Introduction
Keshan disease (KD) is an endemic cardiomyopathy
occurring only in China, involving 327 counties of 16 provinces.
Chronic KD (CKD) has similar clinical characteristics to dilated
cardiomyopathy (DCM), such as acute or chronic episodes of heart
disorder characterized by cardiogenic shock, arrhythmia and
congestive heart failure, with cardiomegaly, as well as multifocal
myocardial necrosis and fibrosis under the electron microscope
(1). A differential diagnosis for the
two diseases in clinical practice is extremely difficult. To
further distinguish, the main characteristics of DCM are increased
heart weight and cardiac hypertrophy, while KD is characterized by
severe myocardial degeneration, evident necrosis and fibrosis, and
even clear geographic characteristics. Previous studies have
identified that the two types of disease were significantly
different in ventricular cavity expansion and ventricular volume by
summarizing the image and ultrasound information of DCM and KD
patients (2). In recent years, certain
studies have proposed that ATM and NFATC2 may be
considered significant differential genetic diagnoses in the two
similar diseases through comparing the KD and DCM patient
peripheral blood mononuclear cells using genome-wide expression
microarray technology (3).
Arachidonic acid (AA) is oxidized through three
major metabolic pathways: Cyclooxygenase (COX), lipoxygenase (LOX)
and cytochrome P450 (CYPs) (4).
Previously, studies have focused on the CYP450 system of AA;
epoxidation and hydroxylation of AA are catalyzed by cytochrome
P450s (CYPs) in heart disease (5).
According to the CYP metabolic pathway followed by AA, the major
metabolites are either epoxyeicosatrienoic acids (EETs) or
20-hydroxyeicosatetraenoic acids (20-HETE), which are primarily
vasodilators or vasoconstrictors, respectively (6). In addition, it has been increasingly
recognized that EETs and 20-HETE have crucial and opposing roles in
the development of cardiac disease. Certain studies have reported
the possible changes in expression levels of the main human CYP450
monooxygenase (2E1 isoform) at DCM progression at the end stage of
heart failure using western blot analysis (7). While for KD, a number of previous studies
have explored the mechanisms of the former two pathways (COX and
LOX pathways of AA), which were abnormal in KD, little evidence is
available regarding the CYP450 pathway in KD. Therefore, a
comparison in the metabolism of AA by the CYP450 enzyme between CKD
and DCM is noteworthy, and may provide more functional information
for the pathogenesis of KD.
In the past several decades, a number of studies
have explored the cause of KD, and its main etiology and
pathogenesis is known to be selenium deficiency. At present, KD is
widely considered as multifactorial environment-gene interaction
complex diseases. KD has been suggested as a ‘mitochondrial
cardiomyopathy’ endemic in China (8).
Our previous study compared the mitochondrial-related gene
expression profiles of peripheral blood mononuclear cells derived
from KD patients and normal controls by mitochondria-focused cDNA
microarray technology. The results indicated that the
enzyme-related genes CYP1A1 and CYP2C19 were upregulated (ratios
≥2.0) (9), and these genes belong to
the cytochrome P450 isoforms, whose metabolites are biologically
active and have critical roles in the maintenance of essential body
functions. Various CYP isoenzymes have been confirmed to have a
role in the metabolism of xenobiotics and endogenous compounds. The
model endogenous substrate of CYP is AA (10).
In the present study, the metabolism of AA by the
CYP450 enzyme was investigated in CKD patients, DCM patients and
health controls by reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) and enzyme-linked immunosorbent assay
(ELISA) to examine the gene expression changes and biochemical
parameters associated with the CYP-AA metabolism. The study may aid
in the understanding of the pathogenesis of KD and distinguish a
possible difference between KD and DCM.
Materials and methods
Sample collection and experiment
group
The present study was approved by the Human Ethics
Committee, Xi'an Jiaotong University (Xi'an, Shaanxi, China) and
all participants provided informed consent. A total of 67 CKD
patients were selected according to ‘The Diagnostic Standard of the
KD in China’ for KD (WS/T 210–2011), who were from areas of
Zhengning and Heshui (Gansu, China). A total of 28 DCM patients who
did not live in endemic areas were diagnosed in three
tertiary-level facilities hospitals in Xi'an in terms of the World
Health Organization/International Society of Federation of
Cardiology proposed standards (11).
Three group subjects were matched by age and gender. These patients
were selected as samples as they were not exhibiting coronary
artery disease, myocarditis, congenital heart disease, pulmonary
heart disease or other cardiovascular diseases. A total of 58
healthy controls without any chronic diseases, such as diabetes,
hypertension and any other heart disease, were collected from the
KD-affected area of Huangling (Shaanxi, China).
Peripheral venous blood (3 ml) from each subject was
collected into vacuum blood tube, and serum was obtained through
centrifugation at 2,683 × g for 8 min and stored at −80°C for
ELISA. Anticoagulant whole blood (2 ml) was acquired into a tube
with 7.5 ml RNA stabilizing solution and mixed prior to storing at
−80°C using for RNA extraction.
Total RNA extraction and cDNA
synthesis
The total RNA from blood was abstracted with
E.Z.N.A. Total RNA kit I (Omega Bio-Tek, Norcross, RA, USA)
according to the manufacturer's protocol. The concentration and
purity of total RNA was evaluated by measuring the absorbance
260/280 ratio with a Thermo Nanodrop 2000 (Thermo Fischer
Scientific, Inc., Waltham, MA, USA). Following this, RNA was
transcribed into cDNA using the PrimeScript RT Reagent kit (Takara
Biotechnology, Dalian, China). The reverse transcription system was
as follows: 0.5 µg of total RNA was added to a mix of 2.0 µl 5X
PrimeScript buffer, 0.5 µl PrimeScript RT Enzyme mix I, 0.5 µl
Oligo dT Primer, 0.5 µl Random 6-mers, finally adding Rnase-Free
dH2O to create a volume of 10 µl. The reaction mixture
was maintained at 37°C for 15 min, and heated for 85°C for 5
sec.
RT-qPCR
RT-qPCR analysis was performed in the Bio-Rad IQ5
PCR Detection system (Bio-Rad, Hercules, CA, USA) using SYBR Premix
Ex Taq™ II (Takara Biotechnology). A 25-µl reaction mixture was
placed into 0.2 ml clear strips with 8-well tubes (Crystalgen,
Commack, NY, USA), which contained 1 µl of 10 µM forward primer and
1 µl of 10 µM reverse primer (0.4 µM final concentration of each
primer), 12.5 µl of SYBR Premix Taq II, 8.5 µl of nuclease-free
water, and 2 µl of the cDNA sample. The specific primer sequences
were CYP1A1 (forward, 5′-CCT CCT CAA CCT CCT GCT AC-3′ and reverse,
5′-AAG CAA ATG GCA CAG AYG AC-3′); CYP2C19 (forward, 5′-GGT CCT TGT
GCT CTG TCT CT-3′ and reverse, 5′-CAT ATC CAT GCA GCA CCA CC-3′);
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (forward, 5′-TGC
ACC ACC AAC TGC TTA GC-3′ and reverse, 5′-GGC ATG GAC TGT GGT CAT
GAG-3′), which were designed and synthesized by AuGCT DNA-SYN
Biotechnology Corporation, Beijing, China. The thermocycling
conditions were set at 95°C for 30 sec, followed by 40 PCR cycles
of denaturation at 95°C for 5 sec, 60°C for 30 sec, and
annealing/extension at 65°C for 15 sec. GAPDH was used as an
internal standard for mRNA levels.
ELISAs
The ELISA kit is an accurate, precise sensitivity
assay, particularly suitable for large sample detection. The levels
of protein expression of CYP1A1 and CYP2C19, AA, EETs and 20-HETE
in all the serum samples were tested using commercially available
ELISA kits (R&D Systems, Inc., Minneapolis, MN, USA) in
accordance with the manufacturer's protocols. A calibration
standard curve on a semi-log scale was subsequently drawn by
collecting and analyzing the corresponding absorbencies at
different dilution extents of standards in duplicate. The Infinite
M2000 multimode microplate reader (Tecan, Männedorf, Switzerland)
was used to test the absorbance of the samples and standards. In
carrying out these assays, the serum of each patient was detected
in two replicates, which prevented from repeated freezing and
thawing of the samples.
Statistical analysis
All the experimental data were analyzed with SPSS
version 18.0 software (IBM, Corp., Armonk, NY, USA). The
gender-matched RT-qPCR sample set and ELISA sample set was
determined using χ2 test and age was compared by one-way
analysis of variance (ANOVA). The 2−ΔΔCT method was used
to calculate the relative fold-changes in gene expression
determined from the RT-qPCR result (12). ELISA data were analyzed using ANOVA,
and were tested by pairwise comparisons [least significant
difference (LSD)-t-test]. P<0.05 was considered to indicate a
statistically significant difference.
Results
Baseline data
The characteristics of the CKD and DCM patients and
the controls are shown in Table I. No
significant difference was apparent between the gender constituent
of the RT-qPCR sample set and ELISA sample among the three groups
by χ2 test (P<0.05). Age distribution in the three
groups was balanced, with no significant difference through one-way
ANOVA (P<0.05). The LSD-t-test analysis of age distribution
indicated no significant difference in any two groups (P<0.05).
These results suggest there were no significant gender and age
differences; and these characteristics were balanced and comparable
among the three groups.
 | Table I.Gender and age characteristics of
research subject. |
Table I.
Gender and age characteristics of
research subject.
| Analysis | Male, n | Female, n | Mean age ± SD,
years |
|---|
| RT-qPCR |
|
|
|
| CKD
(n=6) | 3 | 3 | 52.00±6.00 |
| DCM
(n=6) | 3 | 3 | 49.33±2.08 |
| Controls
(n=6) | 3 | 3 | 48.67±2.52 |
|
Statistical analysis |
χ2=1.706, | P=1 |
F=0.6,
P=0.579 |
| ELISA |
|
|
|
| CKD
(n=67) | 27 | 40 | 54.06±15.72 |
| DCM
(n=28) | 11 | 17 | 58.00±16.64 |
| Controls
(n=58) | 23 | 35 | 50.33±21.36 |
|
Statistical analysis |
χ2=0.825, | P=0.662 | F=0.135, P=0.876 |
Results of RT-qPCR
The concentration of total RNA was examined with the
Thermo Nanodrop 2000. The absorbance value 260/280 was 1.94–2.09,
maintained at ~2.0. Therefore, the RNA samples were of a good
purity and quality. RT-qPCR was performed to assess the mRNA levels
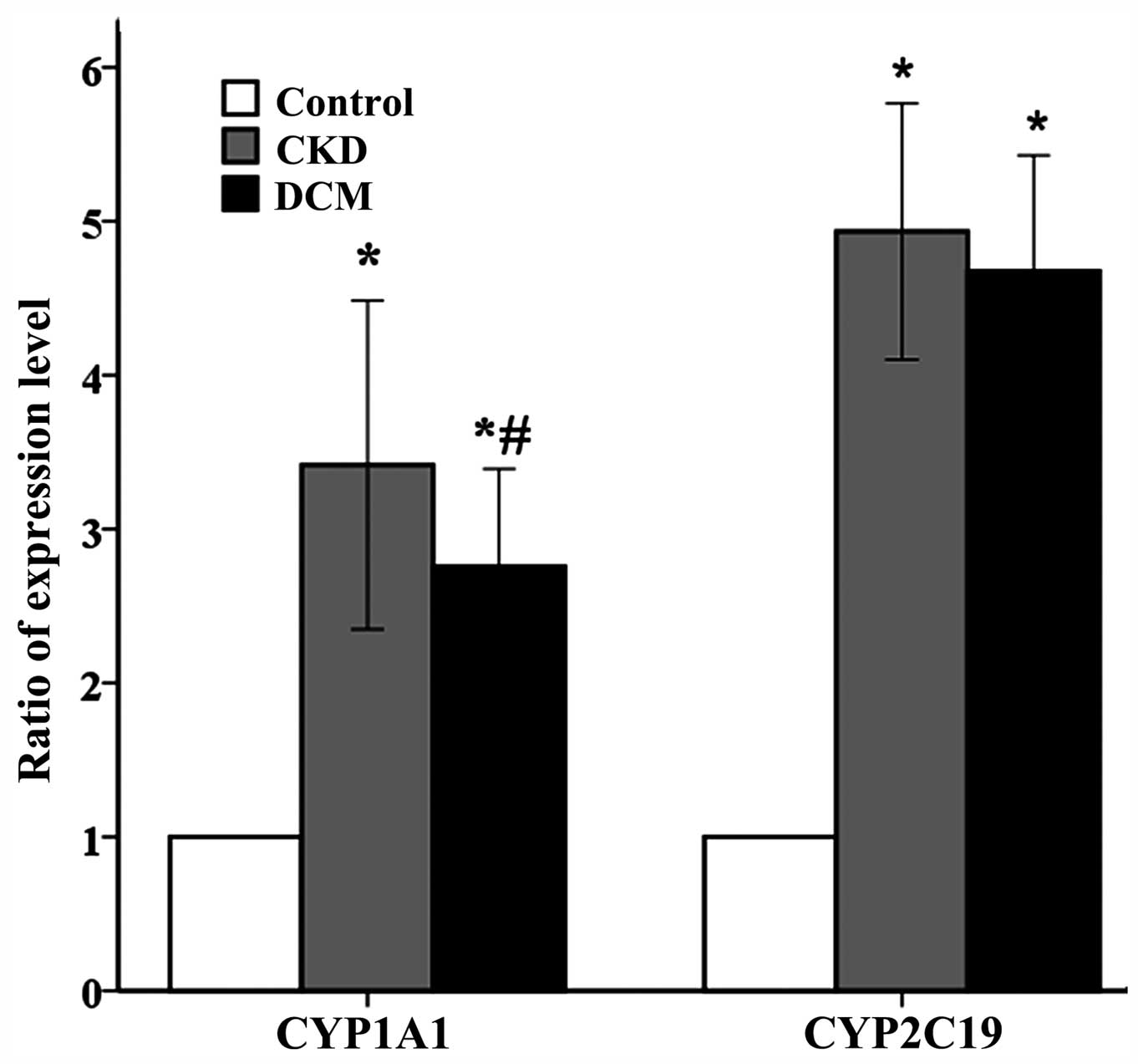
of CYP1A1 and CYP2C19. As shown in Fig.
1, multiple bar diagrams show the expression ratio of the
CYP1A1 and CYP2C19 genes. The expression levels of the two target
genes in the CKD and DCM groups were markedly enhanced compared
with the control group, which were consistent with the former
microarray experiment (5).
ELISA results
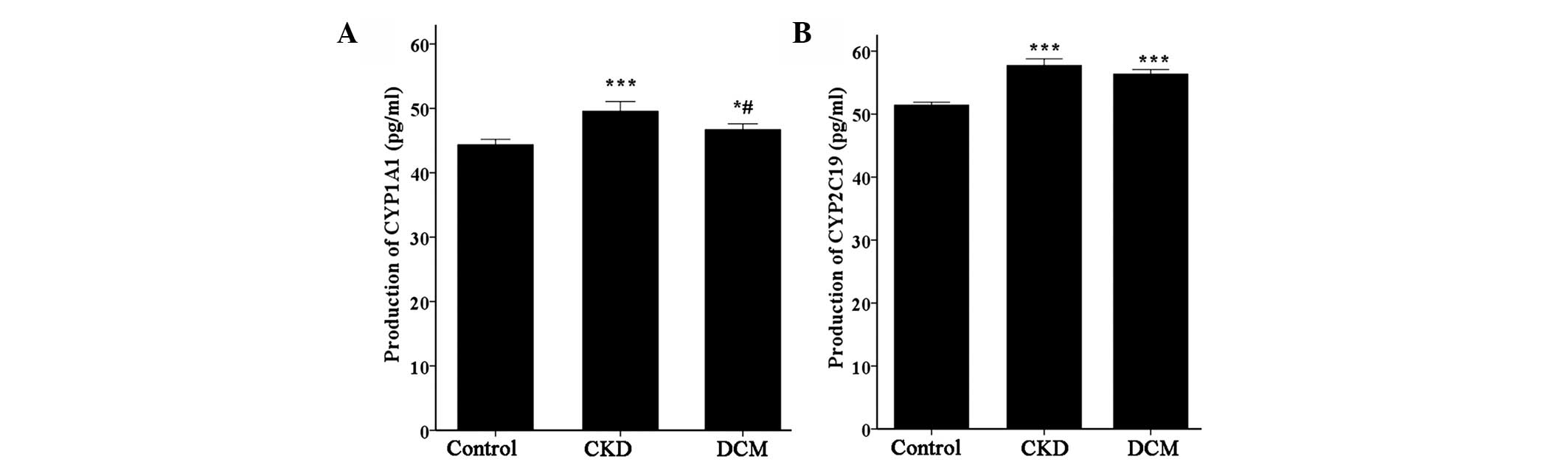
To further investigate the CYP1A1 and CYP2C19
protein expression levels and the relevant metabolites, ELISA kits
were used. Fig. 2 presents the various
expression levels of CYP1A1 and CYP2C19 protein production. The
expression level of the target proteins in the CKD and DCM patients
was significantly increased compared with the healthy controls
(P<0.001), consistent with the former immunohistochemical
results. In addition, between the CKD and DCM groups, the CYP1A1
protein exhibited a significant difference (P<0.05), whereas
there was no difference for the CYP2C19 protein (P>0.05).
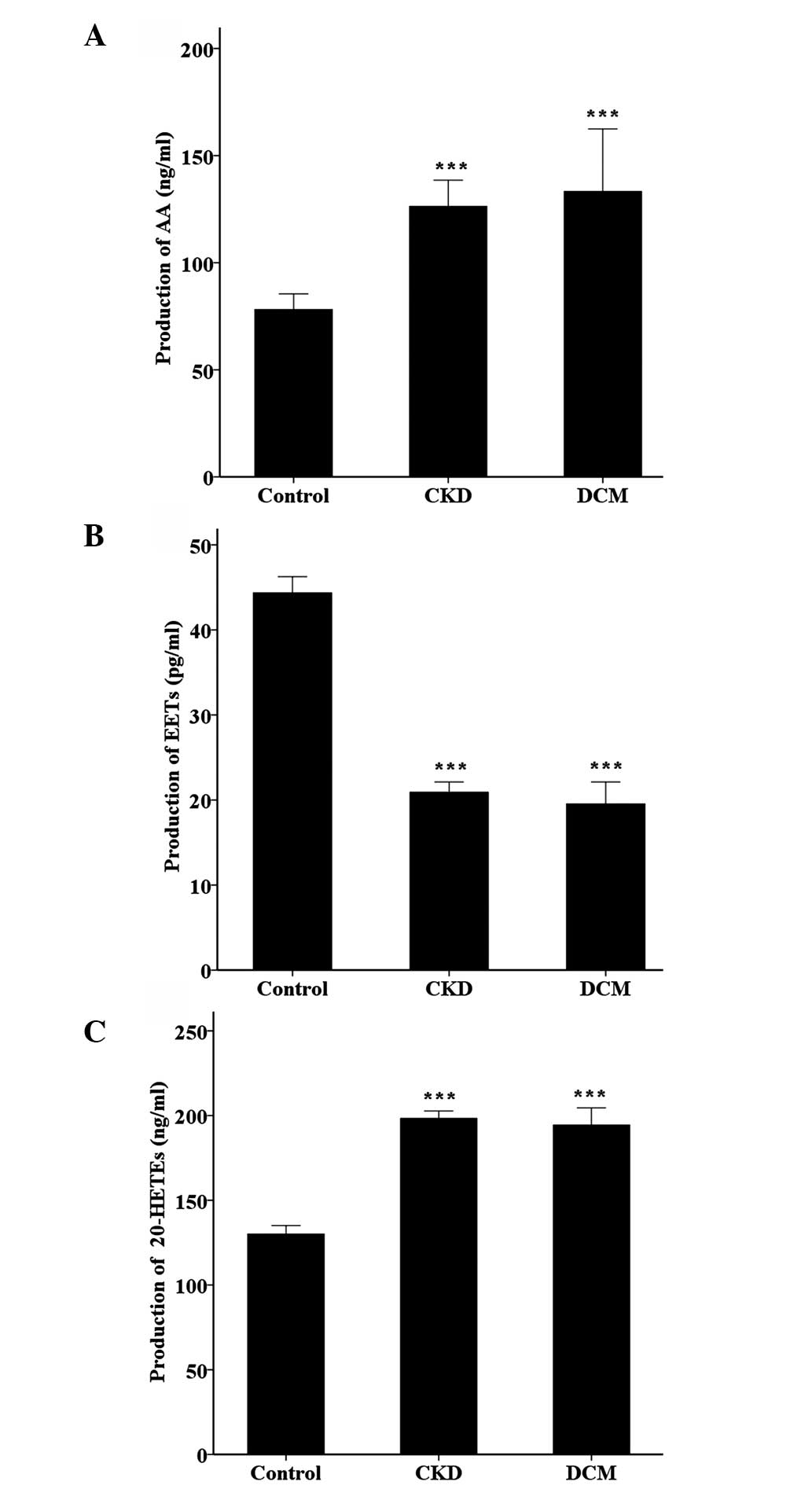
In order to investigate the associated product
expression level of the AA-CYP metabolic pathway, the
concentrations of AA, EETs and 20-HETE in the blood serum were
measured by the ELISA kits. As described in Fig. 3, the content of AA and 20-HETE had a
similar trend, as the content was significantly increased in the
CKD and DCM groups compared to the controls (P<0.001). However,
the expression level of EETs exhibited the opposite tendency, with
a maximum concentration in the healthy control, and a reduction of
the concentration in the CKD and DCM groups (P<0.001).
Discussion
The present study evaluated the possible differences
between CKD and DCM based on the upregulation of CYP1A1 and CYP2C19
genes. RT-qPCR and ELISA were applied to compare the differences in
the CYP1A1 and CYP2C19 gene expression levels and biochemical
parameters associated with CYP-AA metabolism in the CKD patients,
DCM patients and normal controls. The RT-qPCR results were
presented in Fig. 1, and the
expression levels of the two target genes in the CKD and DCM groups
were markedly enhanced compared with the control group, which were
completely consistent with previous research results. In the
healthy human heart, CYP1A1 mRNA is expressed in an extremely low
amount (5), which has a vital
physiological activity on endogenous substrates that are involved
in the metabolism of AA and tryptamine (13). CYP2C19 mainly exists in the hepatic
microsome, which is a type of important liver microsomal enzyme,
mainly involved in a wide variety of drugs' metabolism and
arachidonic acid epoxidation (13,14).
Previous studies have reported that the expression levels of CYP
genes 1A1 and 2C19 in the heart were predominant in the right
ventricle (15). In CKD and DCM
patient blood, mRNA levels for the CYP1A1 and CYP2C19 were higher
in comparison to the healthy control, indicating that certain
changes are caused by genetics in the metabolic pathways mainly
associated with the cardiac structure and function injury, rather
than starting factors, as an approach of original cause leading to
myocardial pathological changes. There is a possible
interrelationship of cause and effect between the pathological
changes of the heart and AA-dependent CYP metabolic pathways.
Several years ago, a number of studies manifested
that low selenium and vitamin E could increase the AA produced by
enhancing the activity of PLA2 in the initial stage of AA
metabolism, thereby affecting the final products level of AA
metabolites. As expected, the present study also identified that
the content of AA in the two patient groups was notably higher
compared to the normal control group. KD is mainly distributed in
the selenium deficient areas of China from southeast to northwest,
which may result in a higher AA concentration in KD. In addition,
lipid peroxide levels were enhanced in dietary vitamin E deficiency
in the heart (16). AA is one of the
diverse free fatty acids, that not only participates in myocardium
energy supply, but AA and its CYP-derived metabolites may also act
as secondary messenger molecules for protein kinases, including
protein kinase C, mitogen-activated protein kinase, protein kinase
A, Erk1/2, and Akt (13,17,18). AA is
synthesized from linoleic acid, and previous studies have reported
that dietary linoleic acid supplementation was beneficial to DCM
patients. This may explain the reason for a higher AA content in
DCM patients. However, more linoleic acid content may aggravate
myocardial ischemia and debase the cardiac function. Thus,
excessive AA may lead to severe cardiac function reduction of DCM.
There was no difference between the content of AA in the two
patient groups. Therefore, a higher AA content may have a certain
protective effect on the heart, but may cause side effects. This
effect has no association with the geographical environment
factors.
There is evidence that CYP450 modulates
contractility of cardiomyocytes through metabolism of AA (14). CYPs are known to metabolize AA to HETEs
and EETs through ω-hydroxylases and epoxidation, respectively
(19). Biological activities of EETs
and the opposing actions of 20-HETE within the heart are now well
established (6). EETs can be applied
to a variety of ion channels of cardiovascular cells, and protect
myocardial cells by regulating the expression of genes and proteins
associated with apoptosis (20).
20-HETE has detrimental effects, particularly in the heart
ischemia. A previous study reported that inhibition of 20-HETE can
reduce the infarct size in canines (21). In the present results, the content of
EETs in CKD and DCM was clearly lower compared to the healthy
control. Therefore, EETs may have a key role during the development
and/or progression of KD and DCM. CYP2C are the predominant
epoxygenase isoforms involved in EETs formation, and they are
abundantly expressed in the endothelium, myocardium and kidney in
humans (20). As a consequence, the
CYP2C19 gene was upregulated and the frequent protein expression
may be a protective compensation reaction. According to certain
studies, CYP1A1 metabolizes AA to different regioisomers of HETEs
(22). Additionally, previous studies
have shown that the induction of CYP1A1 in the hypertrophied hearts
caused a significant increase in the formation of 20-HETE (23). In the present study, the experimental
data also documented that the concentration of 20-HETE was
significantly increased compared to the healthy control subjects.
Therefore, an increase in the expression level of CYP1A1 may
exacerbate myocardial injury. The findings summarized above
demonstrate that AA metabolites are disordered in endemic DCM and
idiopathic DCM. The purpose of these changes may adapt to the
pathological and physiological of the heart through regulating
content, structure and stability of cell membrane lipid, as well as
AA metabolites. A wide cardioprotective role of fatty acids has
been studied thus far, particularly CYP-mediated AA metabolites may
have the more complex role in KD and DCM.
In conclusion, there is a strong correlation between
P450-mediated AA metabolites and the pathogenesis of dilated
cardiomyopathy. The overall balance of CYP genes changes have
resulted in a higher production of 20-HETE and lower production of
EETs in the KD and DCM hearts. Speculation regarding the
therapeutic implications of automatically selective expression of
CYP450 enzymes should be made with caution. Further studies are
required to investigate the mechanisms by which KD modulates P450
gene expression.
Acknowledgements
The authors thank all the research subjects who
submitted peripheral blood. The present study was supported by the
National Natural Scientific Foundation of China (grant no.
81273008) and the National Natural Scientific Foundation of China
(grant no. 30872192).
References
|
1
|
Lei C, Niu X, Wei J, Zhu J and Zhu Y:
Interaction of glutathione peroxidase-1 and selenium in endemic
dilated cardiomyopathy. Clin Chim Acta. 399:102–108. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hwang JJ, Allen PD, Tseng GC, Lam CW,
Fananapazir L, Dzau VJ and Liew CC: Microarray gene expression
profiles in dilated and hypertrophic cardiomyopathic end-stage
heart failure. Physiol Genomics. 10:31–44. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xiaohui S, He S and Hong TW: Comparative
analysis of gene expression profile between Keshan disease and
dilated. Experimental and Clinical Cardiology. 20:1373–1384.
2014.
|
|
4
|
Proctor KG, Falck JR and Capdevila J:
Intestinal vasodilation by epoxyeicosatrienoic acids: Arachidonic
acid metabolites produced by a cytochrome P450 monooxygenase. Circ
Res. 60:50–59. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chaudhary KR, Batchu SN and Seubert JM:
Cytochrome P450 enzymes and the heart. IUBMB Life. 61:954–960.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fer M, Dréano Y, Lucas D, Corcos L, Salaün
J, Berthou F and Amet Y: Metabolism of eicosapentaenoic and
docosahexaenoic acids by recombinant human cytochromes P450. Arch
Biochem Biophys. 471:116–125. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sidorik L, Kyyamova R, Bobyk V, Kapustian
L, Rozhko O, Vigontina O, Ryabenko D, Danko I, Maksymchuk O,
Kovalenko VN, et al: Molecular chaperone, HSP60 and cytochrome P450
2E1 co-expression in dilated cardiomyopathy. Cell Biol Int.
29:51–55. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fuyu Y: Keshan disease and mitochondrial
cardiomyopathy. Sci China C Life Sci. 49:513–518. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He SL, Tan WH, Zhang ZT, Zhang F, Qu CJ,
Lei YX, Zhu YH, Yu HJ, Xiang YZ and Guo X: Mitochondrial-related
gene expression profiles suggest an important role of PGC-1alpha in
the compensatory mechanism of endemic dilated cardiomyopathy. Exp
Cell Res. 319:2604–2616. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Elbekai RH and El-Kadi AO: Cytochrome P450
enzymes: Central players in cardiovascular health and disease.
Pharmacol Ther. 112:564–587. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Maisch B: Classification of
cardiomyopathies according to the WHO/ISFC Task Force - more
questions than answers. Med Klin (Munich). 93:199–209. 1998.(In
German). View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gottlieb RA: Cytochrome P450: Major player
in reperfusion injury. Arch Biochem Biophys. 420:262–267. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Raucy JL, Mueller L, Duan K, Allen SW,
Storm S and Lasker JM: Expression and induction of CYP2C P450
enzymes in primary cultures of human hepatocytes. J Pharmacol Exp
Ther. 302:475–482. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Thum T and Borlak J: Gene expression in
distinct regions of the heart. Lancet. 355:979–983. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Melin AM, Carbonneau MA, Thomas MJ, et al:
Dietary vitamins A and E affect differently lipid peroxidation in
rat heart and testis. J Clin Biochem Nutr. 18:19–33. 1995.
View Article : Google Scholar
|
|
17
|
Westphal C, Konkel A and Schunck WH:
CYP-eicosanoids - a new link between omega-3 fatty acids and
cardiac disease? Prostaglandins Other Lipid Mediat. 96:99–108.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zeldin DC: Epoxygenase pathways of
arachidonic acid metabolism. J Biol Chem. 276:36059–36062. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Seubert JM, Zeldin DC, Nithipatikom K and
Gross GJ: Role of epoxyeicosatrienoic acids in protecting the
myocardium following ischemia/reperfusion injury. Prostaglandins
Other Lipid Mediat. 82:50–59. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu X, Zhang XA and Wang DW: The roles of
CYP450 epoxygenases and metabolites, epoxyeicosatrienoic acids, in
cardiovascular and malignant diseases. Adv Drug Deliv Rev.
63:597–609. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gross ER, Nithipatikom K, Hsu AK, Peart
JN, Falck JR, Campbell WB and Gross GJ: Cytochrome P450
omega-hydroxylase inhibition reduces infarct size during
reperfusion via the sarcolemmal KATP channel. J Mol Cell Cardiol.
37:1245–1249. 2004.PubMed/NCBI
|
|
22
|
Zordoky BN and El-Kadi AO:
2,3,7,8-Tetrachlorodibenzo- p-dioxin and beta-naphthoflavone induce
cellular hypertrophy in H9c2 cells by an aryl hydrocarbon
receptor-dependant mechanism. Toxicol In Vitro. 24:863–871. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zordoky BN, Aboutabl ME and El-Kadi AO:
Modulation of cytochrome P450 gene expression and arachidonic acid
metabolism during isoproterenol-induced cardiac hypertrophy in
rats. Drug Metab Dispos. 36:2277–2286. 2008. View Article : Google Scholar : PubMed/NCBI
|