Introduction
Acute kidney injury (AKI) is a common clinical
syndrome that is characterized by the rapid loss of kidney function
and abrupt kidney damage. It is associated with high morbidity and
mortality worldwide (1). Acute tubular
necrosis (ATN) is the most common and severe pathological
manifestation of AKI. Although necrosis has long been considered to
be unregulated, previous studies have revealed various types of
regulated necrosis that are independent of caspase activities
(2). Necroptosis is an important type
of regulated necrosis and is involved in various pathological
conditions (3–7). Receptor-interacting protein kinase
(RIP)-1 receives signals from various death stimuli, and
subsequently recruits RIP-3 via RIP homotypic interaction motif
domain-mediated interactions and promotes RIP-3 phosphorylation
(8,9).
Phosphorylated RIP-3 mediates the phosphorylation of mixed-lineage
kinase domain-like (MLKL), ultimately leading to rupture of the
plasma membrane (7). Previous studies
have identified necroptosis in ATN induced by various stimuli,
including ischemia/reperfusion, contrast medium and cisplatin
(10–12). RIP-1 inhibition by necrostatin-1
(Nec-1), or knockout of RIP-3 or MLKL all prevent ATN (10–12).
Reactive oxygen species (ROS) promote necroptosis in
cardiomyocytes, liver cells, Jurkat cells and lung adenoma cells
(13–16). However, the effect of ROS on the
necroptosis of renal tubular epithelium remains unknown. The
present study hypothesized that excessive ROS production may
promote necroptosis in HK-2 human kidney cells. Diphenyleneiodonium
(DPI), an NADPH oxidase inhibitor, has been demonstrated to reduce
ROS production and exert a protective role in numerous cell types
(17–19). The present study investigated the
protective effect of DPI in a renal tubular epithelium necroptosis
model, which was established in our previous study (20).
Materials and methods
Cell culture
The HK-2 human kidney proximal tubular cell line
(American Type Culture Collection, Rockville, MD, USA) was
maintained in a mixture of Gibco Ham's F12 and Dulbecco's modified
Eagle's medium (DMEM/F12; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 10% fetal bovine serum (FBS) and 1%
penicillin/streptomycin (Gibco; Thermo Fisher Scientific, Inc.) in
a humidified atmosphere of 5% CO2 at 37°C. The cells
were seeded at an appropriate cell density for different assays and
allowed to grow to 80% confluence. Cell synchronization was
routinely performed by incubating cells in serum-free medium at
37°C for 24 h prior to each experiment. Finally, the cells were
exposed to different experimental conditions.
Cell treatment
Cells were divided into four groups as follows: i)
Control group, cells were maintained in DMEM/F12 supplemented with
10% FBS for 2 h; ii) tumor necrosis factor-α (TNF-α),
benzyloxycarbonyl-Val-Ala-Asp-fluoromethylketone (z-VAD-fmk) and
antimycin A group [TZA group; HK-2 cells were treated with 10 ng/ml
TNF-α (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and 50 µM
z-VAD-fmk (Sigma-Aldrich; Merck KGaA) for 2 h, and with 10 µM
antimycin A (Santa Cruz Biotechnology, Inc., Dallas, TX, USA) for 1
h], as described in our previous study (20); iii) TNF-α, z-VAD-fmk, antimycin A and
Nec-1 group [TZA + Nec-1 group; cells were pretreated with 50 µM of
Nec-1 (Sigma-Aldrich; Merck KGaA) for 6 h, and continuously treated
with 10 ng/ml TNF-α and 50 µM z-VAD-fmk for 2 h, and with 10 μM
antimycin A for 1 h]; and iv) TNF-α, z-VAD-fmk, antimycin A and DPI
group [TZA + DPI group; cells were pretreated with 500 µM of DPI
(Santa Cruz Biotechnology, Inc.) for 6 h, and continuously treated
with 10 ng/ml TNF-α and 50 µM z-VAD-fmk for 2 h, and with 10 µM
antimycin A for 1 h].
ROS measurements
ROS content was quantified using an ROS assay kit
(cat. no. S0033; Beyotime Institute of Biotechnology, Shanghai,
China). The cells were harvested using trypsin (cat. no. 25300054;
Thermo Fisher Scientific, Inc.) and washed with PBS. The harvested
cells were exposed to 10 µM 2′,7′-dichlorodihydrofluorescein
diacetate (DCFH-DA) for 20 min at 37°C, and the labeled cells were
washed twice in PBS and subjected to flow cytometry (Beckman
Coulter, Inc., Brea, CA, USA) for ROS detection using the 488 nm
laser for excitation and detected at 535 nm. The ROS level was
represented by the median fluorescence intensity.
Western blot analysis
HK-2 cells subjected to the different experimental
conditions were lysed in radioimmunoprecipitation assay lysis
buffer (cat. no. P0013B; Beyotime Institute of Biotechnology). The
total protein concentration from the resultant supernatant was
determined using a Pierce™ BCA Protein Assay kit (Thermo Fisher
Scientific, Inc.). Samples of equal protein content (35 µg/lane)
were separated by SDS-PAGE using 8% gels (130 V for 90 min) and
transferred to polyvinylidene difluoride membranes (90 V for 60
min). Membranes were blocked with 3% bovine serum albumin in
TBS-Tween-20 (cat. no. ST825; Beyotime Institute of Biotechnology)
and incubated overnight at 4°C with antibodies against RIP-3
phospho-S227 (cat. no. ab209384 at 1:2,000; Abcam, Shanghai, China)
and MLKL phospho-S358 (cat. no. ab187091 at 1:1,000; Abcam).
β-actin (cat. no. 8457; 1:1,000; Cell Signaling Technology, Inc.,
Danvers, MA, USA) was used as loading control. After being washed
in TBS, blots were incubated with horseradish peroxidase-conjugated
goat anti-rabbit antibodies (cat. no. 7077; 1:2,000; Cell Signaling
Technology, Inc.) for 1 h at room temperature. After a final wash
as described above, the corresponding secondary antibodies were
visualized using enhanced Pierce™ ECL Western Blotting Substrate
(Thermo Fisher Scientific, Inc.). Signals were quantified with a
chemiluminescence detector (Tanon 4200; Tanon Science &
Technology Co., Ltd., Shanghai, China) and the accompanying
densitometry software.
Annexin V-fluorescein isothiocyanate
(FITC) and propidium iodide (PI) staining
Annexin V and PI staining were performed using an
Annexin V-FITC Apoptosis Detection kit (cat. no. PF032; EMD
Millipore, Billerica, MA, USA). Cells were trypsinized, washed
twice with ice-cold PBS and resuspended in 1X binding buffer The
cell concentration was adjusted to 1×106/ml, and 400 µl
cell suspensions were added to 5-ml centrifuge tubes, followed by
the addition of 1.25 µl Annexin V-FITC. Cells were incubated for 30
min at room temperature and centrifuged at 1,000 × g for 3 min at
room temperature. Following resuspension in 400 µl 1X binding
buffer, 5 ml PI was added to each cell suspension and incubated for
5 min in the dark at 2–8°C. The rates of necrosis and apoptosis of
the HK-2 cells were analyzed by flow cytometry (Beckman Coulter,
Inc., Brea, CA, USA).
Statistical analysis
All values are expressed as the mean ± standard
deviation. Multiple comparisons among the groups were conducted by
one-way ANOVA followed by a least significant difference multiple
comparison test. All statistical analyses were performed using SPSS
software (version 21.0; IBM SPSS, Armonk, NY, USA), with P<0.05
considered to indicate a statistically significant difference.
Results
Construction and validation of the
necroptosis model in HK-2 cells
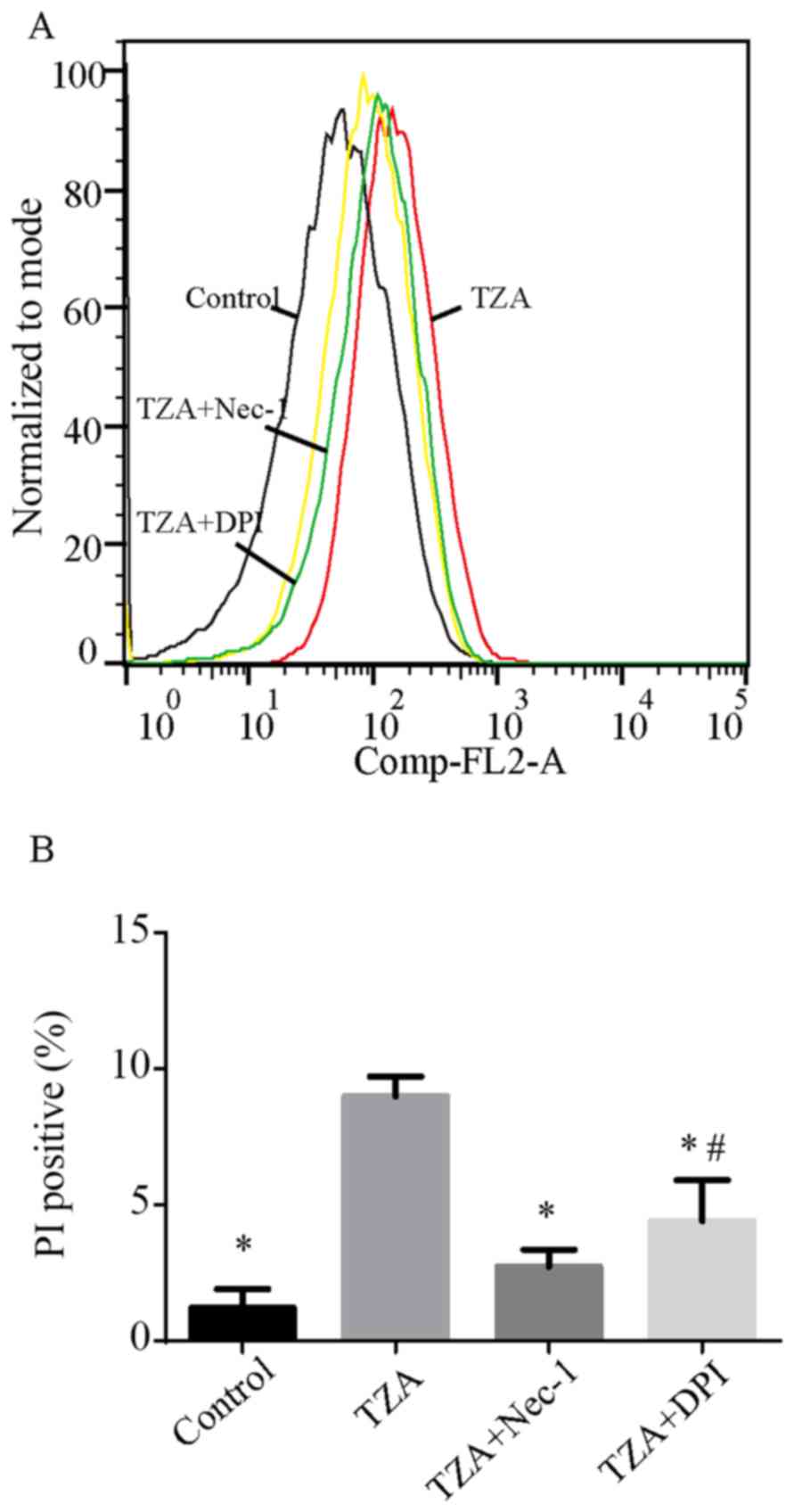
Following the treatment of HK-2 cells with TNF-α,
zVAD-fmK and antimycin A, the percentages of PI-positive cells
increased from 1.22±0.69 to 8.98±0.73% (P<0.001; Fig. 1). However, TZA treatment did not
increase the percentage of Annexin V-positive cells (control group,
0.87±0.15% vs. TZA group, 0.89±0.13%; P=0.098). Administration of
Nec-1 was observed to reduce the rate of TZA-induced necrosis
markedly (PI-positive cells, 2.74±0.62%; P<0.001 vs. TZA group).
This result indicates that TZA-induced necrosis may be regulated by
Nec-1.
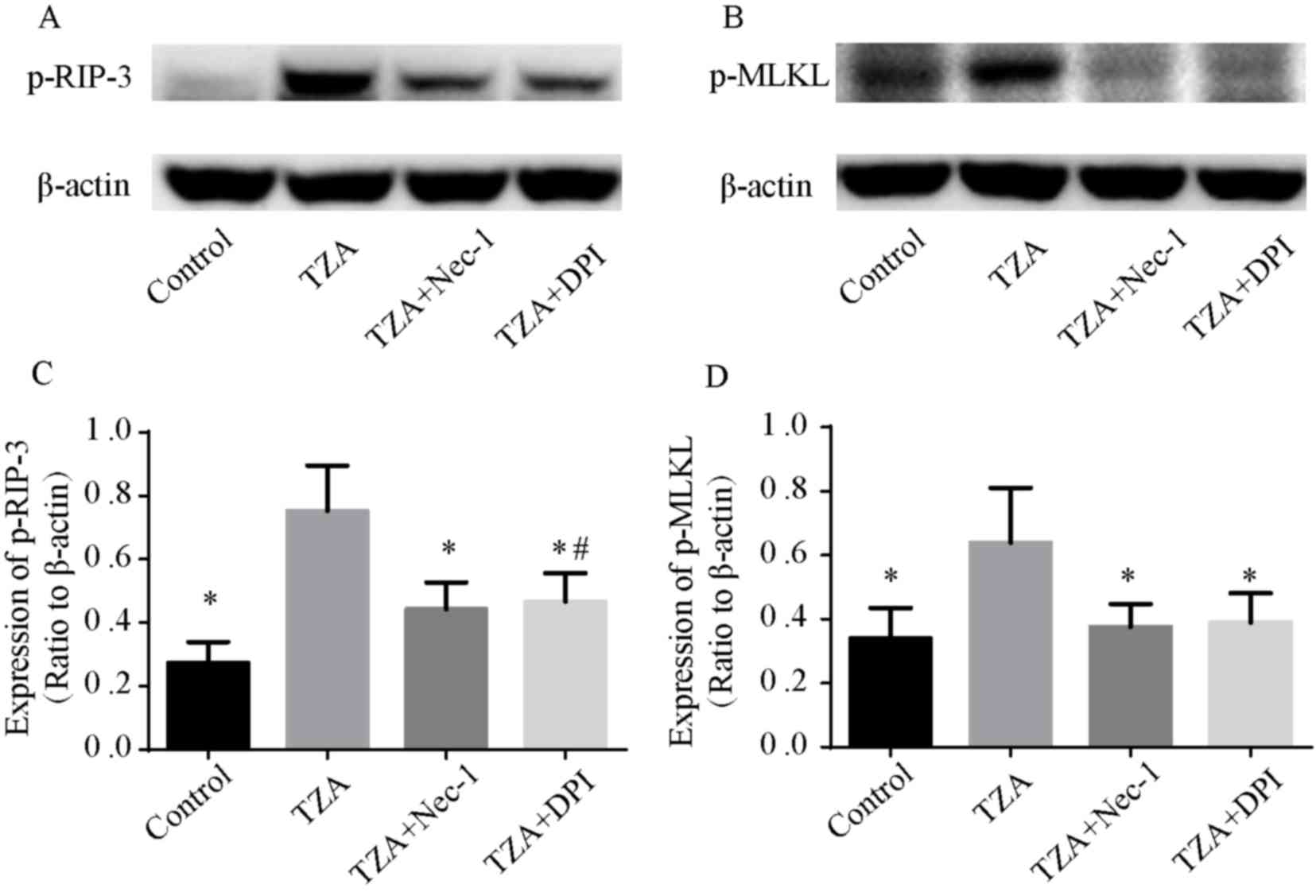
As RIP-3 and MLKL are essential for the necroptosis
pathway, the phosphorylation of RIP-3 (p-RIP-3) and MLKL (p-MLKL)
were assessed by western blotting to confirm that the observed cell
death was by necroptosis. p-RIP-3 and p-MLKL expression levels were
elevated in the TZA group, and Nec-1 reversed the increased p-RIP-3
and p-MLKL expression levels (Fig. 2).
These results indicate that the cell death induced by TZA is
necroptosis.
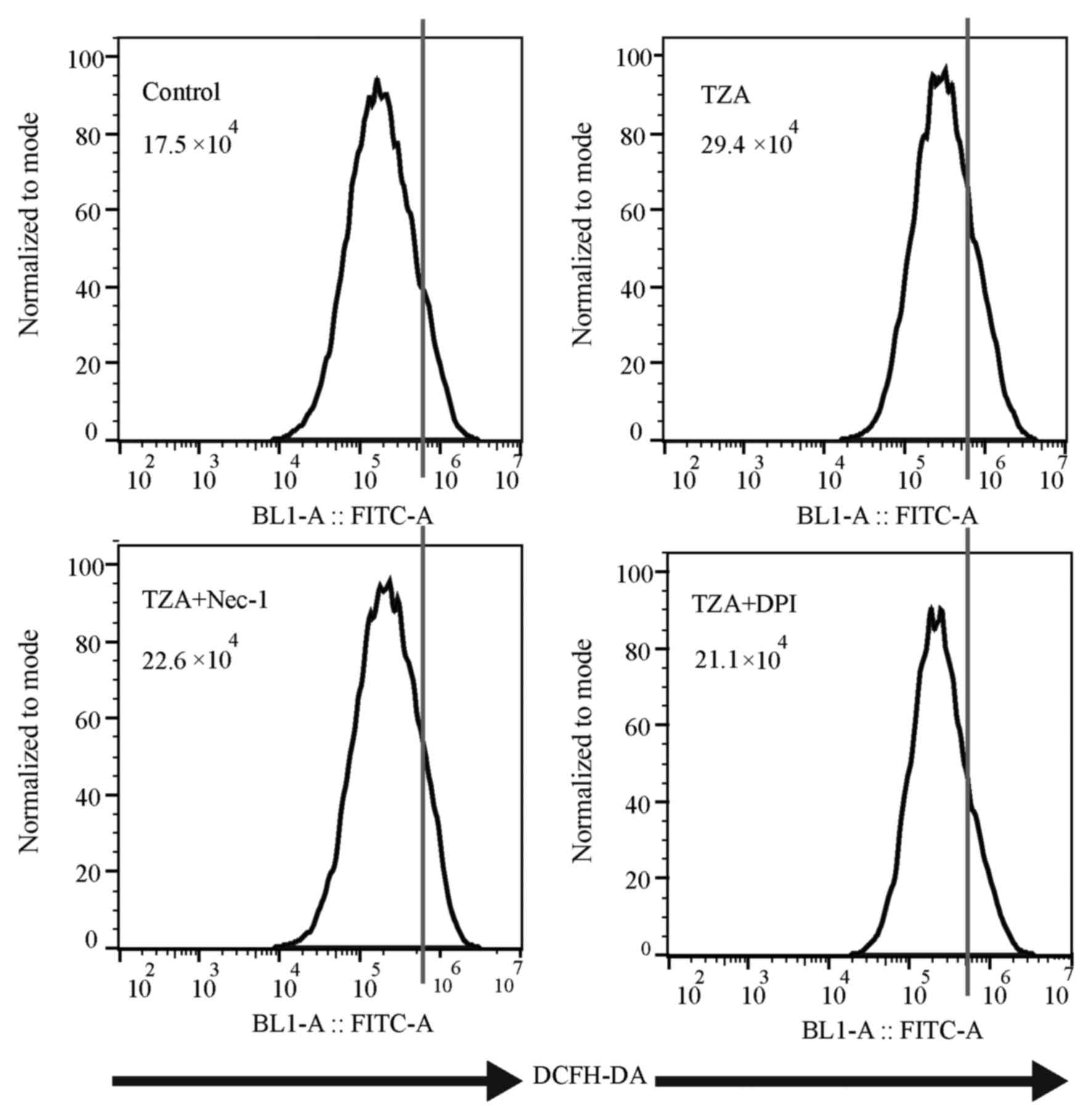
ROS production is augmented in
TZA-induced HK-2 necroptosis
Total ROS production, as measured by DCFH-DA
fluorescence intensity (Fig. 3), was
increased in the TZA group compared with the control group
(29.4×104 vs. 17.5×104). Additionally, the
increase in ROS generation could be reversed by Nec-1 treatment
(22.6×104). Thus, it was concluded that ROS production
increases in HK-2 cells undergoing TZA-induced necroptosis.
NADPH oxidase inhibitor, DPI protects
against necroptosis via inhibition of ROS production
To determine the effect of the NADPH oxidase
inhibitor, DPI on necroptosis in HK-2 cells, DPI was administered
to the TZA-induced necroptosis cell model. DPI inhibited ROS
generation in the TZA-induced necroptosis (TZA + DPI group,
21.1×104 vs. TZA group, 29.4×104; Fig. 3) and ameliorated TZA-induced necrosis
(PI-positive cells: TZA + DPI group, 4.40±1.51% vs. TZA group,
8.98±0.73%; P<0.001; Fig. 1). In
addition, p-RIP-3 and p-MLKL levels were decreased by DPI
pre-treatment (Fig. 2). These results
indicate that the NADPH oxidase inhibitor, DPI may reduce ROS
production in HK-2 cells and prevent HK-2 cell necroptosis.
Discussion
AKI caused by ATN is a common clinical syndrome with
high rates of morbidity and mortality (1). Although necrosis used to be considered as
an unregulated form of cell death, necroptosis, a type of regulated
necrosis, has been identified in ATN, as well as in drug-induced
liver injury and ischemic brain injury (10–12).
Mechanistically, RIP-1 phosphorylates RIP-3 and forms a necrosome
(8,9).
Subsequently, the necrosome phosphorylates MLKL, resulting in
necroptosis in various types of cells (7,21).
Inhibition of RIP-1 by Nec-1 and knockout of RIP-3 or MLKL prevents
ATN in response to various stimuli, such as cisplatin, sepsis and
ischemia/reperfusion injury (10–12).
ROS are chemically reactive species containing
oxygen, including peroxides, superoxides, hydroxyl radicals and
singlet oxygen. Although ROS are formed as a natural byproduct of
the normal metabolism of oxygen, and have important roles in cell
signaling and homeostasis in biological conditions, excessive ROS
are detrimental. Various studies have demonstrated that excessive
ROS promotes necroptosis in various cell types, whereas inhibiting
ROS production reduces necroptosis (13–16). In
addition, previous studies revealed that ROS production is
augmented in AKI, and that ROS lead to renal tubular epithelium
injury via inflammasome activation, mitochondrion damage and
tubular epithelium apoptosis (22–25). To the
best of our knowledge, there are no studies that have investigated
the effect of ROS on necroptosis in renal tubular epithelium. The
current study hypothesized that excessive ROS may promote
necroptosis in HK-2 cells, and that DPI may reduce ROS production
and thus exert a protective role against necroptosis in HK-2
cells.
A necroptosis model in HK-2 cells, established in
our previous study, was validated in the present study, and
revealed that ROS production increased in HK-2 cells undergoing
necroptosis induced by TZA. Nec-1, a necroptosis inhibitor
decreased ROS production in necroptosis. Similarly, pre-treatment
of HK-2 cells with DPI, an NADPH oxidase inhibitor, decreased ROS
generation, and ameliorated TZA-induced necroptosis, and attenuated
p-RIP-3 and p-MLKL expression levels.
In conclusion, the results of the current study
indicate that excessive ROS levels augment HK-2 cell necroptosis
and that DPI may protect HK-2 cells against necroptosis by
inhibiting ROS generation. Currently, the mechanism underlying
precisely how ROS participates in necroptosis remains unclear. The
molecular mechanism of ROS-mediated necroptosis was not
investigated in the present study and further investigations of
this in the renal tubular epithelium are required. Further research
may provide a novel therapeutic strategy for the treatment of
AKI.
Acknowledgements
The present study was supported by the National
Natural Science Foundation (grant no. 81570609), the Natural
Science Foundation of Guangdong Province (grant no. 2014A030313545)
and the National Clinical Key Specialty Construction Preparatory
Projects.
References
|
1
|
Susantitaphong P, Cruz DN, Cerda J,
Abulfaraj M, Alqahtani F, Koulouridis I and Jaber BL: Acute Kidney
Injury Advisory Group of the American Society of Nephrology: World
incidence of AKI: A meta-analysis. Clin J Am Soc Nephrol.
8:1482–1493. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Galluzzi L, Vitale I, Abrams JM, Alnemri
ES, Baehrecke EH, Blagosklonny MV, Dawson TM, Dawson VL, El-Deiry
WS, Fulda S, et al: Molecular definitions of cell death
subroutines: Recommendations of the Nomenclature Committee on Cell
Death 2012. Cell Death Differ. 19:107–120. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Oerlemans MI, Liu J, Arslan F, den Ouden
K, van Middelaar BJ, Doevendans PA and Sluijter JP: Inhibition of
RIP1-dependent necrosis prevents adverse cardiac remodeling after
myocardial ischemia-reperfusion in vivo. Basic Res Cardiol.
107:2702012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Degterev A, Huang Z, Boyce M, Li Y, Jagtap
P, Mizushima N, Cuny GD, Mitchison TJ, Moskowitz MA and Yuan J:
Chemical inhibitor of nonapoptotic cell death with therapeutic
potential for ischemic brain injury. Nat Chem Biol. 1:112–119.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bonnet MC, Preukschat D, Welz PS, van Loo
G, Ermolaeva MA, Bloch W, Haase I and Pasparakis M: The adaptor
protein FADD protects epidermal keratinocytes from necroptosis in
vivo and prevents skin inflammation. Immunity. 35:572–582. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Günther C, Martini E, Wittkopf N, Amann K,
Weigmann B, Neumann H, Waldner MJ, Hedrick SM, Tenzer S, Neurath
MF, et al: Caspase-8 regulates TNF-α-induced epithelial necroptosis
and terminal ileitis. Nature. 477:335–339. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang H, Sun L, Su L, Rizo J, Liu L, Wang
LF, Wang FS and Wang X: Mixed lineage kinase domain-like protein
MLKL causes necrotic membrane disruption upon phosphorylation by
RIP3. Mol Cell. 54:133–146. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang DW, Shao J, Lin J, Zhang N, Lu BJ,
Lin SC, Dong MQ and Han J: RIP3, an energy metabolism regulator
that switches TNF-induced cell death from apoptosis to necrosis.
Science. 325:332–336. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He S, Wang L, Miao L, Wang T, Du F, Zhao L
and Wang X: Receptor interacting protein kinase-3 determines
cellular necrotic response to TNF-alpha. Cell. 137:1100–1111. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Linkermann A, Bräsen JH, Himmerkus N, Liu
S, Huber TB, Kunzendorf U and Krautwald S: Rip1
(receptor-interacting protein kinase 1) mediates necroptosis and
contributes to renal ischemia/reperfusion injury. Kidney Int.
81:751–761. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Linkermann A, Heller JO, Prókai A,
Weinberg JM, De Zen F, Himmerkus N, Szabó AJ, Bräsen JH, Kunzendorf
U and Krautwald S: The RIP1-kinase inhibitor necrostatin-1 prevents
osmotic nephrosis and contrast-induced AKI in mice. J Am Soc
Nephrol. 24:1545–1557. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu Y, Ma H, Shao J, Wu J, Zhou L, Zhang Z,
Wang Y, Huang Z, Ren J, Liu S, et al: A role for tubular
necroptosis in cisplatin-induced AKI. J Am Soc Nephrol.
26:2647–2658. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Takemoto K, Hatano E, Iwaisako K, Takeiri
M, Noma N, Ohmae S, Toriguchi K, Tanabe K, Tanaka H, Seo S, et al:
Necrostatin-1 protects against reactive oxygen species
(ROS)-induced hepatotoxicity in acetaminophen-induced acute liver
failure. FEBS Open Bio. 4:777–787. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Song KJ, Jang YS, Lee YA, Kim KA, Lee SK
and Shin MH: Reactive oxygen species-dependent necroptosis in
Jurkat T cells induced by pathogenic free-living Naegleria fowleri.
Parasite Immunol. 33:390–400. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang Y, Shan S, Chi L, Zhang G, Gao X, Li
H, Zhu X and Yang J: Methyl methanesulfonate induces necroptosis in
human lung adenoma A549 cells through the PIG-3-reactive oxygen
species pathway. Tumour Biol. 37:3785–3795. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang T, Zhang Y, Cui M, Jin L, Wang Y, Lv
F, Liu Y, Zheng W, Shang H, Zhang J, et al: CaMKII is a RIP3
substrate mediating ischemia- and oxidative stress-induced
myocardial necroptosis. Nat Med. 22:175–182. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Farrow MA, Chumbler NM, Lapierre LA,
Franklin JL, Rutherford SA, Goldenring JR and Lacy DB: Clostridium
difficile toxin B-induced necrosis is mediated by the host
epithelial cell NADPH oxidase complex. Proc Natl Acad Sci USA.
110:18674–18679. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ostergaard M, Christensen M, Nilsson L,
Carlsen I, Frøkiær J and Nørregaard R: ROS dependence of
cyclooxygenase-2 induction in rats subjected to unilateral ureteral
obstruction. Am J Physiol Renal Physiol. 306:F259–F270. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Das R, Xu S, Quan X, Nguyen TT, Kong ID,
Chung CH, Lee EY, Cha SK and Park KS: Upregulation of mitochondrial
Nox4 mediates TGF-β-induced apoptosis in cultured mouse podocytes.
Am J Physiol Renal Physiol. 306:F155–F167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liang X, Chen Y, Zhang L, Jiang F, Wang W,
Ye Z, Liu S, Yu C and Shi W: Necroptosis, a novel form of
caspase-independent cell death, contributes to renal epithelial
cell damage in an ATP-depleted renal ischemia model. Mol Med Rep.
10:719–724. 2014.PubMed/NCBI
|
|
21
|
Moujalled DM, Cook WD, Murphy JM and Vaux
DL: Necroptosis induced by RIPK3 requires MLKL but not Drp1. Cell
Death Dis. 5:e10862014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Quintavalle C, Brenca M, De Micco F, Fiore
D, Romano S, Romano MF, Apone F, Bianco A, Zabatta MA, Troncone G,
et al: In vivo and in vitro assessment of pathways involved in
contrast media-induced renal cells apoptosis. Cell Death Dis.
2:e1552011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Winterberg PD, Wang Y, Lin KM, Hartono JR,
Nagami GT, Zhou XJ, Shelton JM, Richardson JA and Lu CY: Reactive
oxygen species and IRF1 stimulate IFNα production by proximal
tubules during ischemic AKI. Am J Physiol Renal Physiol.
305:F164–F172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ahn JM, You SJ, Lee YM, Oh SW, Ahn SY, Kim
S, Chin HJ, Chae DW and Na KY: Hypoxia-inducible factor activation
protects the kidney from gentamicin-induced acute injury. PLoS One.
7:e489522012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao WY, Zhang L, Sui MX, Zhu YH and Zeng
L: Protective effects of sirtuin 3 in a murine model of
sepsis-induced acute.
|