Introduction
The ionophore salinomycin is a polyether antibiotic
that is applied in veterinary medicine as an antiprotozoal agent
(1). Specifically, the use of
salinomycin in stockbreeding, particularly poultry, may have
harmful side effects due to its severe toxicity. The data regarding
human health hazards is poor; however, certain case reports show
life-threatening side effects (2–4). By
consuming animal products, there may be a chronic human exposure to
salinomycin. Mortier et al (5)
investigated egg samples from eight different countries in Europe
for the presence of antibiotics and 20% of all samples were
contaminated with salinomycin. Thus, the consequences on human
health potentially caused by this chronic exposure to salinomycin
appeared to warrant investigation. However, to the best of our
knowledge, there are no studies on salinomycin intoxication
following consumption of animal products. Szkudlarek-Mikh et
al (6) reported that salinomycin
inhibited the differentiation of preadipocytes into adipocytes, and
that salinomycin was the most potent inhibitor. This was not due to
apoptosis or inhibition of cell proliferation, but was closely
associated with suppression of the transcriptional activity of
adipogenesis. The latter was explained by the suppression of the
CCAAT/enhancer-binding proteins and the peroxisome
proliferator-activated receptor (6).
This result raises the question of whether chronic salinomycin
exposure interferes with the differentiation capability of human
cells, such as mesenchymal stem cells (MSC) as well. MSC are
undifferentiated cells and there are no specific markers for their
identification. According to the International Society of Cellular
Therapy, MSC have to meet the following criteria: Adherent growth
behavior, be positive for cluster of differentiation (CD) 105, CD90
and CD73 and negative for CD14, CD11b and HLAII cell surface
markers. Furthermore, MSC must have the ability to differentiate
into bone, cartilage or fat (7,8).
In our previous study, MSC were exposed to
salinomycin for 24 h. Salinomycin did not affect any essential
properties of MSC. However, dose-dependent cytotoxic effects of
salinomycin on MSC were observed. In the present study, the
following functional impairments of MSC following chronic
salinomycin exposure were examined: Alteration of cell surface
markers and differentiation capability, genotoxicity, migration,
proliferation capacity and cytoskeletal architecture.
Materials and methods
MSC isolation and salinomycin
treatment
MSC were isolated from the human bone marrow of 5
voluntary patients undergoing surgery in the Department of
Orthopedic Surgery, University of Wuerzburg. The study was approved
by the Ethics Committee of the Medical Faculty, University of
Wuerzburg (12/06; Bavaria, Germany), and informed consent was
obtained from all the individuals included. The isolation of MSC
was performed according to previous studies using Ficoll density
gradient centrifugation (30 min, 227 × g, density=1,077 g/ml;
Biochrom AG, Berlin, Germany) (9,10).
Subsequent to the collection of the cells from the interphase,
several washing steps with phosphate-buffered saline (PBS) (Roche
Diagnostics GmbH, Mannheim, Germany) containing 2% fetal calf serum
(FCS) (Linaris, Wertheim-Bettingen, Germany) were performed. The
isolated cells were resuspended in Dulbecco's modified Eagle's
medium-expansion medium (DMEM-EM) (Gibco Invitrogen, Thermo Fisher
Scientific, Karlsruhe, Germany) with 10% FCS, 1%
penicillin/streptomycin (Sigma-Aldrich, Schnelldorf, Germany).
After incubation at 37°C and 5% CO2 in DMEM-EM
overnight, the tissue culture plates were washed to remove residual
non-adherent cells. Every other day the medium was changed. Cell
morphology was investigated by inverted microscopy (Leica DMI 4000B
Inverted Microscope; Leica Microsystems, Wetzlar, Germany). MSC
were treated with 100 nM of salinomycin for 4 weeks (MSC-Sal). The
medium containing 100 nM salinomycin was changed every other day.
MSC cultivated in DMEM-EM served as the control (MSC-control).
Expression of the cell surface
marker
The cell surface profile of MSC-Sal was investigated
by flow cytometry (BD FACSCanto™; BD Biosciences, Heidelberg,
Germany) as follows: Before cells were incubated on ice with 5% FCS
for 1 h, they were first trypsinized and washed with PBS.
Subsequently, the cells were washed with PBS and incubated with
anti-CD90 (cat no. 559869), anti-CD73 (cat no. 550257), anti-CD44
(cat no. 555478), and anti-CD34 (cat no. 555820; all from BD
Biosciences). The results were compared to MSC-control.
Multi-differentiation capacity
MSC-Sal were incubated with adipogenic medium, which
consisted of DMEM-EM containing 10 µM dexamethasone
(Sigma-Aldrich), and 10 ng/ml recombinant human insulin. The
osteogenic medium contained DMEM-EM, 10-7 M dexamethasone, 10 mM
β-glycerophosophate, 50 µg/ml ascorbic acid-2-phosphate (all from
Sigma-Aldrich). Cells were incubated for 3 weeks. The medium was
changed every third day. The differentiation potential was examined
by histological staining. Oil Red O was conducted to demonstrate
the adipogenic differentiation. The von Kossa method was used to
confirm osteogenic differentiation. For gene expression analysis,
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) was conducted. In this technique, the expression of
specific marker genes for osteogenic and adipogenic differentiation
was evaluated. All the samples were analyzed in triplicate for each
patient. Total RNA was extracted using the RNAeasy Mini kit
(Qiagen, Hilden, Germany), following the manufacturer's
instructions. Subsequently, total RNA was reverse transcribed to
cDNA using the high-capacity RNA-to-cDNA Master Mix (Applied
Biosystems, Darmstadt, Germany). The RT-qPCR device (Applied
Biosystems) was used. The PCR primers were purchased from Life
Technologies (Darmstadt, Germany). For adipogenic differentiation
gene expression of lipoprotein lipase [LPL; accession number (AN):
NM_000237.2] and leptin (LEP; AN: NM_002303.5) were measured. The
osteogenic differentiation was confirmed by the analysis of
alkaline phosphatase (ALP; AN: NM_000478.4), bone
γ-carboxyglutamate protein (BGLAP; AN: NM_199173.4) and
runt-related transcription factor 2 (RUNX-2/cbfa-1; NM_004348.3).
The amplifications for gene quantification were: 50°C for 2 min;
95°C for 10 min, and 40 cycles at 95°C for 15 sec and 60°C for 1
min. ∆CT values are presented as relative quantification, which
were normalized to the gene expression of the reference gene GAPDH
(AN: NM_002046.3).
Cell migration
The scratch assay was used to clarify a possible
alteration in MSC migration induced by chronic exposure to 100 nM
salinomycin. MSC cells (1×105 cells/ml) were cultivated in a
12-well round bottom plate at 37°C. After 24 h, a straight-line
wound was induced with a sterile 1-ml pipette tip. Following this,
culture plates were washed with PBS, images were captured (t=0 h)
and incubated for another 24 h at 37°C with 5% CO2. To
analyze the relative migration, images of the plates were captured
(t=24 h) (Leica DMI 4000B Inverted Microscope) and the percentage
of the wound closure was evaluated. The area between the wound
borders at time t=24 h was defined as the percentage of wound
closure compared to the wound borders at time t=0 h. The
calculation was investigated using ImageJ (version 1.43u). MSC
cultivated with DMEM-EM served as the control. The experiment was
performed in triplicate using MSC of all patients (n=5).
Immunocytochemical analysis of
α-tubulin
MSC-Sal were washed several times with PBS and fixed
with 4% paraformaldehyde for 30 min at room temperature (RT).
Another 5 min of fixation with acetone at RT was performed.
Subsequently, cells were incubated with 10% bovine serum albumin
(BSA) (Carl Roth GmbH, Karlsruhe, Germany) in Tris-buffered saline
200 mM Tris-base (pH 8), 8% NaCl and 1% Tween-20 (TBS-T) (all from
Sigma-Aldrich). Incubation of MSC in TBS-T containing 1% BSA plus a
mouse monoclonal antibody against α-tubulin (1:500; Sigma-Aldrich)
was followed for 10 h. After three washing steps with TBS-T (each
step 5 min), MSC were incubated for 1 h in 1% BSA at RT with Alexa
488 goat anti-mouse secondary antibody 1:500 (Gibco Invitrogen) and
5 mg/ml DAPI (Sigma-Aldrich). A fluorescence microscope (Leica DMI
4000B Inverted Microscope) was used for cell examination.
Proliferation analysis
The expression of Ki67 was investigated via
immunocytochemical analysis and RT-qPCR. A mouse monoclonal
antibody against Ki67 (1:500; Sigma-Aldrich) was used. The gene
expression of Ki67 (AN: NM_002417.4) (Applied Biosystems) was
measured by RT-qPCR.
Genotoxicity
The alkaline single-cell microgel electrophoresis
(comet) assay was performed for verification, for example, of
possible DNA strand breaks. MSC were treated with 100 nM of
salinomycin for 4 weeks. The slides were prepared as previously
described by Buehrlen et al (11). The slides were observed by fluorescence
microscopy (magnification, ×400; DMLB Leica Microsystems) with the
following filters: A green excitation filter (515–560 nm band
pass), a dichromatic beam splitter (580 nm long pass), and an
emission filter (590 nm long pass). In every sample, 100 cells were
counted. For the DNA fragmentation analysis, the KOMET 5.5 image
system (Kinetic Imaging, Liverpool, UK) was used. Tail DNA (TD),
tail length (TL) and Olive tail moment (OTM) as a product of the
median migration distance and the percentage of DNA in the tail
were measured, as previously described by Olive et al
(12). The comet assay was performed
for all MSC-Sal (n=5) and MSC-control (n=5).
Statistical analysis
All the data were transferred to standard
spreadsheets and analyzed by statistical analysis (GraphPad Prism
5.0 software; GraphPad, Inc., La Jolla, CA, USA). The
Kruskal-Wallis test was carried out to evaluate statistical
significance. P<0.05 was considered to indicate a statistically
significant difference and are shown by an asterisk.
Results
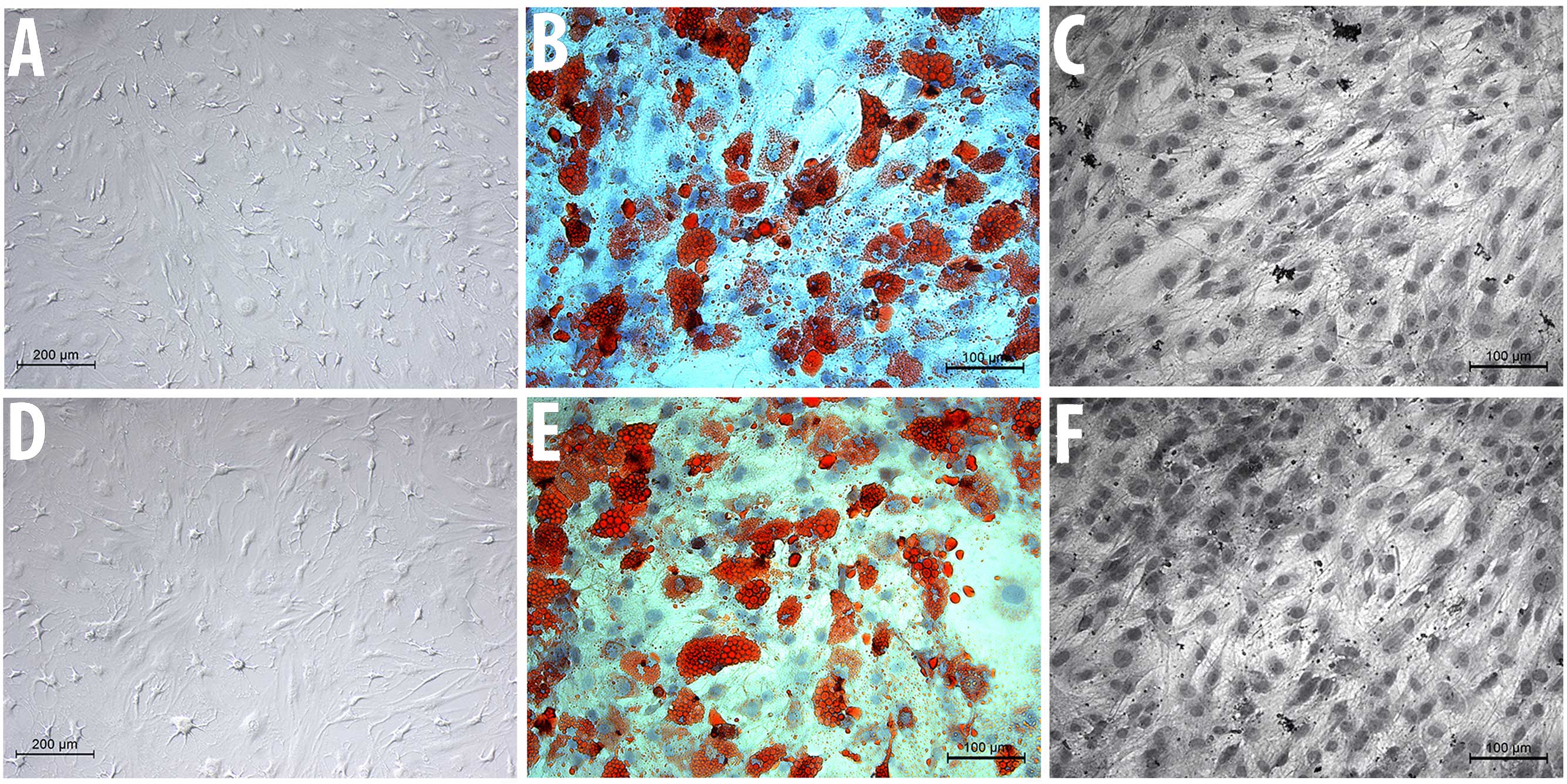
Cell morphology and
multidifferentiation capacity
The two cell groups, MSC-Sal and MSC-control,
maintained their typical spindle-shaped fibroblast-like morphology.
Salinomycin did not induce any signs of apoptosis or senescence.
The cell surface analysis revealed that MSC-Sal and MSC-control
were positive for CD90, CD73 and CD44, and negative for CD34
(Fig. 1).
MSC-Sal and MSC-control were cultivated in
osteogenic and adipogenic medium. The multidifferentiation capacity
was examined via staining methods, as well as with RT-qPCR. The Oil
Red O and the von Kossa staining showed that MSC-Sal and
MSC-control were able to differentiate into adipocytes and
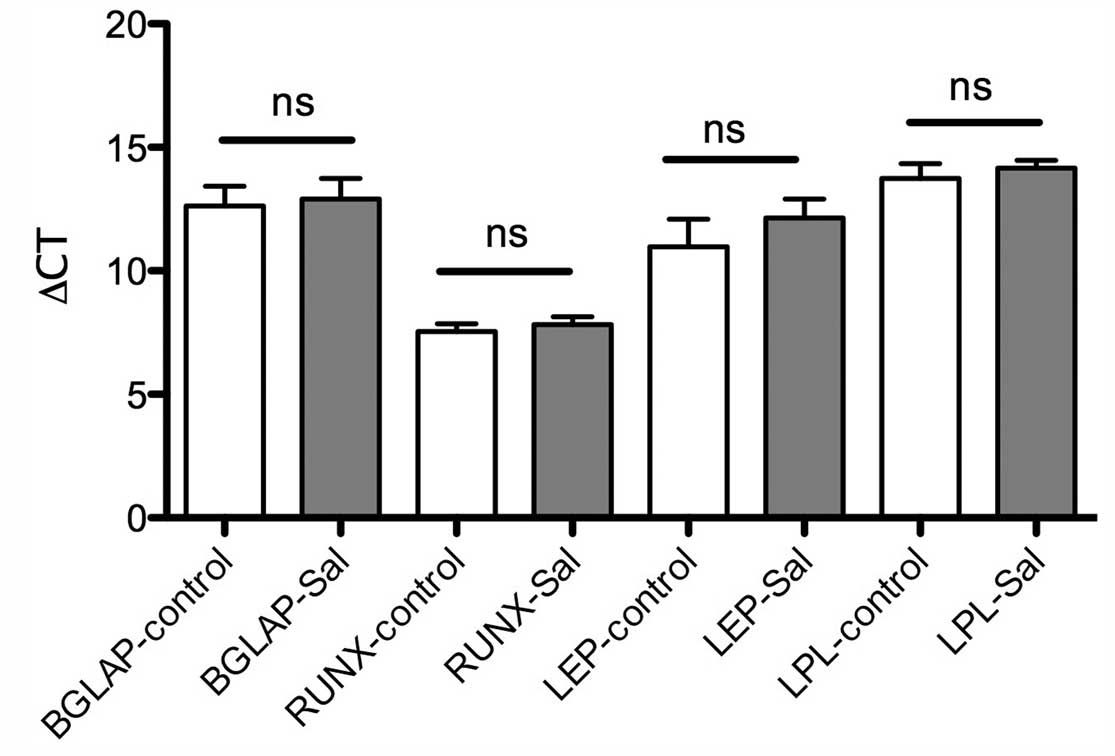
osteocytes. These results were confirmed by RT-qPCR. The gene
expression for osteogenic and adipogenic differentiation was
detectable in MSC-Sal and MSC-control following cultivation in
adipogenic and osteogenic medium. The ∆CT values showed no
significant differences (Fig. 2).
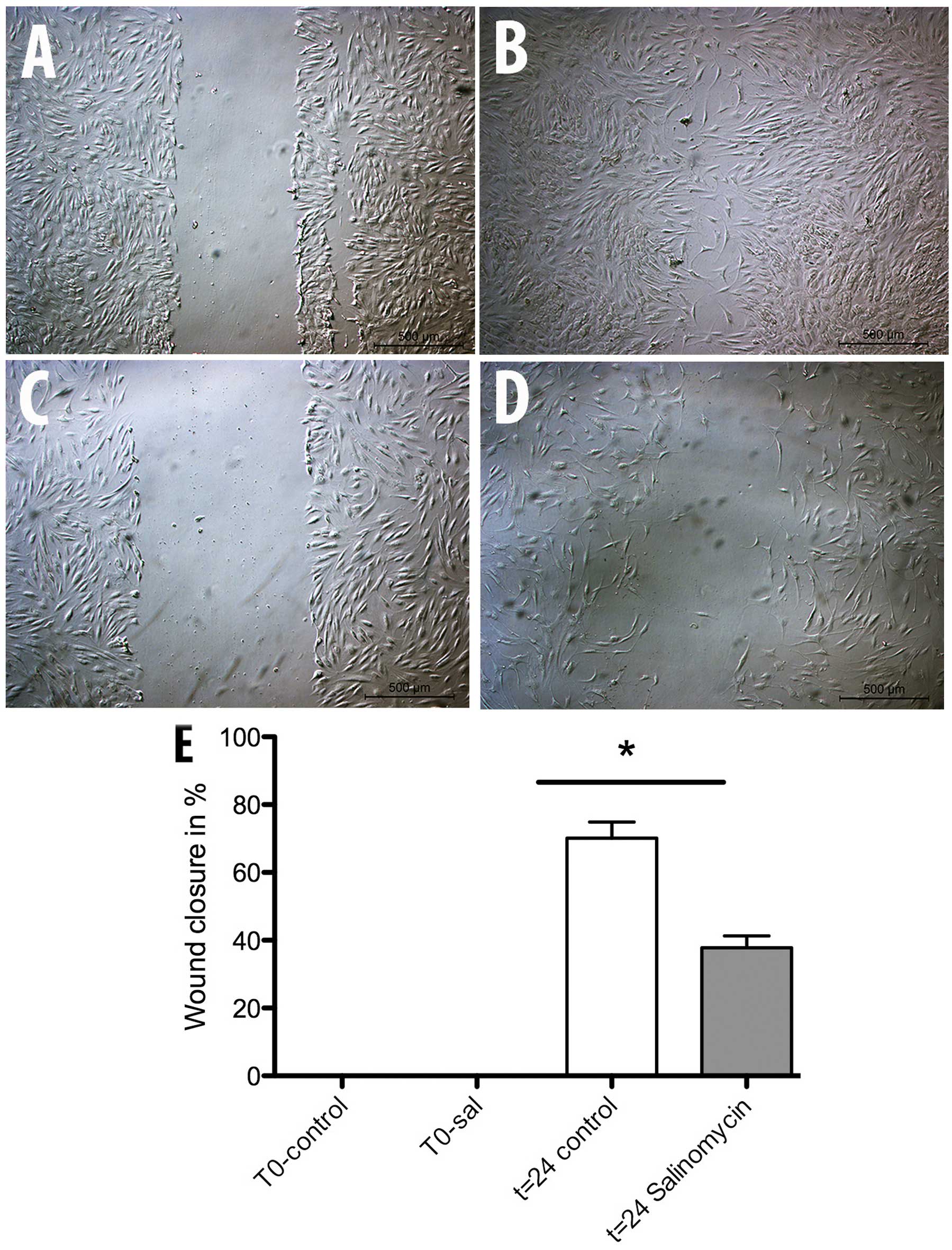
Migration capability
The scratch assay was used to determine a possible
influence of chronic salinomycin exposure on MSC migration. After a
repair period of 24 h at 37°C, the wound area was controlled and
the level of wound closure measured. The assay showed a significant
inhibition of MSC-Sal ability in closing the wound area compared to
the MSC-control (Fig. 3). MSC-control
induced 70% wound closure while MSC-Sal was only able to show a
wound closure of 40%.
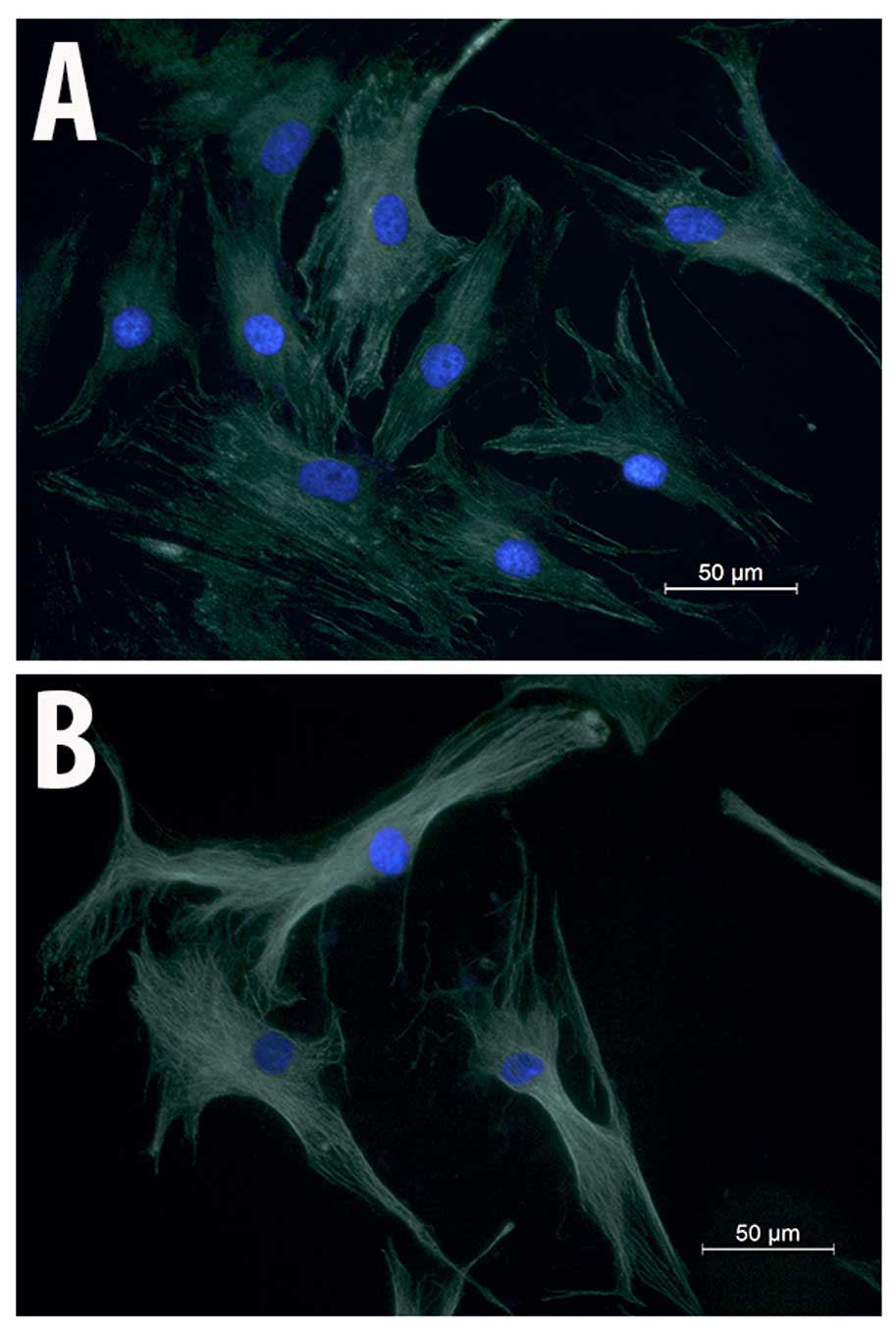
Immunocytochemical analysis of
α-tubulin
To examine whether the migration impairment induced
by salinomycin is associated with changes in MSC cytoskeletal
structure, immunocytochemical analysis of α-tubulin was performed.
MSC-Sal and MSC-control showed no effect on their cell cytoskeletal
structures. There were no signs for disruption of the tubulin
fibers or structural instability in MSC-Sal and MSC-control. The
shapes of the two cell types (MSC-Sal and MSC-control) did not show
any differences (Fig. 4).
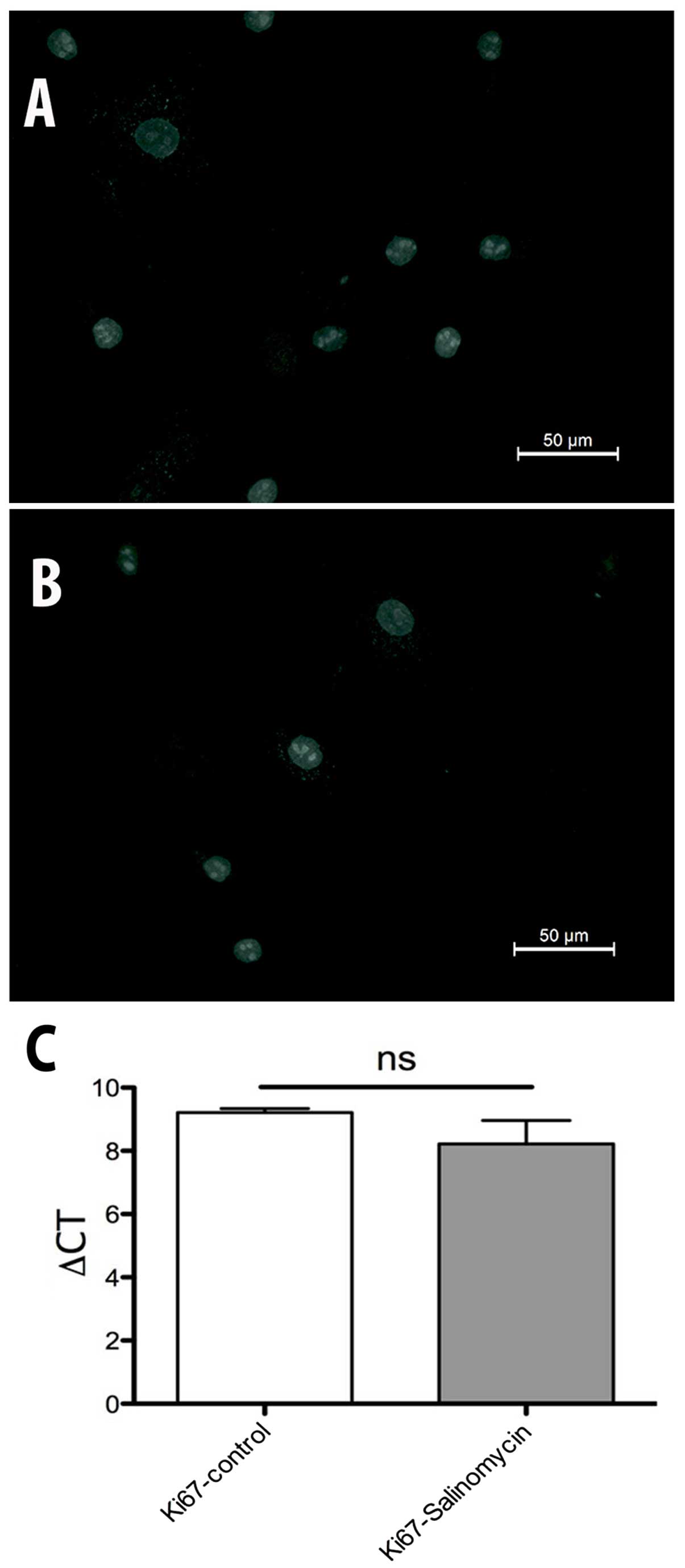
Proliferation analysis
The expression of Ki67 was investigated to evaluate
possible changes in the activity of cell proliferation. The
immunohistochemical analysis of Ki67 revealed no differences
between MSC-Sal and MSC-control. The results were confirmed by
RT-qPCR. ∆CT values in Fig. 5 show no
statistical significant difference between MSC-Sal and MSC-control
(Fig. 6).
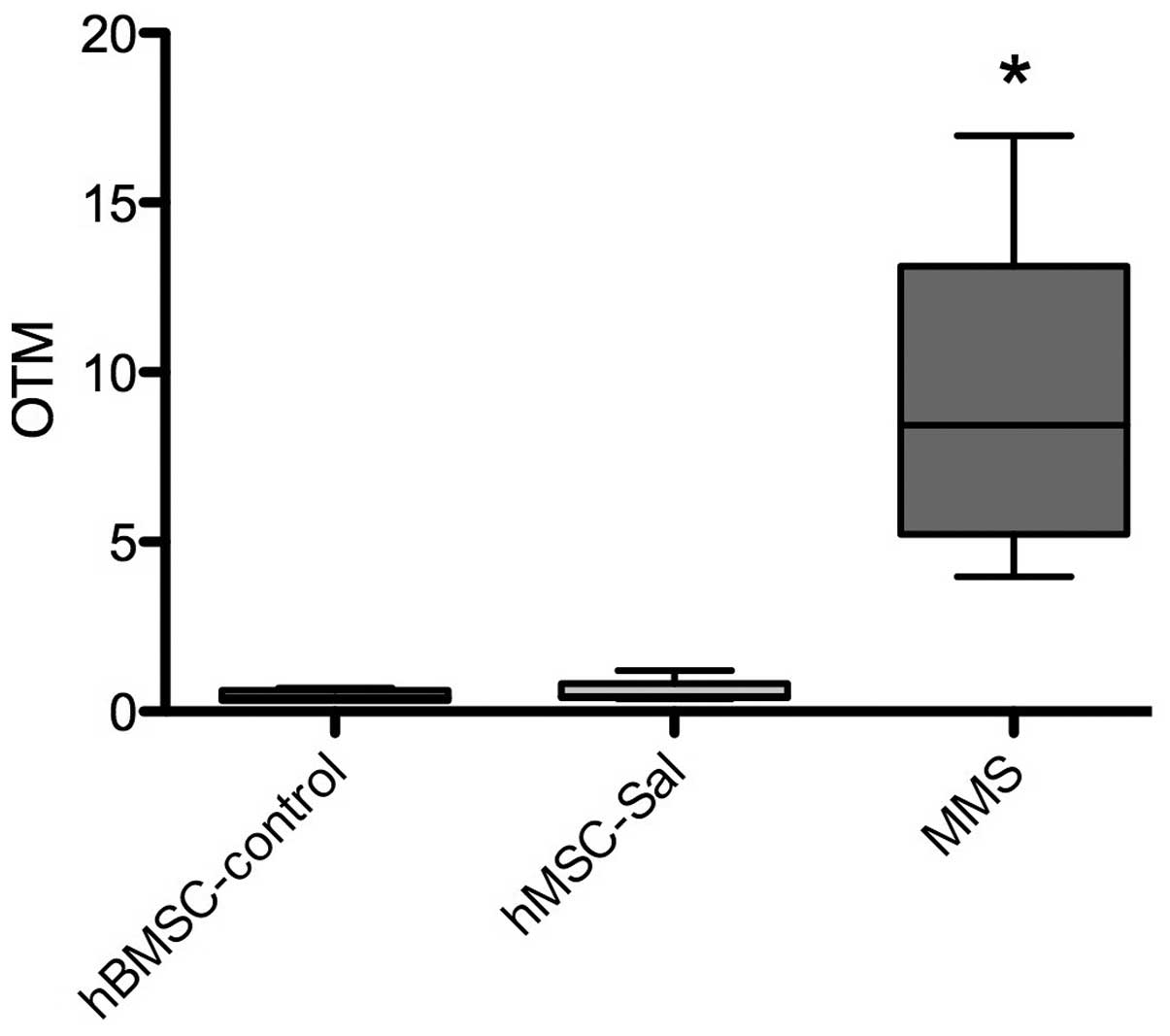
Genotoxicity
The comet assay was performed in MSC-Sal and
MSC-control. In all samples, no significant increase in the OTM, as
an indicator for genotoxic effects, was observed. MMS as a positive
control induced a significant increase in DNA breakage. Data are
presented in Fig. 5.
Discussion
Ionophores, such as salinomycin, induce toxicity by
increasing the intracellular sodium concentration, which
consecutively increases the intracellular Ca2+
concentration due to the Na+/Ca2+ exchange in
the plasma membrane and mitochondria (13). This proton exchange leads to an
alteration in pH, and thereby increases osmotic pressure inside the
cell, which may lead to apoptosis (14). Ionophores show a broad spectrum of
biological activity, such as antibacterial activity, particularly
for Gram-positive bacteria and also including antibiotic-resistant
S. aureus, as well as antiviral and antiparasitic activity
(15). In target animal species, such
as poultry, salinomycin did not exhibit any toxic side effects. The
toxicity of ionophores, such as monensin, has also been
investigated. Todd et al (16)
could identify acute toxic symptoms, such as anorexia,
hypoactivity, skeletal muscle weakness, ataxia, diarrhea, decreased
weight gain and delayed fatalities induced by monensin. Chronic
exposure also resulted in anorexia, weakness, ataxia and increased
serum muscle enzyme combined with severe skeletal muscle
degeneration and necrosis (16). The
data concerning salinomycin toxicity in humans is extremely poor
and divergent. In a clinical pilot study of a patient with
metastatic invasive ductal breast cancer, systemic salinomycin
application of 200 µg/kg every other day was tolerated extremely
well (17), and resulted in a
significant reduction in the cancer. The same effects were observed
in patients suffering from head and neck cancer as well as from
ovarian cancer. The side effects of intravenous salinomycin
following administration were tachycardia and mild tremor (17). By contrast, there are certain case
reports postulating harmful side effects induced by salinomycin,
such as rhabdomyolysis and renal failure (2,4). These
divergences between the results of case reports and clinical pilot
studies may be due to differences in the dosage of salinomycin. In
our recent study, 10–20 µM salinomycin induced cytotoxic effects in
human peripheral lymphocytes and nasal mucosa cells in vitro
(18). However, no genotoxic effects
were detectable at subcytotoxic levels. In the clinical studies,
the dosage of salinomycin ranged from 200–250 µg/kg. All the
patients experienced a reduction in the cancer, but complete
remission was not achieved. Hypothetically, this may be due to the
dosages of salinomycin applied. Higher amounts of salinomycin could
have a more potent effect on the cancer. Story and Doube (2) described an accidental ingestion of
salinomycin by a farmer. The dosage was not determined, however, it
was estimated that 1 mg/kg of the body weight was swallowed. This
resulted in life-threatening neuropathia, rhabdomyolysis and a
6-week hospitalization (2). Therefore,
the biological hazards in humans induced by salinomycin have to be
explored in greater detail as well.
In our previous study, acute exposure of salinomycin
to MSC revealed a dose-dependent attenuation of cell viability
without affecting the cell migration capability (19). The ability of MSC to differentiate into
osteocytes and adipocytes remained unchanged after treatment with
salinomycin for 24 h. As chronic exposure affects the
differentiation of adipocytes, it was worthwhile to investigate
whether chronic exposure of salinomycin inhibits the multipotency
of MSC as well. In the present study, the staining methods and gene
expression analysis revealed no alteration in MSC multipotency
following chronic salinomycin exposure as compared to the control
group. The cell cytoskeletal structure was also not affected.
Notably, however, the migration capability of MSC attenuated
significantly following chronic exposure to salinomycin. One
possible explanation of this phenomenon may be the regulation of
the C-X-C chemokine receptor type 4 (CXCR4) by salinomycin. CXCR4
is a ligand of the chemokine stromal-derived factor-1, and both
have an important role in the mediation of MSC migration (20). Liu et al (21) demonstrated an enhancement of MSC
migration following gene modification of MSC with CXCR4. Larzaba
et al (22) investigated the
effects of salinomycin on lung primary tumors and metastasis, and
in this study, salinomycin reduced the expression of CXCR4
significantly. CXCR4 is highly expressed by MSC within the bone
marrow and is markedly reduced during in vitro expansion
(23). The chronic exposure of
salinomycin may have induced a reduction in CXCR4. The evaluation
of CXCR4 and possible shortcomings of the present study will be
addressed in future studies: First, an exposure of 4 weeks with
salinomycin may be too short to detect side effects. Second, MSC
have a considerable ability to restore their biological behavior.
In the present study MSC were treated for 4 weeks with salinomycin.
Following this period they were cultured in expansion medium with
supplements either for adipogenic or osteogenic differentiation.
During the differentiation process, MSC were not exposed to
salinomycin. This may have had an impact on the regeneration of the
differentiation capability.
In conclusion, salinomycin induced a significant
attenuation of MSC migration, while differentiation, as well as
cell morphology and proliferation, were unchanged. Further
investigations, for example with different primary human cells, are
required in order to evaluate the potential cytotoxic effects of
acute and chronic salinomycin exposure.
References
|
1
|
Boehmerle W and Endres M: Salinomycin
induces calpain and cytochrome c-mediated neuronal cell death. Cell
Death Dis. 2:e1682011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Story P and Doube A: A case of human
poisoning by salinomycin, an agricultural antibiotic. N Z Med J.
117:U7992004.PubMed/NCBI
|
|
3
|
Caldeira C, Neves WS, Cury PM, Serrano P,
Baptista MA and Burdmann EA: Rhabdomyolysis, acute renal failure
and death after monensin ingestion. Am J Kidney Dis. 38:1108–1112.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kouyoumdjian JA, Morita MP, Sato AK and
Pissolatti AF: Fatal rhabdomyolysis after acute sodium monensin
(Rumensin) toxicity: Case report. Arq Neuropsiquiatr. 59:596–598.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mortier L, Huet AC, Charlier C, Daeseleire
E, Delahaut P and Van Peteghem C: Incidence of residues of nine
anticoccidials in eggs. Food Addit Contam. 22:1120–1125. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Szkudlarek-Mikho M, Saunders RA, Yap SF,
Ngeow YF and Chin KV: Salinomycin, a polyether ionophoric
antibiotic, inhibits adipogenesis. Biochem Biophys Res Commun.
428:487–493. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dominici M, Le Blanc K, Mueller I,
Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A,
Prockop Dj and Horwitz E: Minimal criteria for defining multipotent
mesenchymal stromal cells. The international society for cellular
therapy position statement. Cytotherapy. 8:315–317. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Horwitz EM, Le Blanc K, Dominici M,
Mueller I, Slaper-Cortenbach I, Marini FC, Deans RJ, Krause DS and
Keating A: International Society for Cellular Therapy:
Clarification of the nomenclature for MSC: The international
society for cellular therapy position statement. Cytotherapy.
7:393–395. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee RH, Kim B, Choi I, Kim H, Choi HS, Suh
K, Bae YC and Jung JS: Characterization and expression analysis of
mesenchymal stem cells from human bone marrow and adipose tissue.
Cell Physiol Biochem. 14:311–324. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Scherzed A, Hackenberg S, Froelich K,
Kessler M, Koehler C, Hagen R, Radeloff A, Friehs G and Kleinsasser
N: BMSC enhance the survival of paclitaxel treated squamous cell
carcinoma cells in vitro. Cancer Biol Ther. 11:349–357. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Buehrlen M, Harreus UA, Gamarra F, Hagen R
and Kleinsasser NH: Cumulative genotoxic and apoptotic effects of
xenobiotics in a mini organ culture model of human nasal mucosa as
detected by the alkaline single cell microgel electrophoresis assay
and the annexin V-affinity assay. Toxicol Lett. 169:152–161. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Olive PL, Durand RE, Le Riche J, Olivotto
IA and Jackson SM: Gel electrophoresis of individual cells to
quantify hypoxic fraction in human breast cancer. Cancer Res.
53:733–736. 1993.PubMed/NCBI
|
|
13
|
Dorne JL, Fernández-Cruz ML, Bertelsen U,
Renshaw DW, Peltonen K, Anadon A, Feil A, Sanders P, Wester P and
Fink-Gremmels J: Risk assessment of coccidostatics during feed
cross-contamination: Animal and human health aspects. Toxicol Appl
Pharmacol. 270:196–208. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Antoszczak M, Popiel K, Stefańska J,
Wietrzyk J, Maj E, Janczak J, Michalska G, Brzezinski B and
Huczyński A: Synthesis, cytotoxicity and antibacterial activity of
new esters of polyether antibiotic-salinomycin. Eur J Med Chem.
76:435–444. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huczyński A: Polyether
ionophores-promising bioactive molecules for cancer therapy. Bioorg
Med Chem Lett. 22:7002–7010. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Todd GC, Novilla MN and Howard LC:
Comparative toxicology of monensin sodium in laboratory animals. J
Anim Sci. 58:1512–1517. 1984.PubMed/NCBI
|
|
17
|
Naujokat C and Steinhart R: Salinomycin as
a drug for targeting human cancer stem cells. J Biomed Biotechnol.
2012:9506582012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Scherzad A, Hackenberg S, Schramm C,
Froelich K, Ginzkey C, Hagen R and Kleinsasser N: Geno- and
cytotoxicity of salinomycin in human nasal mucosa and peripheral
blood lymphocytes. Toxicol In Vitro. 29:813–818. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Scherzed A, Hackenberg S, Froelich K, Rak
K, Technau A, Radeloff A, Nöth U, Koehler C, Hagen R and
Kleinsasser N: Effects of salinomycin on human bone marrow-derived
mesenchymal stem cells in vitro. Toxicol Lett. 218:207–214. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wynn RF, Hart CA, Corradi-Perini C,
O'Neill L, Evans CA, Wraith JE, Fairbairn LJ and Bellantuono I: A
small proportion of mesenchymal stem cells strongly expresses
functionally active CXCR4 receptor capable of promoting migration
to bone marrow. Blood. 104:2643–2645. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu N, Tian J, Cheng J and Zhang J:
Migration of CXCR4 gene-modified bone marrow-derived mesenchymal
stem cells to the acute injured kidney. J Cell Biochem.
114:2677–2689. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Larzabal L, El-Nikhely N, Redrado M,
Seeger W, Savai R and Calvo A: Differential effects of drugs
targeting cancer stem cell (CSC) and non-CSC populations on lung
primary tumors and metastasis. PloS One. 8:e797982013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Marquez-Curtis LA and Janowska-Wieczorek
A: Enhancing the migration ability of mesenchymal stromal cells by
targeting the SDF-1/CXCR4 axis. BioMed Res Int. 2013:5610982013.
View Article : Google Scholar : PubMed/NCBI
|