1. Introduction
Cells that manage to elude central endogenous
regulatory mechanisms proliferate uncontrollably, which is how
cancer is defined (1). Cancers may
be categorized according to the organ or tissue from whence they
originated, but the molecular properties of cancer cells are
increasingly being studied and understood (2). Cancer is the second greatest cause of
mortality and a significant public health issue globally, despite
the notable advancements in tumor detection and therapy over the
past few years, as noted by the World Health Organization and
American Cancer Society (3,4). The
GLOBOCAN 2020 update of the global cancer burden considers cancer
incidence and mortality. It predicts that there will be 19.3
million new cases of cancer worldwide (excluding non-melanoma skin
cancer) and >10 million cancer deaths (excluding non-melanoma
skin cancer) in 2023(5). There
were 983,160 new cases of cancer in male patients and 322,090
fatalities in the United States in 2022(6).
In males as a representative population, lung cancer
is the most common type of cancer and the leading cause of
cancer-associated death, followed by prostate cancer and colorectal
cancers (5,6). Carcinogenesis is the process through
which healthy normal cells develop into cancer cells, where
oncogenes and tumor suppressor genes have a role in the development
of cancer (7). In addition to
acting as nuclear transcription factors and signal transducers,
oncogenes may also act as growth factors (8,9).
Various oncogenes in mammals regulate proper cell differentiation
and proliferation (8,9).
Along with oncogenes, tumor-suppressor genes also
have a significant role in the regulation of the growth and
differentiation of normal cells and prevent the development of
cancer (8,9). The vast majority of tumor-suppressor
genes function together to prevent the development of neoplasia in
an organism. A tumor-suppressor gene must be dormant in both copies
inherited from each parental cell for a cancer cell to continue to
grow or survive (7).
Tumor-suppressor genes are present dispersed across the human
genome and are involved in the development of the diverse forms of
human neoplasia (7). A single cell
that has developed malignant characteristics due to cellular DNA
damage is the first step in the genesis of a malignant tumor in an
otherwise healthy tissue (10).
However, only a relatively small number of valuable cancer
biomarkers are being identified and confirmed for diagnostic and
therapeutically helpful screening. Given the increasing incidence
and mortality of cancer worldwide, there is a growing demand for
precise biomarkers for improved detection, diagnosis, prognosis and
monitoring (11,12).
Protein post-translational modifications (PTMs) have
a significant impact on how proteins function and are required for
virtually all biological processes in cells (13). However, PTMs and genetic
alterations are substantial in the onset and development of
carcinogenesis and cancer, as evidenced by the detrimental effects
of several PTMs (13). Protein
ubiquitination is a post-translational alteration that controls
several physiological processes (14). It involves binding of ubiquitin, a
highly conserved 76-amino-acid polypeptide, to lysine residues of
target substrates (15,16). The functions of protein
ubiquitination are listed below, with the primary role of protein
ubiquitination being the eventual breakdown of proteins (17). The two principal mechanisms that
control the ubiquitination of proteins and, consequently, the size
of protein pools, are proteasomal and lysosomal degradation
(18). According to estimates,
lysosomal-mediated degradation accounts for 20% of the total
degradation of ubiquitinated proteins, with the proteasome
contributing 80% (19,20).
Three necessary enzymes known as E1, E2 and E3 are
responsible for tagging target proteins with ubiquitin chains
(21,22). The E1 enzyme is a
ubiquitin-activating enzyme that uses ATP to adenylate a lysine at
the C-terminus of a ubiquitin molecule (23). The ubiquitin-conjugating enzyme (E2
enzyme) then transfers active ubiquitin to a cysteine residue,
forming E2-S-ubiquitin intermediates (23). An E3 ubiquitin ligase enzyme then
transfers the active ubiquitin to interact with an amino
(NH2) group in the lysine residue of the target protein
through the C-terminal domain of the ubiquitin signal that is
attached to the E2 enzyme (24).
Only two E1 enzymes, ~35 E2 enzymes and >600 E3 ubiquitin
ligases are encoded in the human genome and they can target
different compounds in different pathways (25). Of note, the protein ubiquitination
cascade's rate-limiting stage is assumed to be E3-ligase-mediated
ubiquitination.
As previously established, E3-ubiquitin ligases have
a crucial role in the rate-limiting stage of protein ubiquitination
before directing it to the proteasome or other minor routes for
degradation. According to the method by which activated ubiquitin
is transferred, the E3-ubiquitin families can be divided into
subfamilies. The two most essential subfamilies are known as really
interesting new gene (RING)-type E3 ubiquitin ligases and
homologous to E6Ap C-terminus (HECT)-domain E3 ubiquitin ligases
(23,26,27).
Direct ubiquitin transfer is made possible by RING E3s building
bridges across the ring finger motif between E2 enzymes and the
target protein (23,26-28).
HECT E3 ligases, on the other hand, have a HECT
domain that joins with the E2 and forms a thioester bond with the
target protein, transferring ubiquitin from the cystine residue to
the protein (29). Nearly 95% of
all E3-ubiquitin ligases belong to the largest subfamily, the RING
E3s. With only 30 members described thus far, the HECT-domain
subfamily is substantially smaller. There are further subfamilies,
such as the recently identified RING/HECT hybrid subfamily and UFD2
(or U-box) homologous proteins (30). In addition, there are three other
subcategories under the HECT domain subfamily, including neural
precursor cell expressed developmentally downregulated protein 4
(NEDD4), HECT and RLD domain containing E3 ubiquitin protein ligase
and other residual HECTs (23,31).
From yeast to humans, NEDD4 is a highly conserved HECT-domain E3
ligase subfamily (32-35).
A lipid-binding N-terminal C2 domain, an auto-ubiquitination HECT
ligase domain and 2-4 tryptophan-tryptophan (WW) fields are
necessary for interaction with adaptors or substrates, making up
its conserved structure (36-38).
A total of 9 NEDD4 E3 ligases are present in the human genome and
have been demonstrated to be crucial for viral budding, protein
trafficking and target protein degradation (37).
NEDD4 (also known as NEDD4-1), NEDD4 like E3
ubiquitin protein ligase (NEDD4L; also known as NEDD4-2), WW
domain-containing E3 ubiquitin-protein ligase 1 (WWP1), WWP2, itchy
E3 ubiquitin protein ligase, SMAD-specific E3 ubiquitin-protein
ligases (Smurf1), Smurf2, HECT, C2 and WW domain containing E3
ubiquitin protein ligase 1 [NEDL1 (HECW1)] and NEDL2 (HECW2) are
the 9 ligases that constitute the NEDD4 family (39). Although the structures of these
ligases are similar, they have different roles due to their unique
WW domains, which are crucial for binding to target proteins
through their proline-rich sequence domains or
phospho-serine/threonine domains (38). Previous studies have indicated a
strong association between NEDD4L expression and tumor progression
in various malignancies, including prostate cancer, gastric cancer,
hepatoma and gallbladder carcinoma (40-42).
In addition, it controls several cellular functions by degrading
certain proteins. It has been acknowledged that ubiquitination is a
crucial stage in the development and progression of various
diseases in humans (39,43). To find new therapeutic targets for
a variety of illnesses, including atherosclerosis, cancer and
neurodegenerative disorders (44-46),
the ubiquitin proteasomal system has become increasingly studied
(47). This has led to the
development of treatments for various pathological conditions with
proteasomal inhibitors. For instance, the proteasomal inhibitor
bortezomib (Velcade®) is used to treat multiple myeloma
(48).
2. NEDD4 and NEDD4L in cancer
Lung cancer
Lung cancer is one of the most globally prevalent
cancers and the leading cause of cancer-related death in both men
and women worldwide, given its often rapidly occurring metastasis
and its aggressiveness (49,50).
In recent years, there has been a sharp rise in the incidence of
lung cancer and patients only have a 19% 5-year survival rate,
despite the extensive research and improved understanding of the
disease (51). Lung cancer is
categorized into small cell lung cancer (SCLC) and non-SCLC
(NSCLC). NSCLC includes three major histological subtypes:
Adenocarcinoma (ADC), extensive cell lung cancer and squamous cell
carcinoma (51-54).
Around 85% of all lung cancer cases are of the NSCLC subtype and
60% of these are of the ADC subtype (55). NEDD4 and NEDD4L play a part in the
proliferation, apoptosis, migration and medication resistance of
lung cancer cells (56). In-depth
research is therefore urgently required to increase the survival
rate of lung cancer patients.
The role of NEDD4 in lung cancer has received
increasing attention recently. It has been proposed that NEDD4
contributes to the development and spread of lung cancer tumors and
is overexpressed in a sizeable fraction of NSCLCs (57). A previous study has indicated that
lung cancer tissues have higher levels of NEDD4 and its silencing
was linked to a reduction in lung cancer cell proliferation in
vitro and tumor growth in vivo (56). NEDD4 augments the malignant
features of lung epithelial cells and promotes cell proliferation,
migration and invasion through phosphatase and tensin homolog
(PTEN) degradation (56). The PTEN
gene is a tumor suppressor that produces a lipid phosphatase and
counteracts the effects of PI3K/Akt. Lung cancer cells frequently
exhibit PTEN and PI3K/Akt signaling pathway loss and this is
associated with malignant transformation of lung cells and
resistance to chemotherapeutic drugs. In 55-74% of NSCLC cases,
PTEN expression is decreased or has been lost (57). NEDD4 negatively impacts PTEN levels
by polyubiquitination and proteolysis, although its impact on lung
cancer has remained elusive (52,57).
It has been found that the HER receptor tyrosine
kinase family member, epidermal growth factor receptor (EGFR) and
NEDD4 are able to interact. This signaling system is essential for
processes including lung cancer cell growth, migration, invasion
and apoptosis, as well as relapse (58). In addition, NEDD4 is activated by
EGFR signaling in NSCLC tumor tissues. NEDD4 knockdown
significantly inhibits the migration of NSCLC cells in response to
EGF and NEDD4 promotes EGF-induced cathepsin B lysosomal secretion,
indicating that NEDD4 mediates a novel EGFR migration signaling
pathway in lung cancer. Another study found that NEDD4 is more
readily degraded when prostate transmembrane protein, androgen
induced 1 (TMEAI) interacts with NEDD4 and binds to the
transforming growth factor-β (TGF-β)-type I receptor.
NEDD4 is required to deliver TMEPAI to the lysosome
(59). However, it is unclear
exactly how NEDD4 affects EGFR-dependent lung cancer cell migration
(58). In addition to controlling
the expression of NEDD4, EGFR signaling also regulates the
expression of NEDD4L (60). As a
crucial player in the development of lung cancer, NEDD4L may serve
as a prognostic indicator due to its association with aggressive
clinicopathological tumors and poor overall survival (60). It is recommended to use NEDD4L to
build a bridge between the EGFR and the mammalian target of
rapamycin (mTOR) pathways, where mTOR is another important oncogene
in solid tumors, the signaling of which regulates protein
synthesis, cell proliferation and survival (61). Thus, the EGFR-NEDD4L-mTOR pathway
is crucial for determining the outcome of NSCLC.
NEDD4 is implicated in the control of cancer cell
drug resistance, which may be a significant impediment to effective
treatment. According to a study on HCC827/ER lung cells, NEDD4 may
increase acquired resistance to erlotinib in NSCLC cells by
suppressing the expression of PTEN (62). Furthermore, in nude mice with
xenografted HCC827/ER cells, suppression of NEDD4 reduced tumor
development and tumor weight (62). Another in vitro study
revealed that NEDD4L expression was downregulated in NSCLC, where
it functions as a tumor suppressor, and NEDD4L knockdown increased
cell proliferation, invasion and migration (63,64).
In another study, NEDD4L expression was inhibited to demonstrate
the oncogenic role of microRNA (miR)-93 in lung cancer (65). miR-93 is a member of the
miR-106b-25 cluster, whose expression increases in NSCLC, and
upregulation of miR-93 was observed to be associated with poor
survival in patients with lung cancer (65). When miR-93 directly binds to the
3'-UTR of the NEDD4L mRNA, NEDD4L expression is downregulated.
Consequently, miR-93 overexpression may be a useful technique in
the pathogenesis of lung cancer (65). Reduced NEDD4L accelerated
TGF-induced epithelial-mesenchymal transition, decreased
SMAD2/SMAD3 degradation and enhanced TGF signal transmission
(63,64). Conversely, A549 cancer cells
exhibited NEDD4 expression, a specific E3 ligase for
serine/threonine-protein kinase (GCN2), which increases the
aggressiveness of tumors (58,59).
Through the action of arrestin, a GCN2-arrestin-NEDD4L axis
facilitated ubiquitin-mediated GCN2 degradation and thus protects
cells from cancer (58,59).
Another crucial factor in the role of NEDD4 in lung
cancer is the serine/threonine kinase mitogen-activated protein
kinase (MAPK) kinase kinase 5 (MEKK5), also known as apoptosis
signal-regulating kinase 1 and a member of the MAPK kinase family
apoptosis signal-regulating kinase (63,66).
It has been shown that NEDD4 is able to interact with MEKK5 through
its WW domain and overexpression and downregulation of MEKK5 had
opposite effects on NEDD4 ubiquitination (63,67).
This suggests that the MEKK5 and NEDD4 interaction may serve as a
possible inhibitory mechanism for NEDD4 migration signaling. In
addition, NEDD4 is associated with the action of nitidine chloride
(NC), an antioxidant and anti-cancer phytochemical alkaloid, and
the downregulation of NEDD4 increased NC-induced antitumor effects
(50). Although NC targets
multiple signaling pathways in different types of cancer to have a
tumor-suppressing impact, its significance in lung cancer is has
remained elusive (50). In
addition, NEDD4L is associated with the action of
dimethyl-acryl-alkannin (ALCAP2), a naturally occurring substance
and traditional Chinese medicine obtained from the roots of the
plant Lithospermum erythrorhizon (53). ALCAP2 has been shown to play a role
in cell cycle arrest and apoptosis promotion. ADC was prevented
from proliferating, migrating and invading by NEDD4L-stimulated
ubiquitination of β-catenin, which decreased cellular β-catenin and
activated the transcription of survivin, cyclin D1 and matrix
metalloproteinase-9(53). Thus,
NEDD4 and NEDD4L inhibitors may serve as novel therapeutics for
treating lung cancer, since NEDD4 and NEDD4L both have critical
roles in lung carcinogenesis and tumor progression.
Bladder cancer (BCa)
According to GLOBOCAN 2020, BCa is one of the most
common urogenital malignancies and accounts for 3% of all cancer
diagnoses worldwide (68).
Muscle-invasive BCa (MIBC) is a metastatic type of cancer with
recurrence-related characteristics. Although it is possible to
recover from non-MIBC (NMIBC), various NMIBC patients may exhibit
recurrence of NMIBC (69).
Therefore, identifying the mechanism of BCa tumorgenicity is
crucial.
The suppression of PTEN expression and the
activation of Notch-1 signaling has shown that overexpression of
the NEDD4 protein increased RT4 BCa cell proliferation and
invasion. In addition, NEDD4 inhibition suppressed Notch-1 while
activating PTEN. Therefore, the downregulation of NEDD4 in BCa
cells resulted in increased apoptosis, decreased cell migration and
decreased cell proliferation (70,71).
This emphasizes the significance of NEDD4 inhibitor research for
managing human BCa. Another proposed mechanism is that NEDD4
stabilizes Kruppel-like factor 8 expression, which reduces the
inhibitory effect of miR-132 on nuclear respiratory factor 1 and
boosts BCa cell survival, tumor development and migratory capacity
(72). Jing et al (73) hypothesized that NEDD4 caused BCa by
upregulating PDL-1 expression, which attenuates the action of
antitumor T-lymphocytes, allowing cancer cells to evade immune
monitoring more readily and enhancing their capacity for
proliferation and invasion. Fibroblast growth factor receptor 3
(FGFR3) activation enhanced NEDD4, which then regulated PD-L1
ubiquitination (73). Mutations
that activate the FGFR3 gene are highly prevalent in BCa. Jing
et al (73) sought to
elucidate the relationship between FGFR3, NEDD4 and PD-L1 during
immune surveillance. It was found that when FGFR3 was activated,
PD-L1 was downregulated in BCa cells and FGFR3 phosphorylated the
E3 ubiquitin ligase NEDD4. This accelerated NEDD4 ubiquitination
and degradation, thereby decreasing the amount of PD-L1 present at
the cell membrane. NEDD4 activity is also required for FGFR3 to
regulate the stability of PD-L1, and the stabilizing effect of
FGFR3 on PD-L1 was lost when NEDD4 was silenced or inhibited,
resulting in increased PD-L1 levels (73). Thus, NEDD4 is required for FGFR3 to
modulate the expression of PD-L1.
Furthermore, NEDD4L has a tumor-suppressing
function; downregulation of NEDD4L is observed in clear-cell renal
cell carcinoma (ccRCC), where it may have an anticancer effect by
inhibiting TGF1 signaling (74).
The modulation of tumor energy metabolism is another proposed
anticancer mechanism of NEDD4L in ccRCC (74).
Prostate cancer
Prostate cancer is the second leading cause of
cancer-associated male deaths worldwide (5). Given the improved therapeutic options
and early detection of prostate cancer utilizing prostate-specific
antigen testing, the 5-year survival rate of patients with prostate
cancer has increased significantly (74-77).
The three main types of treatment currently available for prostate
cancer are surgery, radiation and hormonal ablation therapy
(78). However, androgen
deprivation therapy resistance has led to the development of
metastatic castrate-resistant prostate cancer, which has resulted
in poor survival rates among patients with prostate cancer
(79,80). Consequently, identifying novel
therapeutic methods to treat prostate cancer is essential.
Numerous studies have supported the carcinogenic
function of NEDD4 and its overexpression in prostate cancer and
other human malignancies (75,81).
Of note, the fact that NEDD4 overexpression promotes cell growth,
migration and invasion while NEDD4 downregulation inhibits cell
growth and motility of prostate cancer cells supports the notion
that NEDD4 functions as an oncoprotein in this disease (75). It has been found that the
downregulation of androgenic receptors and PTEN causes NEDD4 to
have an oncogenic effect (82). As
a result, NEDD4 may be targeted for the treatment of prostate
cancer using a variety of inhibitors, including natural substances
such as diosgenin, a potential NEDD4 inhibitor in prostate cancer
cells, by inhibiting cell proliferation (83). In addition, aggressive prostate
tumors have lower levels of NEDD4L than benign prostatic
hyperplasia (75).
Colorectal cancer
Colorectal cancer is the third most common type of
cancer worldwide and one of the most common malignancies in both
men and women (84). Colon cancer
affects 1.2 million individuals worldwide and accounts for
>600,000 deaths annually (85).
Both hereditary and environmental factors have a significant impact
on the genesis of colorectal cancer (86). Compared to other types of cancer,
the etiology of colorectal cancer involves genetic and epigenetic
changes resulting in tumor cell growth (87). Even though 80% of patients with
colon cancer are diagnosed at an early stage, the high mortality
and recurrence rates in those with advanced-stage colorectal cancer
still pose a challenge for scientists and clinicians (88). It should be emphasized that surgery
and chemotherapy are the primary methods of treatment for colon
cancer (89). Despite the advances
in colon cancer treatment over the past few years, which have
included the adoption of cutting-edge drugs, such as
immunotherapeutics and targeted therapies, the prognosis for the
disease remains bleak (90). Thus,
there is an urgent need to develop novel individualized
medications.
There are several hypotheses explaining how NEDD4
and colon cancer are related. In 80% of colorectal cancers, NEDD4
is present and induces the ubiquitination of PTEN, which results in
the degradation of PTEN and thus increased tumor growth (91). PTEN serves as an essential negative
regulator of the PI3K/AKT signaling pathway and suppresses AKT
activation by dephosphorylating PIP3(91). Phosphoinositide 3-kinase (PI3K) is
a protein kinase that promotes cell development by phosphorylating
PIP2 to create PIP3 by activating receptor tyrosine kinases
(91). In colorectal cancer, PTEN
is frequently deleted, changed, or even hypermethylated, which
results in the PI3K/AKT signaling pathway becoming activated
(91). One study, which used
HCT-15 and LoVo colon cancer cells, revealed that PTEN and PI3K/AKT
signaling was not required for NEDD4 to increase colon cancer cell
growth (91). In their
investigation into the relationship between NEDD4 and colon cancer,
it was established that NEDD4/forkhead box A1
(FOXA1)/miR-340-5p/activating transcription factor-1 (ATF1) has a
novel function in the emergence of colon cancer. FOXA1 is known to
be necessary for the growth and development of specific
endoderm-derived organs, but its significance in the genesis of
cancer remains unclear. FOXA1 may, however, contribute to the
growth of cancer and the repression of critical cellular functions
(92,93). They proposed that NEDD4 resulted in
the ubiquitination and degradation of FOXA1, which alters the
expression of miR-340-5p and ATF1 and inhibits the progression of
colon cancer. Thus, this may shed light on the pathophysiology of
colon cancer and aid in creating novel gene-based treatments.
N-myc downstream regulated 1 (NDRG1) was
downregulated in CRC tissues, and both in vitro and in
vivo studies revealed a beneficial relationship between NDRG1
and p21(94). Zhang et al
(94) proposed that E3 ligase
NEDD4 may directly bind with and degrade p21, a cyclin-dependent
kinase inhibitor; however, NDRG1 could oppose the NEDD4-mediated
ubiquitylation of p21, increasing the effective p21 expression and
thus reducing tumor growth. Although NDRG1 is a cytoplasmic and
nuclear protein, it has been discovered to be intricately involved
in cell adhesion, differentiation, tumor growth and metastasis.
Furthermore, it is involved in autophagy and cell cycle regulation,
as it has been shown to cause cancer cell cycle arrest at
G0/G1(94). In addition, NDRG1 was
associated with improved survival rates in several malignancies,
including breast, colon, prostate and pancreatic cancer (94). Thus, this may be a target mechanism
for treating colorectal cancer.
Lu et al (95) also investigated the relationship
between NEDD4 and the WNT signaling pathway, which has various
roles in several physiological and cellular processes, including
cellular proliferation, fate and embryonic development. For
instance, it has been demonstrated that NEDD4 blocks the
transcription factors lymphoid enhancer binding factor 1 and YY1 to
at least partially limit colonic WNT signaling and tumor growth
(95).
Tanksley et al (96) reported that NEDD4L was considerably
downregulated, whereas NEDD4 was significantly upregulated in all
stages of colorectal cancer. According to the hypothesized
mechanism of action, NEDD4L acted as a tumor suppressor in
colorectal cancer by inhibiting WNT signaling at or downstream of
β-catenin (96). Unexpectedly,
patients with high NEDD4L expression have been demonstrated to
exhibit prolonged disease-specific survival. However, NEDD4L was
discovered to be highly upregulated in colon cancer (91).
Another study by Eide et al (91) linking NEDD4 to colon cancer did not
focus on how NEDD4 affected signaling mechanisms but instead showed
that NEDD4 has a significant role in regulating the actin
cytoskeleton in colon cancer cells. It was observed that the actin
cytoskeleton and changes in cell shape were brought about by NEDD4
knockdown (91). Considering all
available research, more work is necessary to fully determine the
role of NEDD4 in CRC.
Fig. 1 illustrates
the general molecular and signaling pathways of NEDD4-1 and NEDD4L
in tumor development and drug resistance.
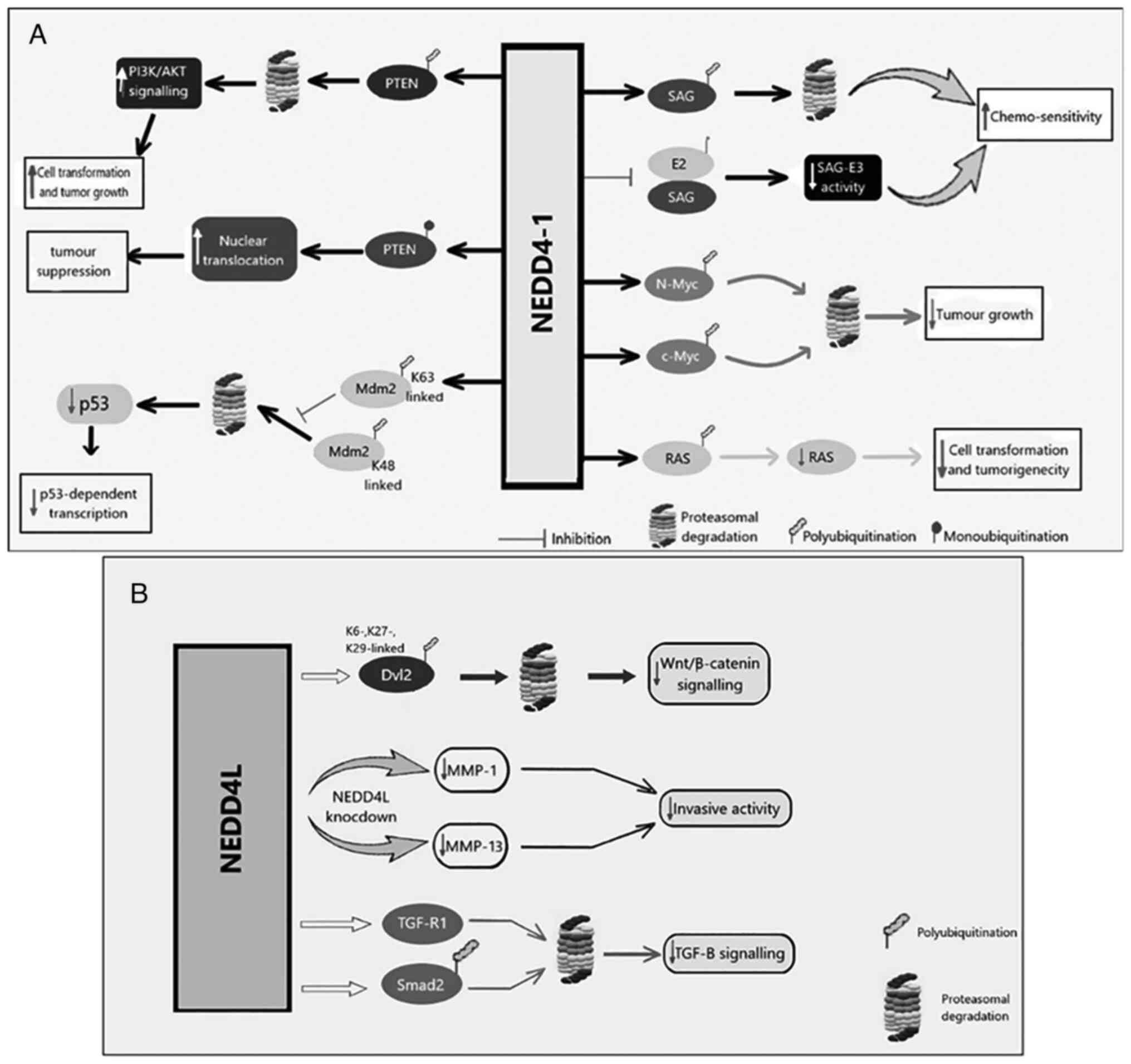 | Figure 1Molecular and signaling pathways of
NEDD4-1 and NEDD4L in tumor development and drug resistance
(93). (A) NEDD4-1 promotes the
proteasomal degradation of PTEN, thereby enhancing PI3K/AKT
signaling and tumor development (93). In addition, NEDD4-1 can
monoubiquitinate PTEN, which facilitates PTEN nuclear
translocation, possibly protects PTEN from degradation and boosts
the tumor-suppressive effects of PTEN (93). By interfering with RAS, NEDD4-1 has
an essential function in normal cells, inhibiting the
transformation and tumorigenicity (93). By increasing Mdm2-mediated
K63-linked polyubiquitination, NEDD4-1 inhibits Mdm2
autoubiquitination and reduces p53-dependent transcription
(93). NEDD4-1 also enhances
chemosensitivity by inhibiting SAG, another E3, by promoting its
degradation and preventing its binding to ubiquitin-bound E2.
Oncoproteins n-Myc and c-Myc are also destabilized by the
tumor-suppressing NEDD4-1. (B) NEDD4L downregulates TGF signaling
by ubiquitinating TGF-β-R1 and Smad2 and directing them toward
degradation. Depletion of NEDD4L can decrease MMP-1 and MMP-13
levels and, consequently, the invasiveness of gallbladder cancer
(93). Negative regulation of
Wnt/β-catenin signaling is observed when NEDD4L promotes the K6,
K27 and K29-linked polyubiquitination of Dvl2, resulting in its
degradation (93). PTEN,
phosphatase and tensin homolog; NEDD4-1, neural precursor cell
expressed developmentally downregulated protein 4; NEDD4L,
NEDD4-like E3 ubiquitin protein ligase; TGF-β-R1, transforming
growth factor-β-receptor 1; SAG, sensitive to apoptosis; Dvl2,
dishevelled segment polarity protein 2. |
Clinical studies on the regulation of
NEDD4 and NEDD4L in lung and colorectal cancers
In preclinical studies, targeting NEDD4 and NEDD4L
as a method of cancer treatment demonstrated promise. NEDD4 and
NEDD4L may function as therapeutic targets for the treatment of
various types of cancer as well as a biomarker for poor prognosis
(73,97-99).
NEDD4 may function as a tumor suppressor and an oncogene, and
NEDD4L may also function as a tumor suppressor and an oncogene, in
both cases depending on the type of cancer being assessed (97-102),
necessitating the use of NEDD4 and/or NEDD4L inhibitors or
activators in cancer therapy.
It was discovered that NEDD4 is an oncogene as it
inhibits PTEN, a known tumor suppressor; thus, PTEN levels are
decreased and NEDD4 levels are elevated in a variety of human
cancer cell lines (100). PTEN is
a phosphatase that inhibits the PI3K/Akt signaling pathway, which
is essential for cancer cell survival, to prevent the development
of tumors by cancer cells. It has been demonstrated that NEDD4 is a
PTEN-specific E3 ligase, which decreases PTEN protein levels, and
thus increases Akt signaling. Therefore, NEDD4 has been suggested
as a potential oncoprotein and tumor suppressor.
In various cancer models, inhibiting NEDD4 has been
shown to reduce tumor growth and make cancer cells more responsive
to therapy (97). Targeting NEDD4
can be accomplished by a variety of mechanisms. These include using
small chemical inhibitors and RNA interference technology (101). To prevent the degradation of
tumor suppressor proteins or to interfere with the activation of
oncogenic pathways, small molecule inhibitors may be developed to
bind to NEDD4 and limit its activity.
Although numerous preclinical studies indicate that
the NEDD4 protein possesses both oncogenic and antitumor
properties, only a small number of clinical studies support this
assertion. These clinical studies (52,54,101-103),
summarized in Table I, demonstrate
that the NEDD4 and NEDD4L proteins can be potential therapeutic
targets for preventing and treating lung and colorectal cancer.
There are currently no clinical outcomes for prostate and BCa, and
additional clinical research and trials are required.
 | Table IClinical studies on the regulation of
NEDD4 in lung cancer and CRC. |
Table I
Clinical studies on the regulation of
NEDD4 in lung cancer and CRC.
| A, Lung cancer |
|---|
| Tissue type | NEDD4/NEDD4L
expression levels | (Refs.) |
|---|
| NSCLC | NEDD4 high | (52) |
| Lung
adenocarcinoma | NEDD4 high | (54) |
| NSCLC | NEDD4L low | (101) |
| B, Colorectal
cancer |
| Tissue type | NEDD4/NEDD4L
expression levels | (Refs.) |
| CRC | NEDD4 high | (102,103) |
| CRC | NEDD4 high and
NEDD4L low | (95) |
3. Conclusions and future perspective
It is evident from combining studies that NEDD4 and
NEDD4L ligases are essential to the signaling pathways that enhance
the development of various types of cancer. However, their
importance in certain types of cancers remains to be determined. In
most cancer types, both of NEDD4 and NEDD4L slow carcinogenesis by
increasing the ubiquitination and degradation of their substrates,
which play vital oncogenic and tumor-suppressing roles in a range
of malignancies. Given their functions in carcinogenesis, NEDD4 and
NEDD4L are therefore potential targets for developing novel
treatments for the management of anti-cancer agents. Treatment
strategies that interfere with NEDD4 and NEED4L interacting with
their substrates with no/few adverse effects may be preferable to
those that directly target NEDD4 and NEDD4L activity, given the
wide range of targets and the duality of their roles. However,
additional research is required to fully determine the significance
of NEDD4 in carcinogenesis.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
AZA designed and performed the narrative review, and
drafted and proofread the article critically. KA helped in writing
the section on colorectal cancer and critically revised the
manuscript. GBH helped in writing the conclusion section and
revised the article critically for intellectual content. RAS
contributed to writing the section on lung cancer. Rawan AD helped
in writing the section on BCa. MO helped in writing the section on
prostate cancer. AA and YAE assisted in reviewing the manuscript
critically. All authors have read and approved the final version of
the manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cooper GM: The Cell: A Molecular Approach.
2nd edition. Sunderland (MA): Sinauer Associates. The Development
and Causes of Cancer. https://www.ncbi.nlm.nih.gov/books/NBK9963/. 2000.
|
|
2
|
Hanahan D: Hallmarks of cancer: New
dimensions. Cancer Discov. 12:31–46. 2022.PubMed/NCBI View Article : Google Scholar
|
|
3
|
American Cancer Society. Cancer Statistics
Center. http://cancerstatisticscenter.cancer.org. Accessed Dec
25, 2022.
|
|
4
|
World Health Organization (WHO): Editorial
Style Manual WHO, Geneva, p83-91, 1993.
|
|
5
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249.
2021.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Siegel RL, Miller KD, Fuchs HE and Jemal
A: Cancer statistics, 2022. CA Cancer J Clin. 72:7–33.
2022.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Bertram JS: The molecular biology of
cancer. Mol Aspects Med. 21:167–223. 2000.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kontomanolis EN, Koutras A, Syllaios A,
Schizas D, Mastoraki A, Garmpis N, Diakosavvas M, Angelou K,
Tsatsaris G, Pagkalos A, et al: Role of oncogenes and
tumor-suppressor genes in carcinogenesis: A review. Anticancer Res.
40:6009–6015. 2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Welti J, Loges S, Dimmeler S and Carmeliet
P: Recent molecular discoveries in angiogenesis and antiangiogenic
therapies in cancer. J Clin Invest. 123:3190–3200. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
10
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kamali H, Golmohammadzadeh S, Zare H,
Nosrati R, Fereidouni M and Safarpour H: The recent advancements in
the early detection of cancer biomarkers by DNAzyme-assisted
aptasensors. J Nanobiotechnology. 20(438)2022.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Pan S and Chen R: Pathological implication
of protein post-translational modifications in cancer. Mol Aspects
Med. 86(101097)2022.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Hershko A and Ciechanover A: The ubiquitin
system. Annu Rev Biochem. 67:425–479. 1998.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Weissman AM: Themes and variations on
ubiquitylation. Nat Rev Mol Cell Biol. 2:169–178. 2001.PubMed/NCBI View
Article : Google Scholar
|
|
16
|
Komander D and Rape M: The ubiquitin code.
Annu Rev Biochem. 81:203–229. 2012.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Salas-Lloret D and González-Prieto R:
Insights in post-translational modifications: Ubiquitin and SUMO.
Int J Mol Sci. 23(3281)2022.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Liebl MP and Hoppe T: It's all about
talking: Two-way communication between proteasomal and lysosomal
degradation pathways via ubiquitin. Am J Physiol Cell Physiol.
311:C166–C178. 2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Tu Y, Chen C, Pan J, Xu J, Zhou ZG and
Wang CY: The Ubiquitin Proteasome Pathway (UPP) in the regulation
of cell cycle control and DNA damage repair and its implication in
tumorigenesis. Int J Clin Exp Pathol. 5:726–738. 2012.PubMed/NCBI
|
|
20
|
Klionsky DJ and Schulman BA: Dynamic
regulation of macroautophagy by distinctive ubiquitin-like
proteins. Nat Struct Mol Biol. 21:336–345. 2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Kerscher O, Felberbaum R and Hochstrasser
M: Modification of proteins by ubiquitin and ubiquitin-like
proteins. Annu Rev Cell Dev Biol. 22:159–180. 2006.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Ciechanover A and Kwon YT: Degradation of
misfolded proteins in neurodegenerative diseases: Therapeutic
targets and strategies. Exp Mol Med. 47(e147)2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Berndsen CE and Wolberger C: New insights
into ubiquitin E3 ligase mechanism. Nat Struct Mol Biol.
21:301–307. 2014.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Kravtsova-Ivantsiv Y and Ciechanover A:
Non-canonical ubiquitin-based signals for proteasomal degradation.
J Cell Sci. 125(Pt 3):539–548. 2012.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Buetow L and Huang DT: Structural insights
into the catalysis and regulation of E3 ubiquitin ligases. Nat Rev
Mol Cell Biol. 17:626–642. 2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Rotin D and Kumar S: Physiological
functions of the HECT family of ubiquitin ligases. Nat Rev Mol Cell
Biol. 10:398–409. 2009.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Metzger MB, Hristova VA and Weissman AM:
HECT and RING finger families of E3 ubiquitin ligases at a glance.
J Cell Sci. 125(Pt 3):531–537. 2012.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Joazeiro CA and Weissman AM: RING finger
proteins: Mediators of ubiquitin ligase activity. Cell.
102:549–552. 2000.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Bernassola F, Karin M, Ciechanover A and
Melino G: The HECT family of E3 ubiquitin ligases: Multiple players
in cancer development. Cancer Cell. 14:10–21. 2008.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Dove KK, Stieglitz B, Duncan ED, Rittinger
K and Klevit RE: Molecular insights into RBR E3 ligase ubiquitin
transfer mechanisms. EMBO Rep. 17:1221–1235. 2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Li W and Ye Y: Polyubiquitin chains:
Functions, structures, and mechanisms. Cell Mol Life Sci.
65:2397–2406. 2008.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Dinudom A, Harvey KF, Komwatana P, Young
JA, Kumar S and Cook DI: Nedd4 mediates control of an epithelial
Na+ channel in salivary duct cells by cytosolic Na+. Proc Natl Acad
Sci USA. 95:7169–7173. 1998.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Goulet CC, Volk KA, Adams CM, Prince LS,
Stokes JB and Snyder PM: Inhibition of the epithelial Na+ channel
by interaction of Nedd4 with a PY motif deleted in Liddle's
syndrome. J Biol Chem. 273:30012–30017. 1998.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Harvey KF, Dinudom A, Komwatana P,
Jolliffe CN, Day ML, Parasivam G, Cook DI and Kumar S: All three WW
domains of murine Nedd4 are involved in the regulation of
epithelial sodium channels by intracellular Na+. J Biol Chem.
274:12525–12530. 1999.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Harvey KF, Dinudom A, Cook DI and Kumar S:
The Nedd4-like protein KIAA0439 is a potential regulator of the
epithelial sodium channel. J Biol Chem. 276:8597–8601.
2001.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Kanelis V, Farrow NA, Kay LE, Rotin D and
Forman-Kay JD: NMR studies of tandem WW domains of Nedd4 in complex
with a PY motif-containing region of the epithelial sodium channel.
Biochem Cell Biol. 76:341–350. 1998.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ingham RJ, Gish G and Pawson T: The Nedd4
family of E3 ubiquitin ligases: Functional diversity within a
common modular architecture. Oncogene. 23:1972–1984.
2004.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Dodson EJ, Fishbain-Yoskovitz V,
Rotem-Bamberger S and Schueler-Furman O: Versatile communication
strategies among tandem WW domain repeats. Exp Biol Med (Maywood).
240:351–360. 2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Scheffner M and Kumar S: Mammalian HECT
ubiquitin-protein ligases: Biological and pathophysiological
aspects. Biochim Biophys Acta. 1843:61–74. 2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Lee IH, Dinudom A, Sanchez-Perez A, Kumar
S and Cook DI: Akt mediates the effect of insulin on epithelial
sodium channels by inhibiting Nedd4-2. J Biol Chem.
282:29866–29873. 2007.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Goel P, Manning JA and Kumar S: NEDD4-2
(NEDD4L): The ubiquitin ligase for multiple membrane proteins.
Gene. 557:1–10. 2015.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Lu X, Xu H, Xu J, Lu S, You S, Huang X,
Zhang N and Zhang L: The regulatory roles of the E3 ubiquitin
ligase NEDD4 family in DNA damage response. Front Physiol.
13(968927)2022.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Chaugule VK and Walden H: Specificity and
disease in the ubiquitin system. Biochem Soc Trans. 44:212–227.
2016.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Herrmann J, Soares SM, Lerman LO and
Lerman A: Potential role of the ubiquitin-proteasome system in
atherosclerosis: Aspects of a protein quality disease. J Am Coll
Cardiol. 51:2003–2010. 2008.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Potjewyd FM and Axtman AD: Exploration of
Aberrant E3 ligases implicated in Alzheimer's disease and
development of chemical tools to modulate their function. Front
Cell Neurosci. 15(768655)2021.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Sun T, Liu Z and Yang Q: The role of
ubiquitination and deubiquitination in cancer metabolism. Mol
Cancer. 19(146)2020.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Petroski MD: The ubiquitin system,
disease, and drug discovery. BMC Biochem. 9 (Suppl
1)(S7)2008.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Orlowski RZ and Kuhn DJ: Proteasome
inhibitors in cancer therapy: Lessons from the first decade. Clin
Cancer Res. 14:1649–1657. 2008.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Thandra KC and Barsouk A, Saginala K,
Aluru JS and Barsouk A: Epidemiology of lung cancer. Contemp Oncol
(Pozn). 25:45–52. 2021.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Zhang J, Cao R, Lian C, Cao T, Shi Y, Ma
J, Wang P and Xia J: Nitidine chloride suppresses NEDD4 expression
in lung cancer cells. Aging (Albany NY). 13:782–793.
2020.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Rajdev K, Siddiqui AH, Ibrahim U,
Patibandla P, Khan T and El-Sayegh D: An unusually aggressive large
cell carcinoma of the lung: Undiagnosed until autopsy. Cureus.
10(e2202)2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Amodio N, Scrima M, Palaia L, Salman AN,
Quintiero A, Franco R, Botti G, Pirozzi P, Rocco G, De Rosa N and
Viglietto G: Oncogenic role of the E3 ubiquitin ligase NEDD4-1, a
PTEN negative regulator, in non-small-cell lung carcinomas. Am J
Pathol. 177:2622–2634. 2010.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Zhang W, Zhang R, Zeng Y, Li Y, Chen Y,
Zhou J, Zhang Y, Wang A, Zhu J, Liu Z, et al: ALCAP2 inhibits lung
adenocarcinoma cell proliferation, migration and invasion via the
ubiquitination of β-catenin by upregulating the E3 ligase NEDD4L.
Cell Death Dis. 12(755)2021.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Song YH, Zhang CQ, Chen FF and Lin XY:
Upregulation of neural precursor cell expressed developmentally
downregulated 4-1 is associated with poor prognosis and
chemoresistance in lung adenocarcinoma. Chin Med J (Engl).
131:16–24. 2018.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Chen Z, Fillmore CM, Hammerman PS, Kim CF
and Wong KK: Non-small-cell lung cancers: A heterogeneous set of
diseases. Nat Rev Cancer. 14:535–546. 2014.PubMed/NCBI View Article : Google Scholar
|
|
56
|
He H, Huang C, Chen Z, Huang H, Wang X and
Chen J: An outlined review for the role of Nedd4-1 and Nedd4-2 in
lung disorders. Biomed Pharmacother. 125(109983)2020.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Georgescu MM: PTEN tumor suppressor
network in PI3K-Akt pathway control. Genes Cancer. 1:1170–1177.
2010.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Wei C, Lin M, Jinjun B, Su F, Dan C, Yan
C, Jie Y, Jin Z, Zi-Chun H and Wu Y: Involvement of general control
nonderepressible kinase 2 in cancer cell apoptosis by
posttranslational mechanisms. Mol Biol Cell. 26:1044–1057.
2015.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Shao G, Wang R, Sun A, Wei J, Peng K, Dai
Q, Yang W and Lin Q: The E3 ubiquitin ligase NEDD4 mediates cell
migration signaling of EGFR in lung cancer cells. Mol Cancer.
17(24)2018.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Sakashita H, Inoue H, Akamine S, Ishida T,
Inase N, Shirao K, Mori M and Mimori K: Identification of the
NEDD4L gene as a prognostic marker by integrated microarray
analysis of copy number and gene expression profiling in non-small
cell lung cancer. Ann Surg Oncol. 20 (Suppl 3):S590–S598.
2013.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Li G, Song Z, Wu C, Li X, Zhao L, Tong B,
Guo Z, Sun M, Zhao J, Zhang H, et al: Downregulation of NEDD4L by
EGFR signaling promotes the development of lung adenocarcinoma. J
Transl Med. 20(47)2022.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Li R, Hu Z, Sun SY, Chen ZG, Owonikoko TK,
Sica GL, Ramalingam SS, Curran WJ, Khuri FR and Deng X: Niclosamide
overcomes acquired resistance to erlotinib through suppression of
STAT3 in non-small cell lung cancer. Mol Cancer Ther. 12:2200–2212.
2013.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Xie S, Xia L, Song Y, Liu H, Wang ZW and
Zhu X: Insights into the biological role of NEDD4L E3 ubiquitin
ligase in human cancers. Front Oncol. 11(774648)2021.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Lai XN, Li J, Tang LB, Chen WT, Zhang L
and Xiong LX: MiRNAs and LncRNAs: Dual roles in TGF-β
signaling-regulated metastasis in lung cancer. Int J Mol Sci.
21(1193)2020.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Qu MH, Han C, Srivastava AK, Cui T, Zou N,
Gao ZQ and Wang QE: miR-93 promotes TGF-β-induced
epithelial-to-mesenchymal transition through downregulation of
NEDD4L in lung cancer cells. Tumour Biol. 37:5645–5651.
2016.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Bai X, Jing L, Li Y, Li Y, Luo S, Wang S,
Zhou J, Liu Z and Diao A: TMEPAI inhibits TGF-β signaling by
promoting lysosome degradation of TGF-β receptor and contributes to
lung cancer development. Cell Signal. 26:2030–2039. 2014.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Sun A, Zhu J, Xia S, Li Y, Wu T, Shao G,
Yang W and Lin Q: MEKK5 interacts with and negatively regulates the
E3 ubiquitin ligase NEDD4 for mediating lung cancer cell migration.
Life (Basel). 11(1153)2021.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Saginala K and Barsouk A, Aluru JS, Rawla
P, Padala SA and Barsouk A: Epidemiology of bladder cancer. Med Sci
(Basel). 8(15)2020.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Sanli O, Dobruch J, Knowles MA, Burger M,
Alemozaffar M, Nielsen ME and Lotan Y: Bladder cancer. Nat Rev Dis
Primers. 3(17022)2017.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Wen W, Li J, Wang L, Xing Y, Li X, Ruan H,
Xi X, Xiong J and Kuang R: Inhibition of NEDD4 inhibits cell growth
and invasion and induces cell apoptosis in bladder cancer cells.
Cell Cycle. 16:1509–1514. 2017.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Wang X, Trotman LC, Koppie T, Alimonti A,
Chen Z, Gao Z, Wang J, Erdjument-Bromage H, Tempst P, Cordon-Cardo
C, et al: NEDD4-1 is a proto-oncogenic ubiquitin ligase for PTEN.
Cell. 128:129–139. 2007.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Mao M, Yang L, Hu J, Liu B, Zhang X, Liu
Y, Wang P and Li H: Oncogenic E3 ubiquitin ligase NEDD4 binds to
KLF8 and regulates the microRNA-132/NRF2 axis in bladder cancer.
Exp Mol Med. 54:47–60. 2022.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Jing W, Wang G, Cui Z, Xiong G, Jiang X,
Li Y, Li W, Han B, Chen S and Shi B: FGFR3 Destabilizes PD-L1 via
NEDD4 to Control T-cell-mediated bladder cancer immune
surveillance. Cancer Res. 82:114–129. 2022.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Zhao H, Zhang J, Fu X, Mao D, Qi X, Liang
S, Meng G, Song Z, Yang R, Guo Z, et al: Integrated bioinformatics
analysis of the NEDD4 family reveals a prognostic value of
NEDD4L in clear-cell renal cell cancer. PeerJ.
9(e11880)2021.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Hu XY, Xu YM, Fu Q, Yu JJ and Huang J:
Nedd4L expression is downregulated in prostate cancer compared to
benign prostatic hyperplasia. Eur J Surg Oncol. 35:527–531.
2009.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Negoita S, Feuer EJ, Mariotto A, Cronin
KA, Petkov VI, Hussey SK, Benard V, Henley SJ, Anderson RN, Fedewa
S, et al: Annual report to the nation on the status of cancer, part
II: Recent changes in prostate cancer trends and disease
characteristics. Cancer. 124:2801–2814. 2018.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Butler SS, Muralidhar V, Zhao SG, Sanford
NN, Franco I, Fullerton ZH, Chavez J, D'Amico AV, Feng FY, Rebbeck
TR, et al: Prostate cancer incidence across stage, NCCN risk
groups, and age before and after USPSTF Grade D recommendations
against prostate-specific antigen screening in 2012. Cancer.
126:717–724. 2020.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Miller KD, Siegel RL, Lin CC, Mariotto AB,
Kramer JL, Rowland JH, Stein KD, Alteri R and Jemal A: Cancer
treatment and survivorship statistics, 2016. CA Cancer J Clin.
66:271–289. 2016.PubMed/NCBI View Article : Google Scholar
|
|
79
|
McCrea E, Sissung TM, Price DK, Chau CH
and Figg WD: Androgen receptor variation affects prostate cancer
progression and drug resistance. Pharmacol Res. 114:152–162.
2016.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Zhang Y, Duan H, Zhao H, Qi L, Liu Y,
Zhang Z, Liu C, Chen L, Jin M, Guan Y, et al: Development and
evaluation of a PSMA-targeted nanosystem co-packaging docetaxel and
androgen receptor siRNA for castration-resistant prostate cancer
treatment. Pharmaceutics. 14(964)2022.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Zhao Y, Li J, Chen J, Ye M and Jin X:
Functional roles of E3 ubiquitin ligases in prostate cancer. J Mol
Med (Berl). 100:1125–1144. 2022.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Li H, Xu LL, Masuda K, Raymundo E, McLeod
DG, Dobi A and Srivastava S: A feedback loop between the androgen
receptor and a NEDD4-binding protein, PMEPA1, in prostate cancer
cells. J Biol Chem. 283:28988–28995. 2008.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Zhang J, Xie JJ, Zhou SJ, Chen J, Hu Q, Pu
JX and Lu JL: Diosgenin inhibits the expression of NEDD4 in
prostate cancer cells. Am J Transl Res. 11:3461–3471.
2019.PubMed/NCBI
|
|
84
|
Kim SE, Paik HY, Yoon H, Lee JE, Kim N and
Sung MK: Sex- and gender-specific disparities in colorectal cancer
risk. World J Gastroenterol. 21:5167–5175. 2015.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Xi Y and Xu P: Global colorectal cancer
burden in 2020 and projections to 2040. Transl Oncol.
14(101174)2021.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Rattray NJW, Charkoftaki G, Rattray Z,
Hansen JE, Vasiliou V and Johnson CH: Environmental influences in
the etiology of colorectal cancer: The premise of metabolomics.
Curr Pharmacol Rep. 3:114–125. 2017.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Parmar S and Easwaran H: Genetic and
epigenetic dependencies in colorectal cancer development.
Gastroenterol Rep (Oxf). 10(goac035)2022.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Hsieh MH, Kung PT, Kuo WY, Ke TW and Tsai
WC: Recurrence, death risk, and related factors in patients with
stage 0 colorectal cancer: A nationwide population-based study.
Medicine (Baltimore). 99(e21688)2020.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Mishra J, Drummond J, Quazi SH, Karanki
SS, Shaw JJ, Chen B and Kumar N: Prospective of colon cancer
treatments and scope for combinatorial approach to enhanced cancer
cell apoptosis. Crit Rev Oncol Hematol. 86:232–250. 2013.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Golshani G and Zhang Y: Advances in
immunotherapy for colorectal cancer: A review. Therap Adv
Gastroenterol. 13(1756284820917527)2020.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Eide PW, Cekaite L, Danielsen SA,
Eilertsen IA, Kjenseth A, Fykerud TA, Ågesen TH, Bruun J, Rivedal
E, Lothe RA and Leithe E: NEDD4 is overexpressed in colorectal
cancer and promotes colonic cell growth independently of the
PI3K/PTEN/AKT pathway. Cell Signal. 25:12–18. 2013.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Aikemu B, Shao Y, Yang G, Ma J, Zhang S,
Yang X, Hong H, Yesseyeva G, Huang L, Jia H, et al: NDRG1 regulates
Filopodia-induced Colorectal Cancer invasiveness via modulating
CDC42 activity. Int J Biol Sci. 17:1716–1730. 2021.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Zou X, Levy-Cohen G and Blank M: Molecular
functions of NEDD4 E3 ubiquitin ligases in cancer. Biochim Biophys
Acta. 1856:91–106. 2015.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Zhang S, Yu C, Yang X, Hong H, Lu J, Hu W,
Hao X, Li S, Aikemu B, Yang G, et al: N-myc downstream-regulated
gene 1 inhibits the proliferation of colorectal cancer through
emulative antagonizing NEDD4-mediated ubiquitylation of p21. J Exp
Clin Cancer Res. 38(490)2019.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Lu C, Thoeni C, Connor A, Kawabe H,
Gallinger S and Rotin D: Intestinal knockout of Nedd4 enhances
growth of Apcmin tumors. Oncogene. 35:5839–5849.
2016.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Tanksley JP, Chen X and Coffey RJ: NEDD4L
is downregulated in colorectal cancer and inhibits canonical WNT
signaling. PLoS One. 8(e81514)2013.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Liu J, Shaik S, Dai X, Wu Q, Zhou X, Wang
Z and Wei W: Targeting the ubiquitin pathway for cancer treatment.
Biochim Biophys Acta. 1855:50–60. 2015.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Wang K, Yu Y, Wang W, Jiang Y, Li Y, Jiang
X, Qiao Y, Chen L, Zhao X, Liu J, et al: Targeting the E3 ligase
NEDD4 as a novel therapeutic strategy for IGF1 signal
pathway-driven gastric cancer. Oncogene. 42:1072–1087.
2023.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Ahn Y, Hwang CY, Lee SR, Kwon KS and Lee
C: The tumour suppressor PTEN mediates a negative regulation of the
E3 ubiquitin-protein ligase Nedd4. Biochem J. 412:331–338.
2008.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Ao N, Chen Q and Liu G: The small
molecules targeting ubiquitin-proteasome system for cancer therapy.
Comb Chem High Throughput Screen. 20:403–413. 2017.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Wang X, Duan J, Fu W, Yin Z, Sheng J, Lei
Z and Wang H: Decreased expression of NEDD4L contributes to NSCLC
progression and metastasis. Biochem Biophys Res Commun.
513:398–404. 2019.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Yue M, Yun Z, Li S, Yan G and Kang Z:
NEDD4 triggers FOXA1 ubiquitination and promotes colon cancer
progression under microRNA-340-5p suppression and ATF1
upregulation. RNA Biol. 18:1981–1995. 2021.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Kim SS, Yoo NJ, Jeong EG, Kim MS and Lee
SH: Expression of NEDD4-1, a PTEN regulator, in gastric and
colorectal carcinomas. APMIS. 116:779–784. 2008.PubMed/NCBI View Article : Google Scholar
|














