Introduction
Lung cancer is the leading cause of cancer-related
mortality worldwide, with an exceedingly poor prognosis (1); of these, non-small cell lung cancer
(NSCLC) accounts for ~85% (2). The
most common lung cancer therapies include surgery, chemotherapy,
radiotherapy, targeted therapy and immunotherapy; nevertheless,
these treatments do not significantly improve the long-term
survival of patients (3). In this
view, substantial research has been devoted to developing safe and
effective antitumor drugs, including nanoparticles (4,5) and
Chinese herbs. Ailanthone (AIL) is one of the most significant
active constituents of ailanthus, a traditional Chinese herbal
medicine with anti-inflammatory, anti-allergic, anti-malaria and
antitumor properties. Previous studies have identified that AIL
suppresses various tumors, including glioma, liver carcinoma and
leukemia by augmenting apoptosis, autophagy and cell cycle arrest
(6-8).
However, the effects and mechanisms by which AIL acts on lung
cancer have not been fully elucidated.
Autophagy is a self-eating homeostatic catabolic
process for the final degradation of intracellular components in
lysosomes regulated by the autophagy-related gene (ATG) (9). Autophagy maintains optimal cellular
homeostasis under physiological conditions. However, excessive
autophagy can result in autophagic cell death and the activation of
autophagy signaling pathways (10). Pharmacological agents such as
natural small molecules or Chinese herb extracts have been
demonstrated to cause autophagic cell death in several cancer
types, including NSCLC (11,12).
Ferroptosis is a form of iron-dependent programmed cell death that
differs from necrosis, apoptosis and autophagy by lipid
peroxidation (LPO) and iron accumulation (13). Guo et al (14) revealed that a combination of
cisplatin and the ferroptosis inducer erastin exhibited increased
antitumor activity. In this view, ferroptosis has emerged as an
attractive strategy for cancer treatment, with significant
implications for lowering lung cancer recurrence and mortality
rates, as well as improving patient prognosis. Evidence indicates
that an intricate crosstalk exists between autophagy and
ferroptosis (15), that
ferroptosis is dependent on autophagy, and that excessive autophagy
cell activation can increase ferroptosis by degrading ferritin in
fibroblasts and cancer cells (16).
AMP-activated protein kinase (AMPK) is a highly
conserved serine/threonine protein kinase that acts as an energy
sensor, regulating energy metabolism and the ATP-generating rate
(17). Recent research indicated
that AMPK is implicated in various programmed cell death
mechanisms, including apoptosis, necroptosis, autophagy and
ferroptosis, depending on the environmental stimuli (18). Activation of the AMPK/mTOR/p70S6k
signaling pathway enhances autophagy, promotes ferritin
degradation, increases the labile iron pool, and accelerates the
accumulation of intracellular reactive oxygen species (ROS),
eventually causing ferroptosis (19). These findings demonstrated a
potential link between the AMPK/mTOR/p70S6k signaling pathway with
autophagy and ferroptosis pathways. The present study investigated
the role of AIL in inhibiting cell proliferation, promoting cell
autophagy and ferroptosis, and regulating the AMPK/mTOR/p70S6k
signaling pathway. The findings provide the theoretical groundwork
for the development of a novel drug and the clinical application of
AIL.
Materials and methods
Cell culture
Mouse NSCLC Lewis cells were purchased from
Guangzhou Geneo Biotechnology (https://www.jennio-bio.com/product/829.html?productCateId=59).
Meanwhile, mouse normal lung epithelial TC-1 cells were obtained
from Procell Life Science & Technology Co., Ltd. and used as a
control. All cells were cultured in complete DMEM containing 10%
fetal bovine serum, 100 U/ml penicillin, and 100 µg/ml streptomycin
(cat. no. P1400-100ml; Beijing Solarbio Science & Technology
Co., Ltd.) at 37˚C in a humidified incubator containing 5%
CO2.
MTT assays
Lewis (1.5x104/well) and TC-1 cells
(5.0x103/well) were seeded in a 96-well plate, incubated
for 24 h, and then treated with a gradient concentration of AIL for
24, 36 and 48 h. AIL was purchased from Chengdu Alfa Biological
Technology Co. Ltd. (Chengdu, China). Subsequently, 20 µl of MTT at
5 mg/ml was added to each well and incubated for 4 h. The medium
was then replaced with 100 µl DMSO to solubilize the formazan
crystals. Absorbance was measured at 490 nm wavelength using a
microplate reader (BioTek Instruments Inc.).
Colony formation assay
Lewis cells (1.0x103 cells) were seeded
in each 60 mm culture dish and incubated for 24 h, and then treated
with varying concentrations of AIL (0, 2.5, 5 and 10 µM) for 24 h.
Cells were washed in PBS and incubated at 37˚C in a complete growth
medium for two weeks to form clones. The cells were then fixed with
2 ml methanol and stained using crystal violet solution (0.1%) for
30 min at room temperature. Cell survival ability was evaluated by
counting clones on each plate. The number of clones forming >50
cells was calculated by ImageJ v.1.8.0 software (National
Institutes of Health).
Western blotting
Lewis cells were cultured to 80% confluence in 60 mm
dishes and treated with various concentrations of AIL (0, 2.5, 5
and 10 µM) for 24 h. Cells were lysed in RIPA buffer (Beijing
Solarbio Science & Technology Co., Ltd.) containing PMSF (RIPA:
PMSF=100:1). Protein concentrations were then quantified using a
BCA protein assay kit (cat. no. PC0020-50; Beijing Solarbio Science
& Technology Co., Ltd.). A 20-microgram amount of protein was
loaded on SDS-PAGE and transferred onto a nitrocellulose membrane.
The membrane was blocked with 5% BSA (Beijing Solarbio Science
& Technology Co., Ltd.) at 37˚C for 3 h, and then incubated
overnight at 4˚C with the following primary antibodies: Rabbit
microtubule-associated protein light chain3B monoclonal antibody
(LC3B; 1:2,000), rabbit sequestosome1 monoclonal antibody
(SQSTM1/P62; 1:10,000), rabbit autophagy-related protein 6
polyclonal antibody (ATG6/beclin1; 1:2,000), rabbit
autophagy-related protein 5 monoclonal antibody (ATG5; 1:1,000),
rabbit solute carrier family 7 member 11 monoclonal antibody
(SLC7A11/xCT; 1:10,000), rabbit glutathione peroxidase 4 monoclonal
antibody (GPX4; 1:2,000), rabbit ferritin heavy chain monoclonal
antibody (FTH; 1:2,000), rabbit transferrin receptor monoclonal
antibody (TFRC; 1:2,000), rabbit phospho-AMPK monoclonal antibody
(p-AMPK; 1:500), rabbit AMPK monoclonal antibody (1:500), rabbit
phospho-mammalian target of rapamycin monoclonal antibody (p-mTOR;
1:500), rabbit mTOR monoclonal antibody (1:10,000), rabbit
phospho-ribosomal protein S6 kinase monoclonal antibody (p-p70S6K;
1:2,000), rabbit ribosomal monoclonal antibody (p70S6K; 1:10,000),
rabbit nuclear receptor coactivator 4 monoclonal antibody (NCOA4;
1:2,000), and rabbit β-actin monoclonal antibody (1:10,000). After
washing with Tris-buffered saline containing 0.05% Tween 20 (TBST),
the membranes were incubated with HRP-coupled secondary antibodies
(1:5,000; cat. no. AS014; ABclonal Biotech Co., Ltd.) for 2 h at
room temperature and visualized with the enhanced chemiluminescence
system (Zhongshi Gene Technology Co., Ltd.; http://www.zsgentech.com/). Blots were scanned and
examined using ImageJ v.1.8.0 software (National Institutes of
Health).
Immunofluorescence
Lewis cells were seeded into 24-well plates
(5x104 cells per well) and incubated for 24 h. The cells
were treated with gradient concentrations of AIL (0, 2.5, 5 and 10
µM) for 24 h, fixed with 4% paraformaldehyde for 30 min at room
temperature, and solubilized in 0.1% Triton X-100 for 10 min. The
cells were then blocked with 5% BSA for 1 h at room temperature and
incubated with primary antibodies [rabbit LC3B monoclonal antibody
(1:100) and rabbit P62 monoclonal antibody (1:100)] at 4˚C for 4 h,
and fluorescently labeled with anti-rabbit secondary antibodies
(1:1,000; cat. no. AS011; ABclonal Biotech Co., Ltd.) for 2 h in
the dark. Subsequently, cells were stained with DAPI (5 mg/ml; cat.
no. C1005; Beyotime Institute of Biotechnology) at room temperature
for 3 min, and examined under a fluorescence microscope (Olympus
Corporation).
ROS assay
ROS generation was evaluated using a ROS assay kit
(cat. no. S0033S; Beijing Solarbio Science & Technology Co.,
Ltd.) according to the manufacturer's protocol. Lewis cells were
seeded into 6-well plates (5x105 cells per well) and
treated on the next day with gradient concentrations of AIL (0,
2.5, 5 and 10 µM) for 24 h. Subsequently, 10 µM
2,7-dichlorodi-hydrofluorescein diacetate (DCFH-DA) was added into
each well and incubated in the dark for 20 min. The cells were
washed with serum-free medium 3 times, digested, and transferred to
a 96-well plate at the same density. The fluorescence intensity was
measured at a 488 nm excitation and 525 nm emission wavelength on a
fluorescence microplate reader (BioTek Instruments, Inc.). In
addition, fluorescence was also directly observed under a
fluorescence microscope (Olympus Corporation).
Determination of ferroptosis-related
markers
Intracellular levels of Fe2+, LPO,
malondialdehyde (MDA), glutathione (GSH), total superoxide
dismutase (T-SOD) and catalase (CAT) were determined using the
Fe2+ assay kit (cat. no. A039-2-1), LPO assay kit (cat.
no. A106-1-1), MDA assay kit (cat. no. A003-1-1), GSH assay kit
(cat. no. A061-1-2), T-SOD assay kit (cat. no. A001-1-1) and CAT
assay kit (cat. no. A007-1-1), respectively, following the
manufacturer's protocols. All the kits were purchased from Nanjing
Jiancheng Bioengineering Institute.
Statistical analysis
All experiments were repeated three times. SPSS 22.0
software (IBM Corp.) was employed for statistical analysis. Data
are presented as the mean ± standard deviation. Differences between
AIL concentration groups were compared using one-way ANOVA followed
by Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
AIL inhibits the growth of NSCLC Lewis
cells
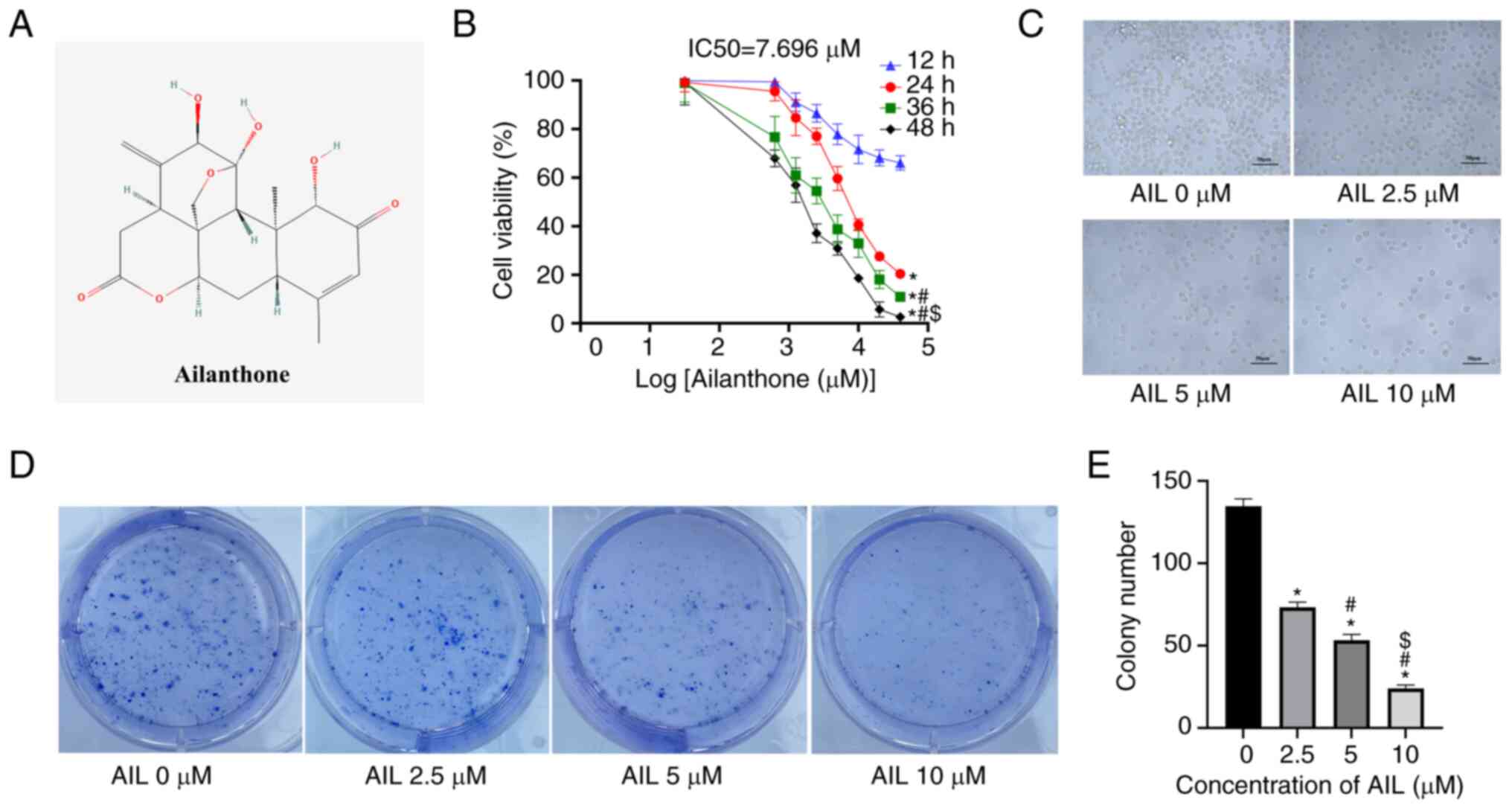
The chemical structure of AIL is depicted in
Fig. 1A (PubChem ID, 72965). Lewis
and TC-1 cells were treated with a gradient concentration of AIL
for different time points (12, 24, 36 and 48 h), and cell viability
was assessed by MTT. The findings demonstrated that AIL suppressed
Lewis cell viability in a dose- and time-dependent manner (Fig. 1B); the IC50 value after
24 h of AIL treatment was 7.696±0.327 µM. Interestingly, AIL did
not alter mouse normal lung epithelial cell (TC-1) growth (Fig. S1). Next, Lewis cells were treated
with 0, 2.5, 5 or 10 µM AIL for 24 h and their morphology was
examined by light microscopy. The cells exhibited poor adhesion and
refractivity ability and uneven-size shrunk cells with an increase
in AIL concentration (Fig. 1C).
Furthermore, the influence of AIL on cell proliferation was
evaluated using the cell colony formation test. The results
demonstrated that the number of cell colony formations decreased
significantly as the AIL dosage increased (F=0.680, P<0.001;
Fig. 1D and E). These findings demonstrated that AIL
significantly inhibited the growth of NSCLC Lewis cells.
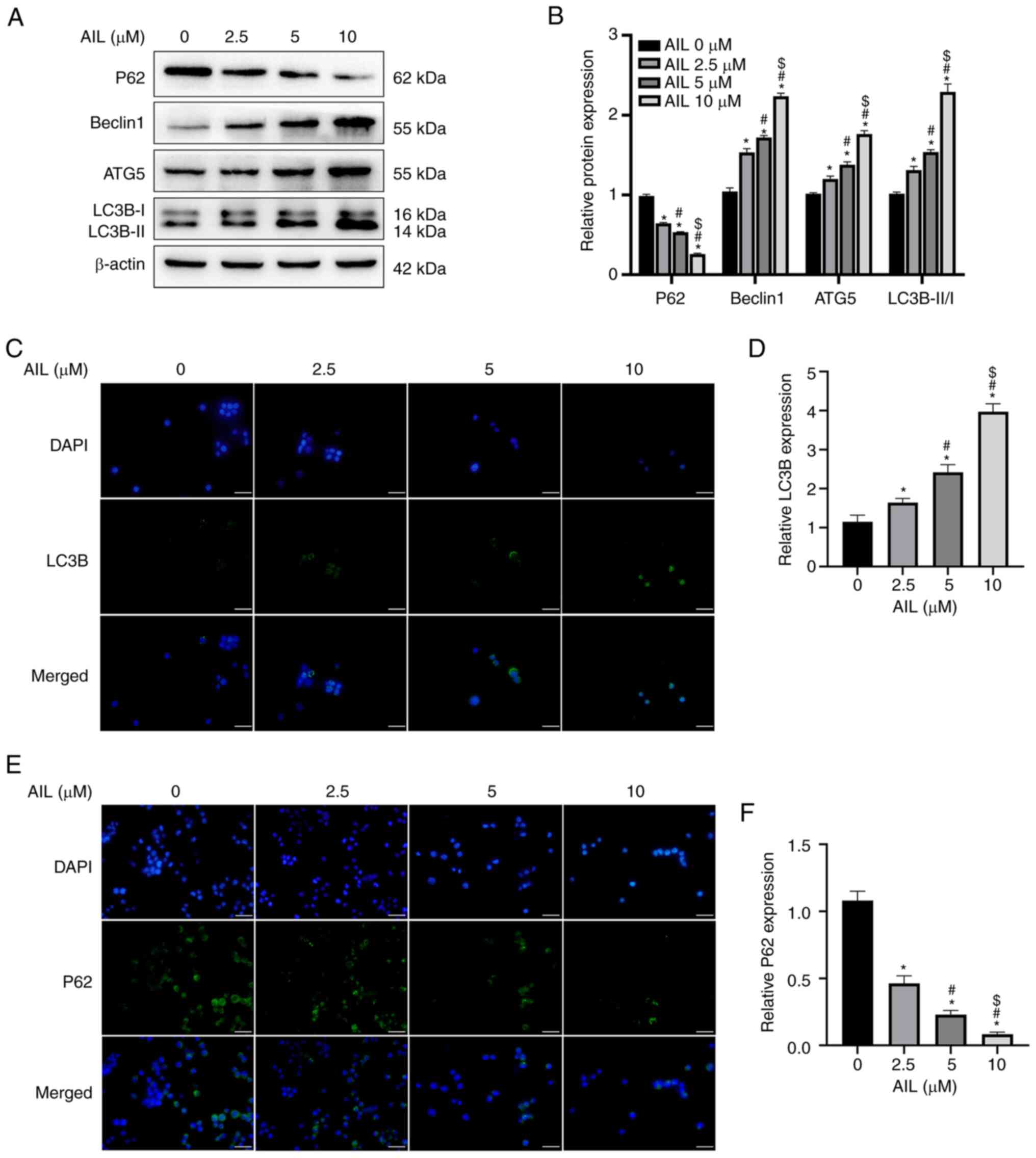
AIL induces autophagy in NSCLC Lewis
cells
The potential of AIL to regulate autophagy was
assessed based on the expression level of several
autophagy-associated proteins in Lewis cells after treatment with
0, 2.5, 5 or 10 µM AIL for 24 h. As illustrated in Fig. 2A and B, the protein expression levels of
Beclin1 (F=456.014; P<0.001), ATG5 (F=247.187; P<0.001) and
LC3B (F=152.740; P<0.001) increased, while the expression of P62
(F=1234.348; P<0.001) decreased with an increase in AIL
concentration. The immunofluorescence results were consistent with
the findings of western blot experiments in which an increase in
AIL concentration increased the green fluorescence intensity of
LC3B (F=152.179; P<0.001; Fig.
2C and D) but decreased the
green fluorescence intensity of P62 (F=255.494; P<0.001;
Fig. 2E and F).
AIL induces ferroptosis in NSCLC Lewis
cells
ROS accumulation is one of the hallmarks of
ferroptosis; therefore, ROS levels in Lewis cells treated with a
gradient concentration of AIL were assessed using the ROS-detecting
fluorescent dye DCFH-DA. The results obtained by fluorescence
microscopy (F=182.200; P<0.001; Fig. 3A and B) and a fluorescent microplate reader
(F=15.863; P<0.001; Fig. 3C)
revealed that the green fluorescence intensity increased with an
increase in AIL concentrations.
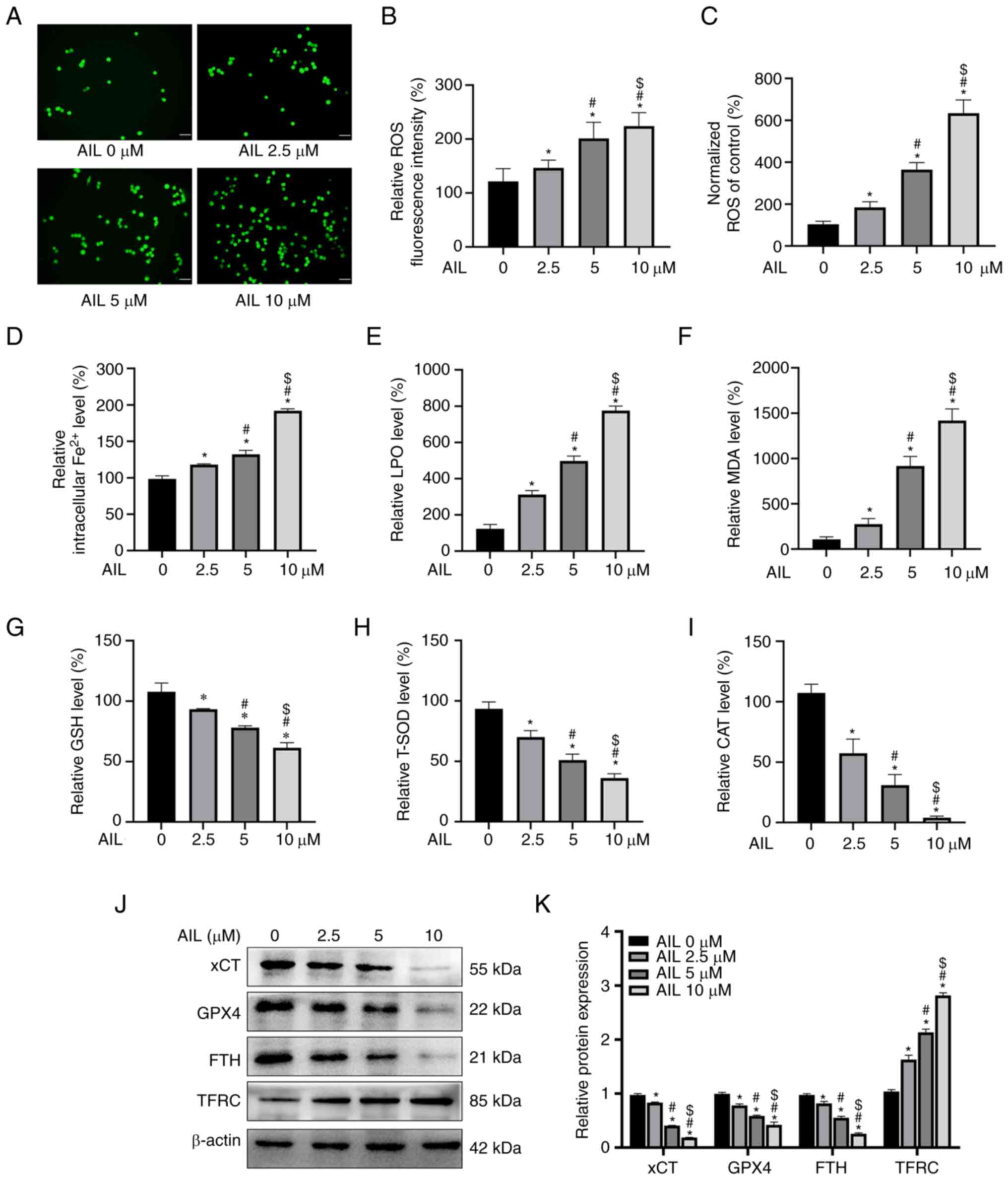 | Figure 3AIL induces ferroptosis in Lewis
cells. (A) Relative fluorescence intensity of ROS in Lewis cells
treated by AIL under a fluorescence microscope. Scale bar, 50 µm.
(B) Semi-quantitative analysis of the results from the fluorescence
intensity. (C) Relative fluorescence levels of ROS in Lewis cells
treated by AIL, as detected by fluorescent enzyme label. (D-I)
Colorimetry detection of the levels of Fe2+, LPO, MDA,
GSH, T-SOD and CAT in Lewis cells treated with gradient
concentrations of AIL. (J) Western blot analysis of
ferroptosis-associated proteins xCT, GPX4, FTH and TFRC in Lewis
cells treated with different concentrations of AIL. (K)
Semi-quantitative analysis of the results from the western blot
assay. *P<0.05 vs. AIL 0 µM group;
#P<0.05 vs. AIL 2.5 µM group and
$P<0.05 vs. AIL 5 µM group. AIL, ailanthone; ROS,
reactive oxygen species; LPO, lipid peroxidation; MDA,
malondialdehyde; GSH, glutathione; T-SOD, total superoxide
dismutase; CAT, catalase; GPX4, glutathione peroxidase 4; FTH,
ferritin heavy chain; TFRC, transferrin receptor. |
Iron overload and redox imbalance are the main
characteristics of ferroptosis. In this view, the intracellular
Fe2+ level was detected using an iron assay kit. As
demonstrated in Fig . 3D, the
intracellular Fe2+ content increased (F=353.507;
P<0.001) with an increase in AIL concentration. In addition, the
effect of AIL on ferroptosis was investigated by assessing the
intracellular redox state using reagent kits. The results revealed
that the concentrations of LPO (F=380.356; P<0.001; Fig. 3E) and MDA (F=230.938; P<0.001;
Fig. 3F) increased, while GSH
(F=67.112; P<0.001; Fig. 3G),
T-SOD (F=74.961; P<0.001; Fig.
3H) and CAT (F=88.188; P<0.001; Fig. 3I) decreased with an increase in AIL
concentration.
In addition, the protein levels of four
ferroptosis-associated genes, xCT, GPX4, FTH and TFRC, were
evaluated. Western blot assay results revealed that AIL decreased
the protein expression levels of xCT (F=2055.887; P<0.001), GPX4
(F=160.192; P<0.001) and FTH (F=402.311; P<0.001) while
increasing the protein expression level of TFRC (F=484.205;
P<0.001) in Lewis cells in a dose-dependent manner (Fig. 3J and K). These results demonstrated that AIL
induces ferroptosis in the NSCLC Lewis cells.
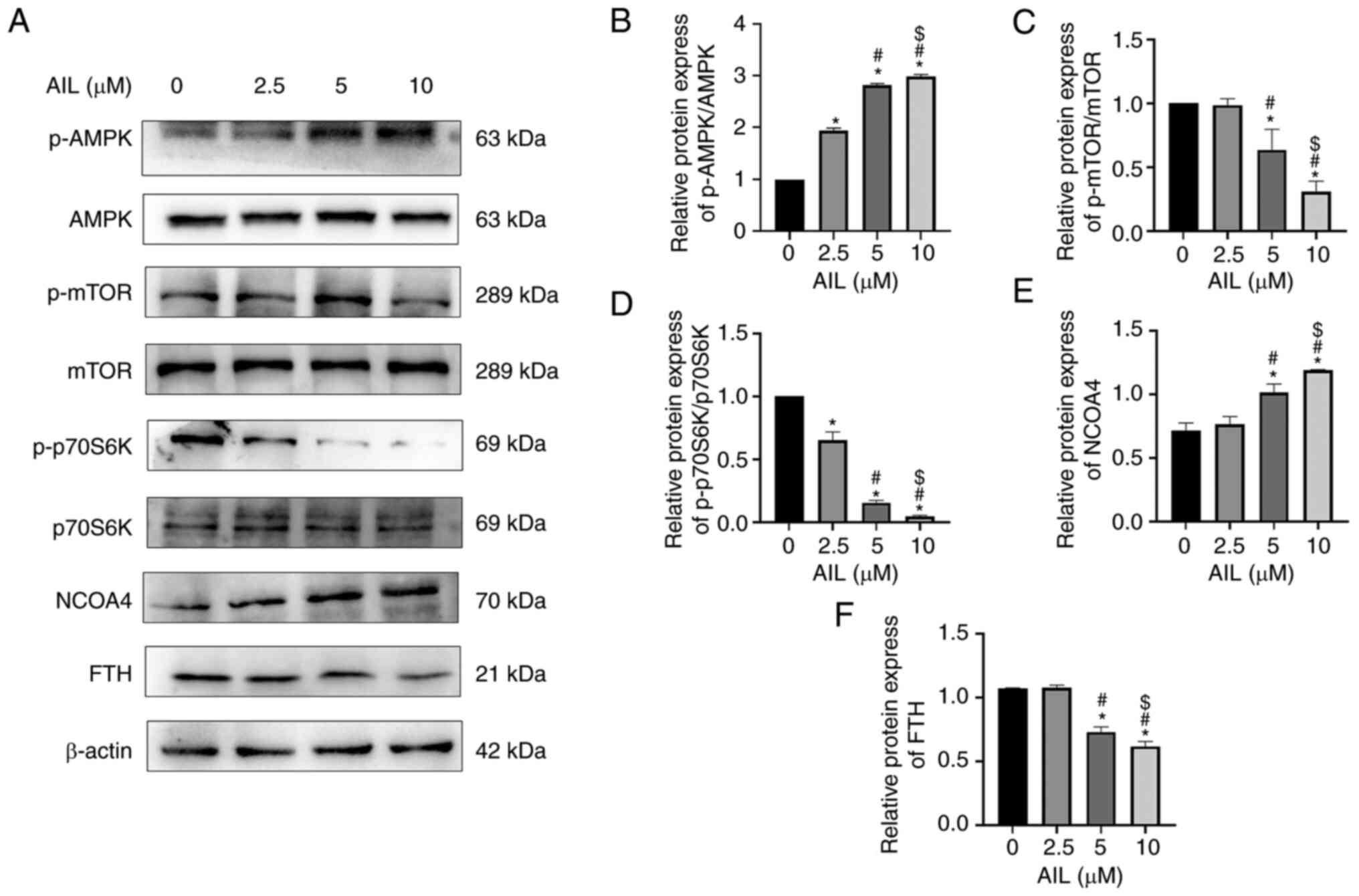
AIL regulates the AMPK/mTOR/p70S6K
signaling pathway
The mechanism by which AIL regulates autophagy and
ferroptosis was also investigated by evaluating the key protein of
the AMPK/mTOR/p70S6K signaling pathway in Lewis cells treated with
a gradient concentration of AIL for 24 h. The results showed that
AIL significantly increased p-AMPK levels and p-AMPK/AMPK ratios
(F=1199.000; P<0.001; Fig. 4A
and B) while decreasing the levels
of p-mTOR and p-mTOR/mTOR ratios (F=61.570; P<0.001; Fig. 4A and C), as well as p-p70S6K and
p-p70S6K/p70S6K ratios (F=680.200; P<0.001; Fig. 4A and D). Meanwhile, AIL upregulated the pathway
downstream protein NCOA4 (F=48.170; P<0.001; Fig. 4A and E), whereas it downregulated FTH protein
(F=181.700; P<0.001; Fig. 4A
and F).
Discussion
Recent research indicated that an increasing number
of Chinese natural medicines have a remarkable antitumor effect
while also having fewer side effects (10). In this view, a combination of
Chinese natural medicines and antitumor drugs has been used in
clinical traditional Chinese medicine antitumor treatment. The
present findings demonstrated that AIL significantly suppressed
cell proliferation in a dose- and time-dependent way. Indeed,
previous research highlighted the anticancer activity of AIL in
lung cancer. Ni et al (20)
found that AIL inhibited cell growth and colony formation in NSCLC
cell lines, while also delaying tumor growth and prolonging overall
survival in subcutaneous and orthotopic xenograft lung tumor mouse
models.
Chen et al (21) demonstrated that AIL therapy either
significantly up- or downregulated the majority of long non-coding
RNAs (lncRNAs) in NSCLC cells. Additionally, AIL-induced
alterations in lncRNA expression were essential for the occurrence
and metastasis of lung cancer. While these findings revealed that
AIL has potent antitumor effects, the underlying mechanism warrants
further investigation. As such, current research is focused on
regulating autophagy and ferroptosis, the two forms of programmed
cell death, in cancer patients (10,20).
Wei et al (22) identified
lower expression of fat atypical cadherin 4 (FAT4) in colorectal
cancer tissues than in non-cancerous tissues, and FAT4 could
decrease cell proliferation, migration and invasion in vitro
by promoting autophagy via the PI3K/AKT/GSK-3β and PI3K/AKT/mTOR
signaling pathways. Zheng et al (23) found that tetrazole blocked lung
cancer cell proliferation by over-activating autophagy, which in
turn deactivated ERK1/2 signaling while promoting mTOR signaling.
Similarly, in the present study it was found that AIL exerted an
anti-lung cancer effect by activating autophagy. Expression of
autophagy-associated protein in Lewis cells treated with a
concentration gradient of AIL was assessed using western blot and
immunofluorescence assays. The two detection methods yielded
consistent results, with an increase in the expression levels of
Beclin1, ATG5 and LC3B and a decrease in P62 protein expression,
and these effects were dose-dependent. Various proteins within the
cell have been implicated in autophagy regulation. Beclin1 is a
crucial autophagy regulator, initiating autophagosome formation by
recruiting other autophagy proteins to pre-autophagosomal
structures (24). Autophagy
activation induces the conversion of LC3-I into LC3-II, which then
associates with autophagic vesicles. Therefore, LC3-II expression
and LC3-I transformation to LC3-II are useful indicators for
intracellular autophagy assessment (25). Previous evidence indicated that
>30 ATGs regulate autophagosome formation, with ATG5 being a
central molecule in autophagy elongation (26). Besides, the level of P62 can be
utilized as an autophagy indicator because P62 is integrated into
mature autophagy and degraded by autophagy. Thus, P62 expression is
negatively correlated with autophagy (27). These data suggested that targeting
autophagy in cancer may be a promising therapeutic strategy. On one
hand, autophagy inducers can prevent the accumulation of damaged
proteins and organelles, causing genomic stability and tumor
suppression; on the other hand, autophagy inhibitors can restore
chemosensitivity and enhance tumor cell death by decreasing stress
tolerance and energy production (28).
Previous research has identified ferroptosis as a
new form of cell death caused by iron accumulation in the cell and
lipid peroxidation (29). Given
its vital role in cancer, ferroptosis therapy has become a viable
cancer target. Hu et al (30) reported that SLC7A11, a critical
ferroptosis regulator, was upregulated in lung adenocarcinoma
patients with KRAS-mutant and was positively correlated with tumor
progression. In another investigation, SLC7A11 inhibitors
significantly decreased cystine uptake and glutathione synthesis in
cells, which increased oxidative stress- and ER stress-induced cell
death in KRAS-mutant cells. ROS plays a crucial role in the
ferroptosis mechanism by inducing lipid peroxidation, which
consequently results in ferroptosis (31). The present study employed the
fluorescent probe DCFH-DA to assess intracellular ROS levels, and
the fluorescence intensity was evaluated using a fluorescence
microscope and a fluorescence microplate reader. The results
revealed higher levels of ROS in Lewis cells treated with different
concentrations of AIL. Of note, it was found that ROS accumulation
and elevated Fe2+ levels induced ferroptosis, and that
the level of Fe2+ increased with an increase in AIL
concentration. Intracellular LPO and MDA are well-known indicators
of lipid peroxidation, whereas GSH, T-SOD and CAT are the most
commonly used indicators to assess antioxidant potential. In this
view, the potential occurrence of ferroptosis in AIL-treated Lewis
cells was evaluated. The results demonstrated that an increase in
AIL concentration elevated the levels of LPO and MDA while
decreasing the levels of GSH, T-SOD and CAT, indicating that AIL
increases LPO while decreasing antioxidant enzyme activities,
resulting in ferroptosis. In addition, it was identified that AIL
treatment decreased the protein expression levels of xCT, GPX4 and
FTH but increased the expression of TFRC in Lewis cells. xCT
promotes GPX4 activity and lipid oxide metabolism by mediating
glutamate export in exchange for cystine (32), ultimately leading to ferroptosis
suppression. FTH deficiency and TFRC upregulation result in
intracellular iron accumulation, which causes ferroptosis in cancer
cells (33). These results
demonstrated that AIL increases both lipid peroxidation and ferrous
iron accumulation, resulting in ferroptosis.
The AMPK/mTOR/p70S6K signaling pathway is a
classical autophagy signaling pathway implicated in ferroptosis. Du
et al (19) found that
dihydroartemisinin increased autophagy by regulating the
AMPK/mTOR/p70S6k pathway, eventually causing ferroptosis in acute
myeloid leukemia. Chen et al (34) demonstrated that amentoflavone
inhibited cell proliferation and accelerated cell death by
activating autophagy-dependent ferroptosis via the AMPK/mTOR
pathway in human glioma. In the present investigation, the
expression of the crucial proteins in the AMPK/mTOR/p70S6K
signaling pathway was detected after treatment with different AIL
concentrations. The results revealed that an increase in AIL
concentration increased the pAMPK/AMPK ratio but decreased the
ratios of pmTOR/mTOR and p-p70S6K/p70S6K, suggesting that AIL
regulates the AMPK/mTOR/p70S6K pathway. Protein phosphorylation
denotes signal activation. The upstream adenosine triphosphate and
protein kinase drive protein phosphorylation, changing the
conformation of receptor protein, which in turn interacts with
downstream signaling molecules, initiating a series of cascade
reactions downstream of the signal transduction. Generally, the
higher the phosphorylation, the stronger the signal activation. In
western blot detection, only phosphorylated proteins are considered
activated proteins; however, the effect of total protein change
cannot be disregarded if only phosphorylated proteins are detected.
Hence, the phosphorylated proteins/total protein ratio reflects a
more accurate result. A high pAMPK/AMPK ratio is the classical
signal of autophagy activation (35). Increased pAMPK/AMPK ratio can
decrease pmTOR/mTOR ratio and p-p70S6K/p70S6K ratio, upregulating
the expression of autophagy-associated proteins LC3II/I ratio, ATG5
and Beclin-1, and promoting P62 protein degradation (36-39).
Meanwhile, it was demonstrated that the ferritinophagy marker NCOA4
was upregulated while the ferroptosis-associated protein, FTH, was
downregulated, indicating that AIL also influences the expression
of ferroptosis-associated proteins downstream of the
AMPK/mTOR/p70S6K pathway.
In conclusion, the present study demonstrated that
AIL inhibits proliferation and induces autophagy and ferroptosis in
NSCLC Lewis cells by potentially regulating the AMPK/mTOR/p70S6K
signaling pathway. However, the link between autophagy and
ferroptosis, and whether AIL has an inhibitory effect on NSCLC
in vivo warrants further investigation.
Supplementary Material
Cell viability of TC-1 cells after
treatment with a gradient concentration of AIL at 12 h
(IC50=512.047±3.502 μM), 24 h
(IC50=168.551±3.502 μM), 36 h
(IC50=122.912±4.456 μM), and 48 h
(IC50=80.945±4.283 μM) by MTT assay. AIL,
ailanthone.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant no. 81703001), the Natural
Science Foundation of Hebei (grant no. H2021406021), the Hebei
Medical Science Research Project (grant nos. 20210247 and 20221335)
and the Hebei Government-Funded Clinical Medical Outstanding
Talents Project, Chengde Medical University Scientific Research
Major Projects (grant no. KY2020005).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HY and XZ conceived and designed the experiments. YL
and XW visualized data and performed the experiments. ZZ, HX and YB
contributed new reagents or analytical tools and designed the
methodology. FL, QC and XB analyzed and interpreted the data. LZ
and LL contributed to study conception and wrote and reviewed the
manuscript. YB and XB confirm the authenticity of all the raw data.
All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sung H, Ferlay J and Siegel RL: Global
cancer statistics 2020: GLOBOCAN estimates of incidence and
mortality worldwide for 36 cancers in 185 countries. CA Cancer J
Clin. 71:209–249. 2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Reck M and Rabe KF: Precision diagnosis
and treatment for advanced non-small-cell lung cancer. N Engl J
Med. 377:849–861. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Zhang J, Späth SS and Marjani SL:
Characterization of cancer genomic heterogeneity by next-generation
sequencing advances precision medicine in cancer treatment. Precis
Clin Med. 1:29–48. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Yan L, Bao Q, Yang S, Yang M and Mao C:
Bionanoparticles in cancer imaging, diagnosis, and treatment. View
20200027, 2022. https://doi.org/10.1002/VIW.20200027.
|
|
5
|
Yin X, Yang J, Zhang M, Wang X, Xu W,
Price CH, Huang L, Liu W, Su H, Wang W, et al: Serum Metabolic
Fingerprints on Bowl-Shaped Submicroreactor Chip for Chemotherapy
Monitoring. ACS Nano. 16:2852–2865. 2022.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhang Y, Zhang C and Min D: Ailanthone
up-regulates miR-449a to restrain acute myeloid leukemia cells
growth, migration and invasion. ExpMol Pathol. 108:114–120.
2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
He Q, Xiao H and Li J: Fingerprint
analysis and pharmacological evaluation of Ailanthus altissima. Int
Mol Med. 41:3024–3032. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zhuo Z, Hu J, Yang X, Chen M, Lei X, Deng
L, Yao N, Peng Q, Chen Z, Ye W and Zhang D: Ailanthone inhibits
Huh7 cancer cell growth via cell cycle arrest and apoptosis in
vitro and in vitro. Sci Rep. 5(16185)2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Choi YJ, Park YJ, Park JY, Jeong HO, Kim
DH, Ha YM, Kim JM, Song YM, Heo HS, Yu BP, et al: Inhibitory effect
of mTOR activator MHY1485 on autophagy: suppression of lysosomal
fusion. PLoS One. 7(e43418)2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Klionsky DJ, Abdel-Aziz AK, Abdelfatah S,
Abdellatif M, Abdoli A, Abel S, Abeliovich H, Abildgaard MH, Abudu
YP, Acevedo-Arozena A, et al: Guidelines for the use and
interpretation of assays for monitoring autophagy (4th edition).
Autophagy. 17:1–382. 2021.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Ye J, Zhang R, Wu F, Zhai L, Wang K, Xiao
M, Xie T and Sui X: Non-apoptotic cell death in malignant tumor
cells and natural compounds. Cancer Lett. 420:210–227.
2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Liu Y, Yang Y, Ye YC, Shi QF, Chai K,
Tashiro S, Onodera S and Ikejima T: Activation of erk-p53 and
erk-mediated phosphorylation of Bcl-2 are involved in autophagic
cell death induced by the c-met inhibitor su11274 in human lung
cancer a549 cells. J Pharmacol Sci. 118:423–432. 2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Tang D, Kang R, Berghe TV, Vandenabeele P
and Kroemer G: The molecular machinery of regulated cell death.
Cell Res. 29:347–364. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Guo J, Xu B, Han Q, Zhou H, Xia Y, Gong C,
Dai X, Li Z and Wu G: Ferroptosis: A novel anti-tumor action for
cisplatin. Cancer Res Treat. 50:445–460. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Bursch W, Ellinger A, Gerner C, Fröhwein U
and Schulte-Hermann R: Programmed cell death (PCD), Apoptosis,
autophagic PCD, or others? Ann N Y Acad Sci. 926:1–12.
2000.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Hou W, Xie Y, Song X, Sun X, Lotze MT, Zeh
HJ Ⅲ, Kang R and Tang D: Autophagy promotes ferroptosis by
degradation of ferritin. Autophagy. 12:1425–1428. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Carling D: The AMP-activated protein
kinase cascade-a unifying system for energy control. Trends Biochem
Sci. 29:18–24. 2004.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kandula N, Kumar S, Mandlem VKK,
Siddabathuni A, Singh S and Kosuru R: Role of AMPK in myocardial
ischemia-reperfusion injury-induced cell death in the presence and
absence of diabetes. Oxid Med Cell Longev.
2022(7346699)2022.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Du J, Wang T, Li Y, Zhou Y, Wang X, Yu X,
Ren X, An Y, Wu Y, Sun W, et al: DHA inhibits proliferation and
induces ferroptosis of leukemia cells through autophagy-dependent
degradation of ferritin. Free Radic Biol Med. 131:356–369.
2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ni Z, Yao C, Zhu X, Gong C, Xu Z, Wang L,
Li S, Zou C and Zhu S: Ailanthone inhibits non-small cell lung
cancer cell growth through repressing DNA replication via
downregulating RPA1. Br J Cancer. 117:1621–1630. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Chen L, Wu C, Wang H, Chen S, Ma D, Tao Y,
Wang X, Luan Y, Wang T, Shi Y, et al: Analysis of long noncoding
RNAs in aila-induced non-small cell lung cancer inhibition. Front
Oncol. 11(652567)2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wei R, Xiao Y, Song Y, Yuan H, Luo J and
Xu W: FAT4 regulates the EMT and autophagy in colorectal cancer
cells in part via the PI3K-AKT signaling axis. J Exp Clin Cancer
Res. 38(112)2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zheng L, Zhang J, Fan J, He Y, Zhan T,
Rong L, Yuan M and Zhang H: Lung cancer growth inhibition and
autophagy activation by tetrazole via ERK1/2 up-regulation and
mTOR/p70S6K signaling down-regulation. Acta Biochim Pol.
69:139–145. 2022.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Kang R, Zeh HJ, Lotze MT and Tang D: The
Beclin 1 network regulates autophagy and apoptosis. Cell Death
Differ. 18:571–580. 2011.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Runwal G, Stamatakou E, Siddiqi FH, Puri
C, Zhu Y and Rubinsztein DC: LC3-positive structures are prominent
in autophagy-deficient cells. Sci Rep. 9(10147)2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Mizushima N, Yoshimori T and Ohsumi Y: The
role of Atg proteins in autophagosome formation. Annu Rev Cell Dev
Biol. 27:107–132. 2011.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Turco E, Witt M, Abert C, Bock-Bierbaum T,
Su MY, Trapannone R, Sztacho M, Danieli A, Shi X, Zaffagnini G, et
al: FIP200 claw domain binding to p62 promotes autophagosome
formation at ubiquitin condensates. Mol Cell. 74:330–346.e11.
2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Yang ZJ, Chee CE, Huang S and Sinicrope
FA: The role of autophagy in cancer: Therapeutic implications. Mol
Cancer Ther. 10:1533–1541. 2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Hu K, Li K, Lv J, Feng J, Chen J, Wu H,
Cheng F, Jiang W, Wang J, Pei H, et al: Suppression of the
SLC7A11/glutathione axis causes synthetic lethality in Kras-mutant
lung adenocarcinoma. Clin Invest. 130:1752–1766. 2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Su Y, Zhao B, Zhou L, Zhang Z, Shen Y, Lv
H, AlQudsy LHH and Shang P: Ferroptosis, a novel pharmacological
mechanism of anti-cancer drugs. Cancer Lett. 483:127–136.
2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Christensen HN: Role of amino acid
transport and countertransport in nutrition and metabolism. Physiol
Rev. 70:43–77. 1990.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Feng H, Schorpp K, Jin J, Yozwiak CE,
Hoffstrom BG, Decker AM, Rajbhandari P, Stokes ME, Bender HG, Csuka
JM, et al: Transferrin receptor is a specific ferroptosis marker.
Cell Rep. 30:3411–3423. 2020.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Chen Y, Li N, Wang H, Wang N, Peng H, Wang
J, Li Y, Liu M, Li H, Zhang Y and Wang Z: Amentoflavone suppresses
cell proliferation and induces cell death through triggering
autophagy-dependent ferroptosis in human glioma. Life Sci.
247(117425)2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ueno T and Komatsu M: Autophagy in the
liver: functions in health and disease. Nat Rev Gastroenterol
Hepatol. 14:170–184. 2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Gu X, Li Y, Chen K, Wang X, Wang Z, Lian
H, Lin Y, Rong X, Chu M, Lin J and Guo X: Exosomes derived from
umbilical cord mesenchymal stem cells alleviate viral myocarditis
through activating AMPK/mTOR-mediated autophagy flux pathway. Cell
Mol Med. 24:7515–7530. 2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ma S, Yin J, Hao L, Liu X, Shi Q, Diao Y,
Yu G, Liu L, Chen J and Zhong J: Exosomes from human umbilical cord
mesenchymal stem cells treat corneal injury via autophagy
activation. Front Bioeng Biotechnol. 10(879192)2022.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Hou ZP, Li YP, Zhao L, Chen YX and Ruan
XZ: Lipopolysaccharide inhibits lipo phagy in HepG2 cells via
activating mTOR pathway. Sheng Li Xue Bao. 73:813–820.
2021.PubMed/NCBI(In Chinese).
|
|
39
|
Pinto AP, da Rocha AL, Cabrera EMB,
Marafon BB, Kohama EB, Rovina RL, Simabuco FM, Bueno Junior CR, de
Moura LP, Pauli JR, et al: Role of interleukin-6 in inhibiting
hepatic autophagy markers in exercised mice. Cytokine.
130(155085)2020.PubMed/NCBI View Article : Google Scholar
|