Introduction
Anxiety and depression are widespread neurological
or psychological illnesses with high prevalence rates worldwide.
According to epidemiological studies, approximately one-third of
people will suffer from anxiety disorders (1). It is well accepted that neurotransmitter
deficiency is associated with mood disorders, specifically
serotonin and other catecholamines. Therefore, antidepressants,
such as the selective serotonin reuptake inhibitor (SSRI),
escitalopram (ESC), represent a fundamental pharmacotherapeutic
option.
Histamine is a neurotransmitter in the brain with
numerous physiological functions; it has also been implicated in
anxiety-like behaviour in animal models (2,3). Previous
evidence indicates a potential role for the influence of
histaminergic neurons in anxiety-associated behaviour via H1, H2
and H3 receptor activation. H3 receptor activation is hypothesized
to be involved in diminishing the release of serotonin, dopamine
and norepinephrine, which are all implicated in anxiety and mood
disorders (3,4). In addition, H3 receptor knockout in mice
demonstrated reduced anxiety (5).
The precise role of antihistamines in anxiety and
depression remains controversial. First generation antihistamines
have been widely used to alleviate anxiety and panic attacks
(6). The anxiolytic and antidepressant
effects of chlorpheniramine (CPA), a first generation
antihistamine, are proposed to be associated with its serotonergic
functions (7). Furthermore, the
anxiolytic and antidepressant effect of CPA was evident in animal
models (8,9). However, these results have been
challenged by previous findings (10)
in which CPA was demonstrated to exert anxiogenic effects in
mice.
In addition to the histaminergic system, growing
evidence from animal and human trials implicates oxidative stress
in anxiety and stress disorders in the central nervous system and
in peripheral tissues (11,12). Oxidative stress is defined as the
overproduction of free radicals, such as reactive oxygen species
and reactive nitrogen species accompanied by the inability of
endogenous defensive antioxidant enzymes to detoxify these free
radicals (11). Nitrates, a nitric
oxide (NO) metabolite present in different tissues and the chief
reactive nitrogen species constituent, is a marker of oxidative
stress and is implicated in associated disorders (13); NO increases in acute anxiety (14,15).
Furthermore, various animal studies have reported higher NOx in the
hippocampus and brain cortex of a stressed animal (16). Furthermore, SSRIs were found to exert
an antioxidant role peripherally (17). Super oxide dismutase (SOD) is an
abundant antioxidant enzyme responsible for super oxide
(O2−) species detoxification to yield
hydrogen peroxide (H2O2). SOD activation
occurs as a response to an oxidative stress status (18).
Clinically, anxiolytics or antidepressants are used
chronically, rather than as a single dose. Although CPA anxiolytic
and antidepressant effects were investigated using a single dose
(8,18,19), their
effects, however, were not evaluated following repeated doses.
Additionally, the role of CPA in oxidative stress modulation has
yet to be addressed. Therefore, the aim of the present study was to
evaluate the protective anxiolytic and antidepressant effects of
three weeks of CPA administration in comparison to ESC, and to
correlate CPA behavioural effects on serum NOx levels and SOD
activity in mice anxiety.
Materials and methods
Study animals and design
Forty BALB/c mice (Animal House Facility of the
Applied Science University, Amman, Jordan) were used in the present
study. The mice were aged 6–8 weeks and weighed ~25 g. The BALB/c
line was selected due to its suitability in anxiety studies
(20). Mice were maintained in
separate cages, at a temperature of ~25°C, with 50–60% humidity and
continuous air ventilation. The research was conducted according to
the international ethical standards for the Care and Use of
Laboratory Animals and the study was approved by the American
University of Madaba research committee (Madaba, Jordan). The mice
were divided into four groups (n=10 per group) as follows: Group 1,
naïve (unstressed and untreated); group 2, control (treated with
distilled water for 3 weeks subsequently stressed); group 3, CPA
(treated for 3 weeks and subsequently stressed); group 4, ESC
(treated for three weeks and subsequently stressed); and group 5
received a combination of 0.5 mg/kg CPA and 10 mg/kg ESC (treated
for three weeks and subsequently stressed). A treatment period of 3
weeks was selected in order to investigate the antidepressant
potential. Following completion of the 3-week treatment, groups 2,
3 and 4 were restrained in Falcon tubes for 6 h between 9:00 and
15:00, then all mice were subjected to behavioural tests. The mice
were sacrificed, blood samples (~1.5 ml) were collected and the
serum was obtained by centrifugation at 2,000 × g for 10 min at
room temperature.
Treatment strategies
The CPA-treated group received a dose of 0.5 mg/kg
intraperitoneally for 3 weeks. The ESC-treated group received a
dose of 10 mg/kg orally for 3 weeks. CPA and ESC were generously
donated by the Arab Pharmaceutical Manufacturing Co., Ltd. (Amman,
Jordan) and the JOSWE Medical (Amman, Jordan), respectively. The
control group received an equal volume of distilled water
intraperitoneally for 3 weeks. The majority of similar studies
employed acute dosing regimens (1 h prior to the behavioural
tests); however, chronic dosing for 3 weeks with ESC has previously
been performed (21). In addition, the
effect of chronic dosing (at least 2 weeks) was evaluated in the
present study to establish the possible antidepressant effect that
may resemble clinical settings.
Acute immobility stress
In order to induce anxiety in mice, an acute
immobility stress test was performed according to the method of
Machawal and Kumar (22) with slight
modifications. The current findings indicate that acute immobility
stress increases serum levels of nerve growth factor (data not
published), which is associated with anxiety (23).
In the present study, the mice were immobilised
individually for 6 h in a 50-ml Falcon tube while proper
ventilation was maintained. In the naïve group, the mice were
maintained in an animal cage with soft bedding under the same
experimental conditions. After performing the immobility stress
test, the mice were subjected to behavioural tests.
Behavioural tests
Forced swim test (FST)
The FST is the most commonly used behavioural model
for screening antidepressant-like activity in rodents (24). Mice were individually forced to swim
for 5 min in an open glass chamber (25×15×25 cm3)
containing fresh water to a height of 15 cm and maintained at
26±1°C. Floating time (FT) was defined as the time in which mice
stop moving completely while in the water.
Elevated plus maze (EPM)
The EPM test, a model for screening anxiolytics, was
performed as described previously (25) with certain modifications. The apparatus
was elevated 25 cm above the floor. The maze is composed of two
closed arms (30×5×10 cm) and two open arms (30×5cm). Mice were
placed at the centre, facing the closed arm, and allowed to move
freely for 10 min. The frequency of open arm exists (OAE) and the
open arm time (OAT) spent were recorded by an experienced
technician.
Open field test (OFT)
An OFT was performed to assess the locomotion
activity and sedation of the mice (26). Briefly, the mice were placed in a
central square and allowed to move freely for 5 min. The field was
located in a test room and lit by indirect lighting. The procedure
was performed in an empty room to minimise noise and distractions.
The open field maze was cleaned between each mouse, using 70% ethyl
alcohol. The locomotion activity (represented by the number of
lines crossed) and the sedation (represented by the rearing
frequency) were recorded by an experienced technician.
Biochemical tests
Subsequent to performing the behavioural tests, the
mice were sacrificed, blood (~1.5 ml) was collected and serum was
obtained using standard protocols. The accumulation of nitrate, an
indicator of the production of NO, was determined using a
colorimetric assay with a Griess reagent (27). Serum nitrate was assayed using a Nitric
Oxide Assay kit (cat. no. ab65328; Abcam, Cambridge, MA, USA)
according to the manufacturer's instructions. The nitrate
concentration was obtained according to the standard curve
generated after measuring absorbance at a wavelength of 540 nm
using a Multiskan™ FC Microplate Photometer (Thermo Fisher
Scientific, Waltham, MA, USA). All samples and standards were
processed in duplicate.
SOD activity was assayed using an SOD Assay kit
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany; cat. no. 19160) as
previously described (28). The kit
utilises Dojindo's highly water-soluble tetrazolium salt, WST-1
[2-(4-Iodophenyl)- 3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-
2H-tetrazolium, monosodium salt], which produces a water-soluble
formazan dye upon reduction with a superoxide anion. The rate of
the reduction with O2 is linearly associated with the
xanthine oxidase activity, and is inhibited by SOD. The SOD
activity, as an inhibition activity, is quantified by measuring the
decrease in the colour development at a wavelength of 440 nm.
Statistical analysis
All data obtained from the behavioural tests, and
the NO and SOD activity were analyzed with a one-way ANOVA and a
subsequent Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference and quantitative
data are presented as the mean ± standard error of the mean.
Results
FST
The results of the FST are presented in Fig. 1. The results of the FT revealed that
the CPA- and the ESC-treated groups exhibited a similar and
significant decrease when compared with the control group. The
result of combination therapy did not vary when compared with that
of each therapy alone (P>0.05).
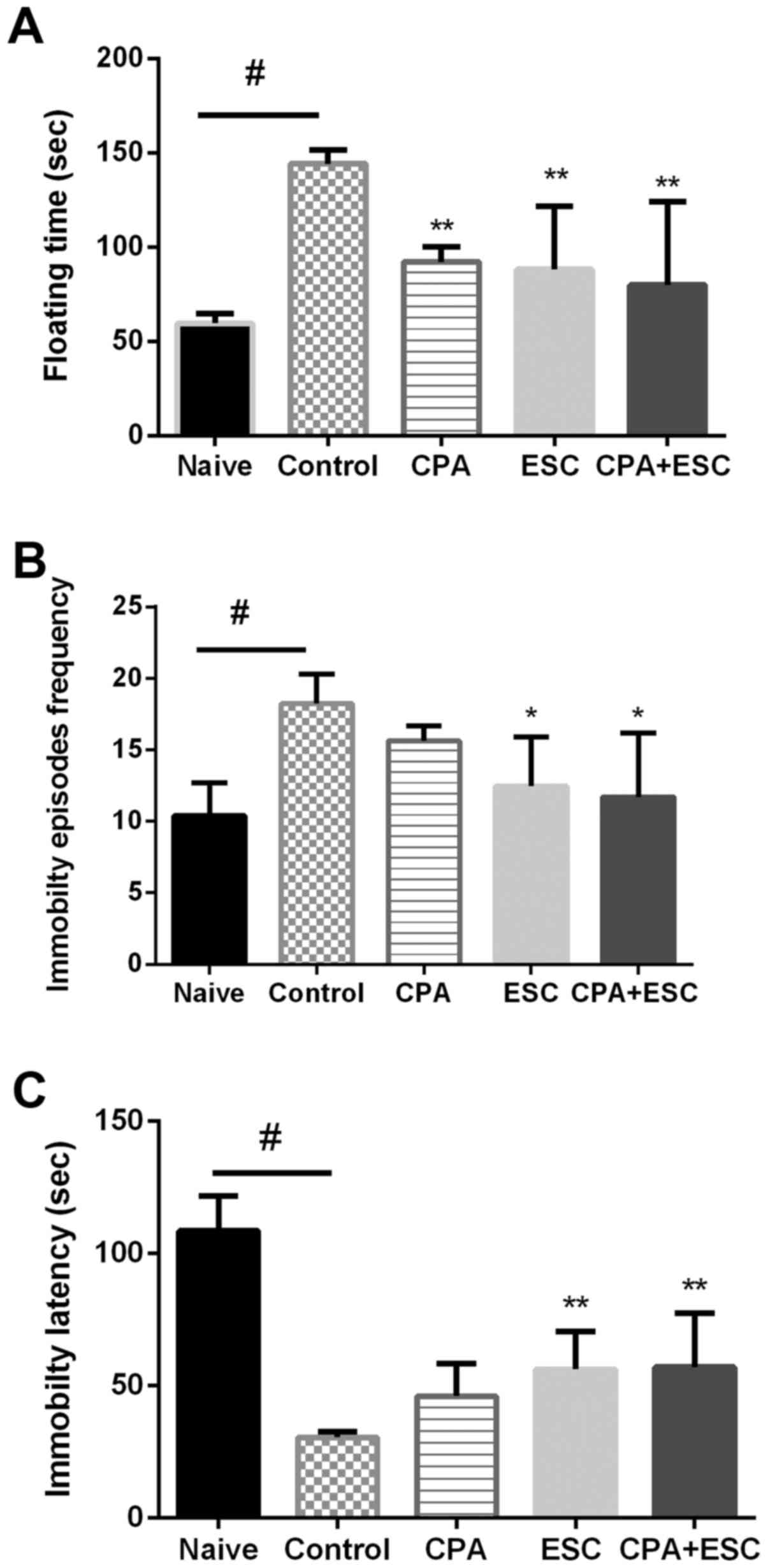 | Figure 1.(A) Effect of 3 weeks of treatment
with CPA (0.5 mg/kg), ESC (10 mg/kg) and a combination of the two
on FT following immobilization stress. Values are expressed as the
mean ± SEM (ANOVA followed by Tukey's test). F(4, 45)=10.83;
**P<0.05 and #P<0.00001. (B) Effect of treatment
with CPA (0.5 mg/kg), ESC (10 mg/kg) and a combination of the two
for 3 weeks on the immobility episodes frequency. Each bar
represents the mean ± SEM (ANOVA followed by Tukey's test). F(4,
39)=5.037; #P=0.002 and *P<0.05. (C) Effect of
treatment with CPA (0.5 mg/kg), ESC (10 mg/kg) and a combination of
the two for 3 weeks on immobility latency. Each bar represents the
mean ± SEM (ANOVA followed by Tukey's test). F(4, 43)=41.83;
#P=0.0001 and **P<0.001. CPA, chlorpheniramine; ESC,
escitalopram; FT, floating time; SEM, standard error of the
mean. |
In addition, the control group demonstrated
increased FT when compared with the naïve group (P<0.00001) and
the CPA group demonstrated higher FT than the naïve group (Fig. 1A; P<0.05). Regarding the immobility
episodes frequency (Fig. 1B), the
control group demonstrated a higher frequency of immobility
episodes when compared with the naïve group (P<0.05). The
ESC-treated group demonstrated a significant decrease in FT when
compared with the control group (P<0.05), the CPA-treated group
exhibited lower, but non-significant immobility episodes when
compared with the control, with no difference between the CPA- and
ESC-treated groups (P>0.05). The result of combination therapy
did not vary when compared with that of each therapy alone
(P>0.05).
The results of the immobility latency (Fig. 1C) revealed significantly lower
immobility latency in the control group when compared with the
naïve group (P<0.05). The ESC-treated group and the combination
therapy group significantly improved the immobility latency
(P<0.05). The CPA-treated group demonstrated higher, but
non-significant immobility latency when compared with the control
group. The result of combination therapy did not vary when compared
with that of each therapy alone (P>0.05).
EPM
The OAT and OAE were significantly higher in the
naïve group when compared with the control group (P<0.05). The
administration of CPA (0.5 mg/kg), ESC (10 mg/kg) and a combination
of the two for 3 weeks did not alter the OAT and OAE measurements
when compared with the control group (P>0.05; Fig. 2A and B).
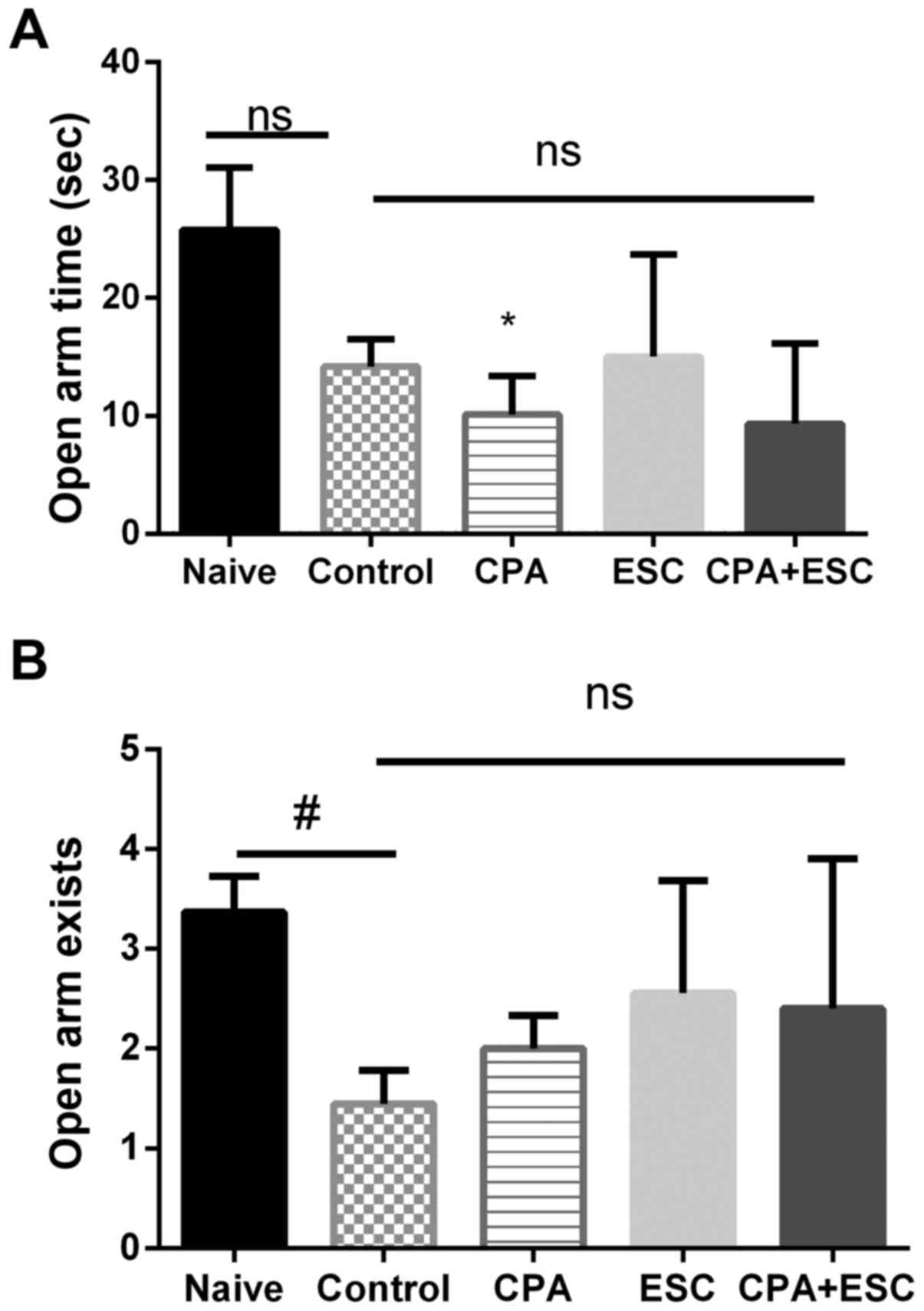 | Figure 2.(A) Effect of 3 weeks of treatment
with CPA (0.5 mg/kg), ESC (10 mg/kg) and a combination of the two
on OAT following immobilization stress. Values are expressed as the
mean ± SEM (ANOVA followed by Tukey's test). F(4, 38)=3.648;
*P<0.05. (B) Effect of treatment with CPA (0.5 mg/kg), ESC (10
mg/kg) and a combination of the two for 3 weeks on OAE. ANOVA
followed by Tukey's test. F(4, 39)=3.725; #P<0.05.
CPA, chlorpheniramine; ESC, escitalopram; FT, floating time; OAE,
open arm exists; SEM, standard error of the mean; ns,
non-significant. |
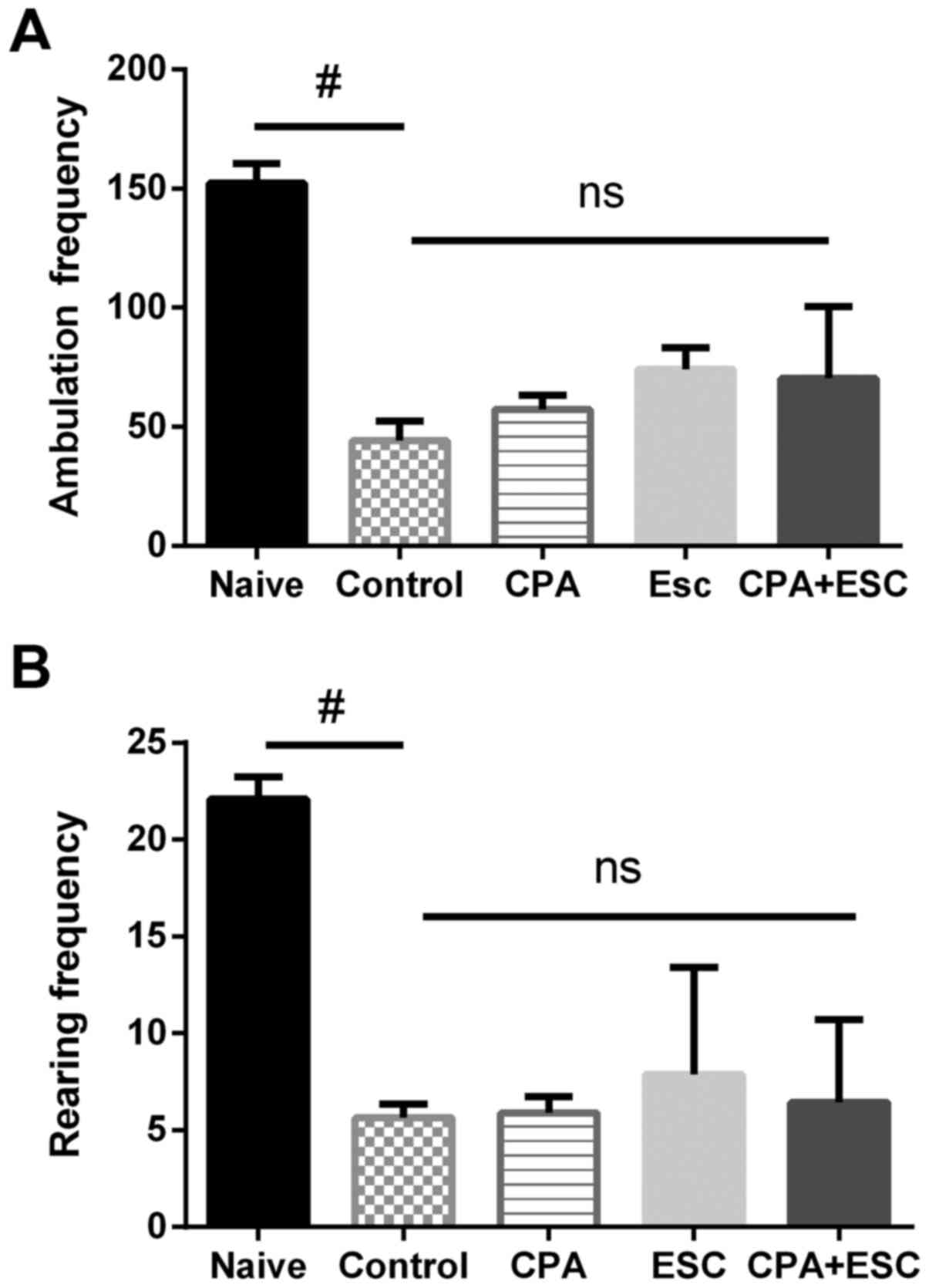
OFT
The results demonstrated a significant decrease in
the locomotion activity in the control group when compared with the
naïve group (P<0.0001). The administration of CPA (0.5 mg/kg),
ESC (10 mg/kg) and a combination of the two for 3 weeks did not
increase the ambulation frequency (P>0.05). Similar results were
observed for the rearing frequency. Acute restraint significantly
decreased rearing in the control group when compared with the naïve
group (P<0.0001). The administration of CPA (0.5 mg/kg), ESC (10
mg/kg) and a combination of the two for 3 weeks did not increase
the rearing frequency (P=0.98; Fig. 3A and
B).
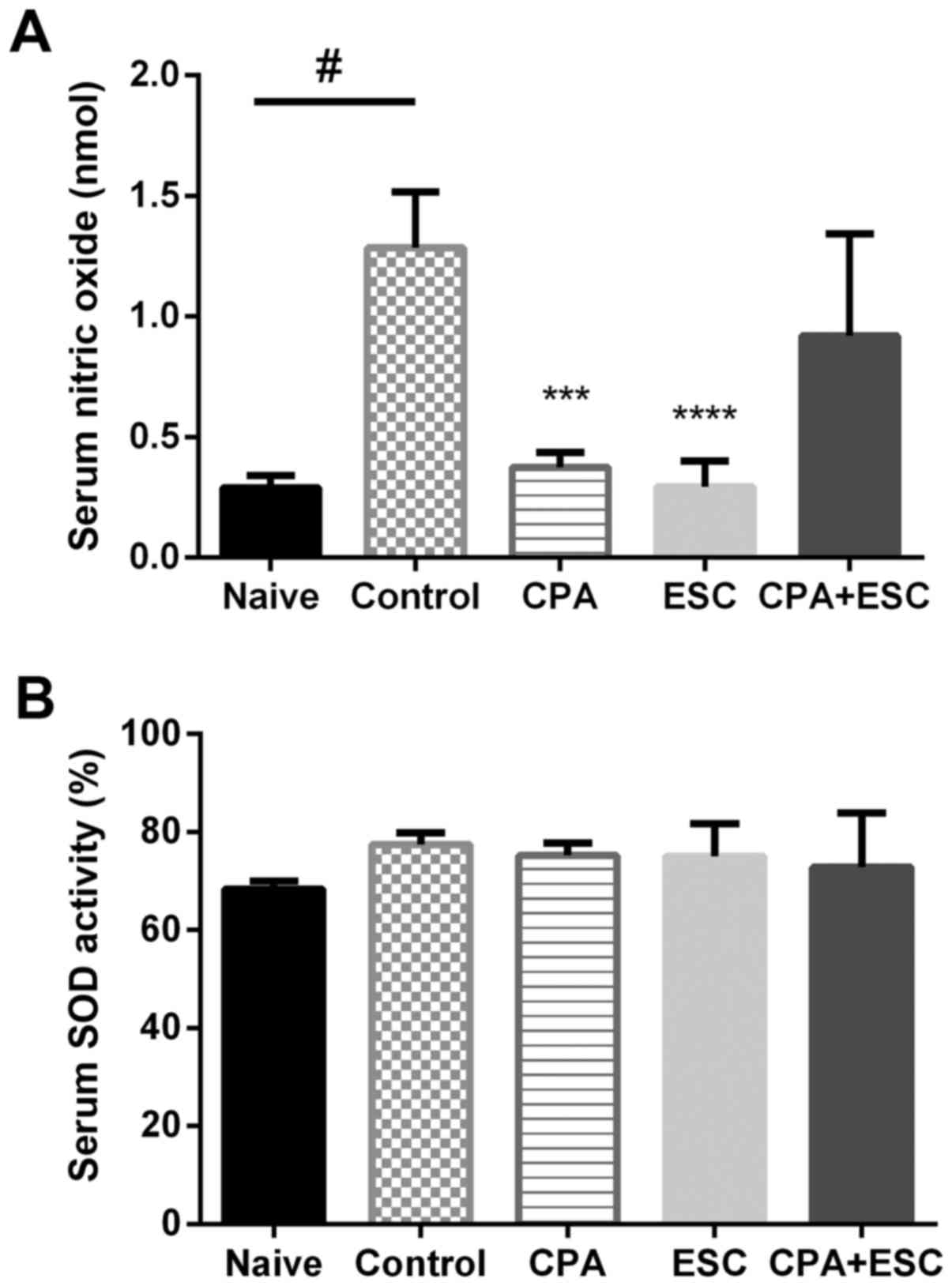
Biochemical tests
Serum NO
The immobility stress resulted in a significant
increase in serum NOx in the control group when compared with the
naïve group (P=0.0002). The CPA- and the ESC-treated groups
demonstrated lower NOx when compared with the control group
(P<0.05). However, the combination therapy did not result in any
reduction (P>0.05; Fig. 4A).
SOD activity
SOD serum activity did not vary among the study
groups (P>0.05; Fig. 4B).
Discussion
The current study revealed an antidepressant, but
not anxiolytic role of CPA following 3 weeks of administration in
mice. Additionally, CPA prevented stress-induced NOx increase,
although it did not alter SOD activity with respect to the
control.
In the current study, CPA treatment demonstrated
antidepressant action similar to that of ESC treatment. A possible
explanation could be attributed to its possible serotonergic
activity, as CPA shares structural properties with SSRIs (29). Another explanation is that CPA exerts
an antidepressant role by enhancing neurotransmitters additional to
serotonin, such as norepinephrine (30). A previous st udy (8) demonstrated a CPA antidepressant effect
via an FST in rodents following repeated acute doses of 7.5 and 15
mg/kg that were administered in the 24 h prior to the FST. In the
current study, a lower dose of CPA (0.5 mg/kg) was used for 3
weeks, which demonstrated antidepressant properties. A reduced dose
of CPA was selected in order to minimise the cholinergic and
sedative side effects (31). Notably,
the combination treatment of CPA with ESC was not synergistic.
Typically, synergism is achieved by combining drugs that act on
different receptors. Although further studies are required to
explain this finding, one hypothesis is that CPA may be competing
for the same serotonin transporter as ESC, based on its postulated
serotonergic activity.
An unexpected finding was that CPA failed to exert
anxiolytic effects in the EPM and the OFT. These findings are
consistent with Serafim et al (10) who demonstrated the anxiogenic effect of
CPA in mice at various doses. It was proposed that the H1 receptor
antagonists supress acetylcholine release from the ventral
striatum; furthermore, acetylcholine is associated with
anxiety-like behaviours in rodents (10).
Conversely, certain studies reported the anxiolytic
role of CPA in animals (25,32). Indeed, it has been suggested that CPA
anxiolysis may be attributed to modulation of the serotonergic and
cholinergic systems (7). In fact, CPA
inhibits the serotonin (5-hydroxytryptamine) transporter (6) and, therefore, inhibits serotonin
reuptake.
Our findings may be due to methodological
differences; for example, prior to performing the anxiety
behavioural tests in the present study, the mice were restrained.
Previous findings indicate that acute restraint induces anxiety
(22). Therefore, it is hypothesized
that the low dose of CPA employed in the present study was
insufficient to alleviate anxiety-like behaviours. This explanation
is supported by the fact that previous studies did not use
immobility stress. Furthermore, other studies employed higher CPA
doses (8). Therefore, it may be
inferred that a low dose of CPA is only useful in mild anxiety.
Acute anxiety provokes an increase in NOx in various
tissues, including the hippocampus, in blood and in saliva
(13,14). To the best of our knowledge, this is
the first study demonstrating the antioxidant effects of CPA. A
novel finding in the current study was the ability of CPA treatment
to normalise the stress-induced increase in serum NOx. Similarly,
ESC treatment reduced serum NOx. Although the exact mechanism is
yet to be established, this may be due to similarities in the
mechanisms between CPA and ESC.
Although SOD activity was increased following acute
stress induction, neither CPA nor ESC diminished its activity. The
exact cause of the rise in SOD activity due to anxiety is yet to be
clarified. It may be a compensatory mechanism to overcome oxidative
stress. Recently, SOD has been gaining attention in mood disorders.
For example, SOD mRNA levels were upregulated following acute
stress exposure (31) while in another
study, mild chronic stressful events decreased SOD activity in the
hippocampus and cortex of mice (33).
This may indicate a potential role for the antioxidant enzyme in
acute stress. The authors suggest that investigating the
transcripts of antioxidant enzymes could be more beneficial than
analyzing enzymatic activity. It was reported that long term use of
antidepressants upregulated the mRNA expression levels of SOD and
other antioxidant enzymes (34).
The present study presents novel ideas, which may
lay the foundation for future investigations; however, there were
certain limitations. Only a single concentration of CPA was
evaluated for its antidepressant and antioxidant effects. Future
studies should administer different doses that could determine
potential dose-dependent effects and potential synergism between
CPA and ESC. Furthermore, future studies may focus on the potential
role of CPA or ESC in modulating the levels or the activity of the
inducible NO synthase [the enzyme responsible for NOx synthesis
under stressful conditions (13)],
which may demonstrate the cross talk between mood disorders and
oxidative stress in the serum, as well as in the brain cortex,
allowing definitive conclusions to be derived.
In conclusion, this is the first study, to the best
of our knowledge, describing the antidepressant and the potential
antioxidant role of CPA in a mouse model of anxiety. This
preliminary finding provides novel hypotheses for future studies
regarding depression. Thus, further investigations are required to
clarify the underlying mechanisms and the possible implementation
in clinical practice.
Acknowledgements
The authors would like to thank Mr. Salem
Al-Shawabkeh (Department of Pharmacy, Applied Science University,
Amman, Jordan), Dr Amjad Abu Rumaileh (Associate Professor of
Pharmacology, Al-Isra University; Amman, Jordan) and Professor S.
G. Moscati (Department of Pharmacy, American University of Madaba)
for their dedication and efforts. The authors would like to thank
Mr. Rafael Angelo Custode (Department of Logistics, American
University of Madaba), Dr Moscati and Mr. Nour Hamarneh (Department
of Pharmacy, American University of Madaba) for their help with the
current study. The project was funded by the American University of
Madaba.
References
|
1
|
Hawgood J and De Leo D: Anxiety disorders
and suicidal behaviour: An update. Curr Opin Psychiatry. 21:51–64.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brown RE, Stevens DR and Haas HL: The
physiology of brain histamine. Prog Neurobiol. 63:637–672. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kumar KV, Krishna DR and Palit G:
Histaminergic H1 receptors mediate L-histidine-induced anxiety in
elevated plus-maze test in mice. Behav Pharmacol. 18:213–217. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zarrindast MR, Nasehi M, Piri M and Bina
P: Anxiety-like behavior induced by histaminergic agents can be
prevented by cannabinoidergic WIN55,212-2 injected into the dorsal
hippocampus in mice. Pharmacol Biochem Behav. 94:387–396. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rizk A, Curley J, Robertson J and Raber J:
Anxiety and cognition in histamine H3 receptor-/- mice. Eur J
Neurosci. 19:1992–1996. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hellbom E and Humble M: Panic disorder
treated with the antihistamine chlorpheniramine. Ann Allergy Asthma
Immunol. 90:361362, author reply 361–362. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tatsumi M, Groshan K, Blakely RD and
Richelson E: Pharmacological profile of antidepressants and related
compounds at human monoamine transporters. Eur J Pharmacol.
40:249–258. 1997. View Article : Google Scholar
|
|
8
|
Hirano S, Miyata S, Onodera K and Kamei J:
Effects of histamine H(1) receptor antagonists on depressive-like
behavior in diabetic mice. Pharmacol Biochem Behav. 83:214–220.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Onodera K, Sakurai E and Niwa H: Effect of
chlorpheniramine on muricide induced by thiamine deficiency:
Pharmacokinetic and behavioral studies. Agents Actions. 20:229–232.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Serafim KR, Kishi MS, Canto-de-Souza A and
Mattioli R: H1 but not H2 histamine antagonist receptors mediate
anxiety-related behaviors and emotional memory deficit in mice
subjected to elevated plus-maze testing. Braz J Med Biol Res.
46:440–446. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gałecki P, Szemraj J, Bieńkiewicz M,
Zboralski K and Gałecka E: Oxidative stress parameters after
combined fluoxetine and acetylsalicylic acid therapy in depressive
patients. Hum Psychopharmacol. 24:277–286. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wei YC, Zhou FL, He DL, Bai JR, Hui LY,
Wang XY and Nan KJ: The level of oxidative stress and the
expression of genes involved in DNA-damage signaling pathways in
depressive patients with colorectal carcinoma. J Psychosom Res.
66:259–266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen HJC, Spiers JG, Sernia C and Lavidis
NA: Response of the nitrergic system to activation of the
neuroendocrine stress axis. Front Neurosci. 9:32015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gammoh OS, Al-Smadi A, Al-Awaida W, Badr
MM and Qinna NA: Increased salivary nitric oxide and G6PD activity
in refugees with anxiety and stress. Stress Health. 32:435–440.
2016. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jin L, Qin L, Xia D, Liu X, Fan Z, Zhang
C, Gu L, He J, Ambudkar IS, Deng D and Wang S: Active secretion and
protective effect of salivary nitrate against stress in human
volunteers and rats. Free Radic Biol Med. 57:61–67. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Olivenza R, Moro MA, Lizasoain I, Lorenzo
P, Fernández AP, Rodrigo J, Boscá L and Leza JC: Chronic stress
induces the expression of inducible nitric oxide synthase in rat
brain cortex. J Neurochem. 74:785–791. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Matchkov VV, Kravtsova VV, Wiborg O,
Aalkjaer C and Bouzinova EV: Chronic selective serotonin reuptake
inhibition modulates endothelial dysfunction and oxidative state in
rat chronic mild stress model of depression. AJP Regulatory
Integrative and Comparative Physiology. 15:R814–R823. 2015.
View Article : Google Scholar
|
|
18
|
Blokhina O, Virolainen E and Fagerstedt
KV: Antioxidants, oxidative damage and oxygen deprivation stress: A
review. Ann Bot. 91:179–194. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shimazoe T, Shibata S and Ueki S: A new
forced swimming test for the evaluation of antidepressants in rats
by recording vibration of a water tank. J Pharmacobiodyn.
10:639–643. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Roy V, Belzung C, Delarue C and Chapillon
P: Environmental enrichment in BALB/c mice: Effects in classical
tests of anxiety and exposure to a predatory odor. Physiol Behav.
74:313–320. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Matchkov VV, Kravtsova VV, Wiborg O,
Aalkjaer C and Bouzinova EV: Chronic selective serotonin reuptake
inhibition modulates endothelial dysfunction and oxidative state in
rat chronic mild stress model of depression. Am J Physiol Regul
Integr Comp Physiol. 309:R814–R823. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Machawal L and Kumar A: Possible
involvement of nitric oxide mechanism in the neuroprotective effect
of rutin against immobilization stress induced anxiety like
behaviour, oxidative damage in mice. Pharmacol Rep. 66:15–21. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gioiosa L, Iannitelli A and Aloe L:
Stress, anxiety and schizophrenia and neurotrophic factors: The
pioneer studies with nerve growth factor. Riv Psichiatr. 44:88–94.
2009.PubMed/NCBI
|
|
24
|
Porsolt RD, Bertin A and Jalfre M:
‘Behavioural despair’ in rats and mice: Strain differences and the
effects of imipramine. Eur J Pharmacol. 51:291–294. 1978.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Privou C, Knoche A, Hasenöhrl RU and
Huston JP: The H1- and H2-histamine blockers chlorpheniramine and
ranitidine applied to the nucleus basalis magnocellularis region
modulate anxiety and reinforcement related processes.
Neuropharmacology. 37:1019–1032. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dishman RK, Armstrong RB, Delp MD, Graham
RE and Dunn AL: Open-field behavior is not related to treadmill
performance in exercising rats. Physiol Behav. 43:541–546. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Green LC, Wagner DA, Glogowski J, Skipper
PL, Wishnok JS and Tannenbaum SR: Analysis of nitrate, nitrite, and
[15N]nitrate in biological fluids. Anal Biochem. 126:131–138. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Omori A, Yoshimura Y, Deyama Y and Suzuki
K: Rosmarinic acid and arbutin suppress osteoclast differentiation
by inhibiting superoxide and NFATc1 downregulation in RAW 264.7
cells. Biomed Rep. 3:483–490. 2015.PubMed/NCBI
|
|
29
|
Domino EF: History of modern
psychopharmacology: A personal view with an emphasis on
antidepressants. Psychosom Med. 61:591–598. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Page ME, Detke MJ, Dalvi A, Kirby LG and
Lucki I: Serotonergic mediation of the effects of fluoxetine, but
not desipramine, in the rat forced swimming test.
Psychopharmacology (Berl). 147:162–167. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kala M and Nivsarkar M: Role of cortisol
and superoxide dismutase in psychological stress induced
anovulation. Gen Comp Endocrinol. 225:117–124. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Miyata S, Hirano S, Ohsawa M and Kamei J:
Chlorpheniramine exerts anxiolytic-like effects and activates
prefrontal 5-HT systems in mice. Psychopharmacol. 213:441–452.
2011. View Article : Google Scholar
|
|
33
|
Biala G, Pekala K, Boguszewska-Czubara A,
Michalak A, Kruk-Slomka M and Budzynska B: Behavioral and
biochemical interaction between nicotine and chronic unpredictable
mild stress in mice. Mol Neurobiol. 54:904–921. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Schmidt AJ, Heiser P, Hemmeter UM, Krieg
JC and Vedder H: Effects of antidepressants on mRNA levels of
antioxidant enzymes in human monocytic U-937 cells. Prog
Neuropsychopharmacol Biol Psychiatry. 32:1567–1573. 2008.
View Article : Google Scholar : PubMed/NCBI
|