Malignant melanoma (MM) is caused by the
transformation of pigment-producing melanocytes into cancer cells.
It commonly develops in the basal layer of the skin and mucous
membranes, but it can also occur in the uvea, meninges,
gastrointestinal tract, and other tissues (1). The incidence of MM has continued to
rise worldwide, and the 5-year survival rate is only 25% (2–4). Major
treatment options include surgical resection, immunotherapy, and
targeted therapy (4,5). Since MM is prone to lymphatic
metastasis at the early stages, the 12-month progression-free
survival rate after targeted therapy is ~35%, and the median
overall survival is 23 months (6).
Therefore, the remission time and the overall survival rate do not
demonstrate significant improvement (5–8).
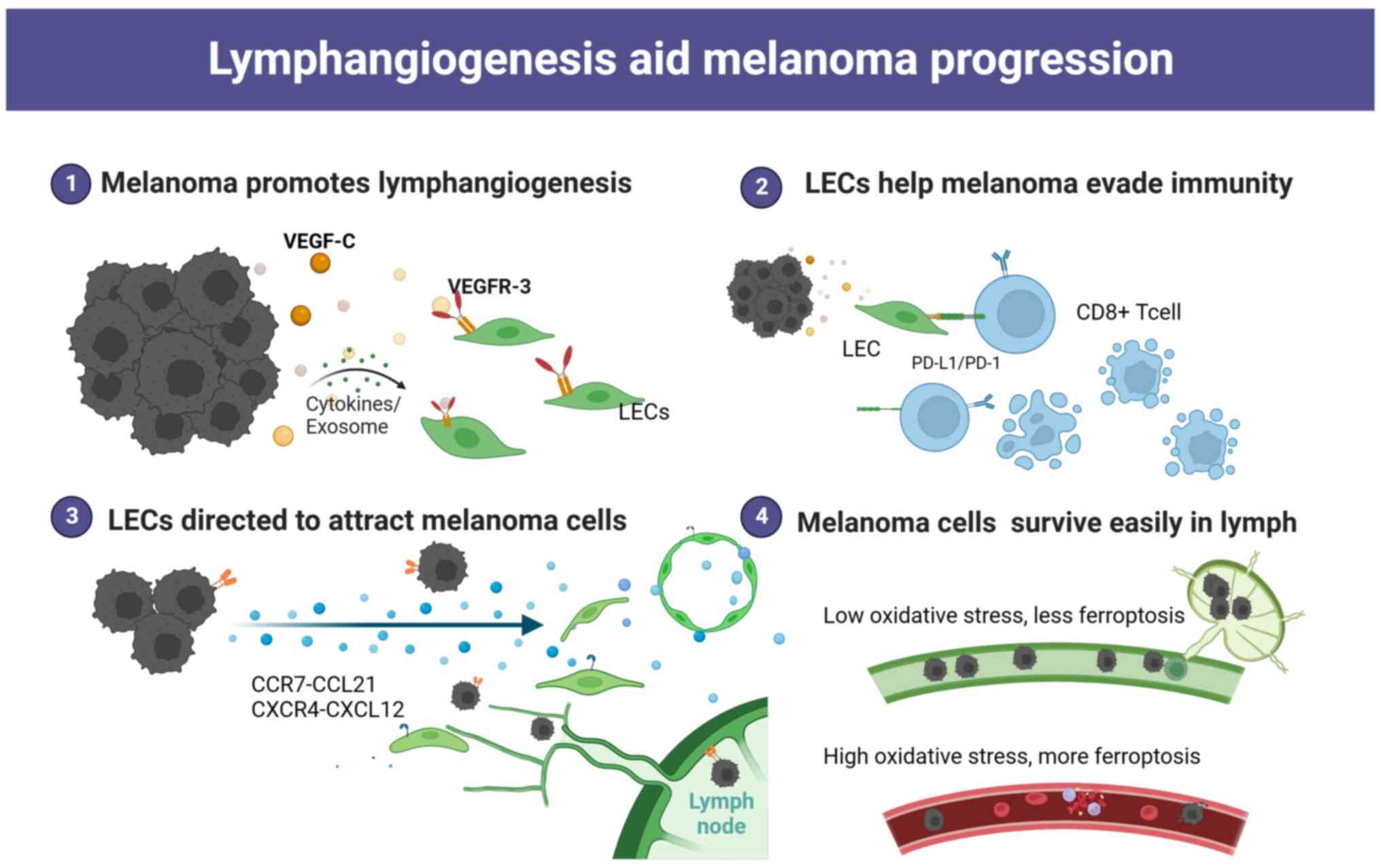
Lymphatic vessel density (LVD) in patients with MM was positively
associated with a poor prognosis (9,10).
This means that when the number of lymph vessels (LVs) adjacent to
the tumour is higher, the probability of tumour metastasis is
higher. At the same time, LVD is also associated with
tumour-infiltrating lymphocytes (TILs), which have predictive value
for therapy. When the number of paracancerous LVs increases, the
number of TILs in the tissue also increases. The increased TIL
number could have a better response to immunotherapy in MM
research, which is beneficial to the prognosis of patients
(6,7). Therefore, lymphatic endothelial cells
(LECs) not only serve as a suitable conduit for metastasis but also
play a multifaceted role in the metastasis of MM and the host
immune response. In the present review, the interactions between
melanoma cells (MMCs) and LECs during MM metastasis were
discussed.
The lymphatic system is a unidirectional circulatory
network involved in maintaining tissue fluid balance, absorbing
lipids and conducting immune surveillance (11). Capillary lymphatic vessels (CLVs) of
blind-ended origin are permeable, with only a loosely single
endothelial layer (12,13). Thin fibrous structures attached to
the surfaces of LECs on CLVs can sense interstitial pressure,
further increasing the permeability of CLVs in inflammatory or
tumour situations (11). All of the
aforementioned features make nascent CLVs the best channel for
tumour cells to invade. In addition, a high endothelial venules
(HEVs) of the lymph node (LN) also serves as a convenient exit for
metastatic MMCs (13). In a mouse
model, B16 melanoma was found to enter the blood circulation
through LNs and finally metastasize to the lung (14).
It is well documented that LECs have multiple
functions, including the secretion of chemokines that actively
recruit cells. Single-cell sequencing of LNs confirmed the presence
of six functional types of LECs mapped to specific locations,
lining the floor and ceiling of the subcapsular sinuses (SCSs),
medullary sinuses (MSs), and valve, expressing different chemokines
(15–17). LECs of the SCSs and MSs express high
levels of neutrophil chemoattractants to maintain chemokine
signaling gradients (15,16). Chemokines and their ligands play
crucial roles in leukocyte trafficking and are involved in the
metastasis of cancer to specific organs. In MM, LECs serve as the
primary source for the chemokine CCL21. This chemokine attracts
CCR7+ MMCs towards CCL21-expressing LECs but not blood
endothelial cells (18,19). As a result, it further promotes LN
metastasis. Secreting CXCL12 by tumour-associated LECs at
metastatic sites has been reported to attract MMCs expressing CXCR4
and promote the growth of metastases. This process can also convert
tumour immunogenicity into immune tolerance, thereby promoting
tumour progression (20,21). Production of the chemokine CCL1 by
the lymphatic sinus within the LN mediates the entry of MMCs
expressing CCR8 into the LNs. Blocking CCR8 has been shown to
reduce LN metastasis (22). At the
same time, high expression of the chemokines CXCL5, CXCR3 and CXCR4
has been found in a variety of melanoma experiments (23–25).
Further blockade of these chemokines or their receptors with
antagonists or neutralizing antibodies reduced the metastasis of
MMCs (23–25).
A previous study showed that lymph from patients
with MM is a rich source of tumour-derived factors including
melanoma biomarkers such as LDH, S100B and S100A8,
metastasis-associated proteins such as CSF-1, galectin-3, MMP-2 and
MMP-9, tumor-derived factors such as IL-6, IL-8, IL-1β, IL-4,
IL-10, TNF-α and extracellular vesicles. This can offer a valuable
proteomic signatures in comparison to plasma contents (26). Low levels of free iron along with
high levels of glutathione and oleic acid in lymph protect MMCs
from oxidative stress and ferroptosis, thereby preventing
subsequent metastasis (27,28). Compared with the highly oxidized
state in the blood, the lymphatic circulation provides a more
suitable environment for the survival and colonization of MMCs
(27). When MMCs were injected into
mice, the efficiency of tumour cell metastasis following intranodal
injection was notably higher compared with that following
intravenous injection (27). The
efficiency of intravenous metastasis also increased after MMCs had
been disposed of in lymph. The lymphatic system, to some extent,
provides a favorable environment for tumour metastasis.
To facilitate invasion and metastasis, MMCs,
tumour-associated macrophages (TAMs) and stromal cells in the
tumour microenvironment can release multiple cytokines that promote
the proliferation of LECs and induce lymphangiogenesis (29–31).
There is a large number of micro-LVs in MM paracancerous tissues,
and quantitative studies have shown that the mean LVD in MM nests
and paracancerous areas is 6.3 and 12.5 per mm2, respectively
(32,33). The LVD in the paracancerous region
is notably higher than that in central areas, and the LVD in
metastatic MM is higher than that in primary MM (34,35).
Although the link between lymphangiogenesis and metastasis has
received strong support, the precise molecular mechanisms driving
tumour lymphangiogenesis remain poorly understood.
VEGF-C/D-VEGFR3 is the most prominent and
well-investigated signaling pathway that plays an important role in
lymphangiogenesis. VEGF-C and VEGF-D are growth factors that
stimulate LEC proliferation and lymphatic remodeling. These factors
have been found to be upregulated in MMCs (36,37).
Inhibition of lymphangiogenesis by blocking VEGFR-3 or VEGF-C/D
could reduce LN colonization and distant metastasis (38,39).
In addition to VEGF family members, CXCL5 is
upregulated in T4-stage MMCs, leading to a notable increase in
lymphangiogenesis (25). Other
factors that can also directly or indirectly promote
lymphangiogenesis include angiopoietins, SRY-box transcription
factor 18, fibroblast growth factor and epidermal growth factor
(10,40,41).
Under physiological conditions, LVs control immune
cell trafficking and initiate the immune response to inhibit tumour
progression. However, tumour-associated LECs display a remarkable
degree of phenotypic plasticity that regulates immunotolerance
(Fig. 1). LECs isolated from highly
metastatic tumours showed a unique expression profile and
transcriptional program compared with those from non-tumour tissues
(50). Characterization of the
tumour-associated LEC secretome by RNA sequencing and cytokine
array revealed that IL6 is one of the most markedly regulated
molecules in promoting primary tumour growth, and its production is
negligible in unexposed LECs (51).
In several MM models, LECs maintain peripheral
tolerance by directly upregulating PD-L1, which interacts with PD-1
on T cells to inhibit autoreactive T cells (52). The observed upregulation of PD-L1 on
LECs may be stimulated by IFN-γ released in the tumour
microenvironment (52,53). LECs are capable of scavenging and
cross-presenting tumour-associated antigens on MHC-I, which in turn
causes dysfunctional activation of CD8+ T cells
(54). It was shown that LECs in LN
could cross-present the exogenous tumour antigen OVA to
CD8+ T cells, and naive LECs scavenge and cross-present
OVA in vitro, leading to loss of function in a B16F10
melanoma model (55). In other
aspects, LECs play an immunosuppressive role by dampening dendritic
cell (DC) maturation, thereby reducing the ability of DCs to
activate effector T cells (56,57).
LECs upregulate the expression of MHC-II in human melanoma
specimens and depend on IFN-γ to promote Treg proliferation and
exert immunosuppressive effects in the tumour microenvironment
(54,58–60).
All results suggest that LECs could play a critical role in
developing an immunosuppressive environment.
MM is an immunogenic malignancy, and there have been
attempts to exploit this specificity to develop novel therapeutic
strategies. A study found a notable number of immune cells,
including different subsets of T cells, DCs, lymphocytes,
macrophages, neutrophils and other cells, infiltrating around the
MM tissue. This infiltration may be attributed to tumour-host
interactions (61). There is a
growing interest in the antitumor immune response exerted by
tumour-infiltrating immune cells (TIICs) (62–64).
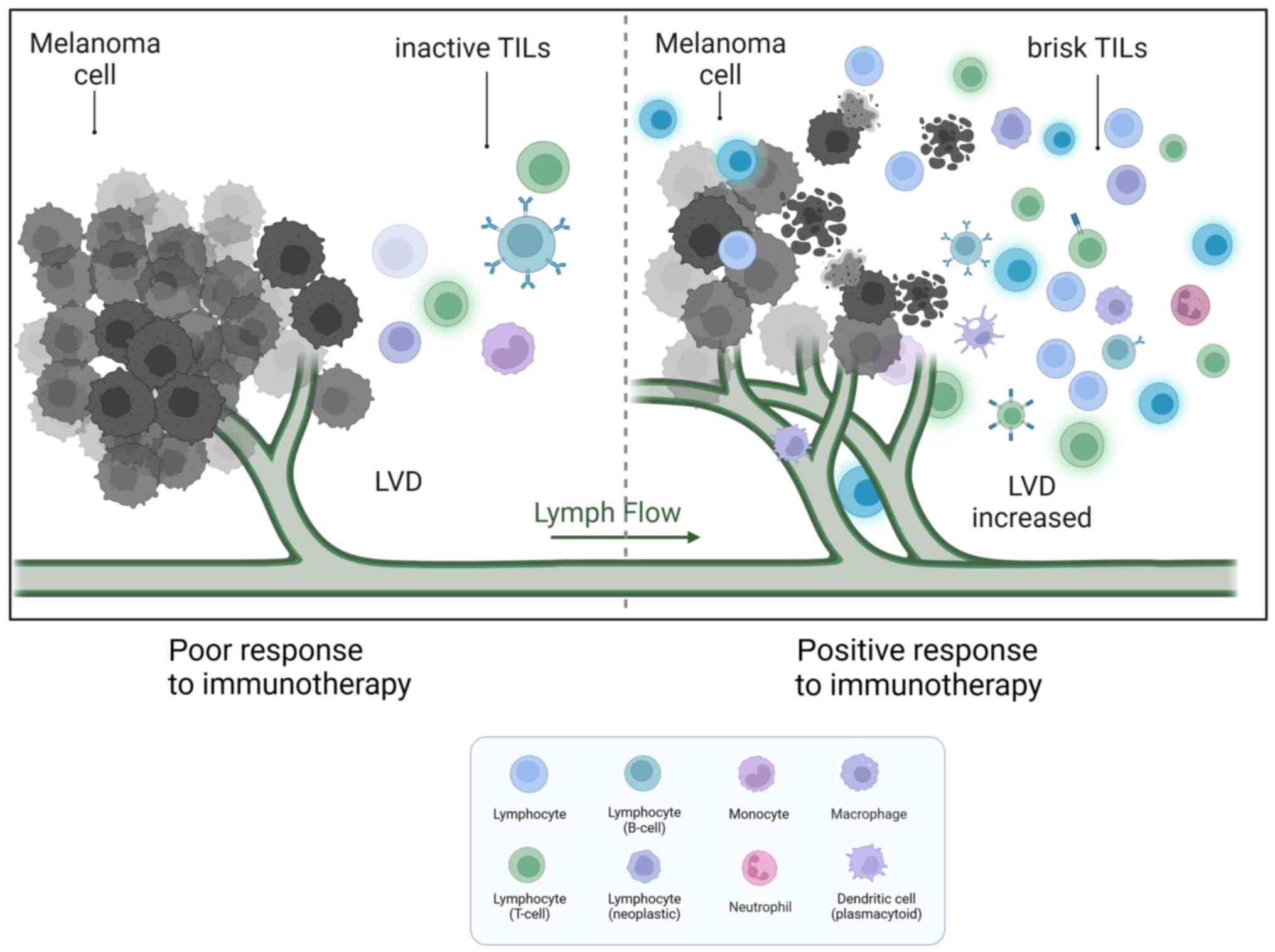
TIIC proportion notably varies among different individuals. As a
result, a certain percentage of patients with melanoma do not
respond to checkpoint immunotherapy (Fig. 2). A study cohort of 2,624 patients
with cutaneous melanoma found that TILs were an important
histopathological characteristic reflecting host immune response. A
total of 16.5% of patients had no TILs, 73.0% had inactive TILs and
10.4% had active TILs. The 5-year survival rate was 71.0% among
patients without TILs and 85.2% among patients with brisk TILs.
Brisk TILs were notably associated with improved overall survival
(OS) (65). The presence of various
subpopulations of TIICs has also been reported to predict patient
response to immune checkpoint therapy.
MMCs manipulate their heterogeneity and plasticity
in some recurrent cases, leading to the loss of expression of
multiple tumour antigens or complete loss of HLA class I expression
which allows them to evade functional antigen-specific immune
recognition (66–70). The effectiveness of T-cell
cytotoxicity requires proper antigen presentation by DCs.
Insufficiently presented antigens cannot activate T cells and
induce immunological ignorance. Altered expression of MHC-I is
frequently observed on MMCs, which allows them to evade recognition
by NK cells and reduces their cytolytic activity (70). Mediators including IL-8, IL-10,
TGF-1 and VEGF released by MMCs or TAMs limit the maturation of
normal DCs and, as a result, hinder their ability to present and
activate CD8+ T cells (70,71).
Activation of negative immune checkpoint molecules
on MMCs shields them from immune attacks and enables further
proliferation. PD-L1 is expressed at high levels on MMCs (72), and the combination of PD-1 and PD-L1
can initiate CD8+ T-cell apoptosis and stimulate the
differentiation of CD4+ T cells into Tregs, allowing
tumour cells to evade the immune system (72). CD8+ T cells/Tregs in the
tumour microenvironment could be used to predict the survival of
patients with MM (73). Increased
expression and higher affinity of cytotoxic T-lymphocyte associated
protein 4 (CTLA-4) molecules on activated effector T cells in MM
can prevent CD28 receptor binding to B7.1 and B7.2 on antigen
presenting cells, leading to T lymphocyte deactivation (74). M2 macrophages have low
antigen-presenting activity, inhibit CD8+ T-cell and NK
cell activity, and promote tumour cell migration (44,75).
In addition, Tregs are deregulated in MM and suppress the immune
system by overproducing TGF-β, IL-10 and IDO. This excessive
production hampers the function of CD4+ and
CD8+ T cells, as well as NK cells (44). MMC-derived exosomes also carry
PD-L1, which can bind to T cells to suppress antitumor immunity
(31,46,76).
Although increased LVs can promote MM spread,
increased tumour-associated LECs can assist MMCs in escaping host
immunity. The paradox is that tumour lymphangiogenesis promotes
T-cell infiltration and potentiates immunotherapy in melanoma
(77,78). There were strong positive
associations between the expression levels of the lymphatic genes
PDPN, LYVE1 and VEGFC, and the immune cell-specific genes CD45,
CD11B, F480, KLRB1, CD3D, CD8A, CD4 and FOXP3 in human metastatic
melanoma (79). Immune cell
infiltration was notably reduced after MM implantation in a mouse
model lacking dermal CLVs. In addition, inflammatory cytokines were
markedly lower, and MM was found to be more susceptible to
CD8+ T-cell attack. These findings suggest that
lymphangiogenesis in MM is associated with immunosuppression
(80). In human metastatic
melanoma, the expression of VEGF-C showed a strong association with
CCL21 and T-cell inflammation. Additionally, serum concentrations
of VEGF-C were found to be associated with both T-cell activation
and expansion (77,80).
The formation of ectopic lymphatic aggregates found
in tumour or inflammatory tissues is defined as tertiary lymphoid
structures (TLS). They are anatomically similar to LNs and contain
T-cell areas, germinal centers with proliferating B cells, and high
endothelial venules (HEVs), among others (81). TLS are typically found in the
paracancerous or stromal regions rather than the core of the
tumour. They also express chemokines, adhesion molecules, and
integrins such as intercellular adhesion molecule (ICAM) 2, ICAM3,
vascular cell adhesion molecule 1 and integrins (αL, α4, and αD),
as well as CCL21, CXCL13, CCL17, CCL22 and IL-16 to facilitate
lymphocyte recruitment (81). In
human MM, it has been found that the presence of functional TLS
activates local anti-melanoma immune responses and generally
indicates a positive prognosis. The histological evaluation
highlighted that metastatic melanoma contains B-cell lymphoid
follicles, indicating the presence of complete TLS. By contrast,
primary melanoma does not contain B-cell lymphoid follicles in TLS
(82,83). Other studies have shown that TLS
does occur in primary melanomas, although at a lower frequency
compared with what has been reported in metastases (83–85).
Using clinical samples of metastatic melanomas, it was found that
B-cell markers in TLS were the most differentially expressed genes
in distinguishing patients with MM with and without
immunotherapeutic response, as determined by bulk RNA sequencing.
Additionally, TLS also influence various T-cell phenotypes and play
a crucial role in enhancing the survival of patients with MM
(85,86). Therefore, the induction of
B-cell-rich TLS formation to enhance the tumour response to
immunotherapy can be explored as a novel strategy for treating
MM.
During MM progression, MMCs reduce the expression of
tumour-associated antigens to avoid their presentation and
recognition. However, they also express multiple inhibitory
antigens to induce immune tolerance when interacting with immune
cells. MM could enhance access to tumour drainage by stimulating
lymphangiogenesis through various mechanisms, which leads to
increased proliferation of LECs and elevated paracancerous LVD. In
the tumour microenvironment, the increased number of
tumour-associated LECs recruits MMCs and immune cells, assisting
MMCs in evading immune surveillance. TILs were found to have a
positive association with LVD and played a role in enhancing the
effectiveness of immunotherapy in MM.
In summary, increased LVD in MM promotes tumour
drainage and increases TILs. While increased drainage can lead to a
poor prognosis, TILs enhance the patients' response to
immunotherapy and improve OS. The interaction between
lymphangiogenesis and MM is complex and dynamic, and the precise
mechanisms remain an open question. It may also be extended to
other malignant tumours that are prone to lymphatic metastasis,
such as breast cancer and squamous cell carcinoma.
Not applicable.
The present review was supported by the National Natural Science
Foundation of China (grant no. 81641174), the Jiangsu Province
Postgraduate Practice Innovation Program (grant no. SJCX22_1879),
the National Natural Science Foundation of China (grant no.
82173380), the Doctoral Program for Entrepreneurship and Innovation
of Jiangsu Province (grant no. JSSCBS20211596) and the Social
Development Project of Zhenjiang Key Research and Development
Program (grant no. SH2021073).
Not applicable.
WJ, XHY, ZXY conceived and designed the study. WJ,
HHC, WZ, DML, WZ collected information. WJ and HHC drew images and
wrote the manuscript. All authors have read and approved the final
version of the manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Hussein Al-Janabi M, Mohammad JG, Mohsen
AY, Saad A and Issa R: Metastatic melanoma to the gallbladder
presented as a polyp with acute cholecystitis: A case report from
Syria. Ann Med Surg (Lond). 76:1035142022.PubMed/NCBI
|
|
2
|
Eddy K and Chen S: Overcoming immune
evasion in melanoma. Int J Mol Sci. 21:89842020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schadendorf D, van Akkooi ACJ, Berking C,
Griewank KG, Gutzmer R, Hauschild A, Stang A, Roesch A and Ugurel
S: Melanoma. Lancet. 392:971–984. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Falk Delgado A, Zommorodi S and Falk
Delgado A: Sentinel lymph node biopsy and complete lymph node
dissection for melanoma. Curr Oncol Rep. 21:542019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hartman RI and Lin JY: Cutaneous
melanoma-a review in detection, staging, and management. Hematol
Oncol Clin North Am. 33:25–38. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ribas A, Hamid O, Daud A, Hodi FS, Wolchok
JD, Kefford R, Joshua AM, Patnaik A, Hwu WJ, Weber JS, et al:
Association of pembrolizumab with tumor response and survival among
patients with advanced melanoma. JAMA. 315:1600–1609. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Knackstedt T, Knackstedt RW, Couto R and
Gastman B: Malignant melanoma: Diagnostic and management update.
Plast Reconstr Surg. 142:202e–216e. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Faries MB, Thompson JF, Cochran AJ,
Andtbacka RH, Mozzillo N, Zager JS, Jahkola T, Bowles TL, Testori
A, Beitsch PD, et al: Completion dissection or observation for
sentinel-node metastasis in melanoma. N Engl J Med. 376:2211–2222.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pasquali S, van der Ploeg APT, Mocellin S,
Stretch JR, Thompson JF and Scolyer RA: Lymphatic biomarkers in
primary melanomas as predictors of regional lymph node metastasis
and patient outcomes. Pigment Cell Melanoma Res. 26:326–337. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ma Q, Dieterich LC, Ikenberg K, Bachmann
SB, Mangana J, Proulx ST, Amann VC, Levesque MP, Dummer R, Baluk P,
et al: Unexpected contribution of lymphatic vessels to promotion of
distant metastatic tumor spread. Sci Adv. 4:eaat47582018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Oliver G, Kipnis J, Randolph GJ and Harvey
NL: The lymphatic vasculature in the 21st century: Novel functional
roles in homeostasis and disease. Cell. 182:270–296. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Petrova TV and Koh GY: Organ-specific
lymphatic vasculature: From development to pathophysiology. J Exp
Med. 215:35–49. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Johnson LA: In sickness and in health: The
immunological roles of the lymphatic system. Int J Mol Sci.
22:44582021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pereira ER, Kedrin D, Seano G, Gautier O,
Meijer EFJ, Jones D, Chin SM, Kitahara S, Bouta EM, Chang J, et al:
Lymph node metastases can invade local blood vessels, exit the
node, and colonize distant organs in mice. Science. 359:1403–1407.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Takeda A, Hollmén M, Dermadi D, Pan J,
Brulois KF, Kaukonen R, Lönnberg T, Boström P, Koskivuo I, Irjala
H, et al: Single-cell survey of human lymphatics unveils marked
endothelial cell heterogeneity and mechanisms of homing for
neutrophils. Immunity. 51:561–572.e5. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rodda LB, Lu E, Bennett ML, Sokol CL, Wang
X, Luther SA, Barres BA, Luster AD, Ye CJ and Cyster JG:
Single-cell RNA sequencing of lymph node stromal cells reveals
niche-associated heterogeneity. Immunity. 48:1014–1028.e6. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fujimoto N, He Y, D'Addio M, Tacconi C,
Detmar M and Dieterich LC: Single-cell mapping reveals new markers
and functions of lymphatic endothelial cells in lymph nodes. PLoS
Biol. 18:e30007042020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang L, Zhu L, Yao X, Lou X, Wan J, Duan
X, Pan L, Li A, Gu Z, Wang M, et al: Paclitaxel treatment enhances
lymphatic metastasis of B16F10 melanoma cells via CCL21/CCR7 axis.
Int J Biol Sci. 18:1476–1490. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cristiani CM, Turdo A, Ventura V, Apuzzo
T, Capone M, Madonna G, Mallardo D, Garofalo C, Giovannone ED,
Grimaldi AM, et al: Accumulation of circulating CCR7+
natural killer cells marks melanoma evolution and reveals a
CCL19-dependent metastatic pathway. Cancer Immunol Res. 7:841–852.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mendt M and Cardier JE: Activation of the
CXCR4 chemokine receptor enhances biological functions associated
with B16 melanoma liver metastasis. Melanoma Res. 27:300–308. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
McConnell AT, Ellis R, Pathy B, Plummer R,
Lovat PE and O'Boyle G: The prognostic significance and impact of
the CXCR4-CXCR7-CXCL12 axis in primary cutaneous melanoma. Br J
Dermatol. 175:1210–1220. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Korbecki J, Grochans S, Gutowska I,
Barczak K and Baranowska-Bosiacka I: CC chemokines in a tumor: A
review of pro-cancer and anti-cancer properties of receptors CCR5,
CCR6, CCR7, CCR8, CCR9, and CCR10 ligands. Int J Mol Sci.
21:76192020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Alimohammadi M, Rahimi A, Faramarzi F,
Alizadeh-Navaei R and Rafiei A: Overexpression of chemokine
receptor CXCR4 predicts lymph node metastatic risk in patients with
melanoma: A systematic review and meta-analysis. Cytokine.
148:1556912021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Doron H, Amer M, Ershaid N, Blazquez R,
Shani O, Lahav TG, Cohen N, Adler O, Hakim Z, Pozzi S, et al:
Inflammatory activation of astrocytes facilitates melanoma brain
tropism via the CXCL10-CXCR3 signaling axis. Cell Rep.
28:1785–1798.e6. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Soler-Cardona A, Forsthuber A, Lipp K,
Ebersberger S, Heinz M, Schossleitner K, Buchberger E, Gröger M,
Petzelbauer P, Hoeller C, et al: CXCL5 facilitates melanoma
cell-neutrophil interaction and lymph node metastasis. J Invest
Dermatol. 138:1627–1635. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Broggi MAS, Maillat L, Clement CC, Bordry
N, Corthésy P, Auger A, Matter M, Hamelin R, Potin L, Demurtas D,
et al: Tumor-associated factors are enriched in lymphatic exudate
compared to plasma in metastatic melanoma patients. J Exp Med.
216:1091–1107. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ubellacker JM, Tasdogan A, Ramesh V, Shen
B, Mitchell EC, Martin-Sandoval MS, Gu Z, McCormick ML, Durham AB,
Spitz DR, et al: Lymph protects metastasizing melanoma cells from
ferroptosis. Nature. 585:113–118. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Piskounova E, Agathocleous M, Murphy MM,
Hu Z, Huddlestun SE, Zhao Z, Leitch AM, Johnson TM, DeBerardinis RJ
and Morrison SJ: Oxidative stress inhibits distant metastasis by
human melanoma cells. Nature. 527:186–191. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Habenicht LM, Kirschbaum SB, Furuya M,
Harrell MI and Ruddell A: Tumor regulation of lymph node lymphatic
sinus growth and lymph flow in mice and in humans. Yale J Biol Med.
90:403–415. 2017.PubMed/NCBI
|
|
30
|
Peppicelli S, Bianchini F and Calorini L:
Inflammatory cytokines induce vascular endothelial growth factor-C
expression in melanoma-associated macrophages and stimulate
melanoma lymph node metastasis. Oncol Lett. 8:1133–1138. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Leary N, Walser S, He Y, Cousin N, Pereira
P, Gallo A, Collado-Diaz V, Halin C, Garcia-Silva S, Peinado H and
Dieterich LC: Melanoma-derived extracellular vesicles mediate
lymphatic remodelling and impair tumour immunity in draining lymph
nodes. J Extracell Vesicles. 11:e121972022. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dadras SS, Paul T, Bertoncini J, Brown LF,
Muzikansky A, Jackson DG, Ellwanger U, Garbe C, Mihm MC and Detmar
M: Tumor lymphangiogenesis: A novel prognostic indicator for
cutaneous melanoma metastasis and survival. Am J Pathol.
162:1951–1960. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pastushenko I, Van den Eynden GG,
Vicente-Arregui S, Prieto-Torres L, Alvarez-Alegret R, Querol I,
Dirix LY, Carapeto FJ, Vermeulen PB and Van Laere SJ: Increased
angiogenesis and lymphangiogenesis in metastatic sentinel lymph
nodes is associated with nonsentinel lymph node involvement and
distant metastasis in patients with melanoma. Am J Dermatopathol.
38:338–346. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ayubi E and Safiri S: Lymphatic vessel
density and VEGF-C expression as independent predictors of melanoma
metastases: Methodological issues. J Plast Reconstr Aesthet Surg.
71:604–605. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pastushenko I, Vermeulen PB, Carapeto FJ,
Van den Eynden G, Rutten A, Ara M, Dirix LY and Van Laere S: Blood
microvessel density, lymphatic microvessel density and lymphatic
invasion in predicting melanoma metastases: Systematic review and
meta-analysis. Br J Dermatol. 170:66–77. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Špirić Z, Eri Ž and Erić M: Lymphatic
vessel density and VEGF-C expression as independent predictors of
melanoma metastases. J Plast Reconstr Aesthet Surg. 70:1653–1659.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Skobe M, Hamberg LM, Hawighorst T,
Schirner M, Wolf GL, Alitalo K and Detmar M: Concurrent induction
of lymphangiogenesis, angiogenesis, and macrophage recruitment by
vascular endothelial growth factor-C in melanoma. Am J Pathol.
159:893–903. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang M, Xu Y, Wen GZ, Wang Q and Yuan SM:
Rapamycin suppresses angiogenesis and lymphangiogenesis in melanoma
by downregulating VEGF-A/VEGFR-2 and VEGF-C/VEGFR-3 expression.
Onco Targets Ther. 12:4643–4654. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lee JY, Hong SH, Shin M, Heo HR and Jang
IH: Blockade of FLT4 suppresses metastasis of melanoma cells by
impaired lymphatic vessels. Biochem Biophys Res Commun.
478:733–738. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Korhonen EA, Murtomäki A, Jha SK, Anisimov
A, Pink A, Zhang Y, Stritt S, Liaqat I, Stanczuk L, Alderfer L, et
al: Lymphangiogenesis requires Ang2/Tie/PI3K signaling for VEGFR3
cell-surface expression. J Clin Invest. 132:e1554782022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rezzola S, Sigmund EC, Halin C and Ronca
R: The lymphatic vasculature: An active and dynamic player in
cancer progression. Med Res Rev. 42:576–614. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wouters J, Kalender-Atak Z, Minnoye L,
Spanier KI, De Waegeneer M, Bravo González-Blas C, Mauduit D, Davie
K, Hulselmans G, Najem A, et al: Robust gene expression programs
underlie recurrent cell states and phenotype switching in melanoma.
Nat Cell Biol. 22:986–998. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Arozarena I and Wellbrock C: Phenotype
plasticity as enabler of melanoma progression and therapy
resistance. Nat Rev Cancer. 19:377–391. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Reticker-Flynn NE, Zhang W, Belk JA, Basto
PA, Escalante NK, Pilarowski GOW, Bejnood A, Martins MM, Kenkel JA,
Linde IL, et al: Lymph node colonization induces tumor-immune
tolerance to promote distant metastasis. Cell. 185:1924–1942.e23.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
García-Silva S, Benito-Martín A, Nogués L,
Hernández-Barranco A, Mazariegos MS, Santos V, Hergueta-Redondo M,
Ximénez-Embún P, Kataru RP, Lopez AA, et al: Melanoma-derived small
extracellular vesicles induce lymphangiogenesis and metastasis
through an NGFR-dependent mechanism. Nat Cancer. 2:1387–1405. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gowda R, Robertson BM, Iyer S, Barry J,
Dinavahi SS and Robertson GP: The role of exosomes in metastasis
and progression of melanoma. Cancer Treat Rev. 85:1019752020.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wakisaka N, Hasegawa Y, Yoshimoto S, Miura
K, Shiotani A, Yokoyama J, Sugasawa M, Moriyama-Kita M, Endo K and
Yoshizaki T: Primary tumor-secreted lymphangiogenic factors induce
pre-metastatic lymphvascular niche formation at sentinel lymph
nodes in oral squamous cell carcinoma. PLoS One. 10:e01440562015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li L, Wu J, Abdi R, Jewell CM and Bromberg
JS: Lymph node fibroblastic reticular cells steer immune responses.
Trends Immunol. 42:723–734. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rovera C, Berestjuk I, Lecacheur M,
Tavernier C, Diazzi S, Pisano S, Irondelle M, Mallavialle A,
Albrengues J, Gaggioli C, et al: Secretion of IL1 by
dedifferentiated melanoma cells inhibits JAK1-STAT3-driven
actomyosin contractility of lymph node fibroblastic reticular
cells. Cancer Res. 82:1774–1788. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Clasper S, Royston D, Baban D, Cao Y,
Ewers S, Butz S, Vestweber D and Jackson DG: A novel gene
expression profile in lymphatics associated with tumor growth and
nodal metastasis. Cancer Res. 68:7293–7303. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Van de Velde M, Ebroin M, Durré T, Joiret
M, Gillot L, Blacher S, Geris L, Kridelka F and Noel A: Tumor
exposed-lymphatic endothelial cells promote primary tumor growth
via IL6. Cancer Lett. 497:154–164. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Dieterich LC, Ikenberg K, Cetintas T,
Kapaklikaya K, Hutmacher C and Detmar M: Tumor-associated lymphatic
vessels upregulate PDL1 to inhibit T-cell activation. Front
Immunol. 8:662017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lane RS, Femel J, Breazeale AP, Loo CP,
Thibault G, Kaempf A, Mori M, Tsujikawa T, Chang YH and Lund AW:
IFNγ-activated dermal lymphatic vessels inhibit cytotoxic T cells
in melanoma and inflamed skin. J Exp Med. 215:3057–3074. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Nörder M, Gutierrez MG, Zicari S, Cervi E,
Caruso A and Guzmán CA: Lymph node-derived lymphatic endothelial
cells express functional costimulatory molecules and impair
dendritic cell-induced allogenic T-cell proliferation. FASEB J.
26:2835–2846. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lund AW, Duraes FV, Hirosue S, Raghavan
VR, Nembrini C, Thomas SN, Issa A, Hugues S and Swartz MA: VEGF-C
promotes immune tolerance in B16 melanomas and cross-presentation
of tumor antigen by lymph node lymphatics. Cell Rep. 1:191–199.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
de Winde CM, Munday C and Acton SE:
Molecular mechanisms of dendritic cell migration in immunity and
cancer. Med Microbiol Immunol. 209:515–529. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Swartz MA and Lund AW: Lymphatic and
interstitial flow in the tumour microenvironment: Linking
mechanobiology with immunity. Nat Rev Cancer. 12:210–219. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Dubrot J, Duraes FV, Harlé G, Schlaeppi A,
Brighouse D, Madelon N, Göpfert C, Stokar-Regenscheit N, Acha-Orbea
H, Reith W, et al: Absence of MHC-II expression by lymph node
stromal cells results in autoimmunity. Life Sci Alliance.
1:e2018001642018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li CY, Park HJ, Shin J, Baik JE, Mehrara
BJ and Kataru RP: Tumor-associated lymphatics upregulate MHC-II to
suppress tumor-infiltrating lymphocytes. Int J Mol Sci.
23:134702022. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Lukacs-Kornek V, Malhotra D, Fletcher AL,
Acton SE, Elpek KG, Tayalia P, Collier AR and Turley SJ: Regulated
release of nitric oxide by nonhematopoietic stroma controls
expansion of the activated T cell pool in lymph nodes. Nat Immunol.
12:1096–1104. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Antohe M, Nedelcu RI, Nichita L, Popp CG,
Cioplea M, Brinzea A, Hodorogea A, Calinescu A, Balaban M, Ion DA,
et al: Tumor infiltrating lymphocytes: The regulator of melanoma
evolution. Oncol Lett. 17:4155–4161. 2019.PubMed/NCBI
|
|
62
|
Mihm MC Jr and Mulé JJ: Reflections on the
histopathology of tumor-infiltrating lymphocytes in melanoma and
the host immune response. Cancer Immunol Res. 3:827–835. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Durante MA, Rodriguez DA, Kurtenbach S,
Kuznetsov JN, Sanchez MI, Decatur CL, Snyder H, Feun LG,
Livingstone AS and Harbour JW: Single-cell analysis reveals new
evolutionary complexity in uveal melanoma. Nat Commun. 11:4962020.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Li H, van der Leun AM, Yofe I, Lubling Y,
Gelbard-Solodkin D, van Akkooi ACJ, van den Braber M, Rozeman EA,
Haanen JBAG, Blank CU, et al: Dysfunctional CD8 T cells form a
proliferative, dynamically regulated compartment within human
melanoma. Cell. 176:775–789.e18. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yang J, Lian JW, Chin YH, Wang L, Lian A,
Murphy GF and Zhou L: Assessing the prognostic significance of
tumor-infiltrating lymphocytes in patients with melanoma using
pathologic features identified by natural language processing. JAMA
Netw Open. 4:e21263372021. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Khong HT, Wang QJ and Rosenberg SA:
Identification of multiple antigens recognized by
tumor-infiltrating lymphocytes from a single patient: Tumor escape
by antigen loss and loss of MHC expression. J Immunother.
27:184–190. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Maeurer MJ, Gollin SM, Martin D, Swaney W,
Bryant J, Castelli C, Robbins P, Parmiani G, Storkus WJ and Lotze
MT: Tumor escape from immune recognition: Lethal recurrent melanoma
in a patient associated with downregulation of the peptide
transporter protein TAP-1 and loss of expression of the
immunodominant MART-1/Melan-A antigen. J Clin Invest. 98:1633–1641.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Al-Batran SE, Rafiyan MR, Atmaca A,
Neumann A, Karbach J, Bender A, Weidmann E, Altmannsberger HM,
Knuth A and Jäger E: Intratumoral T-cell infiltrates and MHC class
I expression in patients with stage IV melanoma. Cancer Res.
65:3937–3941. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Rodig SJ, Gusenleitner D, Jackson DG,
Gjini E, Giobbie-Hurder A, Jin C, Chang H, Lovitch SB, Horak C,
Weber JS, et al: MHC proteins confer differential sensitivity to
CTLA-4 and PD-1 blockade in untreated metastatic melanoma. Sci
Transl Med. 10:eaar33422018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Passarelli A, Mannavola F, Stucci LS,
Tucci M and Silvestris F: Immune system and melanoma biology: A
balance between immunosurveillance and immune escape. Oncotarget.
8:106132–106142. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Failli A, Legitimo A, Orsini G, Romanini A
and Consolini R: Numerical defect of circulating dendritic cell
subsets and defective dendritic cell generation from monocytes of
patients with advanced melanoma. Cancer Lett. 337:184–192. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Garcia-Diaz A, Shin DS, Moreno BH, Saco J,
Escuin-Ordinas H, Rodriguez GA, Zaretsky JM, Sun L, Hugo W, Wang X,
et al: Interferon receptor signaling pathways regulating PD-L1 and
PD-L2 expression. Cell Rep. 19:1189–1201. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Jacobs JFM, Nierkens S, Figdor CG, de
Vries IJM and Adema GJ: Regulatory T cells in melanoma: The final
hurdle towards effective immunotherapy? Lancet Oncol. 13:e32–e42.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Petrova V, Arkhypov I, Weber R, Groth C,
Altevogt P, Utikal J and Umansky V: Modern aspects of immunotherapy
with checkpoint inhibitors in melanoma. Int J Mol Sci. 21:23672020.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Falleni M, Savi F, Tosi D, Agape E, Cerri
A, Moneghini L and Bulfamante GP: M1 and M2 macrophages'
clinicopathological significance in cutaneous melanoma. Melanoma
Res. 27:200–210. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Chen G, Huang AC, Zhang W, Zhang G, Wu M,
Xu W, Yu Z, Yang J, Wang B, Sun H, et al: Exosomal PD-L1
contributes to immunosuppression and is associated with anti-PD-1
response. Nature. 560:382–386. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Fankhauser M, Broggi MAS, Potin L, Bordry
N, Jeanbart L, Lund AW, Da Costa E, Hauert S, Rincon-Restrepo M,
Tremblay C, et al: Tumor lymphangiogenesis promotes T cell
infiltration and potentiates immunotherapy in melanoma. Sci Transl
Med. 9:eaal47122017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Moussion C and Turley SJ: Tumour lymph
vessels boost immunotherapy. Nature. 552:340–342. 2017. View Article : Google Scholar
|
|
79
|
Lund AW, Wagner M, Fankhauser M, Steinskog
ES, Broggi MA, Spranger S, Gajewski TF, Alitalo K, Eikesdal HP,
Wiig H and Swartz MA: Lymphatic vessels regulate immune
microenvironments in human and murine melanoma. J Clin Invest.
126:3389–3402. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Bordry N, Broggi MAS, de Jonge K,
Schaeuble K, Gannon PO, Foukas PG, Danenberg E, Romano E,
Baumgaertner P, Fankhauser M, et al: Lymphatic vessel density is
associated with CD8+ T cell infiltration and
immunosuppressive factors in human melanoma. Oncoimmunology.
7:e14628782018. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Sautès-Fridman C, Petitprez F, Calderaro J
and Fridman WH: Tertiary lymphoid structures in the era of cancer
immunotherapy. Nat Rev Cancer. 19:307–325. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Helmink BA, Reddy SM, Gao J, Zhang S,
Basar R, Thakur R, Yizhak K, Sade-Feldman M, Blando J, Han G, et
al: B cells and tertiary lymphoid structures promote immunotherapy
response. Nature. 577:549–555. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Cipponi A, Mercier M, Seremet T, Baurain
JF, Théate I, van den Oord J, Stas M, Boon T, Coulie PG and van
Baren N: Neogenesis of lymphoid structures and antibody responses
occur in human melanoma metastases. Cancer Res. 72:3997–4007. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Ladányi A, Sebestyén T, Mohos A, Liszkay
G, Somlai B, Tóth E and Tímár J: Ectopic lymphoid structures in
primary cutaneous melanoma. Pathol Oncol Res. 20:981–985. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Cabrita R, Lauss M, Sanna A, Donia M,
Skaarup Larsen M, Mitra S, Johansson I, Phung B, Harbst K,
Vallon-Christersson J, et al: Tertiary lymphoid structures improve
immunotherapy and survival in melanoma. Nature. 577:561–565. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Maibach F, Sadozai H, Seyed Jafari SM,
Hunger RE and Schenk M: Tumor-infiltrating lymphocytes and their
prognostic value in cutaneous melanoma. Front Immunol. 11:21052020.
View Article : Google Scholar : PubMed/NCBI
|