Introduction
Since human dental pulp stem cells (DPSCs) were
first isolated by Gronthos et al (1,2), these
cells have been widely utilized in regenerative medicine and tissue
engineering applications (3,4), mainly due to their multilineage
differentiation plasticity and high proliferative capacity
(5). Of particular interest are the
potential therapeutic applications of DPSCs in various
neurodegenerative diseases and neural disorders, due to the high
innate propensity of these cells to differentiate into the neural
lineage (6) as a consequence of their
origin from the embryonic neural crest (7).
To date, transplanted DPSCs have demonstrated
promise in the treatment of various neural lesions in animal models
of stroke (8), spinal cord injury
(9), facial nerve injury (10) and optical nerve injury (11), among others. Nevertheless, it must be
noted that these animal studies utilized undifferentiated DPSCs,
and it is probable that some degree of neural induction or neural
lineage commitment of DPSCs in vitro, prior to
transplantation in vivo, could further enhance the
neuroregenerative potential of these adult stem cells (6). This may be facilitated by an improved
understanding of the mechanisms of neurogenesis of DPSCs.
The signalling pathways mediated by the family of
EphB receptors and EphrinB ligands, which are expressed in the
perivascular niche of the dental pulp, are known to serve key roles
in the proliferation, mobilization and mineralization of DPSCs
during the process of tooth regeneration, in response to injury of
the dentin matrix (12). Of
particular interest in the field of neuroregeneration are EphrinB2
and its cognate receptors EphB2 and EphB4, which have been
demonstrated to serve key developmental roles in embryonic
neurogenesis (13–15) as well as in the homeostasis and
mobilization of neural stem cells (16) and neural progenitors (17). EphrinB2 and its cognate receptors
EphB2 and EphB4 are transmembrane receptor tyrosine kinases, which
are established to play crucial roles in cell migration and in the
determination of cell lineage fate (18). Simultaneous bidirectional signals are
generated by the binding interactions of EphrinB2 with its cognate
receptors EphB2 and EphB4, with forward signalling being effected
through EphB2 and EphB4 and reverse signalling being effected
through EphrinB2 (19). Direct
intercellular contact is required to initiate these signalling
pathways, as EphrinB2, EphB2 and EphB4 are all membrane-bound
protein receptors. In a previous study by our group, it was
demonstrated that the EphrinB2 signalling pathway served a key role
in the osteogenic/odontogenic differentiation of DPSCs (20).
At present, the role of EphrinB2 signalling in the
neurogenesis of DPSCs is not clear. Therefore, the present study
aimed to investigate and delineate the roles of EphrinB2 forward
and reverse signalling in the neurogenesis of DPSCs in
vitro. Exogenous recombinant EphrinB2-Fc and EphB4-Fc were
utilized together with specific peptide inhibitors of EphB2 and
EphB4 signalling (SNEW and TNYL-RAW, respectively) (21) as well as transgenic DPSCs
overexpressing the EphrinB2 ligand. The peptide inhibitors SNEW and
TNYL-RAW were previously demonstrated by Chrencik et al
(21) to bind specifically to the
EphB2 and EphB4 receptors respectively, and block the interaction
of these receptors with the EphrinB2 ligand. Both forward and
reverse signalling, transduced either by EphrinB2-EphB4 interaction
(TNYL-RAW) or EphrinB2-EphB2 interaction (SNEW), would thus be
inhibited. In the case of transgenic expression of EphrinB2, this
may stimulate both forward and reverse signalling in DPSCs.
Materials and methods
Chemical reagents, culture media and
supplements
Unless otherwise stated, all chemical reagents were
obtained from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany), all
culture media and supplements were obtained from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA), and all plastic labware
consumables were obtained from BD Biosciences (Franklin Lakes, NJ,
USA).
Isolation, characterization and
culture of DPSCs
The DPSCs utilized in the current study were
isolated from the extracted third molars of young adult patients
below 25 years of age according to previously described procedure
(22). The isolated DPSCs were
characterized by flow cytometric analysis on the expressions of
cluster of differentiation (CD)45, CD73, CD90 and CD105 as well as
by multilineage differentiation assays (osteogenic, adipogenic and
neurogenic induction), as previously described (22). The DPSCs were cultured in T75 culture
flasks with α-minimum essential medium supplemented with 10% (v/v)
foetal bovine serum (FBS) and 1% (v/v) penicillin-streptomycin
antibiotic solution at 37°C in a humidified 5% CO2
incubator. The culture medium was refreshed every 3 to 4 days, and
confluent monolayers were dissociated with 0.5% (w/v) trypsin-EDTA
for serial passage.
Transfection of DPSCs
Recombinant retroviral construct Rsv-NFB2-GFP-Bsd
[human EphrinB2 gene (EFNB2) transgenic lentivirus with green
fluorescent protein-blasticidin dual marker) and the corresponding
empty vector Rsv-Null-Control-GFP-Bsd were obtained from Gentarget,
Inc. (San Diego, CA, USA). Monolayers of DPSCs at 50% confluence
were transduced with either the Rsv-NFB2-GFP-Bsd vector for
transgenic expression of EphrinB2 gene or the empty vector
(Rsv-Null-Control-GFP-Bsd) at a multiplicity of infection of 50 for
a total duration of 24 h within a humidified 5% CO2
incubator at 37°C. Fresh culture medium was then applied, and
stably transduced cells were selected with the addition of 2 µg/ml
Blasticidin (EMD Millipore, Billerica, MA, USA) to the culture
medium. Transduction efficiency was assessed qualitatively under
fluorescence microscopy (Olympus Corporation, Tokyo, Japan).
Neural induction of DPSCs
DPSCs seeded on 6-well culture plates at a density
of 2.5×104 cells/cm2 and were cultured in
neural induction medium composed of Neurobasal-A medium (cat. no.
10888-022; Thermo Fisher Scientific, Inc.) supplemented with 2%
(v/v) B27 supplement (cat. no. 17504-044; Thermo Fisher Scientific,
Inc.), 1% (v/v) penicillin-streptomycin antibiotic solution, 20
ng/ml epidermal growth factor (cat. no. PHG0314; Thermo Fisher
Scientific, Inc.) and 40 ng/ml basic fibroblast growth factor (cat.
no. PHG0024; Thermo Fisher Scientific, Inc.) (23). Depending on the particular experiment,
the neural induction medium was additionally supplemented with
either 2 µg/ml EphrinB2-Fc (cat. no. 7397-EB-05C; R&D Systems,
Minneapolis, MN, USA), 100 µM of specific peptide inhibitor for
EphB4 or EphB2 (TNYL-RAW or SNEW, respectively), 100 µM of the
corresponding control scrambled peptides (SCR-WTL or SCR-EPQ,
respectively), or 2 µg/ml EphB4-Fc (cat. no. 446-B4-200; R&D
Systems). The TNYL-RAW peptide (YNYLFSPNGPIARAW), SNEW peptide
(SNEWIQPRLPQH), and corresponding control scrambled peptides
SCR-EPQ (EPQNHSWPIRQL) and SCR-WTL (WTLAIFARNYNGPSP), respectively
(21), were custom-synthesized by
Thermo Fisher Scientific, Inc. The EphrinB2-Fc and EphB4-Fc were
clustered with goat anti-human Fc antibody (cat. no. H10000; Thermo
Fisher Scientific, Inc.) at a molar ratio of 1:5 for 30 min at 37°C
prior to being added into the culture medium (24). It is established that the EphrinB2
ligand naturally occurs as a multimer in its native membrane-bound
form, and that monomeric soluble EphrinB2 ligand is unable to
elicit forward signalling upon binding to either the EphB2 or EphB4
receptor (24). As such, clustering
of the soluble recombinant EphrinB2-Fc with anti-Fc antibodies is
required to mimic its naturally-occurring multimeric membrane-bound
state, to in turn enable transduction of forward signalling
(24). Neural induction was performed
for a duration of 7 days at 37°C within a humidified 5%
CO2 incubator, after which the samples were collected
for either reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) analysis or immunocytochemistry.
RT-qPCR analysis of neural marker
expression by DPSCs
Total RNA of the cultured DPSCs was extracted with
an RNeasy Plus Mini kit (Qiagen, Inc., Valencia, CA, USA) and
subjected to RT with a SuperScript VILO Master Mix (Thermo Fisher
Scientific, Inc.). RT-qPCR analysis was performed with a StepOne
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.) utilizing SYBR Premix Ex Taq II (cat. no. RR820A; Takara Bio,
Inc., Otsu, Japan). The primer sequences of neural markers
[βIII-tubulin, microtubule-associated protein 2 (MAP2), Musashi1,
neural cell adhesion molecule (NCAM), nestin, neurofilament medium
polypeptide (NFM), neurogenin 2 (NGN2) and neuron-specific enolase
(NSE)] utilized for the RT-qPCR analysis were identical to those of
our previous study (23), and GAPDH
was used as the endogenous reference control gene. The primer
sequences used for EphrinB2, EphB2, EphB4 and GAPDH were as
follows: EphrinB2 forward, 5′-CCTCTCCTCAACTGTGCCAAA-3′ and reverse,
5′-CCCAGAGGTTAGGGCTGAATT-3′; EphB2 forward,
5′-ATGAACACGATCCGCACGTA-3′ and reverse,
5′-TTGGTCCGTAGCCAGTTGTTCT-3′; EphB4 forward,
5′-CCTTCCTGCGGCTAAACGAC-3′ and reverse, 5′-GTTGACTAGGATGTTGCGAG-3′;
and GAPDH forward, 5′-GGCATGGACTGTGGTCATGAG-3′ and reverse,
5′-TGCACCACCAACTGCTTAGC-3′. The following amplification parameters
were utilized for the RT-qPCR analysis: 2 min at 50°C, 20 sec at
95°C, and 40 cycles of 3 sec at 95°C followed by 30 sec at 60°C.
The 2−ΔΔCq method (25)
was used to compute the relative cycle threshold (Cq) values for
each gene, which were then normalized against the endogenous gene
expression of GAPDH. The fold-differences in gene expression by the
DPSCs that were subjected to neural induction in the various
experimental groups were normalized with respect to the DPSCs in
the corresponding control groups on day 7. Overall, three
experimental replicates were performed for each gene analysed by
RT-qPCR.
Immunocytochemistry for the detection
of βIII-tubulin and NGN2 expression by DPSCs
After 7 days of neural induction in the presence or
absence of 2 µg/ml EphrinB2-Fc, the DPSCs were fixed at room
temperature with 4% (v/v) paraformaldehyde for 20 min, followed by
permeabilization with 0.1% (w/v) Triton X-100 for 10 min, and
blocking in PBS with 10% (v/v) FBS for 2 h at room temperature. The
fixed DPSCs were then incubated with primary antibodies (1:200
dilution) specific for mouse anti-βIII-tubulin (cat. no. G712A;
Promega Corporation, Madison, WI, USA) or rabbit ant-NGN2 (cat. no.
sc-50401; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) for 2 h
at room temperature. Excess primary antibodies were removed by
washing in 1% (w/v) bovine serum albumin (BSA)/PBS, and the samples
were incubated with the corresponding secondary antibodies (1:200
dilution), goat anti-mouse conjugated to Alexa Fluor 488 (cat. no.
ab150117; Abcam, Cambridge, UK) or goat anti-rabbit conjugated to
tetramethylrhodamine (TRITC; cat. no. ab6718; Abcam), for 2 h in
the dark at room temperature. Following the removal of excess
secondary antibodies by washing in 1% (w/v) BSA/PBS, DAPI was used
to stain the cell nuclei, and the samples were imaged under
fluorescence microscopy under the specific excitation/emission
wavelengths for TRITC (540/570 nm) and Alexa Fluor 488 (490/520
nm).
Statistical analysis
There were triplicate samples of cell cultures for
all experimental and control groups. The data are expressed as the
means ± standard deviation, and statistically significant
differences between the datasets were evaluated with Student's
t-test using the SPSS 19.0 statistics software (IBM Corp., Armonk,
NY, USA). The threshold for statistical significance was set at
P<0.05.
Results
Effect of EphrinB2-Fc on the neural
differentiation of DPSCs
The RT-qPCR analysis of the neural markers (Fig. 1) revealed that supplementation with 2
µg/ml EphrinB2-Fc within the neural induction milieu resulted in
significantly reduced expression of βIII-tubulin, Musashi1, NCAM,
nestin, NFM and NGN2 in the DPSCs (fold expression: 0.39, 0.16,
0.40, 0.38, 0.06 and 0.39, respectively, with respect to control
level expression in the absence of EphrinB2-Fc; P<0.01 for
Musashi1, Nestin and NFM, P<0.05 for βIII-tubulin, NCAM and
NGN2). However, there was no significant difference in the
expression of MAP2 or NSE between the DPSCs EphrinB2-Fc and those
without.
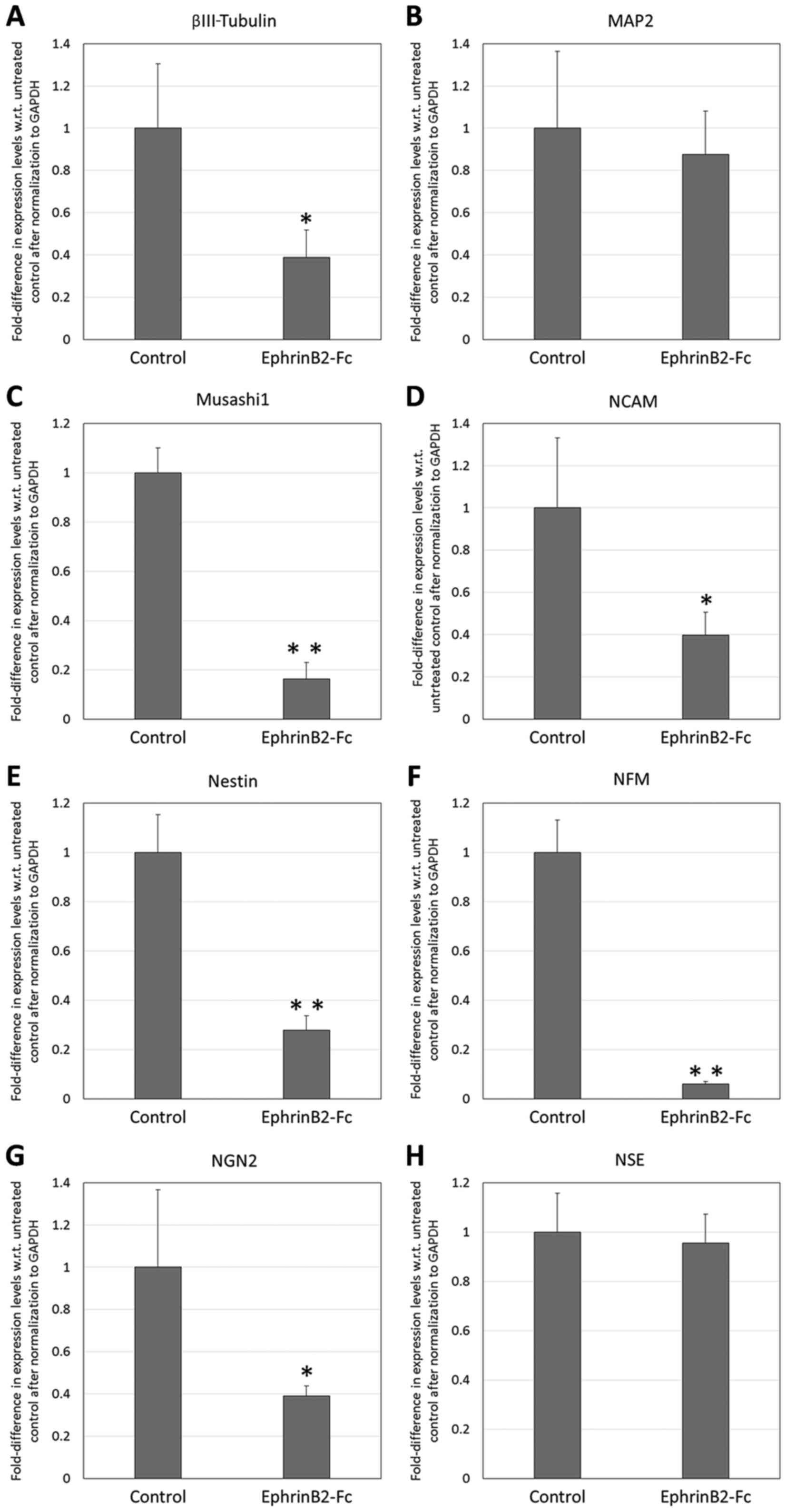 | Figure 1.Reverse transcription-quantitative
polymerase chain reaction analysis of neural marker expression by
dental pulp stem cells following treatment with 2 µg/ml EphrinB2-Fc
in neural induction medium for 7 days. Fold-difference in
expression with respect to untreated controls is shown for (A)
βIII-tubulin, (B) MAP2, (C) Musashi1, (D) NCAM, (E) nestin, (F)
NFM, (G) NGN2 and (H) NSE. *P<0.05 and **P<0.01 vs. Control.
MAP2, microtubule-associated protein 2; NCAM, neural cell adhesion
molecule; NFM, neurofilament medium polypeptide; NGN2, neurogenin
2; NSE, neuron-specific enolase. |
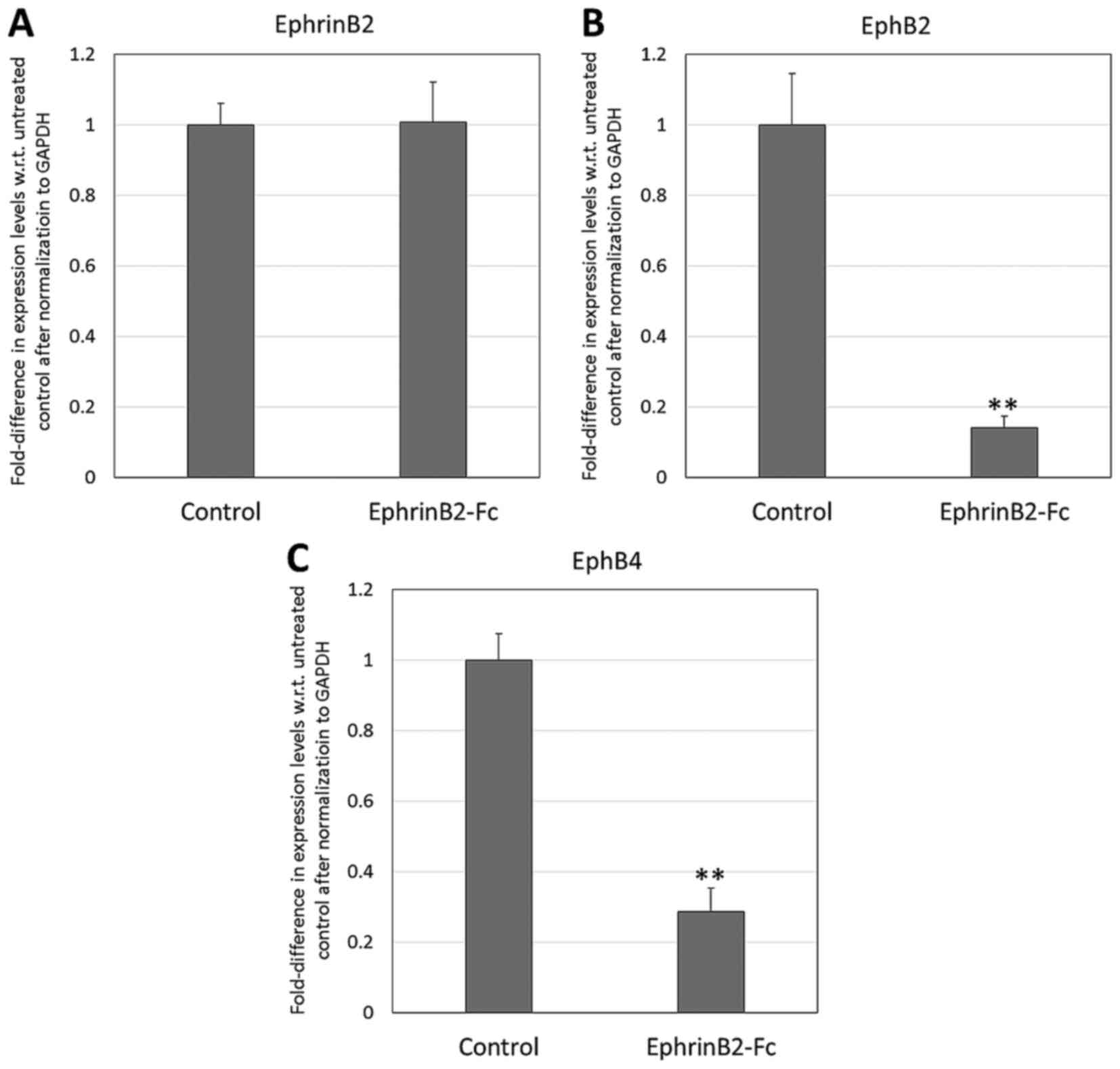
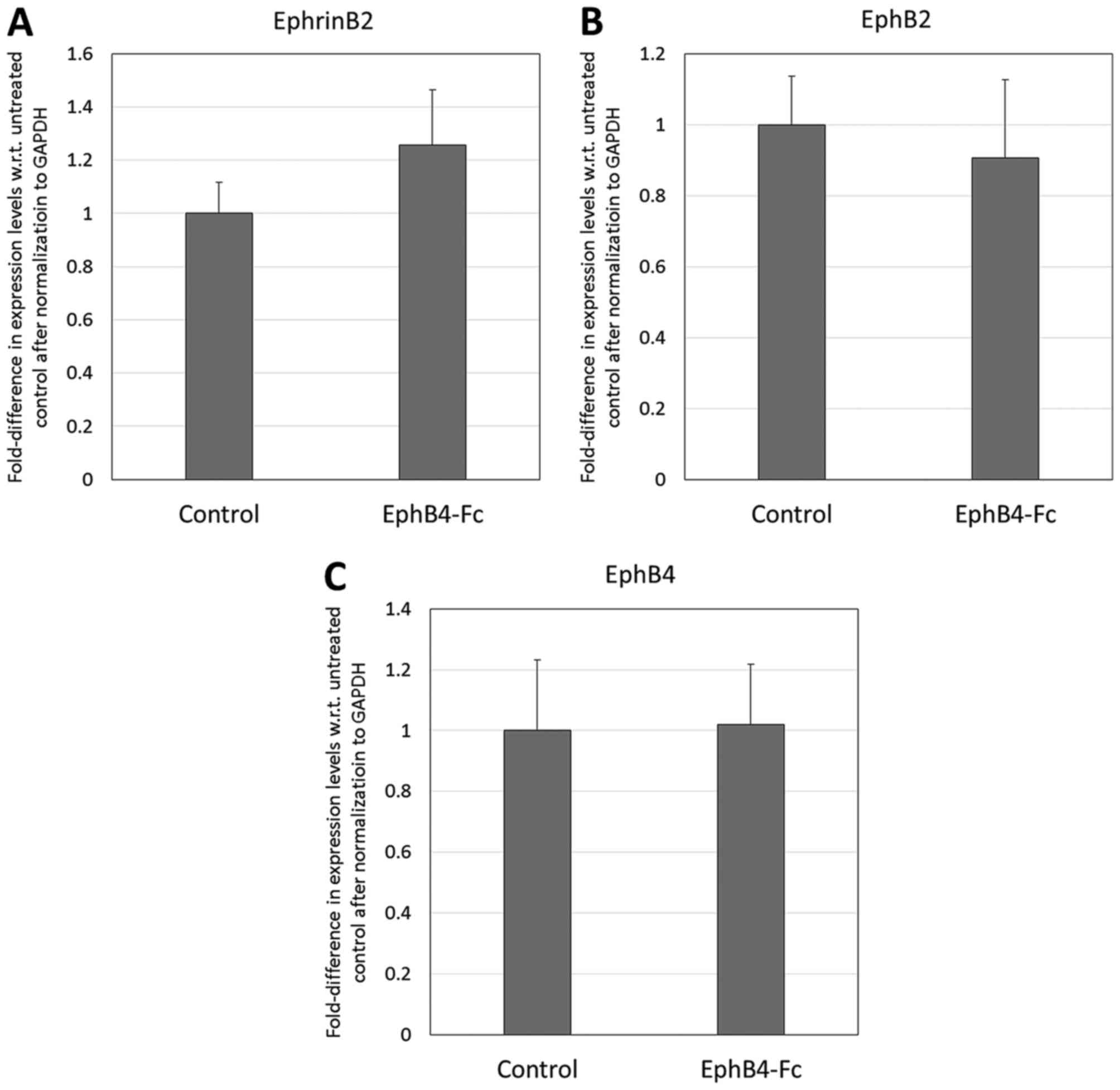
Additionally, as presented in Fig. 2, there was significant downregulation
of EphB2 and EphB4 by the DPSCs that were exposed to EphrinB2-Fc
(fold expression: 0.14 and 0.29, respectively, with respect to
control level expression in the absence of EphrinB2-Fc; P<0.01).
However, there was no significant difference in the expression of
endogenous EphrinB2 by the DPSCs.
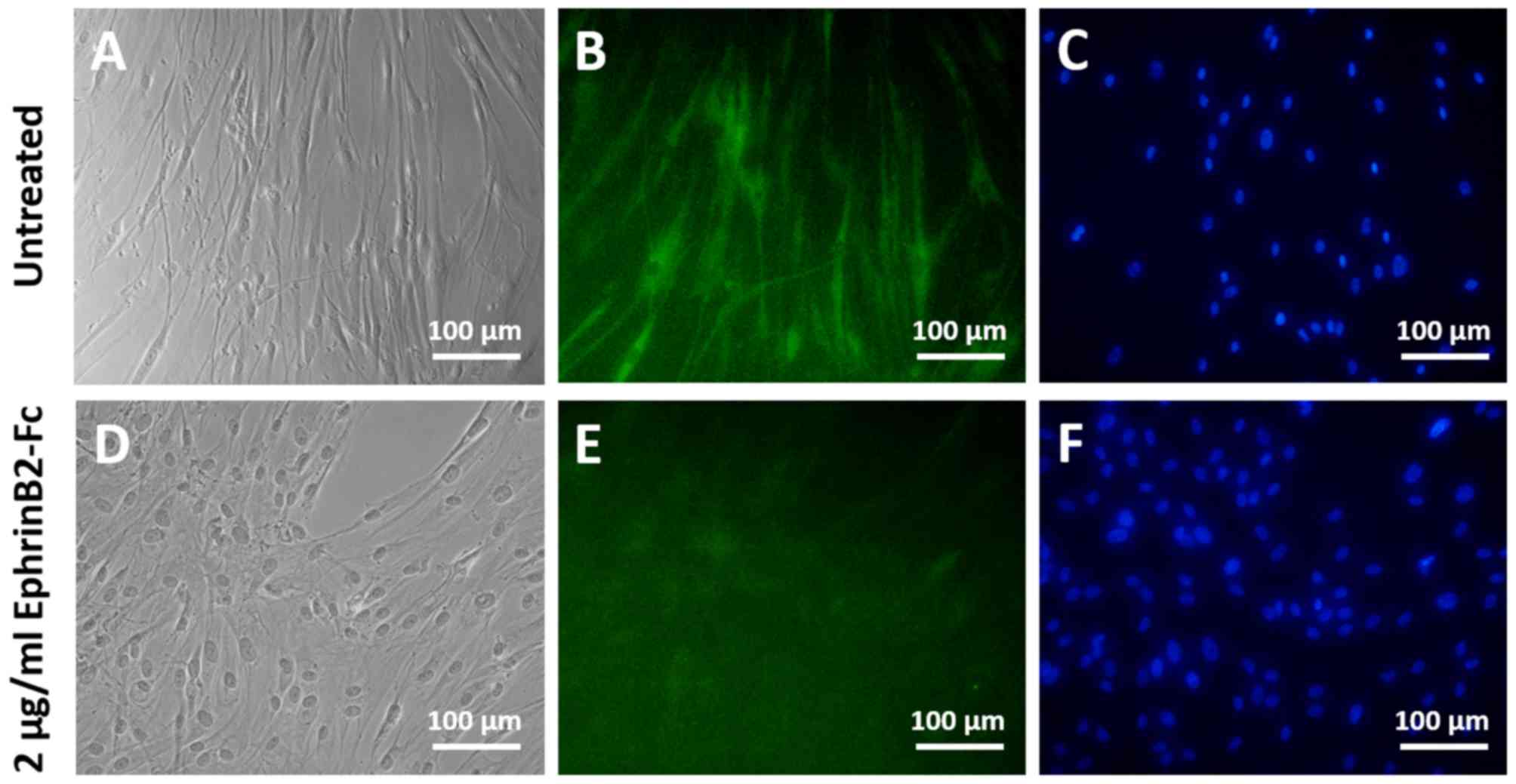
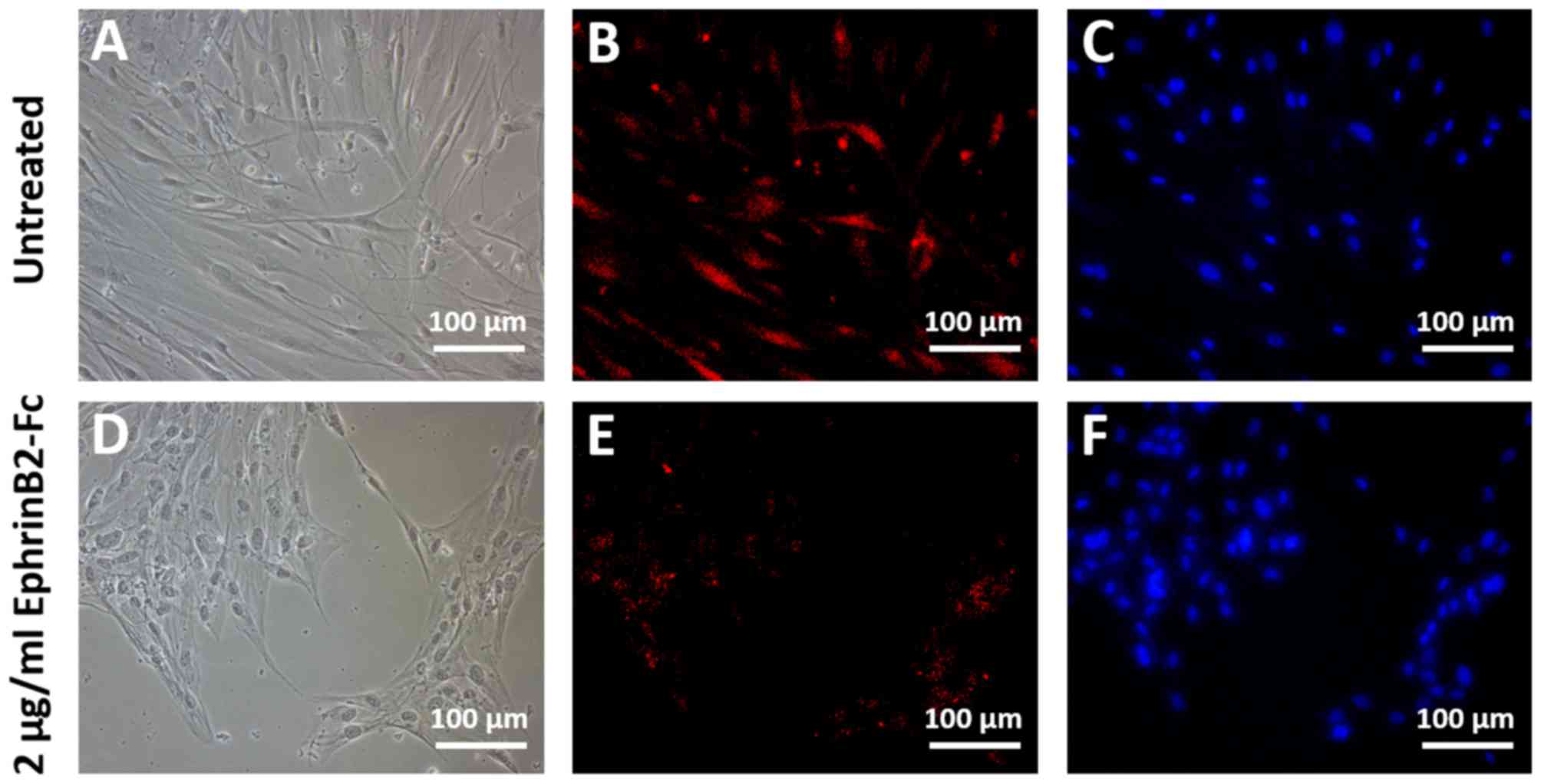
The immunocytochemistry results revealed markedly
lower expression of βIII-tubulin (Fig.
3) and NGN2 (Fig. 4) by the DPSCs
that were exposed to 2 µg/ml EphrinB2-Fc within the neural
induction milieu versus the untreated control. Additionally,
treatment with EphrinB2-Fc led the DPSCs to adopt a more rounded
morphology with fewer neurite outgrowths compared with the
untreated controls (Figs. 3D and
4D). By contrast, the untreated DPSCs
that were subjected to neural induction exhibited elongated
neuron-like morphologies with obvious neurite outgrowths (Figs. 3A and 4A).
Thus, the results demonstrated that treatment with
recombinant EphrinB2-Fc inhibited the neurogenic differentiation of
DPSCs.
Effects of the TNYL-RAW and SNEW
peptides on the neural differentiation of DPSCs
The TNYL-RAW and SNEW peptides bind specifically to
the EphB4 and EphB2 receptors, respectively, block their
interactions with EphrinB2, and thus suppress forward signalling
(20). The RT-qPCR analysis of the
neural markers (Fig. 5) revealed that
the supplementation of the neural induction milieu with 100 µM
TNYL-RAW peptide resulted in significantly increased expression of
MAP2, Musashi1, NGN2 and NSE by the DPSCs (fold expression: 3.01,
1.87, 1.81, and 1.95, respectively, with respect to SCR-WTL control
level expression; P<0.01 for MAP2, P<0.05 for Musashi1, NGN2
and NSE). By contrast, the supplementation of the neural induction
milieu with 100 µM SNEW peptide did not result in significant
changes in any of the 8 analysed neural markers (P>0.05;
Fig. 6).
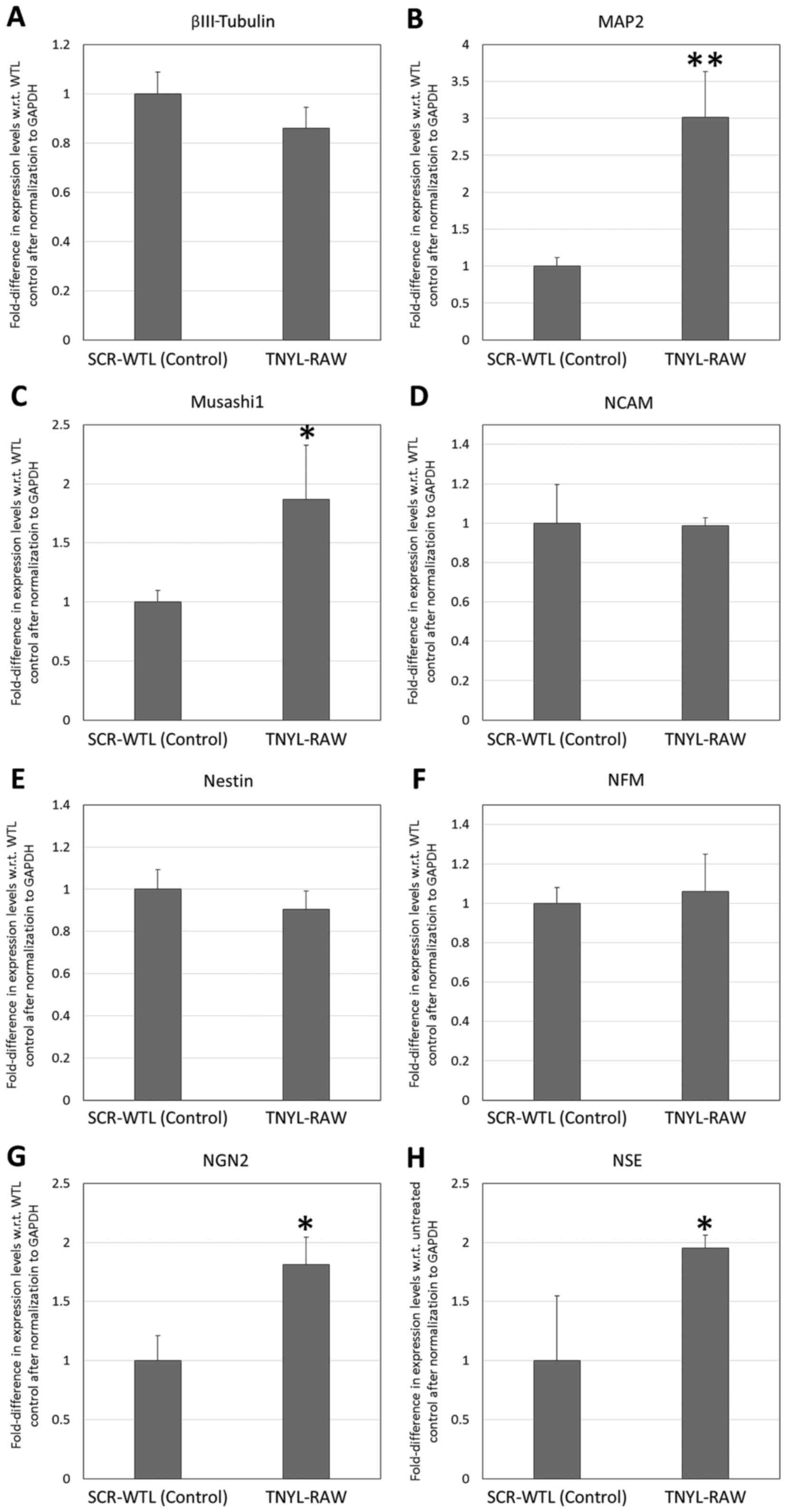 | Figure 5.Reverse transcription-quantitative
polymerase chain reaction analysis of neural marker expression by
dental pulp stem cells following treatment with 100 µM TNYL-RAW or
SCR-WTL (control) peptides in neural induction medium for 7 days.
Fold-difference in expression with respect to untreated controls is
shown for (A) βIII-tubulin, (B) MAP2, (C) Musashi1, (D) NCAM, (E)
nestin, (F) NFM, (G) NGN2 and (H) NSE. *P<0.05 and **P<0.01
vs. Control. MAP2, microtubule-associated protein 2; NCAM, neural
cell adhesion molecule; NFM, neurofilament medium polypeptide;
NGN2, neurogenin 2; NSE, neuron-specific enolase. |
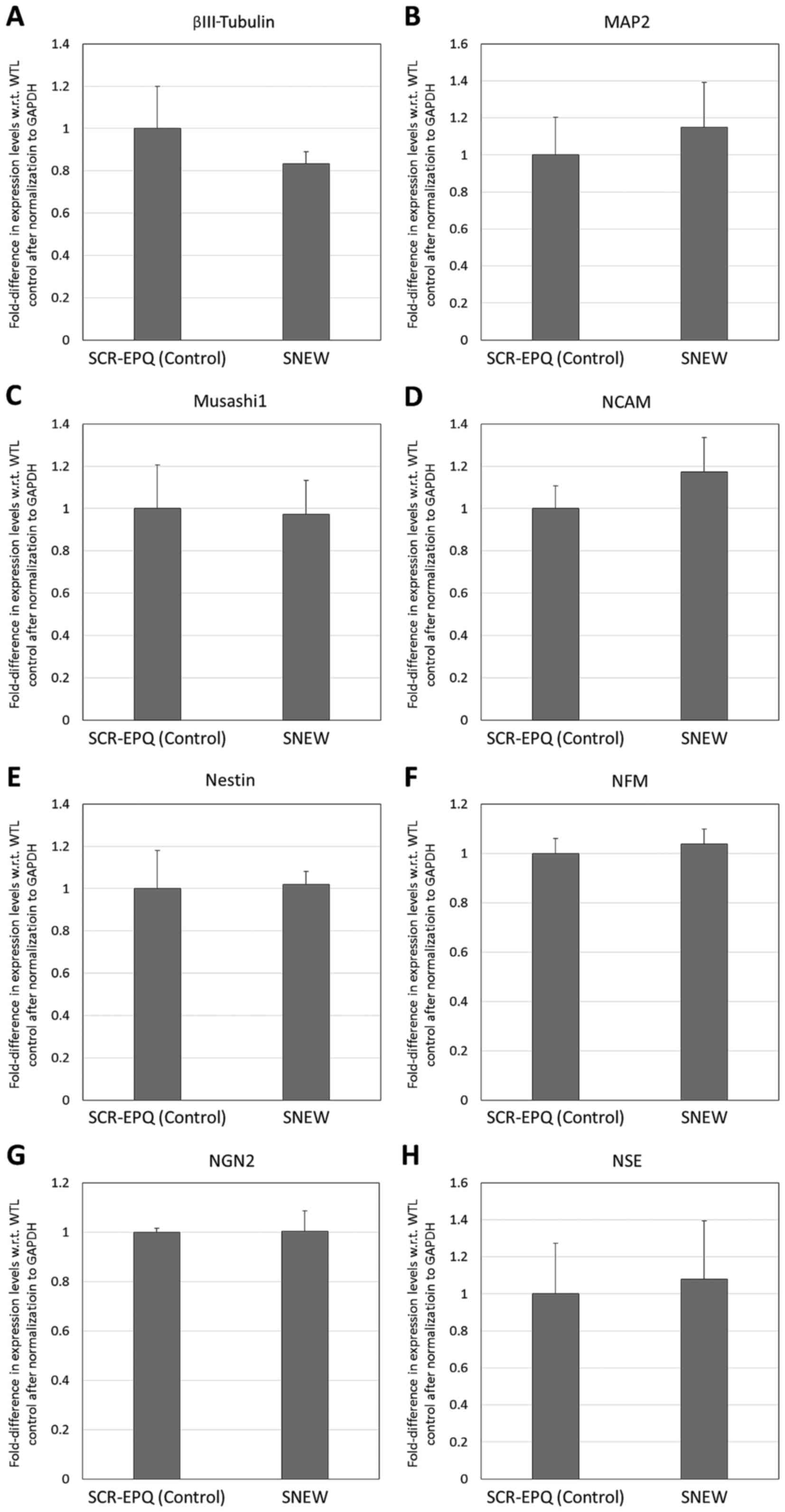 | Figure 6.Reverse transcription-quantitative
polymerase chain reaction analysis of neural marker expression by
dental pulp stem cells following treatment with 100 µM SNEW or
SCR-EPQ (control) peptides in neural induction medium for 7 days.
Fold-difference in expression with respect to untreated controls is
shown for (A) βIII-tubulin, (B) MAP2, (C) Musashi1, (D) NCAM, (E)
nestin, (F) NFM, (G) NGN2 and (H) NSE. There were no significant
differences between the treated group and untreated controls
(P>0.05). MAP2, microtubule-associated protein 2; NCAM, neural
cell adhesion molecule; NFM, neurofilament medium polypeptide;
NGN2, neurogenin 2; NSE, neuron-specific enolase. |
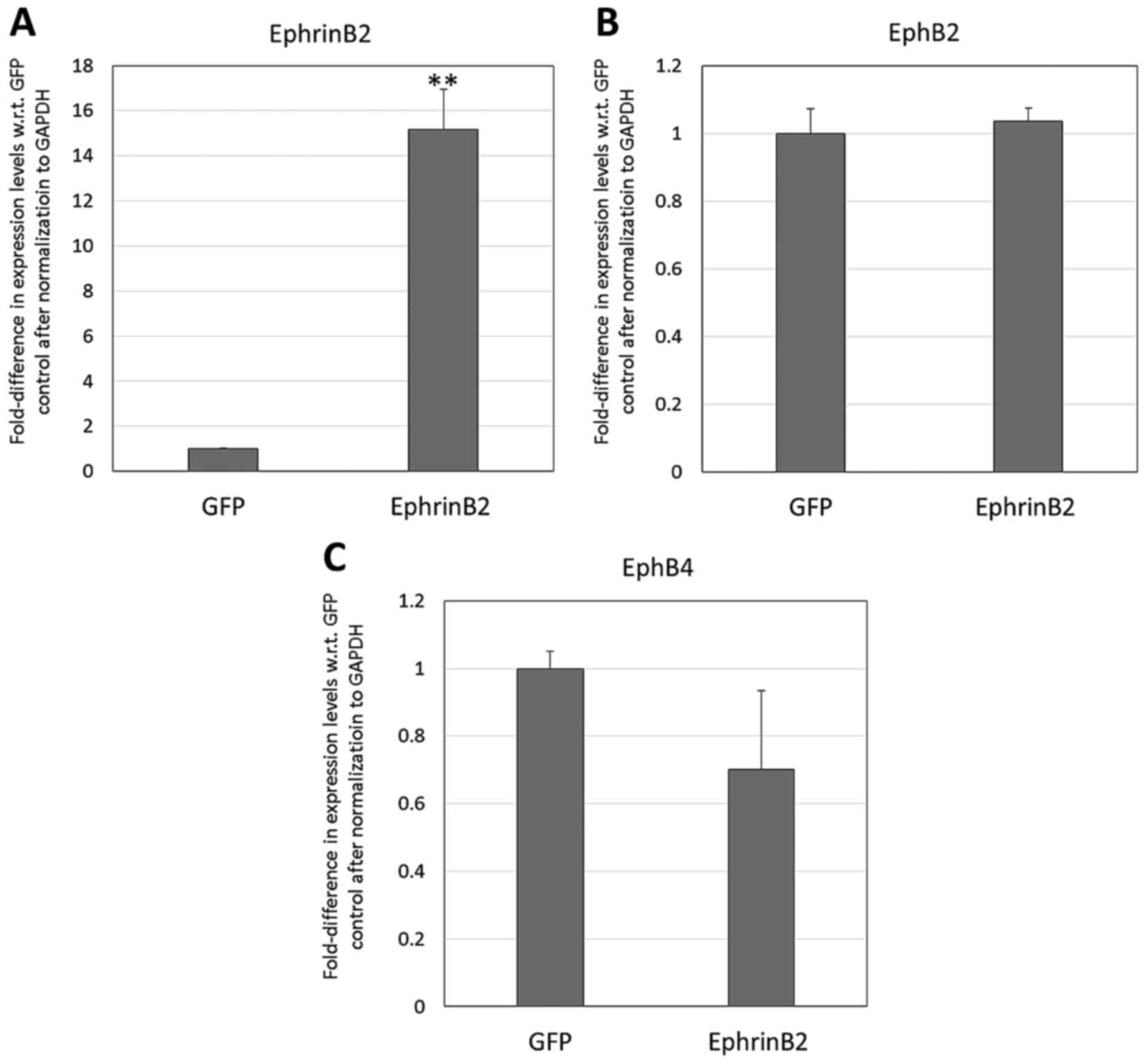
Effect of transgenic expression of
EphrinB2 on the neural differentiation of DPSCs
The RT-qPCR analysis of the neural markers (Fig. 7) revealed significant upregulation of
Musashi1 and NCAM in the transgenic DPSCs compared with in the
null-vector GFP control group (3.86- and 2.41-fold, respectively,
with respect to the null-vector control levels; P<0.01). The
transgenic DPSCs were also identified to overexpress EphrinB2 at a
level that was 15.16±1.78-fold higher than that of the null-vector
controls (P<0.01; Fig. 8A), but
there was no significant difference in the endogenous expression of
EphB2 or EphB4 in the transgenic DPSCs versus the null-vector
controls (P>0.05; Fig. 8B and
C).
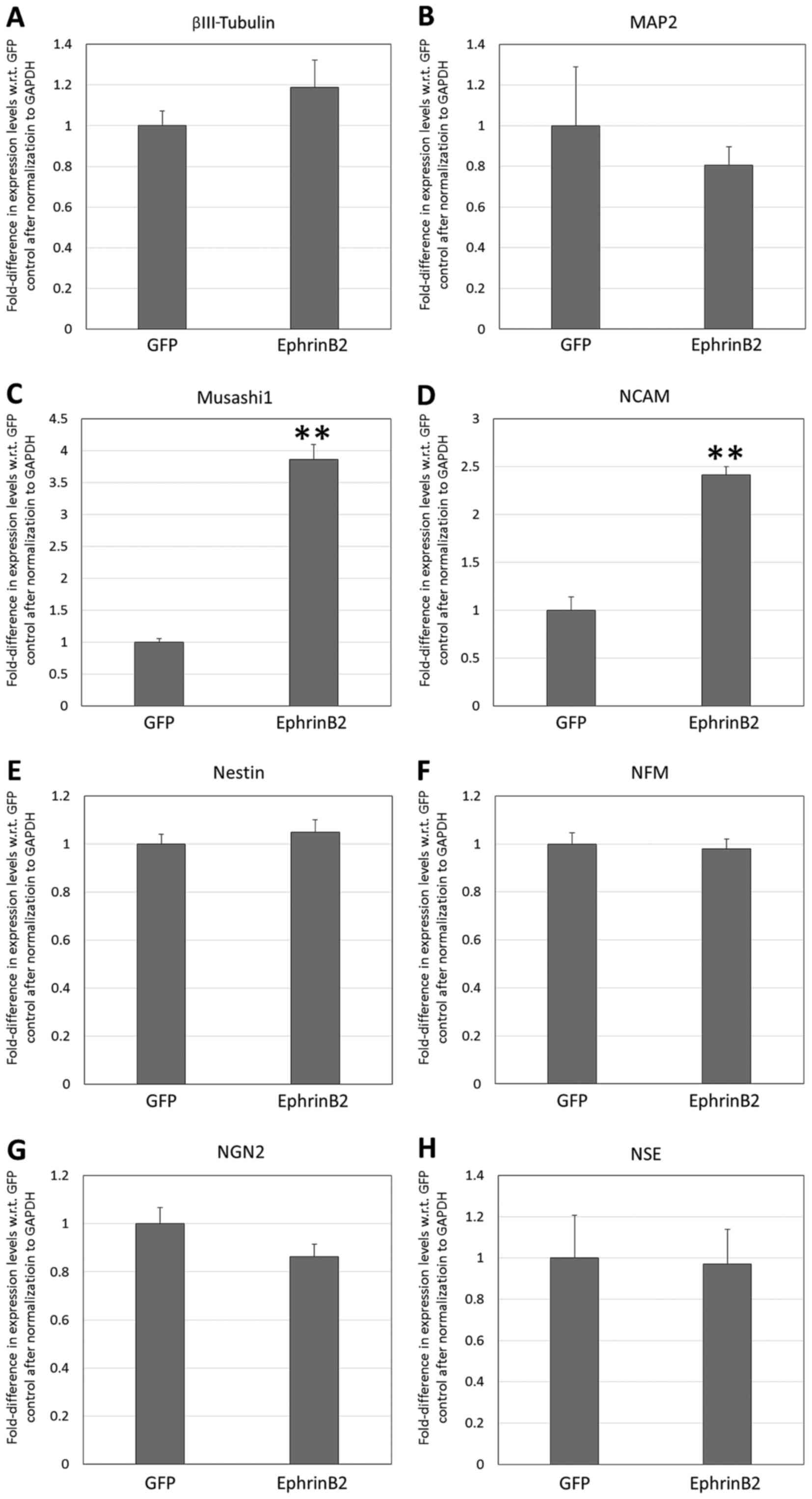 | Figure 7.Reverse transcription-quantitative
polymerase chain reaction analysis of neural marker expression by
transgenic dental pulp stem cells overexpressing recombinant
EphrinB2 following treatment with neural induction medium for 7
days. Fold-difference in expression with respect to untreated
controls is shown for (A) βIII-tubulin, (B) MAP2, (C) Musashi1, (D)
NCAM, (E) nestin, (F) NFM, (G) NGN2, and (H) NSE. **P<0.01 vs.
Control. MAP2, microtubule-associated protein 2; NCAM, neural cell
adhesion molecule; NFM, neurofilament medium polypeptide; NGN2,
neurogenin 2; NSE, neuron-specific enolase. |
Effect of EphB4-Fc on the neural
differentiation of DPSCs
The RT-qPCR analysis of the neural markers (Fig. 9) revealed that the supplementation of
the neural induction milieu with 2 µg/ml EphB4-Fc resulted in
significant upregulation of only Musashi1 (2.00-fold with respect
to the untreated control; P<0.01). The treatment of the DPSCs
with 2 µg/ml EphB4-Fc did not result in any significant changes in
the endogenous expression of EphrinB2, EphB2 or EphB4 (P>0.05;
Fig. 10).
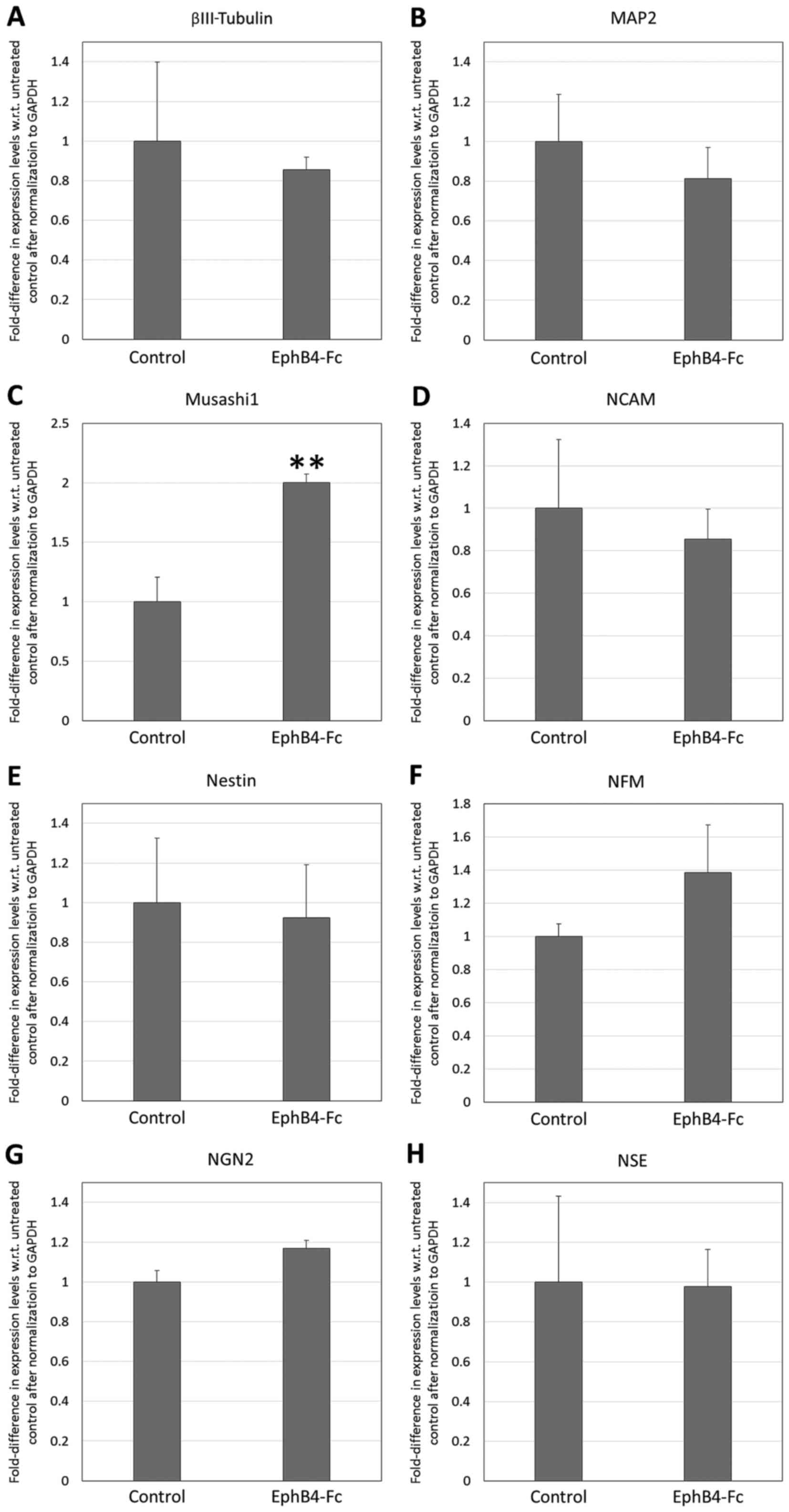 | Figure 9.Reverse transcription-quantitative
polymerase chain reaction analysis of neural marker expression by
dental pulp stem cells following treatment with 2 µg/ml EphB4-Fc in
neural induction medium for 7 days. Fold-difference in expression
with respect to untreated controls is shown for (A) βIII-tubulin,
(B) MAP2, (C) Musashi1, (D) NCAM, (E) nestin, (F) NFM, (G) NGN2 and
(H) NSE. **P<0.01 vs. Control. MAP2, microtubule-associated
protein 2; NCAM, neural cell adhesion molecule; NFM, neurofilament
medium polypeptide; NGN2, neurogenin 2; NSE, neuron-specific
enolase. |
Discussion
Previous studies in animal models have demonstrated
the role of EphrinB2 signalling in the lineage specification of
motor neurons in the spinal cord (13) and dopaminergic neurons in the midbrain
(15). Nevertheless, there are
contradictory and conflicting data within the scientific literature
regarding the role of EphrinB2 signalling in both the promotion and
inhibition of the differentiation of neural stem cells and neural
progenitors. For instance, a study by Ottone et al (16) reported that EphrinB2 ligand expression
by endothelial cells suppressed the differentiation of neural stem
cells and thereby maintained their quiescence and ‘stemness’;
whereas a study by Ashton et al (17) reported that EphrinB2 expression by
hippocampal astrocytes activated β-catenin, upregulated proneural
transcription factors in neural stem cells, and thereby promoted
neural differentiation through juxtacrine signalling. Additionally,
contradictory results regarding the effects of exogenous
recombinant EphrinB2 on neural differentiation have also been
reported in different studies. While Xiaodong et al
(26) reported that soluble
recombinant EphrinB2-Fc maintained the neural progenitor cell state
by inhibiting differentiation, studies by Conway et al
(27) and Conway and Schaffer
(28) demonstrated that the
recombinant ectodomain of EphrinB2 conjugated to biopolymers as
multivalent ligands instead promoted the differentiation of neural
stem cells.
Our results demonstrated that the treatment of DPSCs
with 2 µg/ml EphrinB2-Fc resulted in significant inhibition of
neurogenic differentiation as evidenced by RT-qPCR and
immunocytochemistry results. This finding is consistent with those
of Ottone et al (16) and
Xiaodong et al (26), who
identified that forward EphrinB2 signalling maintained the neural
stem/progenitor cell state by inhibiting neurogenic
differentiation. The observed rounded morphology of DPSCs after 7
days exposure to EphrinB2-Fc under neural inducing conditions, as
compared with the untreated control, may be attributed to
inhibition of cellular adhesion, spreading and migration by
EphrinB2 signalling, as previously demonstrated by Stokowski et
al (29). This involved
initiation of the mitogen-activated protein kinase (MAPK) pathway
by forward signalling, and phosphorylation of the Src family
tyrosine kinases via reverse EphrinB signalling (29).
It may be hypothesized that the observed
downregulation of EphB2 and EphB4 in the DPSCs treated with
EphrinB2-Fc may be due to a negative feedback inhibition loop,
triggered in response to excessive EphrinB2-Fc binding that
activates EphB2 and EphB4 receptors on DPSCs. Previously, Kawano
et al (30) reported that
overstimulation with high dosages of EphrinB2-Fc could trigger a
negative feedback mechanism in the context of T-cell proliferation.
It is possible that the downregulation of EphB2 and EphB4 may form
a part of this negative feedback mechanism, though this requires
further investigation.
The observed inhibitory effects of EphrinB2 forward
signalling on the neurogenic differentiation of DPSCs prompted
investigation into whether specific peptide inhibitors of
EphrinB2-EphB4 and EphrinB2-EphB2 signalling could instead promote
the neurogenic differentiation of DPSCs. The TNYL-RAW and SNEW
peptides, which specifically inhibit the EphB4 and EphB2 receptors,
respectively (21), together with the
corresponding control scrambled peptides (SCR-WTL and SCR-EPQ,
respectively) were utilized. The results indicated that the
TNYL-RAW peptide (specific for EphB4) could enhance neurogenic
differentiation as evidenced by the upregulation of 4 neural
markers (MAP2, Musashi1, NGN2 and NSE). By contrast, the SNEW
peptide (specific for EphB2) did not elicit significant changes in
the expression of any of the 8 analysed neural markers. Thus, it
may be deduced that the EphrinB2-EphB4 interaction serves a more
prominent role in the neurogenesis of DPSCs relative to the
EphrinB2-EphB2 signalling axis.
Subsequently, the present study investigated the
effects of transgenic overexpression of EphrinB2 on the neural
differentiation of DPSCs. The overexpressed EphrinB2 may stimulate
both forward and reverse signalling in the transgenic DPSCs. The
current results demonstrated that there was only a slight
enhancement of neural differentiation in the transgenic DPSCs
overexpressing EphrinB2, and only 2 neural markers (Musashi1 and
NCAM) were upregulated. As the results on EphrinB2-Fc indicated
that forward signalling inhibited neurogenic differentiation in
DPSCs, the marginal enhancement of neurogenesis may possibly arise
from an increase in reverse signalling, mediated via activation of
the overexpressed EphrinB2 by either endogenous EphB2 or EphB4
receptors on adjacent cells. This notion in turn prompted
investigation into whether the treatment of DPSCs with soluble
recombinant EphB4 to stimulate reverse signalling could enhance the
neurogenic differentiation of DPSCs. However, the results revealed
a significant increase in the expression of the neural marker
Musashi1 only.
Therefore, to conclude, stimulation of forward
EphrinB2-EphB4 signalling markedly inhibited the neurogenesis of
DPSCs, whereas suppression of this forward signalling pathway with
the TNYL-RAW peptide inhibitor promoted neural differentiation.
Additionally, stimulation of reverse EphB4-EphrinB2 signalling was
also observed to marginally enhance the neural differentiation of
DPSCs. Future studies by our group will investigate whether
combining the TNYL-RAW peptide with inhibitors of other signalling
pathways can produce a synergistic effect on the promotion of DPSC
neurogenesis. Possible candidates include inhibitors of the
Jagged-Notch signalling pathway, since this particular signalling
pathway was demonstrated in a previous study (16) to work together with forward EphrinB2
signalling to inhibit the differentiation of neural stem cells.
Such peptide or small molecule inhibitors of EphrinB2 forward
signalling may have potential clinical applications in therapies
based on neural tissue engineering with DPSCs.
Acknowledgements
The authors would like to thank Ms. Edith Tong of
the Faculty of Dentistry of the University of Hong Kong for her
assistance in performing the various experiments of the present
study.
Funding
The present study was supported by a grant from the
National Nature Science Foundation of China (grant no. 81470735)
and a General Research Fund (GRF) grant from the Research Grants
Council of Hong Kong (grant no. HKU17126914) to CZ.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BCH performed the experiments, analyzed and
interpreted the data and wrote the draft manuscript. TG, JX and LWL
aided with the experiments and data collection and critically
edited the manuscript. CZ designed and managed the study. All
authors have read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The research protocol was reviewed and approved by
the Human Research Ethics Committee of the University of Hong Kong,
China. All subjects provided written informed consent for
participation in the study.
Consent for publication
All subjects provided written informed consent
permitting the publication of relevant data following anonymization
of personally identifiable information.
Competing interests
All authors declare that they have no competing
interests.
References
|
1
|
Gronthos S, Mankani M, Brahim J, Robey PG
and Shi S: Postnatal human dental pulp stem cells (DPSCs) in vitro
and in vivo. Proc Natl Acad Sci USA. 97:13625–13630. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gronthos S, Brahim J, Li W, Fisher LW,
Cherman N, Boyde A, DenBesten P, Robey PG and Shi S: Stem cell
properties of human dental pulp stem cells. J Dent Res. 81:531–535.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang GT, Gronthos S and Shi S:
Mesenchymal stem cells derived from dental tissues vs. those from
other sources: Their biology and role in regenerative medicine. J
Dent Res. 88:792–806. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nuti N, Corallo C, Chan BM, Ferrari M and
Gerami-Naini B: Multipotent differentiation of Human dental pulp
stem cells: A literature review. Stem Cell Rev. 12:511–523. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sharpe PT: Dental mesenchymal stem cells.
Development. 143:2273–2280. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Heng BC, Lim LW, Wu W and Zhang C: An
overview of protocols for the neural induction of dental and oral
stem cells in vitro. Tissue Eng Part B Rev. 22:220–250. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chai Y, Jiang X, Ito Y, Bringas P Jr, Han
J, Rowitch DH, Soriano P, McMahon AP and Sucov HM: Fate of the
mammalian cranial neural crest during tooth and mandibular
morphogenesis. Development. 127:1671–1679. 2000.PubMed/NCBI
|
|
8
|
Song M, Lee JH, Bae J, Bu Y and Kim EC:
Human dental pulp stem cells are more effective than human bone
marrow-derived mesenchymal stem cells in cerebral ischemic injury.
Cell Transplant. 26:1001–1016. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang C, Li X, Sun L, Guo W and Tian W:
Potential of human dental stem cells in repairing the complete
transection of rat spinal cord. J Neural Eng. 14:0260052017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sasaki R, Aoki S, Yamato M, Uchiyama H,
Wada K, Ogiuchi H, Okano T and Ando T: PLGA artificial nerve
conduits with dental pulp cells promote facial nerve regeneration.
J Tissue Eng Regen Med. 5:823–830. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mead B, Logan A, Berry M, Leadbeater W and
Scheven BA: Intravitreally transplanted dental pulp stem cells
promote neuroprotection and axon regeneration of retinal ganglion
cells after optic nerve injury. Invest Ophthalmol Vis Sci.
54:7544–7556. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Arthur A, Koblar S, Shi S and Gronthos S:
Eph/ephrinB mediate dental pulp stem cell mobilization and
function. J Dent Res. 88:829–834. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Laussu J, Audouard C, Kischel A,
Assis-Nascimento P, Escalas N, Liebl DJ, Soula C and Davy A:
Eph/Ephrin signaling controls progenitor identities in the ventral
spinal cord. Neural Dev. 12:102017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luxey M, Laussu J and Davy A: EphrinB2
sharpens lateral motor column division in the developing spinal
cord. Neural Dev. 10:252015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yue Y, Widmer DA, Halladay AK, Cerretti
DP, Wagner GC, Dreyer JL and Zhou R: Specification of distinct
dopaminergic neural pathways: Roles of the Eph family receptor
EphB1 and ligand ephrin-B2. J Neurosci. 19:2090–2101. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ottone C, Krusche B, Whitby A, Clements M,
Quadrato G, Pitulescu ME, Adams RH and Parrinello S: Direct
cell-cell contact with the vascular niche maintains quiescent
neural stem cells. Nat Cell Biol. 16:1045–1056. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ashton RS, Conway A, Pangarkar C, Bergen
J, Lim KI, Shah P, Bissell M and Schaffer DV: Astrocytes regulate
adult hippocampal neurogenesis through ephrin-B signaling. Nat
Neurosci. 15:1399–1406. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Barquilla A and Pasquale EB: Eph receptors
and ephrins: Therapeutic opportunities. Annu Rev Pharmacol Toxicol.
55:465–487. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nikolov DB, Xu K and Himanen JP:
Eph/ephrin recognition and the role of Eph/ephrin clusters in
signaling initiation. Biochim Biophys Acta. 1834:2160–2165. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Heng BC, Wang S, Gong T, Xu J, Yuan C and
Zhang C: EphrinB2 signaling enhances osteogenic/odontogenic
differentiation of human dental pulp stem cells. Arch Oral Biol.
87:62–71. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chrencik JE, Brooun A, Recht MI, Nicola G,
Davis LK, Abagyan R, Widmer H, Pasquale EB and Kuhn P:
Three-dimensional structure of the EphB2 receptor in complex with
an antagonistic peptide reveals a novel mode of inhibition. J Biol
Chem. 282:36505–36513. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zou T, Dissanayaka WL, Jiang S, Wang S,
Heng BC, Huang X and Zhang C: Semaphorin 4D enhances angiogenic
potential and suppresses osteo-/odontogenic differentiation of
human dental pulp stem cells. J Endod. 43:297–305. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Heng BC, Gong T, Wang S, Lim LW, Wu W and
Zhang C: Decellularized matrix derived from neural differentiation
of embryonic stem cells enhances the neurogenic potential of dental
follicle stem cells. J Endod. 43:409–416. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Davis S, Gale NW, Aldrich TH, Maisonpierre
PC, Lhotak V, Pawson T, Goldfarb M and Yancopoulos GD: Ligands for
EPH-related receptor tyrosine kinases that require membrane
attachment or clustering for activity. Science. 266:816–819. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xiaodong H, Zhen H, Min S, Zhiming C,
Hongyan J, Chong Z, Xuefeng T and Guohua J: Direct inhibition of
cell surface ephrin-B2 by recombinant ephrin-B2/FC. Biochem Biophys
Res Commun. 440:300–305. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Conway A, Vazin T, Spelke DP, Rode NA,
Healy KE, Kane RS and Schaffer DV: Multivalent ligands control stem
cell behaviour in vitro and in vivo. Nat Nanotechnol. 8:831–838.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Conway A and Schaffer DV: Biomaterial
microenvironments to support the generation of new neurons in the
adult brain. Stem Cells. 32:1220–1229. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Stokowski A, Shi S, Sun T, Bartold PM,
Koblar SA and Gronthos S: EphB/ephrin-B interaction mediates adult
stem cell attachment, spreading, and migration: Implications for
dental tissue repair. Stem Cells. 25:156–164. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kawano H, Katayama Y, Minagawa K,
Shimoyama M, Henkemeyer M and Matsui T: A novel feedback mechanism
by Ephrin-B1/B2 in T-cell activation involves a
concentration-dependent switch from costimulation to inhibition.
Eur J Immunol. 42:1562–1572. 2012. View Article : Google Scholar : PubMed/NCBI
|