Introduction
The PI3K-Akt-mTOR pathway is one of the most
frequently altered signaling cascades in breast cancer (1). This pathway is targeted in several
clinical studies (2). The inositol
pyrophosphate (IPP) IP7 is a small phosphate-rich regulator of Akt,
inhibiting membrane translocation and activation of Akt by
competing with membrane-bound phosphoinositide binding to the
pleckstrin homology domain, and thereby disturbing the pyruvate
dehydrogenase kinase 1 (PDK1) -induced phosphorylation at the
Akt-threonine 308 site (3,4). The inositol hexakisphosphate kinases
(IP6K) 1 and 2 are responsible for the conversion of IP6 to
IP7(5).
The IP6K1 and IP6K2 genes are localized at the
chromosomal site 3p21.31, which is a gene-dense region, including
several putative tumor suppressor genes and some oncogenes
(6,7). In the context of Akt inhibition, the
IP6Ks would play a tumor suppressive role as Akt inhibition slows
down tumor proliferation and increases apoptosis (8). Deletion and knock-out studies have
found diverging roles of isoforms of the two IP6Ks in tumors and
between different phases of tumor progression. IP6K1 has been
suggested to be an oncogene, stimulating early cytoskeletal
changes, migration and tumor growth (9), whereas IP6K2 has been suggested to
hold tumor suppressive features by inducing apoptosis in a
p53-dependent manner (10,11). IP6K2 was important for the response
to radiation and cytotoxic drugs in ovarian carcinoma cells, and
loss of IP6K2 predicted higher prevalence of aero-digestive
carcinoma in mice fed with a carcinogen (12,13).
We have previous data indicating that other factors
in the PI3K/Akt/mTOR pathway are altered in the Swedish low-risk
breast cancer cohort (14-16).
IP6Ks have been found in vitro to indirectly, through IP7,
inhibit Akt. The hypothesis was that the IP6K would inhibit the
PI3K/Akt/mTOR pathway and be protective factors in breast cancer.
Therefore, in this study we evaluated the role of IP6K1 and IP6K2
by analyzing their impact on outcome in three separate cohorts of
patients with primary breast cancer. Analyses of gene expression,
protein expression and subcellular localization of proteins were
included. To our knowledge, the IP6Ks have never been evaluated in
cancer patient cohorts, hence this is the first study in this
respect.
Materials and methods
Swedish low-risk cohort
Postmenopausal breast cancer patients at low risk,
with a negative node status and a tumor not exceeding three cm were
randomized to adjuvant tamoxifen, 40 mg/day for two years or no
tamoxifen (17). Patient entry to
the studies was from November 1976 to May 1990, and follow-up data
was available to December 2004. In 1983, tamoxifen-treated
recurrence-free patients were randomized, if consenting, to three
more years of tamoxifen or no tamoxifen. Patients were mainly of
Caucasian ethnicity, and all female. Median age at diagnosis was
62.7, ranging from 45.8 to 76.8. Retrospective studies on archived
tumor tissue, with the purpose to evaluate prognostic and treatment
predicting factors, were approved by the ethics committee at
Karolinska Institute in Stockholm, Sweden. Tumor tissue was
collected on surgical removal of the primary tumor and incubated in
formalin for fixation and paraffin embedded. Three cores of
abundant cancer cell content were selected to represent each tumor
on a tissue microarray (TMA). The ER, progesterone receptor (PgR)
and HER2 status were assessed as previously described (18,19).
Further data on p-Akt and p-S6K were available (14,16).
For all proteins detected, a portion of samples was missing. In the
supplementary table of a previously published paper, missing
samples were compared with the samples on TMA and with samples of
the original cohort (20). The
results show no bias in the missing cases with respect to tumor
size, ER status, or tamoxifen treatment. The present study was
designed and presented with regard to the reporting recommendations
for tumor marker prognostic studies (REMARK) guidelines (21).
Online datasets
Gene-expression data was drawn from a Dutch
retrospective publicly available dataset of 295 breast cancer
patients with a tumor not exceeding five cm at stage I and II,
including node-negative -and node-positive disease (22). Time for diagnosis was during 1984
and 1995, including patients up to the age of 52. Patients were
diagnosed in the Netherlands and were at diagnosis 52 years or
younger. Follow-up time, treatment regimens and gene-expression
data were downloaded from http://bioinformatics.nki.nl/data.php. Additional data
used for analysis was available through personal communication.
Further, prognostic impact of IP6K1 and IP6K2 gene
expression was tested in an additional breast cancer cohort, the
Cancer Genome Atlas (TCGA), previously described (23), and accessed through cBioPortal for
Cancer Genomics https://www.cbioportal.org/ (24). Patients in the TCGA cohort were both
pre-and postmenopausal. Twelve patients were male. Median age was
58, ranging from 26 to 90. Ethnicity distribution was previously
published (25).
Expression of genes at 3p21.31
Formalin fixed and paraffin embedded (FFPE) tumors
from 912 women in the Swedish low-risk group were retrieved.
Messenger RNA was later extracted from FFPE breast tumor tissue and
652 samples were available for microarray gene-expression analysis
using custom-designed arrays, containing approximately 32.1K
probes, detecting approximately 21.5K unique genes (Agilent
Technologies, CA, USA) (26). The
PAM50 intrinsic subtype analysis classifier was used as described
in Parker et al (27).
Expression levels of IP6K1 and IP6K2 were analyzed by quartiles
with low expression defined by the first quartile (Q1). High
expression was defined by the second to fourth quartile
(Q2-Q4).
The IP6K genes are localized in a gene-dense
region. We found six cancer-related genes near the IP6Ks at
3p21.31. SETD2, PTPN23, PLXNB1 and
RASSF1 are potential tumor-suppressor genes and
CDC25A and RHOA are predicted oncogenes according to
their function in tumor cells. The prognostic value of the selected
3p21.31 genes was analyzed in quartiles of gene expression, as with
the IP6Ks. The cut-off for low expression was set at Q1.
Immunohistochemical staining of IP6K1
and IP6K2 proteins
Immunohistochemical analysis was used to detect
IP6K1 protein expression in 731 samples and IP6K2 protein
expression in 724 samples from the Swedish low-risk group. Staining
was evaluated at x400 magnification by two independent observers
(J.S. and A.B.). Cytoplasmic protein expression was evaluated as
negative, weak staining, moderate and strong staining, Fig. S1A-H). For analysis of cytoplasm in
two groups, negative and weak were set as low, whereas moderate and
strong were regarded as high. Nucleus was graded as positive when
>10% of tumor cells showed staining. The PT-link station was
used for deparaffinization and antigen retrieval in a low-pH
buffer, starting at 65˚C, 96˚C for 20 min and cooled down to 65˚C
(DakoCytomation, Glostrup, Denmark). Inactivation of endogenous
peroxidase in 3% hydrogen peroxide in water was followed by
blocking in serum-free protein block for 10 min (Spring
Bioscience). TMAs were incubated in a moisturized chamber at 4˚C
overnight with the IP6K1 and IP6K2 antibodies, diluted 1:800 and
1:500, respectively (IP6K1:HPA040825, RRID:AB_10960426 and
IP6K2:HPA007532, RRID:AB_1851542, Merck KGaA). Secondary rabbit
antibody (DakoCytomation Envision+ HRP system) was applied for 30
min and protein staining was developed with 3,3'-diaminobenzidine
chromogen and substrate buffer (DakoCytomation) and counterstained
with hematoxylin. All wash steps were in phosphate buffered saline
including 0.5% bovine serum albumin. The tissue was dehydrated, and
cover glass was mounted with Pertex (Histolab). All slides were
scanned with ScanScope AT (Aperio) and the images were assessed
with Aperio Imagescope software v.12.4.3.
IP6K1 -and IP6K2 antibody
validation
Antibodies were validated by knock-down of the
specific genes in breast cancer cell lines (Fig. S2). The cell line ZR751
(RRID:CVCL_0588) was transfected with two IP6K2 siRNAs (1. s224205
and 2. s195221, Ambion by ThemoFisher Scientific, MA, USA) and
BT474 (RRID:CVCL_0179) and T47D (RRID:CVCL_0553) were transfected
with IP6K1 siRNA (s18957, Ambion by ThemoFisher Scientific).
Protein detection with western blotting showed specific bands at
the expected 50 kDa that were downregulated after siRNA
transfection. All cell lines were transfected with a scrambled
siRNA (AllStars Negative Control, 102728, Qiagen) and GAPDH siRNA
(4390849, Ambion by Themo Fisher Scientific) and GAPDH antibody
(ab185059, Abcam) served as loading control. Cell line passage
number was kept low. Cells were regularly confirmed negative for
mycoplasma and authenticated by morphology, karyotyping and
STR-assay (ATCC).
Statistical analysis
The Kaplan-Meier product limit method was used to
estimate the cumulative probabilities of distant recurrence-free
interval. For univariable and multivariable analysis of event rates
and P-values, Cox proportional hazard regression was used.
Associations between different variables were assessed by Pearson
χ2 test. P-values <0.05 were considered statistically
significant. Statistical analyses were performed with Statistica
13.5 (TIBCO Software Inc.).
Results
Low IP6K2 gene expression predicts
impaired prognosis in independent cohorts
The prognostic value of gene-expression levels was
tested in the systemically untreated subset of the Swedish low-risk
cohort with distant recurrence-free interval as end point. Patients
with high IP6K2 expression had a better outcome than patients with
low IP6K2 expression (HR: 0.55, 95% CI: 0.35-0.86, P=0.0086,
Fig. 1A). This was confirmed in a
multivariable analysis (HR: 0.48, 95% CI: 0.29-0.79, P=0.0038).
IP6K1 expression (Q1 vs. Q2-4) showed no significant prognostic
value in the univariable analysis (P=0.56), or in the multivariable
analysis correcting for tumor size, ER, PgR and HER2 status
(P=0.14, Fig. 1B). Testing IP6K
gene expression in relation to clinicopathological characteristics
and PI3K/Akt/mTOR-pathway variables we found IP6K2 gene expression
to be associated with small tumor size (P=0.019), ER (P=0.00051),
nuclear p-Akt (P=0.0058) and nuclear p-S6K (P=0.0063) and IP6K1
gene expression to be associated with ER (P=0.014), nuclear p-Akt
(P=0.0058) and with nuclear p-S6K (P=0.045). In addition, IP6K2 and
IP6K1 expression distribution according to PAM50 intrinsic subtypes
are presented (Tables I and
II). IP6K1 and IP6K2 gene
expression were strongly associated (P<0.00001).
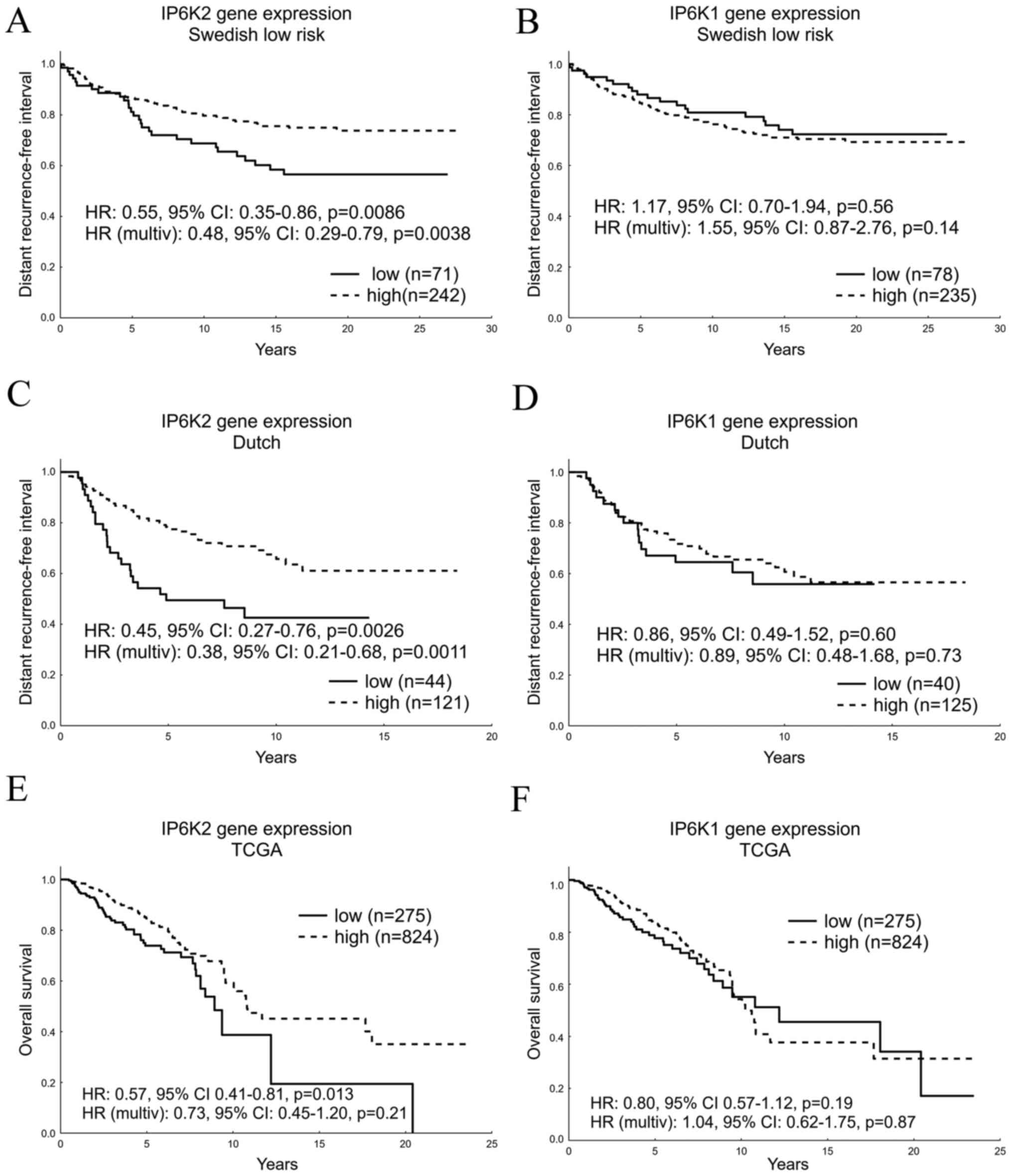 | Figure 1IP6K gene expression as a prognostic
biomarker. Gene expression levels of (A) IP6K2 and (B) IP6K1 in the
Swedish low-risk cohort with the multivariable Cox proportional
hazard model corrected for tumor size, and ER, PgR and HER2 status.
Gene expression levels of (C) IP6K2 and (D) IP6K1 in the Dutch
cohort with the multivariable model corrected for node, ER and HER2
status. Prognostic data was produced from systemically untreated
patients with primary tumors of the two cohorts. Gene expression
levels of (E) IP6K2 and (F) IP6K1 in the TCGA cohort with the
multivariable model corrected for node, ER, PgR, HER2 and TP53
status. IP6K, inositol hexakisphosphate kinase; ER, estrogen
receptor; PgR, progesterone receptor; HR, hazard ratio; TCGA, The
Cancer Genome Atlas; multiv, multivariate. |
 | Table IIP6K2 gene and protein expression,
clinicopathological variables and PI3K/Akt/mTOR-related variables
in the Swedish low-risk cohort. |
Table I
IP6K2 gene and protein expression,
clinicopathological variables and PI3K/Akt/mTOR-related variables
in the Swedish low-risk cohort.
| | IP6K2 mRNA | Cytoplasmic
IP6K2 | Nuclear IP6K2 |
|---|
| Variables | Low, n (%) | High, n (%) | P-value | Low, n (%) | High, n (%) | P-value | Negative, n
(%) | Positive, n
(%) | P-value |
|---|
| Tamoxifen | | | | | | | | | |
|
No
tamoxifen | 71(23) | 242(77) | | 61(17) | 289(83) | | 296(85) | 54(15) | |
|
Tamoxifen | 93(27) | 247(73) | 0.169 | 87(23) | 286(77) | 0.050 | 315(84) | 58(16) | 0.964 |
| Size, mm | | | | | | | | | |
|
<20 | 114(23) | 386(77) | | 116(21) | 433(79) | | 451(82) | 98(18) | |
|
>20 | 47(32) | 98(68) | 0.019 | 27(17) | 131(83) | 0.265 | 147(93) | 11(7) | <0.001 |
| ER | | | | | | | | | |
|
Negative | 47(37) | 81(63) | | 25(16) | 133(84) | | 139(88) | 19(12) | |
|
Positive | 112(22) | 400(78) | <0.001 | 121(22) | 427(78) | 0.087 | 459(84) | 89(16) | 0.195 |
| PgR | | | | | | | | | |
|
Negative | 74(28) | 194(72) | | 53(18) | 248(82) | | 262(87) | 39(13) | |
|
Positive | 74(23) | 247(77) | 0.204 | 81(24) | 260(76) | 0.056 | 295(87) | 46(13) | 0.842 |
| HER2 | | | | | | | | | |
|
Negative | 132(24) | 408(76) | | 127(22) | 461(78) | | 494(84) | 94(16) | |
|
Positive | 20(34) | 38(66) | 0.095 | 10(13) | 68(87) | 0.072 | 72(92) | 6(8) | 0.054 |
| Cyto p-Akt | | | | | | | | | |
|
Negative | 52(24) | 164(76) | | 71(26) | 202(74) | | 220(81) | 53(19) | |
|
Positive | 106(28) | 272(72) | 0.292 | 73(17) | 354(83) | 0.004 | 373(87) | 54(13) | 0.015 |
| Nucl p-Akt | | | | | | | | | |
|
Negative | 87(32) | 188(68) | | 67(22) | 237(78) | | 272(89) | 32(11) | |
|
Positive | 71(22) | 248(78) | 0.010 | 77(19) | 319(81) | 0.400 | 321(81) | 75(19) | 0.002 |
| Cyto p-S6K | | | | | | | | | |
|
Negative | 58(23) | 191(77) | | 81(27) | 215(73) | | 247(83) | 49(17) | |
|
Positive | 95(27) | 252(73) | 0.260 | 63(16) | 331(84) | <0.001 | 340(86) | 54(14) | 0.299 |
| Nucl p-S6K | | | | | | | | | |
|
Negative | 110(29) | 263(71) | | 85(19) | 352(81) | | 380(87) | 57(13) | |
|
Positive | 43(19) | 179(81) | 0.006 | 59(23) | 193(77) | 0.218 | 206(82) | 46(18) | 0.065 |
| PAM50 | | | | | | | | | |
|
Basal
like | 25(40) | 38(60) | | 5(9) | 48(91) | | 48(91) | 5(9) | |
|
Luminal
B | 35(27) | 94(73) | | 23(20) | 90(80) | | 105(93) | 8(7) | |
|
HER2
enriched | 21(36) | 37(64) | | 9(18) | 41(82) | | 46(92) | 4(8) | |
|
Luminal
A | 73(21) | 269(79) | | 55(19) | 229(81) | | 239(84) | 45(16) | |
|
Normal
like | 10(17) | 50(83) | 0.003 | 11(24) | 34(76) | 0.377 | 34(76) | 11(24) | 0.017 |
| Table IIIP6K1 gene and protein expression,
clinicopathological variables and PI3K/Akt/mTOR-related variables
in the Swedish low-risk cohort. |
Table II
IP6K1 gene and protein expression,
clinicopathological variables and PI3K/Akt/mTOR-related variables
in the Swedish low-risk cohort.
| | IP6K1 mRNA | Cytoplasmic
IP6K1 | Nuclear IP6K1 |
|---|
| Variables | Low, n (%) | High, n (%) | P-value | Low, n (%) | High, n (%) | P-value | Negative, n
(%) | Positive, n
(%) | P-value |
|---|
| Tamoxifen | | | | | | | | | |
|
No
tamoxifen | 78(25) | 235(75) | | 74(21) | 276(79) | | 142(41) | 208(59) | |
|
Tamoxifen | 86(25) | 254(75) | 0.912 | 77(20) | 304(80) | 0.756 | 173(45) | 208(55) | 0.187 |
| Size, mm | | | | | | | | | |
|
<20 | 126(25) | 374(75) | | 120(22) | 432(78) | | 212(38) | 340(62) | |
|
>20 | 36(25) | 109(75) | 0.927 | 28(17) | 135(83) | 0.207 | 98(60) | 65(40) | <0.001 |
| ER | | | | | | | | | |
|
Negative | 43(34) | 85(66) | | 27(17) | 130(83) | | 101(64) | 56(36) | |
|
Positive | 118(23) | 394(77) | 0.014 | 122(22) | 431(78) | 0.187 | 207(37) | 346(63) | <0.001 |
| PgR | | | | | | | | | |
|
Negative | 76(28) | 192(72) | | 57(19) | 247(81) | | 158(52) | 146(48) | |
|
Positive | 77(24) | 244(76) | 0.228 | 68(20) | 269(80) | 0.649 | 123(37) | 214(63) | <0.001 |
| HER2 | | | | | | | | | |
|
Negative | 132(24) | 408(76) | | 134(22) | 465(78) | | 251(42) | 348(58) | |
|
Positive | 20(34) | 38(66) | 0.095 | 4(5) | 72(95) | <0.001 | 40(53) | 36(47) | 0.075 |
| Cyto p-Akt | | | | | | | | | |
|
Negative | 48(22) | 168(78) | | 98(35) | 183(65) | | 121(43) | 160(57) | |
|
Positive | 104(28) | 274(72) | 0.155 | 47(11) | 381(89) | <0.001 | 182(43) | 246(57) | 0.888 |
| Nucl p-Akt | | | | | | | | | |
|
Negative | 85(31) | 190(69) | | 75(24) | 240(76) | | 182(58) | 133(42) | |
|
Positive | 67(21) | 252(79) | 0.006 | 70(18) | 324(82) | 0.047 | 121(31) | 273(69) | <0.001 |
| Cyto p-S6K | | | | | | | | | |
|
Negative | 53(21) | 196(79) | | 82(28) | 211(72) | | 127(43) | 166(57) | |
|
Positive | 95(27) | 252(73) | 0.090 | 60(15) | 337(85) | <0.001 | 171(43) | 226(57) | 0.943 |
| Nucl p-S6K | | | | | | | | | |
|
Negative | 103(28) | 270(72) | | 105(24) | 334(76) | | 230(52) | 209(48) | |
|
Positive | 45(20) | 177(80) | 0.045 | 37(15) | 213(85) | 0.004 | 67(27) | 183(73) | <0.001 |
| PAM50 | | | | | | | | | |
|
Basal
like | 19(30) | 44(70) | | 7(13) | 48(87) | | 41(75) | 14(25) | |
|
Luminal
B | 39(30) | 90(70) | | 19(17) | 93(83) | | 53(47) | 59(53) | |
|
HER2
enriched | 20(34) | 38(66) | | 6(12) | 45(88) | | 35(69) | 16(31) | |
|
Luminal
A | 75(22) | 267(78) | | 52(19) | 229(81) | | 96(34) | 185(66) | |
|
Normal
like | 11(18) | 49(82) | 0.070 | 13(30) | 31(70) | 0.164 | 12(27) | 32(73) | <0.001 |
A Dutch set available online with gene-expression
data on breast tumors from systemically untreated patients together
with clinical data was used to test the prognostic value of the
IP6Ks in an independent cohort (22). In line with our gene expression
results from the Swedish low-risk cohort high IP6K2 expression
predicted better outcome than low IP6K2 expression (HR: 0.45, 95%
CI: 0.27-0.76, P=0.0026 and multivariable HR: 0.38, 95% CI:
0.21-0.68, P=0.0011, Fig. 1C).
Likewise, the distant recurrence-free interval among these patients
did not differ based on the IP6K1 levels (univariable P=0.60 and
multivariable P=0.73, Fig. 1D).
Similar results were obtained in the TCGA cohort,
although no treatment data was available and overall survival was
used as end point. High IP6K2 expression indicated better prognosis
in univariable (P=0.013) but not in multivariable analysis
correcting for node-, ER-, PgR-, HER2- and TP53 status
(P=0.21), whereas IP6K1 did not have prognostic value (Fig. 1E and F). Subgrouping on TP53 mutational
status indicated the importance of a functional TP53 for
IP6K2 to be a prognostic biomarker in the TCGA cohort (Table III). No TP53 mutation data was
available in the other cohorts.
| Table IIIIP6K2 and IP6K1 gene expression from
RNA sequencing and TP53 status in The Cancer Genome Atlas
cohort. |
Table III
IP6K2 and IP6K1 gene expression from
RNA sequencing and TP53 status in The Cancer Genome Atlas
cohort.
| | TP53 wt
(n=799) | TP53 mutated
(n=300) |
|---|
| Variable | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| IP6K2 (Q1 vs.
Q2-4) | | | | | | |
|
Univariable | 0.49 | 0.32-0.75 | <0.01 | 0.84 | 0.48-1.48 | 0.55 |
|
Multivariable | 0.51 | 0.28-0.94 | 0.03 | 1.40 | 0.63-3.11 | 0.41 |
| IP6K1 (Q1 vs.
Q2-4) | | | | | | |
|
Univariable | 0.80 | 0.52-1.24 | 0.32 | 0.92 | 0.52-1.62 | 0.76 |
|
Multivariable | 0.89 | 0.46-1.72 | 0.73 | 1.33 | 0.58-3.05 | 0.51 |
Roles of IP6K1 and IP6K2 diverge in
relation to cancer-related genes at 3p21.31
The chromosomal location of IP6K1 and
IP6K2 is at a gene-dense region including several genes with
expected impact on cancer development (Fig. 2 and Table IV). We tested gene expression of
the IP6Ks in relation to six closely located genes and clinical
biomarkers in a multivariable Cox regression analysis in three
cohorts (Table V). In the Swedish
low-risk- and the Dutch cohort IP6K2 expression remained a
protective factor, with a significantly reduced risk of distant
recurrence (Swedish low risk: HR: 0.54, 95% CI: 0.32-0.90, P=0.018,
Dutch: HR: 0.29, 95% CI: 0.15-0.56, P=0.0003), whereas high levels
of IP6K1 significantly increased the risk of distant recurrence
(Swedish low risk: HR: 2.07, 95% CI: 1.07-4.02, P=0.032, Dutch: HR:
2.20, 95% CI: 1.02-4.75, P=0.045).
| Table IVGenes associated with cancer at the
chromosomal region 3p.21.31. |
Table IV
Genes associated with cancer at the
chromosomal region 3p.21.31.
| Gene | Functions |
|---|
| SETD2 | Tumor suppressor
gene coding a histone methyltransferase involved in transcription.
Interacts with p53 and regulates its downstream genes. |
| PTPN23 | Tumor suppressor
gene coding a tyrosine phosphatase suppressing tumor cell motility
and invasion. |
| CDC25A | Oncogene coding a
phosphatase, activating CDK2 in G1 to S phase transition. |
| PLXNB1 | Tumor suppressor
gene coding a cell-surface receptor for semaphorin, controlling
cell adhesion. |
| IP6K2 | Potential tumor
suppressor gene coding a kinase involved in cell adhesion and
p53-regulated apoptosis and through generation of the Akt inhibitor
IP7. |
| RHOA | Oncogene coding a
Ras super family member regulating cell motility and invasion. |
| IP6K1 | Gene coding a
kinase involved in α-actinin and FAK regulated cell migration.
Potential role in cancer through generation of the Akt inhibitor
IP7. |
| RASSF1 | Tumor suppressor
gene coding the Ras-association domain family 1 protein,
downregulated by Akt, inhibiting accumulation of cyclin D1 and
induces cell cycle arrest. |
| Table VDistant recurrence-free interval
analyzed by multivariable Cox regression. |
Table V
Distant recurrence-free interval
analyzed by multivariable Cox regression.
| | Swedish low risk
(n=261) | Dutch (n=165) | TCGA (n=340) |
|---|
| Variable | HR | 95% CI | P-value | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| SETD2 | 1.01 | 0.54-1.89 | 0.97 | 0.54 | 0.29-0.99 | 0.05 | 0.98 | 0.47-2.02 | 0.95 |
| PTPN23 | 0.73 | 0.44-1.23 | 0.24 | 1.01 | 0.49-2.06 | 0.98 | 1.56 | 0.61-4.00 | 0.35 |
| CDC25A | 1.25 | 0.69-2.26 | 0.46 | 1.21 | 0.63-2.33 | 0.56 | 1.45 | 0.66-3.20 | 0.36 |
| PLXNB1 | 1.33 | 0.73-2.42 | 0.35 | 0.84 | 0.43-1.65 | 0.61 | 1.21 | 0.50-2.94 | 0.67 |
| IP6K2 | 0.54 | 0.32-0.90 | 0.02 | 0.29 | 0.15-0.56 | <0.01 | 0.66 | 0.29-1.50 | 0.32 |
| RHOA | 1.40 | 0.73-2.69 | 0.31 | 0.77 | 0.41-1.43 | 0.41 | 1.10 | 0.44-2.77 | 0.84 |
| IP6K1 | 2.07 | 1.07-4.02 | 0.03 | 2.20 | 1.02-4.75 | 0.05 | 1.69 | 0.61-4.65 | 0.31 |
| RASSF1 | 0.64 | 0.38-1.07 | 0.09 | 1.73 | 0.88-3.37 | 0.11 | 0.89 | 0.41-1.94 | 0.77 |
| ER | 0.60 | 0.29-1.24 | 0.17 | 1.24 | 0.61-2.49 | 0.55 | 0.92 | 0.26-3.19 | 0.89 |
| PgR | 2.00 | 1.11-3.59 | 0.02 | - | - | - | 1.09 | 0.37-3.25 | 0.87 |
| HER2 | 2.41 | 1.21-4.81 | 0.01 | 3.43 | 1.78-6.62 | <0.01 | 0.78 | 0.30-1.97 | 0.59 |
| Node status | - | - | - | 1.56 | 0.81-3.00 | 0.19 | 1.97 | 1.00-3.87 | 0.05 |
| Tumor size | 2.48 | 1.50-4.09 | <0.01 | - | - | - | - | - | - |
IP6K1 and IP6K2 protein
expression
Cytoplasmic and nuclear protein detection was
successful in 731 and 724 tumors for IP6K1 and IP6K2, respectively.
No significant prognostic value of IP6Ks in the cytoplasm was
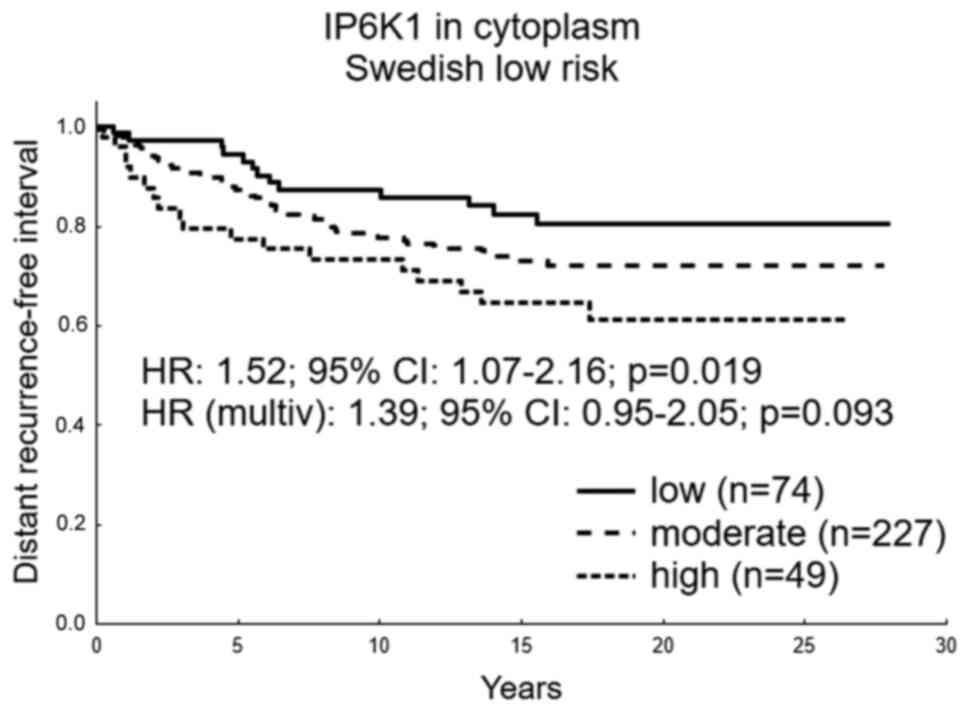
detected in systemically untreated patients (Fig. S3A and B). However, cytoplasmic IP6K1 tended to
indicate a worse prognosis with higher levels of expression
(P=0.080) in the univariate analysis. Therefore, we divided the
IP6K1 expression in three groups; low, medium and high, where a
worse prognosis was seen with increasing expression in the
univariable analysis (HR: 1.52, 95% CI: 1.07-2.16, P=0.019), and a
trend in the multivariable analysis correcting for ER, PgR and HER2
status and tumor size (HR: 1.39, 95% CI: 0.95-2.05, P=0.093,
Fig. 3). No significant prognostic
value was detected with nuclear expression; however, a minor trend
of worse prognosis was seen in patients with tumors lacking IP6K in
the nucleus (Fig. S3C and D).
Testing protein expression of all tumors in relation
to clinicopathological and PI3K/Akt/mTOR-pathway variables IP6K2
cytoplasmic expression was significantly higher in tumors with high
levels of cytoplasmic p-Akt (P=0.0044) and cytoplasmic p-S6K
(P=0.00027, Table I). IP6K2 nuclear
positivity associated with small tumors (P=0.00084) and p-Akt in
the nucleus (P=0.0022). IP6K1 cytoplasmic expression associated
with HER2 overexpression (P=0.00049), cytoplasmic p-Akt
(P<0.00001) and cytoplasmic p-S6K (P=0.00004, Table II). IP6K1 nuclear positivity
associated strongly with small tumor size, ER, PgR and the
PI3K/Akt/mTOR-pathway markers p-Akt and p-S6K in the nucleus, as
well as with the PAM50 luminal molecular subtype. IP6K1 and IP6K2
protein expression were highly associated in cytoplasm (P=0.0011)
and in nuclei (P<0.00001).
Discussion
In this study we present data on IP6K1 and IP6K2 in
tumors from three large sets of patients with follow up after
primary breast cancer. We investigated the prognostic potential of
these genes because of their coupling to the PI3K/Akt/mTOR pathway
that has been previously studied in our group. Recently reviewed
results are varying and exclusively generated from in vitro
and animal studies (28).
Previous studies have reported both common and
separate roles for the two kinases. Both are important for the
generation of IP7 and cell migration. IP6K2 is most prominent in
apoptosis in a variety of cells and more often mentioned in
association with cancer, while IP6K1 is reported to be a kinase
important for insulin secretion in pancreas (11,29,30).
The IP6Ks are known to interfere with the PI3K/Akt/mTOR pathway
through generation of the Akt inhibitor IP7(3). IP6K1 and IP6K2 gene expression as well
as protein nuclear expression associated here with PI3K/Akt/mTOR
activity in the nucleus, not in the cytoplasm. However, high
cytoplasmic protein expression of the IP6Ks associated with an
active PI3K/Akt/mTOR pathway in the cytoplasm. This was not in line
with the expected effect of IP6Ks on the PI3K/Akt/mTOR activity,
thus reflecting that other factors may be involved.
For IP6K2, the hypothesized protective role in
breast cancer was supported in gene-expression analyses in three
independent breast cancer cohorts of which two had systemically
untreated patients. Studies point out IP6K2 as a potential tumor
suppressor. Morrison et al (12,13)
showed that cells with overexpressed IP6K2 had a better response to
radiation treatment than controls and IP6K2 K-O mice exposed to an
oral carcinogen developed oral cancer four times more often than
controls. Interestingly, the protective role of IP6K2 remained true
only when the TP53 was intact in the TCGA cohort, suggesting
that the function of IP6K2 as tumor suppressor is over-ruled by the
oncogenic features generated by TP53 mutation. The p53
expression is regulated by IP7 through catalytically active IP6K
and in addition nuclear IP6K2 binds p53 and controls the
transcription towards cell repair inhibition and apoptosis
activation (11,31). We suggest that in the TP53
mutated cells, this regulation has no effect, as we do not see a
prognostic effect of IP6K2 in patients with TP53 mutated
tumors.
The IP6K1 and IP6K2 genes are located
closely together on 3p21.31, with several other genes commonly
deregulated in breast cancer (7).
Results seen with gene expression could reflect characters of other
genes located close to the IP6Ks. Therefore, we searched the
region and found six cancer-related genes of interest, namely;
SETD2 (32-34),
PTPN23 (35), CDC25A
(36), PLXNB1 (37), RHOA (38,39),
and RASSF1 (40,41). Interestingly, IP6K1 and IP6K2 gene
expression show consistent results in multivariable tests of
prognostic value in the two cohorts with systemically untreated
patients and tendencies in the same direction in the TCGA cohort.
Correcting for the expression of the six genes and clinical
biomarkers, the IP6K2 decreased the risk of recurrence, as expected
of a tumor suppressor gene, and IP6K1 increased the risk of
recurrence, as expected of an oncogene. Apart from the previously
known prognostic factors; HER2, tumor size and node status, that
came out significant in the analysis, only SETD2 indicated a
significant prognostic value as tumor suppressor in one cohort.
These results suggest that gene-expression analysis can be a useful
method to detect prognostic value of individual genes in this
gene-dense region.
Analyzing the molecular profile of the IP6K1/2 loci
in the large cohort TCGA the mutational burden in these genes was
very low. Frequent copy-number variation with more loss than
amplification was seen. Copy number and gene expression shows high
degree of association for both genes. However, gene expression can
be additionally regulated (23). In
squamous cell carcinoma of head and neck no promoter methylation of
the CDC25A gene was found, while this was frequent in other genes
in the region, such as RASSF1A (6).
IP6K2 protein localization has been explored in
functional studies showing that the heat-shock protein HSP90, which
often is active in tumor cells, binds directly to IP6K2, not IP6K1,
and inhibits its nuclear translocation (42). The nuclear IP6K2 binds p53 and
direct the response of DNA damage towards apoptosis rather than
cell cycle arrest and repair (10,11).
Apoptotic functions of IP6K2 have also been described in neuronal
cells (43-45).
In our study we detect no prognostic value of IP6K2 protein levels,
although nuclear IP6K2 was more common in small tumors and a minor
trend of worse prognosis when tumors lack nuclear IP6K2 was
noticed. Localized to the cytoplasmic compartment of the tumor
cells IP6K1 tends to take an oncogenic role, and significantly when
further dividing the expression in three groups, indicating worse
prognosis. This is in line with previous findings describing that
IP6K1 is a driver of cell migration (30,46,47).
In addition, we found high IP6K1 protein expression in the
cytoplasm to be more common in HER2-positive disease and associated
with high activity in the PI3K/Akt/mTOR pathway. Nuclear IP6K1 had
no significant prognostic value, however in contrast to when found
in the cytoplasm the nuclear IP6K1 was more common in small tumors
and ER -and PgR positive tumors.
Data presented here represents new insight in the
role of the IP6K1 and IP6K2 as prognostic biomarkers in breast
cancer. Gene expression of IP6K2 was the most prominent prognostic
marker, demonstrating its protective role in independent cohorts.
IP6K1 stood out as an independent prognostic marker of worse
outcome in relation to other cancer-related genes on 3p21.31.
Targeting the IP6Ks has potential as breast cancer treatment,
although precise selection of patient groups and isoform specific
molecules are of importance as the IP6K1 and IP6K2 seem to diverge
in breast cancer.
Supplementary Material
Immunohistochemical staining of IP6K
proteins. (A) IP6K2 cytoplasm negative and nucleus negative. (B)
IP6K2 cytoplasm weak and nucleus positive. (C) IP6K2 cytoplasm
moderate and nucleus negative. (D) IP6K2 cytoplasm strong and
nucleus positive. (E) IP6K1 cytoplasm negative and nucleus
negative. (F) IP6K1 cytoplasm weak and nucleus positive. (G) IP6K1
cytoplasm moderate and nucleus negative. (H) IP6K1 cytoplasm strong
and nucleus positive. Arrows indicate cytoplasmic and/or nuclear
staining (magnification, x400). IP6K, inositol hexakisphosphate
kinase.
IP6K antibody validation. (A) ZR751
breast cancer cells were used to validate the IP6K2 antibody. Two
siRNAs (IP6K2si1 and IP6K2si2) were tested, and IP6K2si2 exhibited
knockdown of IP6K2, 50 kDa. (B) BT474 breast cancer cells were used
to validate the IP6K1 antibody later used for immunostaining of
formalin-fixed paraffin-embedded breast cancer tumor tissue. One
siRNA was tested and exhibited complete knockdown. (C) T47D breast
cancer cells were used to further validate the IP6K1 antibody later
used for immunostaining. One siRNA was tested and exhibited partial
knockdown of the IP6K1, 50 kDa. An untransfected control and cell
lysate from cells transfected with scrambled siRNA were included
for all cell lines. GAPDH protein detection, 36 kDa, served as
loading control. IP6K, inositol hexakisphosphate kinase; siRNA/si,
small interfering RNA.
Distant recurrence-free interval in
relation to protein expression and localization of the IP6Ks. (A)
IP6K2 in the cytoplasm and (B) IP6K1 cytoplasmic protein expression
in tumor cells. Detection of nuclear expression of (C) IP6K2 and
(D) IP6K1. Prognostic data were produced from systemically
untreated patients with primary tumors of the Swedish low-risk
breast cancer cohort. The multivariable Cox proportional hazard
model corrected for tumor size, and estrogen receptor, progesterone
receptor and HER2 status. IP6K, inositol hexakisphosphate kinase;
HR, hazard ratio; multiv, multivariate.
Acknowledgements
The authors would like to thank Mrs. Birgitta
Holmlund and Mrs. Birgit Olsson, both technicians (Linköping
University, Linköping, Sweden), and Dr Lambert Skoog (Karolinska
Institute, Stockholm, Sweden), for technical assistance and
assessment of clinical biomarkers, respectively.
Funding
The present study was funded by the Swedish Cancer Society
(grant no. 17-0479; to OS), the Region of Östergötland (grant no.
LIO-795201; to OS), the Percy Falk Foundation (grant no.
2017-02-10; to JS) and the Oncology Clinic in Linköping Research
Fund (grant no. 2019-10-29; to JS).
Availability of data and materials
The data from the Swedish cohort are not publicly
available as the study participants did not consent to sharing
their data in a public repository but are available from the
corresponding author on reasonable request. The Dutch public
dataset analyzed in the current study is available at http://bioinformatics.nki.nl/data.php.
The TCGA clinical and genomic data are publicly available from the
open-access resource for cancer genomics, cBioPortal (https://www.cbioportal.org/).
Authors' contributions
OS and JS secured funding. JS, AB, RL and OS
conceived the study, and participated in the study design and
coordination. JS, AB and RL conducted the experiments. JS, LL, GPT
and OS conducted the data analyses and confirm the authenticity of
all raw data. TF and BN initiated the clinical trial, and collected
patient material and follow-up data for the Swedish cohort. All
authors read the manuscript drafts and contributed edits. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures performed in studies involving human
participants were in accordance with the ethical standards of the
institutional and national research committee and with the 1964
Helsinki Declaration and its later amendments or comparable ethical
standards. Ethical approval for the Swedish cohort was obtained
from the Karolinska Institute Ethics Council, with approved
amendments 02-01-2003 and 10-17-2017. According to the ethical
approval, informed consent from the Swedish patients was not
required.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Guerrero-Zotano A, Mayer IA and Arteaga
CL: PI3K/AKT/mTOR: Role in breast cancer progression, drug
resistance, and treatment. Cancer Metastasis Rev. 35:515–524.
2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Paplomata E and O'Regan R: The
PI3K/AKT/mTOR pathway in breast cancer: Targets, trials and
biomarkers. Ther Adv Med Oncol. 6:154–166. 2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Chakraborty A, Koldobskiy MA, Bello NT,
Maxwell M, Potter JJ, Juluri KR, Maag D, Kim S, Huang AS, Dailey
MJ, et al: Inositol pyrophosphates inhibit akt signaling, thereby
regulating insulin sensitivity and weight gain. Cell. 143:897–910.
2010.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Luo HR, Huang YE, Chen JC, Saiardi A,
Iijima M, Ye K, Huang Y, Nagata E, Devreotes P and Snyder SH:
Inositol pyrophosphates mediate chemotaxis in dictyostelium via
pleckstrin homology domain-PtdIns(3,4,5)P3 interactions. Cell.
114:559–572. 2003.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Saiardi A, Erdjument-Bromage H, Snowman
AM, Tempst P and Snyder SH: Synthesis of diphosphoinositol
pentakisphosphate by a newly identified family of higher inositol
polyphosphate kinases. Curr Biol. 9:1323–1326. 1999.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Ghosh S, Ghosh A, Maiti GP, Alam N, Roy A,
Roy B, Roychoudhury S and Panda CK: Alterations of 3p21.31 tumor
suppressor genes in head and neck squamous cell carcinoma:
Correlation with progression and prognosis. Int J Cancer.
123:2594–2604. 2008.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ching HC, Naidu R, Seong MK, Har YC and
Taib NA: Integrated analysis of copy number and loss of
heterozygosity in primary breast carcinomas using high-density SNP
array. Int J Oncol. 39:621–633. 2011.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Manning BD and Cantley LC: AKT/PKB
signaling: Navigating downstream. Cell. 129:1261–1274.
2007.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Jadav RS, Kumar D, Buwa N, Ganguli S,
Thampatty SR, Balasubramanian N and Bhandari R: Deletion of
inositol hexakisphosphate kinase 1 (IP6K1) reduces cell migration
and invasion, conferring protection from aerodigestive tract
carcinoma in mice. Cell Signal. 28:1124–1136. 2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Rao F, Cha J, Xu J, Xu R, Vandiver MS,
Tyagi R, Tokhunts R, Koldobskiy MA, Fu C, Barrow R, et al: Inositol
pyrophosphates mediate the DNA-PK/ATM-p53 cell death pathway by
regulating CK2 phosphorylation of Tti1/Tel2. Mol Cell. 54:119–132.
2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Koldobskiy MA, Chakraborty A, Werner JK
Jr, Snowman AM, Juluri KR, Vandiver MS, Kim S, Heletz S and Snyder
SH: p53-Mediated apoptosis requires inositol hexakisphosphate
kinase-2. Proc Natl Acad Sci USA. 107:20947–20951. 2010.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Morrison BH, Bauer JA, Hu J, Grane RW,
Ozdemir AM, Chawla-Sarkar M, Gong B, Almasan A, Kalvakolanu DV and
Lindner DJ: Inositol hexakisphosphate kinase 2 sensitizes ovarian
carcinoma cells to multiple cancer therapeutics. Oncogene.
21:1882–1889. 2002.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Morrison BH, Haney R, Lamarre E, Drazba J,
Prestwich GD and Lindner DJ: Gene deletion of inositol
hexakisphosphate kinase 2 predisposes to aerodigestive tract
carcinoma. Oncogene. 28:2383–2392. 2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Bostner J, Karlsson E, Pandiyan MJ,
Westman H, Skoog L, Fornander T, Nordenskjöld B and Stål O:
Activation of akt, mTOR, and the estrogen receptor as a signature
to predict tamoxifen treatment benefit. Breast Cancer Res Treat.
137:397–406. 2013.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Karlsson E, Perez-Tenorio G, Amin R,
Bostner J, Skoog L, Fornander T, Sgroi DC, Nordenskjöld B, Hallbeck
AL and Stål O: The mTOR effectors 4EBP1 and S6K2 are frequently
coexpressed, and associated with a poor prognosis and endocrine
resistance in breast cancer: A retrospective study including
patients from the randomised stockholm tamoxifen trials. Breast
Cancer Res. 15(R96)2013.PubMed/NCBI View
Article : Google Scholar
|
|
16
|
Bostner J, Karlsson E, Eding CB,
Perez-Tenorio G, Franzén H, Konstantinell A, Fornander T,
Nordenskjöld B and Stål O: S6 kinase signaling: Tamoxifen response
and prognostic indication in two breast cancer cohorts. Endocr
Relat Cancer. 22:331–343. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Rutqvist LE and Johansson H: Stockhol
Breast Cancer Study Group. Long-Term follow-up of the randomized
stockholm trial on adjuvant tamoxifen among postmenopausal patients
with early stage breast cancer. Acta Oncol. 46:133–145.
2007.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Jerevall PL, Jansson A, Fornander T, Skoog
L, Nordenskjöld B and Stål O: Predictive relevance of HOXB13
protein expression for tamoxifen benefit in breast cancer. Breast
Cancer Res. 12(R53)2010.PubMed/NCBI View
Article : Google Scholar
|
|
19
|
Khoshnoud MR, Löfdahl B, Fohlin H,
Fornander T, Stål O, Skoog L, Bergh J and Nordenskjöld B:
Immunohistochemistry compared to cytosol assays for determination
of estrogen receptor and prediction of the long-term effect of
adjuvant tamoxifen. Breast Cancer Res Treat. 126:421–430.
2011.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Bostner J, Skoog L, Fornander T,
Nordenskjöld B and Stål O: Estrogen receptor-alpha phosphorylation
at serine 305, nuclear p21-activated kinase 1 expression, and
response to tamoxifen in postmenopausal breast cancer. Clin Cancer
Res. 16:1624–1633. 2010.PubMed/NCBI View Article : Google Scholar
|
|
21
|
McShane LM, Altman DG, Sauerbrei W, Taube
SE, Gion M and Clark GM: Statistics Subcommittee of the NCI-EORTC
Working Group on Cancer Diagnostics. Reporting recommendations for
tumour MARKer prognostic studies (REMARK). Br J Cancer. 93:387–391.
2005.PubMed/NCBI View Article : Google Scholar
|
|
22
|
van de Vijver MJ, He YD, van't Veer LJ,
Dai H, Hart AA, Voskuil DW, Schreiber GJ, Peterse JL, Roberts C,
Marton MJ, et al: A gene-expression signature as a predictor of
survival in breast cancer. N Engl J Med. 347:1999–2009.
2002.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Cancer Genome Atlas Research Network.
Weinstein JN, Collisson EA, Mills GB, Shaw KR, Ozenberger BA,
Ellrott K, Shmulevich I, Sander C and Stuart JM: The cancer genome
atlas pan-cancer analysis project. Nat Genet. 45:1113–1120.
2013.PubMed/NCBI View
Article : Google Scholar
|
|
24
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBioPortal. Sci Signal. 6(pl1)2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Spratt DE, Chan T, Waldron L, Speers C,
Feng FY, Ogunwobi OO and Osborne JR: Racial/Ethnic disparities in
genomic sequencing. JAMA Oncol. 2:1070–1074. 2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Esserman LJ, Yau C, Thompson CK, van't
Veer LJ, Borowsky AD, Hoadley KA, Tobin NP, Nordenskjöld B and
Fornander T: Use of molecular tools to identify patients with
indolent breast cancers with ultralow risk over 2 decades. JAMA
Oncol. 3:1503–1510. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Parker JS, Mullins M, Cheang MC, Leung S,
Voduc D, Vickery T, Davies S, Fauron C, He X, Hu Z, et al:
Supervised risk predictor of breast cancer based on intrinsic
subtypes. J Clin Oncol. 27:1160–1167. 2009.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chakraborty A: The inositol pyrophosphate
pathway in health and diseases. Biol Rev Camb Philos Soc.
93:1203–1227. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Illies C, Gromada J, Fiume R, Leibiger B,
Yu J, Juhl K, Yang SN, Barma DK, Falck JR, Saiardi A, et al:
Requirement of inositol pyrophosphates for full exocytotic capacity
in pancreatic beta cells. Science. 318:1299–1302. 2007.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Fu C, Xu J, Cheng W, Rojas T, Chin AC,
Snowman AM, Harraz MM and Snyder SH: Neuronal migration is mediated
by inositol hexakisphosphate kinase 1 via α-actinin and focal
adhesion kinase. Proc Natl Acad Sci USA. 114:2036–2041.
2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Gu C, Nguyen HN, Ganini D, Chen Z, Jessen
HJ, Gu Z, Wang H and Shears SB: KO of 5-insP7 kinase activity
transforms the HCT116 colon cancer cell line into a hypermetabolic,
growth-inhibited phenotype. Proc Natl Acad Sci USA.
114:11968–11973. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Al Sarakbi W, Sasi W, Jiang WG, Roberts T,
Newbold RF and Mokbel K: The mRNA expression of SETD2 in human
breast cancer: Correlation with clinico-pathological parameters.
BMC Cancer. 9(290)2009.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Roberti A, Dobay MP, Bisig B, Vallois D,
Boéchat C, Lanitis E, Bouchindhomme B, Parrens MC, Bossard C and
Quintanilla-Martinez L: Type II enteropathy-associated T-cell
lymphoma features a unique genomic profile with highly recurrent
SETD2 alterations. Nat Commun. 7(12602)2016.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Duns G, van den Berg E, van Duivenbode I,
Osinga J, Hollema H, Hofstra RM and Kok K: Histone
methyltransferase gene SETD2 is a novel tumor suppressor gene in
clear cell renal cell carcinoma. Cancer Res. 70:4287–4291.
2010.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhang S, Fan G, Hao Y, Hammell M,
Wilkinson JE and Tonks NK: Suppression of protein tyrosine
phosphatase N23 predisposes to breast tumorigenesis via activation
of FYN kinase. Genes Dev. 31:1939–1957. 2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Sadeghi H, Golalipour M, Yamchi A,
Farazmandfar T and Shahbazi M: CDC25A pathway toward tumorigenesis:
Molecular targets of CDC25A in cell-cycle regulation. J Cell
Biochem. 120:2919–2928. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Malik MF, Ye L and Jiang WG: Reduced
expression of semaphorin 4D and plexin-B in breast cancer is
associated with poorer prognosis and the potential linkage with
oestrogen receptor. Oncol Rep. 34:1049–1057. 2015.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Zhang C, Wang HJ, Bao QC, Wang L, Guo TK,
Chen WL, Xu LL, Zhou HS, Bian JL, Yang YR, et al: NRF2 promotes
breast cancer cell proliferation and metastasis by increasing
RhoA/ROCK pathway signal transduction. Oncotarget. 7:73593–73606.
2016.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Jeong D, Park S, Kim H, Kim CJ, Ahn TS,
Bae SB, Kim HJ, Kim TH, Im J, Lee MS, et al: RhoA is associated
with invasion and poor prognosis in colorectal cancer. Int J Oncol.
48:714–722. 2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Agathanggelou A, Cooper WN and Latif F:
Role of the ras-association domain family 1 tumor suppressor gene
in human cancers. Cancer Res. 65:3497–3508. 2005.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Blanchard TG, Lapidus R, Banerjee V,
Bafford AC, Czinn SJ, Ahmed H and Banerjee A: Upregulation of
RASSF1A in colon cancer by suppression of angiogenesis signaling
and akt activation. Cell Physiol Biochem. 48:1259–1273.
2018.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Chakraborty A, Koldobskiy MA, Sixt KM,
Juluri KR, Mustafa AK, Snowman AM, van Rossum DB, Patterson RL and
Snyder SH: HSP90 regulates cell survival via inositol
hexakisphosphate kinase-2. Proc Natl Acad Sci USA. 105:1134–1139.
2008.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Nagata E, Saiardi A, Tsukamoto H, Okada Y,
Itoh Y, Satoh T, Itoh J, Margolis RL, Takizawa S, Sawa A and Takagi
S: Inositol hexakisphosphate kinases induce cell death in
huntington disease. J Biol Chem. 286:26680–26686. 2011.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Nagata E, Nonaka T, Moriya Y, Fujii N,
Okada Y, Tsukamoto H, Itoh J, Okada C, Satoh T, Arai T, et al:
Inositol hexakisphosphate kinase 2 promotes cell death in cells
with cytoplasmic TDP-43 aggregation. Mol Neurobiol. 53:5377–5383.
2016.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Moriya Y, Nagata E, Fujii N, Satoh T,
Ogawa H, Hadano S and Takizawa S: Inositol hexakisphosphate kinase
2 is a presymptomatic biomarker for amyotrophic lateral sclerosis.
Tokai J Exp Clin Med. 42:13–18. 2017.PubMed/NCBI
|
|
46
|
Rao F, Xu J, Fu C, Cha JY, Gadalla MM, Xu
R, Barrow JC and Snyder SH: Inositol pyrophosphates promote tumor
growth and metastasis by antagonizing liver kinase B1. Proc Natl
Acad Sci USA. 112:1773–1778. 2015.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Izaguirre G, Aguirre L, Hu YP, Lee HY,
Schlaepfer DD, Aneskievich BJ and Haimovich B: The
cytoskeletal/non-muscle isoform of alpha-actinin is phosphorylated
on its actin-binding domain by the focal adhesion kinase. J Biol
Chem. 276:28676–28685. 2001.PubMed/NCBI View Article : Google Scholar
|