell. 158:1402–1414. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Marks PA, Richon VM, Miller T and Kelly
WK: Histone deacetylase inhibitors. Adv Cancer Res. 91:137–168.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jenner AM, Rafter J and Halliwell B: Human
fecal water content of phenolics: The extent of colonic exposure to
aromatic compounds. Free Radic Biol Med. 38:763–772. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Waldecker M, Kautenburger T, Daumann H,
Busch C and Schrenk D: Inhibition of histone-deacetylase activity
by short-chain fatty acids and some polyphenol metabolites formed
in the colon. J Nutr Biochem. 19:587–593. 2008. View Article : Google Scholar
|
|
15
|
Zhang LP and Ji ZZ: Synthesis,
antiinflammatory and anticancer activity of cinnamic acids, their
derivatives and analogues. Yao Xue Xue Bao. 27:817–823.
1992.PubMed/NCBI
|
|
16
|
Akao Y, Maruyama H, Matsumoto K, Ohguchi
K, Nishizawa K, Sakamoto T, Araki Y, Mishima S and Nozawa Y: Cell
growth inhibitory effect of cinnamic acid derivatives from propolis
on human tumor cell lines. Biol Pharm Bull. 26:1057–1059. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Foti MC, Daquino C and Geraci C:
Electron-transfer reaction of cinnamic acids and their methyl
esters with the DPPH(*) radical in alcoholic solutions.
J Org Chem. 69:2309–2314. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Song F, Li H, Sun J and Wang S: Protective
effects of cinnamic acid and cinnamic aldehyde on
isoproterenol-induced acute myocardial ischemia in rats. J
Ethnopharmacol. 150:125–130. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
El-Sayed el SM, Abd El-Raouf OM, Fawzy HM
and Manie MF: Comparative study of the possible protective effects
of cinnamic acid and cinnamaldehyde on cisplatin-induced
nephrotoxicity in rats. J Biochem Mol Toxicol. 27:508–514. 2013.
View Article : Google Scholar
|
|
20
|
Conti BJ, Búfalo MC, Golim Mde A, Bankova
V and Sforcin JM: Cinnamic Acid is partially involved in propolis
immunomodu-latory action on human monocytes. Evid Based Complement
Alternat Med. 2013:1098642013. View Article : Google Scholar
|
|
21
|
Adisakwattana S, Sompong W, Meeprom A,
Ngamukote S and Yibchok-Anun S: Cinnamic acid and its derivatives
inhibit fructose-mediated protein glycation. Int J Mol Sci.
13:1778–1789. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kasetti RB, Nabi SA, Swapna S and Apparao
C: Cinnamic acid as one of the antidiabetic active principle(s)
from the seeds of Syzygium alternifolium. Food Chem Toxicol.
50:1425–1431. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen YL, Huang ST, Sun FM, Chiang YL,
Chiang CJ, Tsai CM and Weng CJ: Transformation of cinnamic acid
from trans- to cis-form raises a notable bactericidal and
synergistic activity against multiple-drug resistant Mycobacterium
tuberculosis. Eur J Pharm Sci. 43:188–194. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Barros MP, Lemos M, Maistro EL, Leite MF,
Sousa JP, Bastos JK and Andrade SF: Evaluation of antiulcer
activity of the main phenolic acids found in Brazilian Green
Propolis. J Ethnopharmacol. 120:372–377. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kong YH, Jo YO, Cho CW, Son D, Park S, Rho
J and Choi SY: Inhibitory effects of cinnamic acid on melanin
biosynthesis in skin. Biol Pharm Bull. 31:946–948. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Niero EL and Machado-Santelli GM: Cinnamic
acid induces apoptotic cell death and cytoskeleton disruption in
human melanoma cells. J Exp Clin Cancer Res. 32:312013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsai CM, Sun FM, Chen YL, Hsu CL, Yen GC
and Weng CJ: Molecular mechanism depressing PMA-induced invasive
behaviors in human lung adenocarcinoma cells by cis- and
trans-cinnamic acid. Eur J Pharm Sci. 48:494–501. 2013. View Article : Google Scholar
|
|
28
|
Wheatley NC, Andrews KT, Tran TL, Lucke
AJ, Reid RC and Fairlie DP: Antimalarial histone deacetylase
inhibitors containing cinnamate or NSAID components. Bioorg Med
Chem Lett. 20:7080–7084. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Halley F, Reinshagen J, Ellinger B, Wolf
M, Niles AL, Evans NJ, Kirkland TA, Wagner JM, Jung M, Gribbon P
and Gul S: A bioluminogenic HDAC activity assay: validation and
screening. J Biomol Screen. 16:1227–1235. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang LT, Liou JP, Li YH, Liu YM, Pan SL
and Teng CM: A novel class I HDAC inhibitor, MPT0G030, induces cell
apoptosis and differentiation in human colorectal cancer cells via
HDAC1/PKCδ and E-cadherin. Oncotarget. 5:5651–5662. 2014.
View Article : Google Scholar : PubMed/NCBI
|