Introduction
Platelets are involved in multiple immune responses,
including hemostasis, thrombosis and wound healing. However, recent
studies have highlighted their distinct functions in inflammatory
responses. Upon infection, platelets first bind to infectious
pathogens, such as bacteria or viruses. Then, the platelets secrete
various immunoregulatory cytokines and express receptors to mediate
various immune effects and regulatory functions (1,2),
including regulation of macrophage functions, stimulation or
inhibition of regulatory T cells, and secretion of pro-resolving
mediators (3,4). However, their regulatory functions can
be complex: Both pro-inflammatory and anti-inflammatory regulations
are modulated by the degree and phase of injury and inflammation,
which vary on a case-by-case basis (2-6).
The novel severe acute respiratory syndrome
coronavirus 2 (SARS-CoV-2) is the causative agent of the current
coronavirus disease-2019 (COVID-19) pandemic. The virus continues
to pose a significant threat to global health. Like most other
coronaviruses, SARS-CoV-2 is comprised of the standard biological
components, including an envelope, a membrane and spike proteins
(7). The spike protein recognizes
and binds to the host cell surface receptor, angiotensin-converting
enzyme 2 (ACE2), which facilitates entry of SARS-CoV-2 into host
cells (7), including platelets. It
is hypothesized that the behavior of the virus is similar to the
influenza virus (1,8,9).
Initially, coronavirus infection elevates fibrinogen and D-dimer
levels to evoke systemic hypercoagulability and frequent venous
thromboembolic events (10). This
could be considered a severe proinflammatory state, which
consequently induces pro-coagulopathy through endothelial
activation or damage (10). As
anticipated, recent studies have found that the number of
activated, circulating platelets increase in COVID-19 patients
(10-13).
This suggests that platelets may contribute to COVID-19 severity
(13), although the role of
platelets in the pathogenesis of SARS-CoV-2 is not yet well
understood.
Several COVID-19 vaccines are now widely available
and others are still in production. The World Health Organization
(WHO) and the U.S. Food and Drug Administration (FDA) approved the
Pfizer/BioNTech mRNA vaccine (BNT162b2) as one of the first
COVID-19 vaccines on the 23rd of August, 2021 (14,15).
Due to these approvals, COVID-19 vaccines based on mRNA technology
dominate the seven major markets (7MM), comprised of the US,
Germany, France, Italy, Spain, the UK and Japan, which held a
collective market share of 95.4% in 2021(16). Although the market shares vary among
individual countries at present (17), like other major countries including
other members of the 7MM, the Japanese government approved and
provided the Pfizer/BioNTech vaccine, first to medical staff as a
priority followed by older citizens. The Moderna mRNA vaccine
(mRNA-1273) has since been approved as the second major vaccine,
now widely available to all applicants.
Both the Pfizer/BioNTech and Moderna mRNA vaccines
are designed to produce the spike protein of SARS-CoV-2 and then to
induce formation of spike protein-specific neutralizing antibodies,
which are expected to protect against infection by the target
viruses through immunological reactions (18,19).
As these vaccines mimic the initial phase of reaction to SARS-CoV-2
infection, these agents can also cause various side-effects that
are consistent with the symptoms of infection by
SARS-CoV-2(20). Thus, it is
possible that the vaccines stimulate platelet activation and
degranulation, to modify inflammation and immunological reactions,
likely involving polyphosphate (polyP).
Among platelet regulatory molecules, platelet polyP
has recently received increasing attention, not only as a
coagulation factor, but also as a potent modulator of inflammation
(21-23).
polyP is a linear polymer of orthophosphates linked by high-energy
phospho-anhydride bonds, and is stored primarily in platelet dense
granules and released upon activation of platelets (21,24).
polyP can act as a proinflammatory factor or an anti-inflammatory
factor by triggering bradykinin release and inhibiting complement
activation, respectively (25,26).
Thus, it is hypothesized that platelets, activated by infection by
pathogens, release pro-inflammatory and anti-inflammatory
biomolecules, including polyP, thereby exacerbating or suppressing
inflammation, respectively. To determine the possible involvement
of platelet polyP in the COVID-19 mRNA vaccine-induced
side-effects, including the immune responses, the effects of the
COVID-19 mRNA vaccine on platelet polyP levels were examined.
Materials and methods
Preparation of the platelet
suspensions
The study design and consent forms for all
procedures (approval no. 2019-0423) were approved by the Ethics
Committee for Human Participants at the Niigata University School
of Medicine (Niigata, Japan), and complied with the Helsinki
Declaration of 1964, as revised in 2013 (27,28).
Vaccination with mRNA vaccines, BNT162b2 and
mRNA-1273, was controlled by the Japanese government and local
governments (29). In principle,
the residents were categorized into several groups depending on
various factors, such as age, severity of pre-existing diseases,
area of residence and type of occupation. The government scheduled
vaccinations without or upon the requests of individuals, and
designated a vaccination date. The first priority for vaccination
was given to medical workers, ambulance crews and public health
center officials.
Healthy Japanese subjects (male: first vaccination,
n=6, median age, 55.5, age range, 47-60); second vaccination, n=8,
median age, 55.0, age range, 47-72; and female: first vaccination,
n=14, median age, 29.5, age range, 20-57; second vaccination, n=23,
median age, 32.0, age range, 21-66), who were selected from medical
workers that were not receiving continuous medical treatment and
would be receiving the Pfizer/BioNTech vaccine (BNT162b2)
participated in this study. As for the inclusion criteria, the
subjects were required to have no limitations on their activities
of daily living, regardless of lifestyle-related diseases or
regular medication. Subjects who had cancer or received estrogen
replacement therapy were excluded. All subjects were required to
declare that they were free of human immunodeficiency virus,
hepatitis B virus, hepatitis C virus, or syphilis infections; any
subjects who had been infected with any of these viruses were
excluded from the study. No acute reactions were observed in the
subjects during or immediately after blood samples were taken.
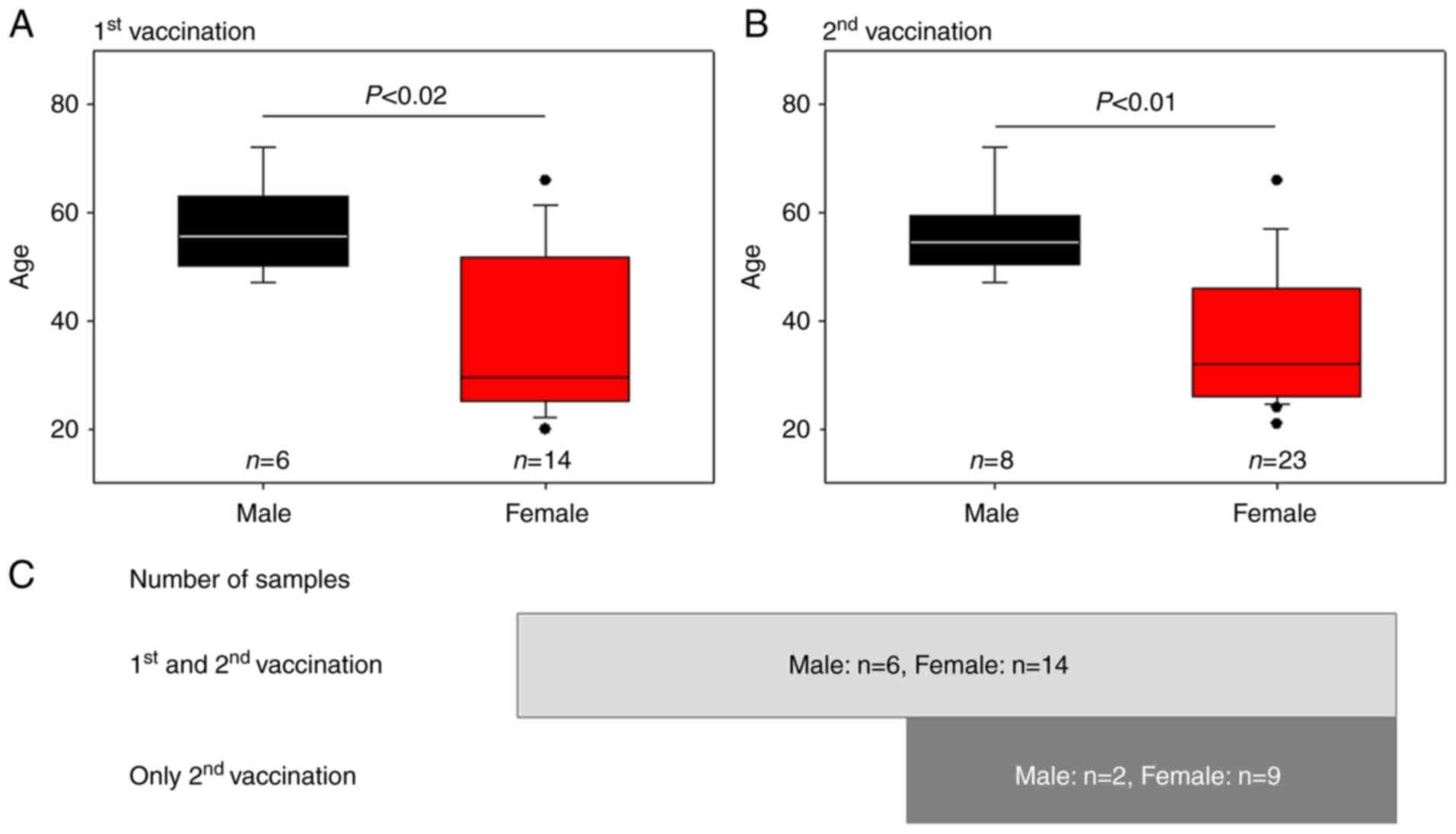
The age distributions of the sample groups and the
participants' constitutions (male numbers vs. female numbers) are
shown in Fig. 1. The female groups
(first dose, 36.3±5.0 years; second dose, 36.3±12.3 years, mean ±
SD) were significantly younger than the respective male groups
(first dose, 56.8±8.8 years; second dose, 56.0±7.8 years). All
participants (male, n=6; female, n=14) provided blood samples after
both the first and second vaccinations. Additional participants
(male, n=2; female, n=9) provided blood samples only after the
second vaccination.
When the participants were available during the
daytime on weekdays, after an appropriate interval of time
following receipt of the vaccine (first vaccination, 4-10 days,
median, 8 days; second vaccination, 4-15 days, median, 7 days),
blood withdrawal was performed by venipuncture of the median
cubital vein using 21G wing needles (NIPRO). Blood was collected in
vacutainer A-formulation of acid-citrate-dextrose (ACD-A) glass
tubes (BD Biosciences). Blood cells from each sample were counted
using an automated hematology analyzer (pocH-100i V Diff; Sysmex)
and intermittently rotated at 22-25˚C until use.
Prior to centrifugation, 0.5 ml ACD-A (Terumo) was
added to each blood sample, then the samples were incubated for 10
min to prevent unexpected coagulation, which is often observed in
female blood samples (30).
Platelets were collected using the dual-spin protocol of
pure-platelet rich plasma (PRP) preparation (1st soft spin: 472 x g
for 10 min; 2nd hard spin: 578 x g for 5 min, at 22-25˚C). The
resulting platelet pellets were then gently suspended in PBS,
incubated for 15 min at 22-25˚C, and fixed with ThromboFix Platelet
Stabilizer (Beckman-Coulter, Inc.). The fixed samples were stored
at 4˚C for at least 24 h prior to polyP quantification. It should
be noted that the centrifugation conditions were mild, thus
minimizing platelet activation among the preparation conditions
performed (31).
Additionally, healthy Japanese subjects who received
the Moderna vaccine (mRNA-1273) were selected (5 females; median
age 37.0; age range 34-48 years), using the same inclusion and
exclusion criteria as above.
Fluorometric measurement of platelet
polyphosphate using DAPI
After storage at 4˚C for 24 h, the fixed platelets
were centrifuged at 578 x g for 5 min at 22-25˚C, after which the
supernatants were carefully aspirated. The resulting platelet
pellets were gently suspended in Milli-Q water, as previous
observations found that Milli-Q water does not increase background
of fluorescence signal quantification, and enables DAPI to invade
the cytoplasm without perforation (30). Platelets were counted using an
automated hematology analyzer and then adjusted to the appropriate
range (<4.5x107/ml) for polyP quantification. DAPI
(Dojindo Molecular Technologies, Inc.) was added to each tube at a
concentration of 4 µg/ml and incubated for 30 min at 22-25˚C.
Fluorescence intensity was measured using a fluorometer (FC-1;
Tokai Optical Co., Ltd.) with excitation and emission wavelengths
of 425 and 525 nm, respectively (30).
Due to the lack of specific probes, it is difficult
to accurately quantify platelet polyP levels in an efficient
manner. Therefore, our methods are modeled using previous studies
(30,32,33)
showing that the right shift of excitation and emission wavelengths
(350 and 470 nm) from those wavelengths used for DNA examination
increases the specificity of DAPI against polyP, with some
modifications. Specifically, quantification and visualization of
platelet polyP using DAPI was optimized through the determination
of the optimum light wavelengths for fluorescence, as well as for
sample fixation and choice of the aqueous solution. Although the
possible count of DAPI-bound, non-polyP substances may be included
in the quantification process, this method enables rapid
quantification of polyP levels without sacrificing recovery or
reproducibility.
The polyP ratio was calculated for each individual,
and all polyP ratios were combined for statistical analysis.
Determination of mitochondrial
activity using a Cell Counting Kit-8 (CCK-8) assay
The number of platelets suspended in PBS was
adjusted to the density range of 2.5-5.0x107/100 µl,
mixed with 10 µl CCK-8 solution (Dojindo Molecular Technologies,
Inc.), and incubated for 1 h at 22-25˚C. After centrifugation (578
x g, 5 min, 22-25˚C), the supernatants were analyzed using a
spectrophotometer at 450 nm (SmartSpec Plus; Bio-Rad Laboratories,
Inc.).
Evaluation of side-effects based on
individual self-declarations
The score sheet of the side-effects are shown in
Table I. The perception of vaccine
side-effects was examined by asking the subjects the following
questions: ‘Have you had any of these side effects? Please score
each side-effect on a scale where: 0, not experienced; 1,
experienced; and 2, experienced strongly (more important or severe
for you). If you took painkillers, please score 1. Also, please
mention the number of days the side-effects lasted and the
frequency of taking medicines treating them.’ These questions were
introduced as described in the questionnaire to the donors.
 | Table IList of vaccine-induced side-effects
and scoring of severity based on patient declarations. |
Table I
List of vaccine-induced side-effects
and scoring of severity based on patient declarations.
| Side-effects | Severity score
(a) | Number of days or
doses of medicine taken (b) | Final score (a x
b) |
|---|
| Systemic
side-effects | | | |
|
Fever,
temperature >37.5˚C | 2 | | |
|
Slight
fever, temperature 37.2-37.5˚C | 1 | | |
|
Fatigue | 1 | | |
|
Headache | 1 | | |
|
Others
including chills, arthralgia, myalgia, nausea, etc. | 1 | | |
| Local
side-effects | | | |
|
Pain | 1 | | |
|
Redness | 1 | | |
|
Swelling | 1 | | |
|
Others,
including tenderness, itch, warmth, bruising, etc. | 1 | | |
| Allergic
reaction | | | |
|
Rash | 1 | | |
|
Skin
burning | 1 | | |
|
Red welts on
face and lips | 1 | | |
| Analgesics
dose | | | |
|
Type of
analgesicsa | 1 | | |
Statistical analysis
Quantification was performed in triplicate. Data are
presented as the mean ± standard deviation. Data plotted in box
plots represent the median values, the upper and lower quartiles
(box), and maximum and minimum values (whiskers). A Mann-Whitney U
test was performed to compare the two groups (SigmaPlot version
13.0; Systat Software, Inc.). To confirm the statistical difference
provided by the Mann-Whitney U test, particularly given the fact
that the score values were not continuous variables, data for the
apparent reduction of platelet polyP levels (<0.9) and the
apparent side-effect scores (>8.2) in individual data were also
analyzed using a Fisher's exact test. These border values were
derived from the mean values of the female group's polyP ratio
post-first vaccination (0.9%) and the female group's score
post-second vaccination (8.2). P<0.05 was considered to indicate
a statistically significant difference.
Linear regression analysis and calculation of
correlation coefficient values were performed. Absolute R values
ranging from 0.6-0.79 and 0.4-0.59 were considered a ‘strong’ and
‘moderate’ correlation, respectively. R values of 0.2-0.39 and
<0.2 indicated a ‘weak’ and ‘very weak’ correlation,
respectively.
Results
Initially, platelet counts were examined to detect
thrombocytopenia before and after vaccination. As shown in Table II, no significant reduction
following vaccination was found in any of the participant
groups.
| Table IIPlatelet counts in whole-blood
samples. |
Table II
Platelet counts in whole-blood
samples.
| | Male,
x104/µl | Female,
x104/µl |
|---|
| Vaccination
status | First vaccination,
n=6 | Second vaccination,
n=8 | First vaccination,
n=14 | Second vaccination,
n=23 |
|---|
|
Pre-vaccination | 25.8±4.3 | 26.0±4.4 | 25.9±6.2 | 26.9±8.3 |
| Post-1st
vaccination | 26.3±4.7 | - | 25.5±6.3 | - |
| Post-2nd
vaccination | - | 25.7±4.6 | - | 24.8±6.0 |
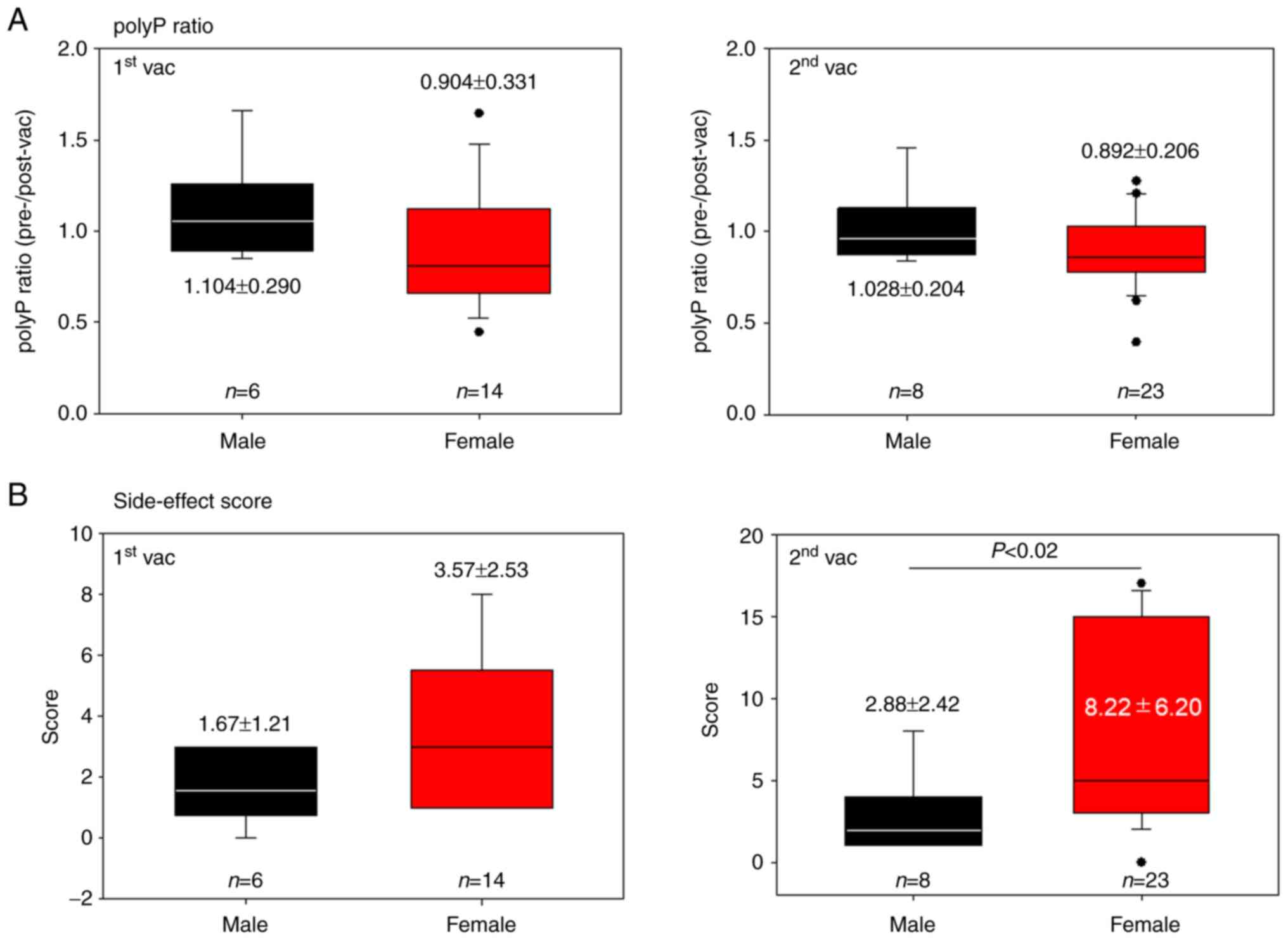
Ratios of polyP levels after vaccination (BNT162b2)
to polyP levels pre-vaccination, that is, vaccine-induced reduction
of polyP levels, are shown in Fig.
2A. There were no significant differences between the male and
female groups after either the first vaccination (1.104±0.290 vs.
0.904±0.331) or the second vaccination (1.028±0.204 vs.
0.892±0.206). Scores for the side-effects in the post-vaccination
period are shown in Fig. 2B. The
scores were generally lower in the male groups than in the female
groups; however, a significant statistical difference in symptoms
between males and females (P<0.02) was found only after the
second vaccination (2.88±2.42 vs. 8.22±6.20) (c.f., first
vaccination, 1.67±1.21 vs. 3.57±2.53) of the vaccine.
To confirm the statistical difference, the data were
analyzed again using a Fisher's exact test. A statistically
significant difference (P=0.042) between males and females was
found in the polyP ratios after the first vaccination (Table III). Border values were adopted by
the mean values of the post-first (polyP=0.9%) and post-second
vaccination (score=0.82) scores in the female groups.
| Table IIINumber of subjects showing apparent
reduction of platelet polyphosphate and apparent exacerbation of
vaccine side-effect scores following vaccination with the
Pfizer-BioNTech vaccine. |
Table III
Number of subjects showing apparent
reduction of platelet polyphosphate and apparent exacerbation of
vaccine side-effect scores following vaccination with the
Pfizer-BioNTech vaccine.
| Vaccine dose | n | Platelet
polyphosphate reduction <0.9 (%) |
P-valuea | Side-effect score
>8.2 |
P-valuea |
|---|
| 1st | | | | | |
|
Male | 6 | 0 (0.0) | 0.042b | 0 (0.0) | - |
|
Female | 14 | 8 (57.1) | | 0 (0.0) | |
| 2nd | | | | | |
|
Male | 8 | 3 (37.5) | 0.412 | 0 (0.0) | 0.032b |
|
Female | 23 | 14 (60.9) | | 10 (39.1) | |
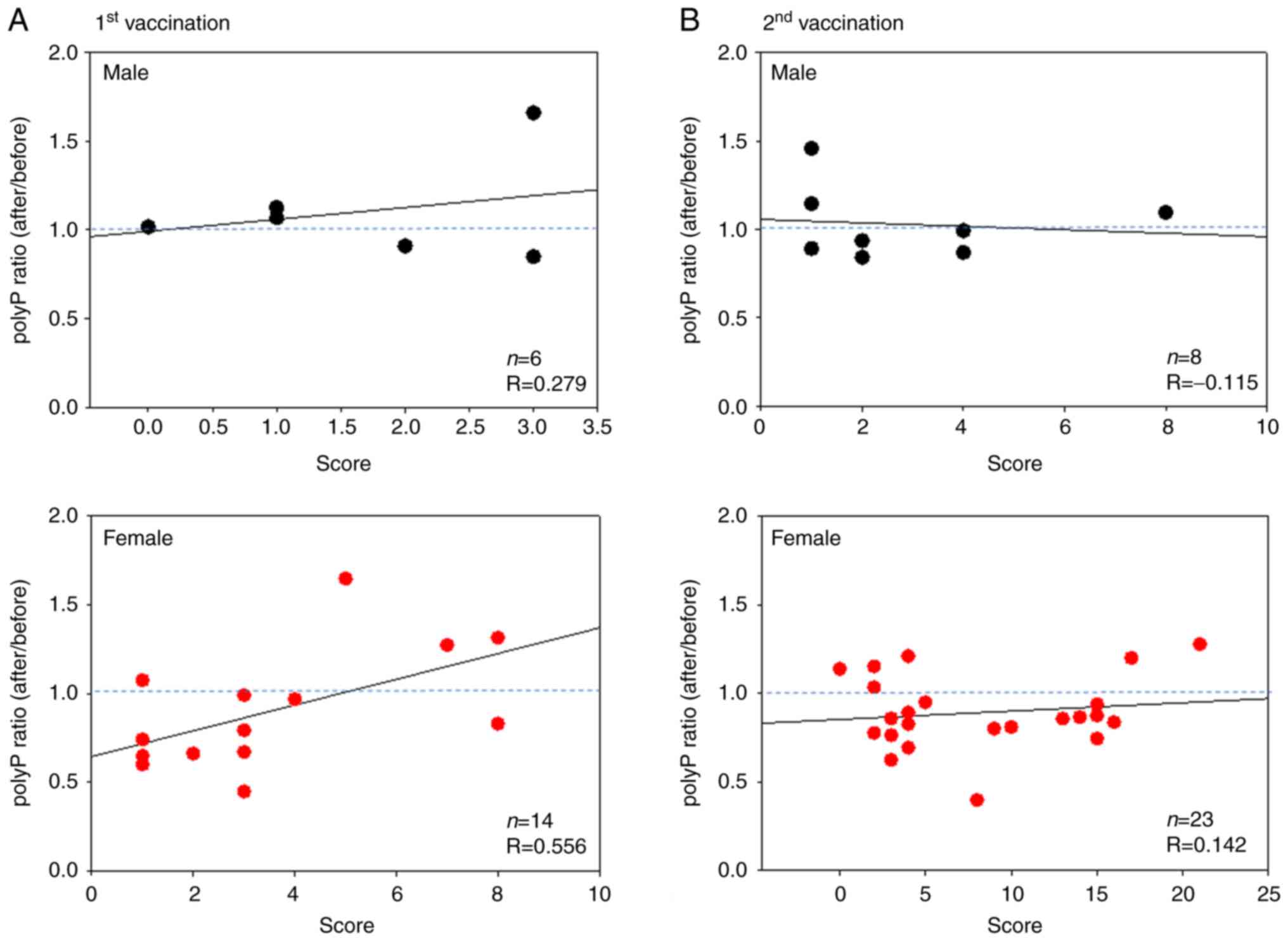
Scatter plots of the relationship between polyP
ratios (pre-vaccination, as well as after the first and second
doses of the vaccinations) and side-effect scores for BNT162b2 are
shown in Fig. 3. In the male
groups, the coefficient correlations (R) were 0.279 and -0.115 for
the first and second vaccinations, respectively. Although the
correlation value was very weak (R=0.142) for the second
vaccination, a moderate correlation (R=0.556) was found for the
first vaccination.
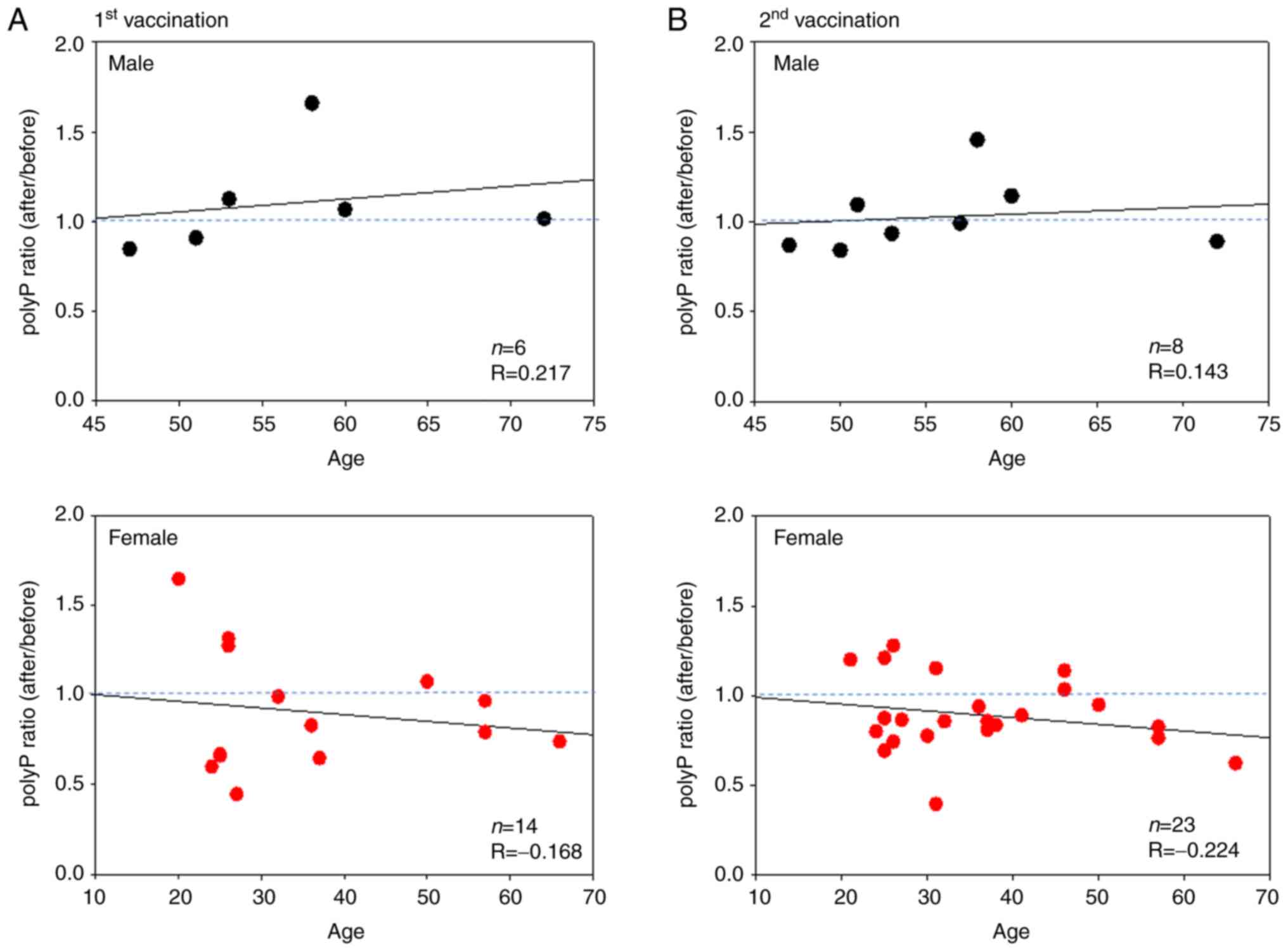
Scatter plots of the relationship between polyP
ratios and age in terms of the Pfizer/BioNTech vaccine (BNT162b2)
are shown in Fig. 4. In the male
group, the correlation coefficients were weak (R=0.217) and very
weak (R=0.143) for the first and second vaccinations, respectively.
In the female groups, the correlations were again negative but very
weak (R=-0.168) and weak (R=-0.224) for the first and second
vaccinations, respectively.
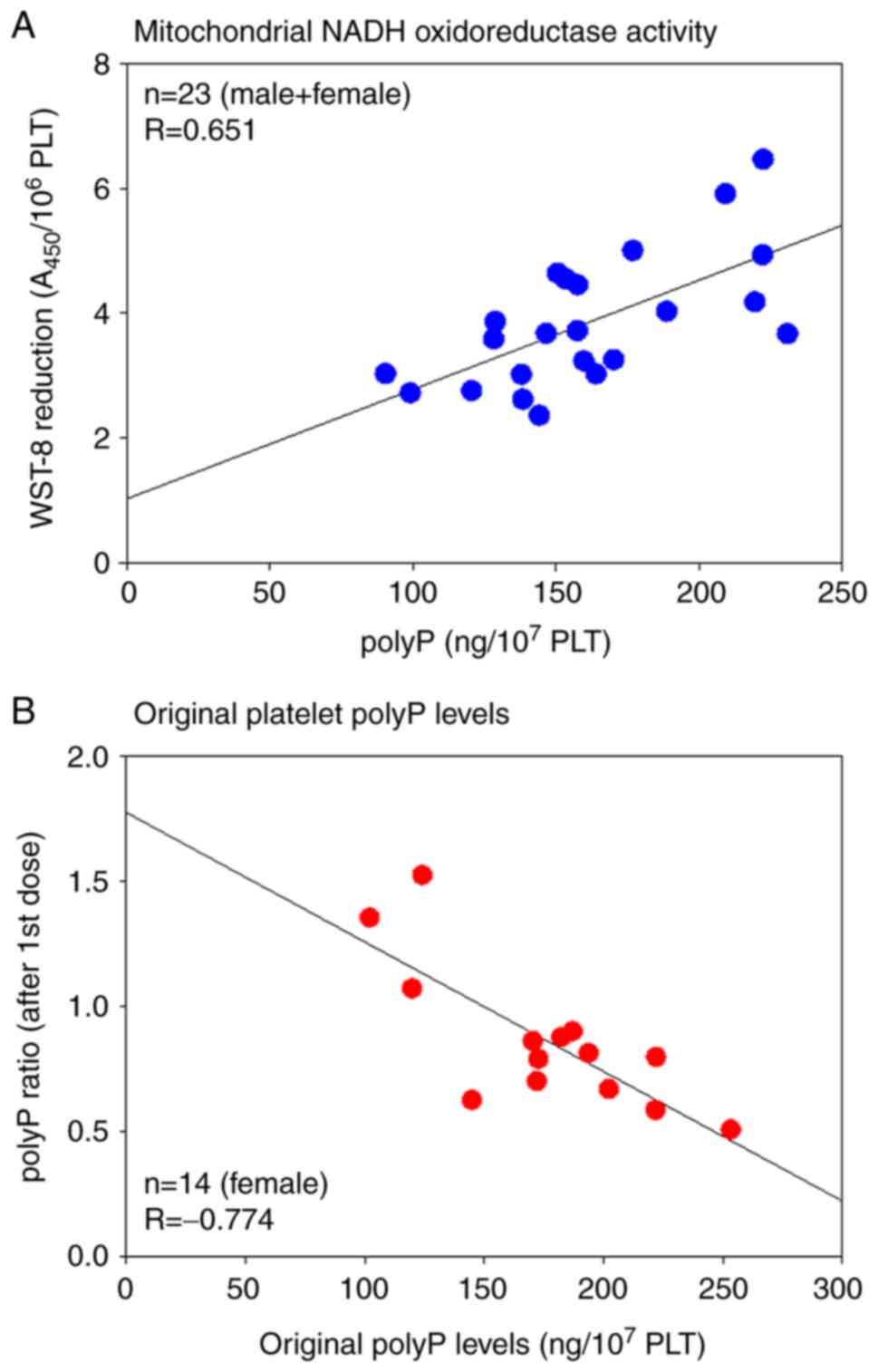
Scatter plots of the relationship between the WST-8
reduction and polyP levels, polyP ratios and pre-vaccination polyP
levels for the Pfizer/BioNTech vaccine (BNT162b2) are shown in
Fig. 5. Although polyP synthesis is
not yet fully understood, several studies have suggested that
mitochondria are involved in polyP synthesis in mammalian cells
(34-36).
To test this hypothesis, the mitochondrial activity and polyP
levels were compared. In the group containing both male and female
subjects, a strong correlation (R=0.651) was found between NADH
activity, which is a proxy for mitochondrial activity, and platelet
polyP levels (Fig. 5A). In the
female group, after the first vaccination, a very strong negative
correlation (R=-0.774) was found between the polyP ratio and the
original platelet polyP levels (Fig.
5B).
The effects of the Moderna vaccine (mRNA-1273) on
polyP levels were examined in a small number of subjects only, due
to its limited supply during our investigation (Table IV). A substantial reduction in
platelet polyP levels was observed in three out of four female
subjects after the first vaccination, whereas the polyP levels
increased in three out of four subjects after the second
vaccination. The oldest female subject was unique, and showed both
a low side-effect score and COVID-19 arm soreness (37), both of which are usually observed
after the first vaccination. A drastic increase in the side-effect
score after the second vaccination was observed in only the
youngest female subject.
| Table IVpolyP ratios and side effect scores
in the patients treated with the Moderna vaccine. |
Table IV
polyP ratios and side effect scores
in the patients treated with the Moderna vaccine.
| Subject | Age range | Sex | Initial dose polyP
ratio | Side-effect
score | Second dose polyP
ratio | Side-effect
score |
|---|
| A | 55-59 | Male | 1.32 | 1 | 1.423 | 2 |
| B | 30-34 | Female | 0.817 | 6 | 1.286 | 24 |
| C | 55-59 | Female | 0.785 | 3 | 0.742 | 8a |
| D | 45-49 | Female | 0.704 | 4a | 1.205 | 8 |
| E | 35-39 | Female | 1.131 | 11 |
Declinedb | - |
Discussion
Prior to this study, it was hypothesized that if a
micro-thrombosis forms in the peripheral tissue, most probably
around the site of vaccine injection in the arm, then platelet
polyP is released (and intra-platelet polyP levels thereafter
decrease) to function as a coagulation factor. However, no symptoms
indicating local thrombocytopenia or purpura were observed upon
visual inspection. Although severe side-effects, classified as
severe fever, headache and pain at the injection site, were
reported in several female subjects after the second vaccination,
it was speculated that the COVID-19 vaccination assessed likely
does not induce platelet-dependent microthrombus formation or
internal bleeding.
Interestingly, the Pfizer/BioNTech vaccine
(BNT162b2) significantly reduced platelet polyP levels after the
first vaccination, along with suppressed side-effects in the
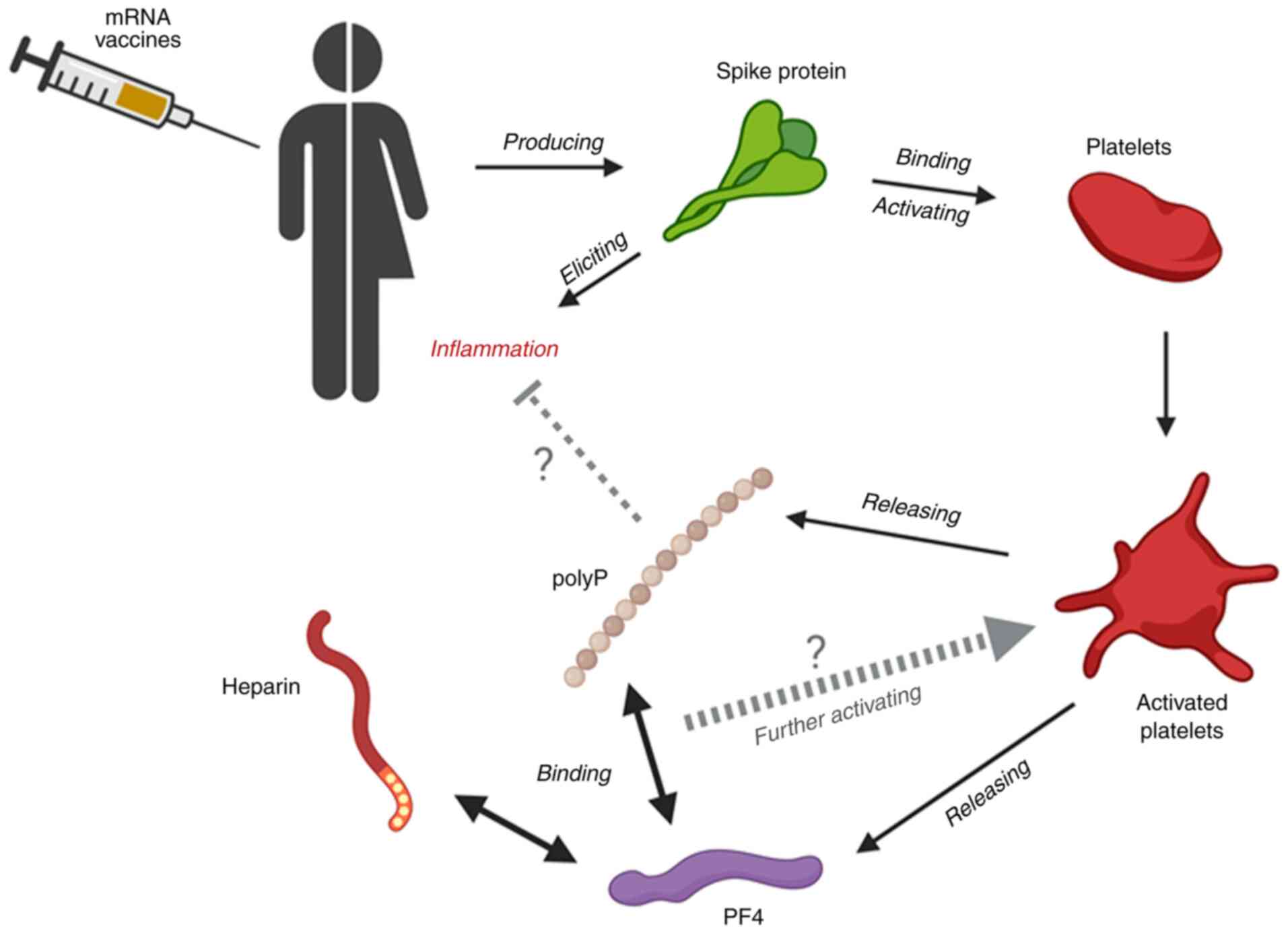
younger female group. Thus, a hypothesis to be considered for
further investigation is that the spike protein produced by the
mRNA vaccines induces inflammation, which is generally more severe
in younger female individuals (38), and that the platelets activated by
this inflammatory response release polyP, along with other
anti-inflammatory molecules, to suppress the inflammation in a
negative feedback loop. This hypothesis is illustrated in Fig. 6. At present, there is no evidence to
suggest how these phenomena are related, necessitating further
investigation to clarify the involvement of polyP in the
inflammatory response initiated by the vaccine.
The dysfunction of this loop system after the second
dose of the vaccine may be due to platelet status. Platelets, which
contain various immunoreceptors, continuously scan for areas of
injury or inflammation as versatile patrollers (39,40).
When platelets recognize intravascular pathogens, they directly
limit pathogen growth and indirectly ensure pathogen clearance
through the activation of immune cells (3,6,40).
Therefore, once platelet signals are received and taken over by
immune cells, inflammation may not be regulated by platelets.
However, if inflammation becomes more severe, platelets may become
dysfunctional. This scenario could be explained by a ‘chicken or
egg’ paradox-like essence, introduced by Portier and Campbell, in
which aberrant platelet activation, which could be caused by
continuous moderate (or severe) inflammation, can lead to
predominantly inflammatory and thrombotic events (40).
It is important to evaluate the expression of
platelet activation markers, such as CD62P, to confirm the release
of polyP by activated platelets. However, to stabilize polyP
levels, it is necessary to incubate blood samples at least
overnight at 22-25˚C (30).
Furthermore, it was difficult to normalize the venipuncture skill,
which can easily influence platelet conditions; thus, platelet
activation was not evaluated in the present study.
The aforementioned explanation assumes that the
reduction of platelet polyP is due to the release of polyP from
activated platelets. Before discussing other possible explanations,
polyP metabolism in platelets should be confirmed. It is generally
accepted that polyP is synthesized from ATP or GTP and degraded by
cellular enzymes in bacteria (26,41,42).
In eukaryotes, this mechanism is not yet fully understood; however,
polyP is suggested to be synthesized and degraded by the same or a
similar mechanism, given that inorganic phosphate (Pi) is
incorporated by a specific transporter into mammalian cells and
used for polyP synthesis in mitochondria non-enzymatically
(24). Although polyP is widely
distributed in the cytoplasm and among organelles, it is most
condensed when stored in dense granules in platelets and released
upon activation (24). Thus, it is
generally accepted that platelet polyP levels are regulated by
three major factors: Activation levels, mitochondrial activity and
extracellular Pi levels.
Since platelets of patients with sepsis or cardiac
shock possess lower mitochondrial NADH levels (43), in this study, the activity of NADH
using was evaluated using WST-8, a tetrazolium salt that is reduced
to soluble formazan (44), and
found a moderate correlation between the activity of this enzyme
and polyP levels. Serum phosphate levels were not examined in this
study, as no declarations for hyperphosphatemia or hypophosphatemia
were obtained from individual participants. Taken together, these
results suggest that basal polyP levels depend on mitochondrial
activity, and influence the rate of reduction of polyP induced by
platelet activation. Considering that platelet activation is a
highly energy-dependent process (45), it is plausible that platelets with
higher mitochondrial activity show stronger immune responses to
released polyP, resulting in polyP reduction.
In clinical observations, SARS-CoV-2 infection
initially produces inflammation, platelet activation, and prominent
elevation of fibrinogen and D-dimer (10-12).
Subsequently, the virus leads to various levels of thrombotic
events and thrombocytopenia (10).
The mRNA vaccines were designed to temporarily produce the spike
protein at lower levels than those observed in SARS-CoV-2 infection
or DNA vaccines. Thus, even though the initial events induced by
SARS-CoV-2 can be reproduced by mRNA vaccines, severe symptoms
observed in COVID-19 patients cannot be produced. However, due to
individual differences in immune activity, vaccines sometimes
induce thrombotic thrombocytopenia (VITT) (46,47).
To date, the incidence of VITT has been much lower in the case of
mRNA vaccines than in DNA vaccines, such as ChAdOx1 nCov-19 and
Ad26.COV2.5, wherein VITT has been reported in various groups and
countries (29).
Recently, the primary mechanism of VITT has been
explained by heparin-induced thrombocytopenia (48). Platelet factor 4 (PF4) released from
activated platelets binds blood heparin to form the complex
(PF4-heparin), which binds to IgG antibodies, then subsequently
binds to platelet FcγRIIa receptors to further activate platelets
to induce thrombosis (49-52).
Interestingly, the authors who reported on this mechanism also
mentioned the possibility that polyanions, such as polyP, may bind
to PF4 to form the PF4-polyP complex and induce thrombosis
(50). This possibility and the
subsequent biological reactions should be further studied.
As described above, the possible anti-inflammatory
effects of polyP, which are thought to be mediated through the
suppression of the complement system, are discussed (53,54).
However, the conflicting possibility that polyP has the potential
to induce thrombosis should be noted. Surprisingly, although the
chain length of polyP may be a matter for further investigation, it
has been reported that polyanions such as polyP can form a complex
with PF4, and that this complex activates platelets, similar to the
PF4-heparin complex (48,50). Platelets also release
anti-inflammatory cytokines, such as IL-4 and IL-10, upon
activation (1); thus, PRP has been
clinically utilized as an anti-inflammatory or a pain-relieving
agent (55). At present, it is
difficult to predict the risk of PF4-polyP complex-induced
thrombosis by the autocrine loop or by exposure to bacterial polyP
after vaccination.
By applying this possibility to the findings of this
study, the biological significance of these conflicting functions
could be interpreted as the unexpected aberrant activation and
destruction of platelets, as well as the uncontrolled exacerbation
of inflammation; thus, the immune system may be protected by the
dual functions of platelets and polyP. However, especially in
younger female subjects who generally exhibit stronger immune
responses, exposure to pathogens or sustained inflammatory
conditions for a long time (i.e., the interval between the first
and second vaccination in this study ~3 weeks) may cause platelets
to lose their ability to detect pathogens and allow the
exacerbation of inflammation, which is recognized as a
side-effect.
There are several limitations in the present study.
First, most allocation guidelines have prioritized high-risk
healthcare workers in most countries (56), including Japan. However, the
vaccination schedule has been strictly controlled by the central
and local governments, limiting the availability of a more diverse
cohort. Second, our limited capacity to attain samples made it
difficult to secure a sufficient number of subjects. As a result,
it was not possible to expand the sample size to what was
originally anticipated. Third, to support the specificity of the
quantification data, it would be better to further examine the
localization and the appearance of DAPI-dependent signals using a
histochemical method that we previously performed for
Ca2+-activated platelets (30,32).
Therefore, further investigation with a larger sample size is
required, along with visualization and image analysis, to reach a
definitive conclusion. However, the present findings regarding sex
differences are essentially consistent with the previous reports of
antibody formation (57-59),
and therefore, this pilot study should be considered important for
the scientific community.
In conclusion, these findings suggest that polyP
released from activated platelets may be involved in the
suppression of severe side-effects after the first vaccination in
young female subjects. This may be related to the result that young
female subjects complained of severe side-effects after the second
vaccination, but not after the first vaccination. To control such
high immune responses, more potent ‘braking’ systems are required
to be balanced with potent ‘accelerating’ systems; PolyP may be a
possible candidate factor.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by Japan Society for the
Promotion of Science (Tokyo, Japan) Grants-in-Aid for Scientific
Research [JSPS KAKENHI (grant no. 21K09932)].
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
TU and TK conceived the study. TK designed the
study. TTs, TTa and KI assisted in the design of the study. TW
analyzed the data. TU, AS, HA, TTs, TW, KI, HK, YK and TK performed
the experiments. HA and YK collected the data. TU, HA, TTs, TTa and
TK wrote the manuscript. TU, TTs, TTa and TK edited the manuscript.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The study design and consent forms for all
procedures (approval no. 2019-0423) were approved by the Ethics
Committee for Human Participants at the Niigata University School
of Medicine (Niigata, Japan) and complied with the Helsinki
Declaration of 1964, as revised in 2013.
Patient consent for publication
Written informed consent was obtained from all the
patients for publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen Y, Zhong H, Zhao Y, Luo X and Gao W:
Role of platelet biomarkers in inflammatory response. Biomark Res.
8(28)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Li C, Li J and Ni H: Crosstalk between
platelets and microbial pathogens. Front Immunol.
11(1962)2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Margraf A and Zarbock A: Platelets in
Inflammation and Resolution. J Immunol. 203:2357–2367.
2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Morrell CN, Aggrey AA, Chapman LM and
Modjeski KL: Emerging roles for platelets as immune and
inflammatory cells. Blood. 123:2759–2767. 2014.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Gómez RM, López Ortiz AO and Schattner M:
New roles of platelets in inflammation. Curr Opin Physiol.
19:99–104. 2021.
|
|
6
|
Gros A, Ollivier V and Ho-Tin-Noé B:
Platelets in inflammation: Regulation of leukocyte activities and
vascular repair. Front Immunol. 5(678)2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Wang MY, Zhao R, Gao LJ, Gao XF, Wang DP
and Cao JM: SARS-CoV-2: Structure, biology, and structure-based
therapeutics development. Front Cell Infect Microbiol.
10(587269)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Cox D: Targeting SARS-CoV-2-platelet
interactions in COVID-19 and vaccine-related thrombosis. Front
Pharmacol. 12(708665)2021.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lichtenberger LM and Vijayan KV: Is
COVID-19-induced platelet activation a cause of concern for
patients with cancer? Cancer Res. 81:1209–1211. 2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Wool GD and Miller JL: The impact of
COVID-19 disease on platelets and coagulation. Pathobiology.
88:15–27. 2021.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Comer SP, Cullivan S, Szklanna PB, Weiss
L, Cullen S, Kelliher S, Smolenski A, Murphy C, Altaie H, Curran J,
et al: COVID-19 induces a hyperactive phenotype in circulating
platelets. PLoS Biol. 19(e3001109)2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Hottz ED, Azevedo-Quintanilha IG, Palhinha
L, Teixeira L, Barreto EA, Pão CRR, Righy C, Franco S, Souza TML,
Kurtz P, et al: Platelet activation and platelet-monocyte aggregate
formation trigger tissue factor expression in patients with severe
COVID-19. Blood. 136:1330–1341. 2020.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Yatim N, Boussier J, Chocron R, Hadjadj J,
Philippe A, Gendron N, Barnabei L, Charbit B, Szwebel TA, Carlier
N, et al: Platelet activation in critically ill COVID-19 patients.
Ann Intensive Care. 11(113)2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
World Health Organization (WHO): WHO
issues its first emergency use validation for a COVID-19 vaccine
and emphasizes need for equitable global access. WHO, Geneva, 2020.
https://www.who.int/news/item/31-12-2020-who-issues-its-first-emergency-use-validation-for-a-covid-19-vaccine-and-emphasizes-need-for-equitable-global-access.
Accessed December 18, 2021.
|
|
15
|
U.S. Food & Drug Administration (FDA):
FDA Approves First COVID-19 Vaccine. FDA, Silver Spring, MD, 2021.
https://www.fda.gov/news-events/press-announcements/fda-approves-first-covid-19-vaccine.
Accessed December 18, 2021.
|
|
16
|
Pharmaceutical Technology: COVID-19
vaccine market set to reach $19.5bn by 2026-register for free
webinar. Pharmaceutical Technology, New York, NY, 2021. https://www.pharmaceutical-technology.com/news/covid-19-vaccine-market-set-to-reach-19-5bn-by-2026-register-for-free-webinar/.
Accessed December 18, 2021.
|
|
17
|
Nikkei Asia: Status of vaccinations around
the world. Nikkei Inc., Tokyo, 2021. https://vdata.nikkei.com/en/newsgraphics/coronavirus-vaccine-status/.
Accessed December 18, 2021.
|
|
18
|
Corbett KS, Edwards DK, Leist SR, Abiona
OM, Boyoglu-Barnum S, Gillespie RA, Himansu S, Schäfer A, Ziwawo
CT, DiPiazza AT, et al: SARS-CoV-2 mRNA vaccine design enabled by
prototype pathogen preparedness. Nature. 586:567–571.
2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Jalkanen P, Kolehmainen P, Häkkinen HK,
Huttunen M, Tähtinen PA, Lundberg R, Maljanen S, Reinholm A,
Tauriainen S, Pakkanen SH, et al: COVID-19 mRNA vaccine induced
antibody responses against three SARS-CoV-2 variants. Nat Commun.
12(3991)2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Menni C, Klaser K, May A, Polidori L,
Capdevila J, Louca P, Sudre CH, Nguyen LH, Drew DA, Merino J, et
al: Vaccine side-effects and SARS-CoV-2 infection after vaccination
in users of the COVID Symptom Study app in the UK: A prospective
observational study. Lancet Infect Dis. 21:939–949. 2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Morrissey JH, Choi SH and Smith SA:
Polyphosphate: An ancient molecule that links platelets,
coagulation, and inflammation. Blood. 119:5972–5979.
2012.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Travers RJ, Smith SA and Morrissey JH:
Polyphosphate, platelets, and coagulation. Int J Lab Hematol. 37
(Suppl 1):S31–S35. 2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Weitz JI and Fredenburgh JC: Platelet
polyphosphate: The long and the short of it. Blood. 129:1574–1575.
2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Abbasian N and Harper MT: High
extracellular phosphate increases platelet polyphosphate content.
Platelets. 32:992–994. 2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Baker CJ, Smith SA and Morrissey JH:
Polyphosphate in thrombosis, hemostasis, and inflammation. Res
Pract Thromb Haemost. 3:18–25. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Morrissey JH: Polyphosphate: A link
between platelets, coagulation and inflammation. Int J Hematol.
95:346–352. 2012.PubMed/NCBI View Article : Google Scholar
|
|
27
|
World Medical Association. World Medical
Association Declaration of Helsinki: Ethical principles for medical
research involving human subjects. JAMA. 310:2191–2194.
2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Kurihara C: The 2013 revision of the
declaration of Helsinki: Challenges for the Next 50 Years. Rinsho
Yakuri/Jap J Clin Pharmacol Ther. 45:41–51. 2014.
|
|
29
|
The Japan Times: Japan's COVID-19 vaccine
plan prioritizes health care workers and older residents. The Japan
Times, Ltd., Tokyo, 2020. https://www.japantimes.co.jp/news/2020/12/25/national/japan-vaccine-older-people/.
Accessed December 18, 2021. (This page is now missing).
|
|
30
|
Watanabe T, Kitamura Y, Aizawa H, Masuki
H, Tsujino T, Sato A, Kawabata H, Isobe K, Nakata K and Kawase T:
Fluorometric Quantification of human platelet polyphosphate using
4',6-diamidine-2-phenylindole Dihydrochloride: Applications in the
Japanese population. Int J Mol Sci. 22(7257)2021.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Vollmar B, Slotta JE, Nickels RM, Wenzel E
and Menger MD: Comparative analysis of platelet isolation
techniques for the in vivo study of the microcirculation.
Microcirculation. 10:143–152. 2003.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Sato A, Aizawa H, Tsujino T, Isobe K,
Watanabe T, Kitamura Y and Kawase T: Fluorescent Cytochemical
detection of polyphosphates associated with human platelets. Int J
Mol Sci. 22(1040)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Aschar-Sobbi R, Abramov AY, Diao C,
Kargacin ME, Kargacin GJ, French RJ and Pavlov E: High sensitivity,
quantitative measurements of polyphosphate using a new DAPI-based
approach. J Fluoresc. 18:859–866. 2008.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Baev AY, Angelova PR and Abramov AY:
Inorganic polyphosphate is produced and hydrolyzed in F0F1-ATP
synthase of mammalian mitochondria. Biochemical J. 477:1515–1524.
2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Lynn WS and Brown RH: Synthesis of
polyphosphate by rat liver mitochondria. Biochem Biophys Res
Commun. 11:367–371. 1963.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Müller WEG, Schröder HC and Wang X:
Inorganic polyphosphates as storage for and generator of metabolic
energy in the extracellular matrix. Chem Rev. 119:12337–12374.
2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Blumenthal KG, Freeman EE, Saff RR,
Robinson LB, Wolfson AR, Foreman RK, Hashimoto D, Banerji A, Li L,
Anvari S and Shenoy ES: Delayed large local reactions to mRNA-1273
Vaccine against SARS-CoV-2. N Engl J Med. 384:1273–1277.
2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Chen PW, Tsai ZY, Chao TH, Li YH, Hou CJ
and Liu PY: Addressing vaccine-induced immune thrombotic
thrombocytopenia (VITT) Following COVID-19 vaccination: A
Mini-review of practical strategies. Acta Cardiol Sin. 37:355–364.
2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Guo L and Rondina MT: The era of
thromboinflammation: Platelets are dynamic sensors and effector
cells during infectious diseases. Front Immunol.
10(2204)2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Portier I and Campbell RA: Role of
platelets in detection and regulation of infection. Arterioscler
Thromb Vasc Biol. 41:70–78. 2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Simbulan-Rosenthal CM, Carney BC, Gaur A,
Moghe M, Crooke E, Moffatt LT, Shupp JW and Rosenthal DS: Inorganic
polyphosphates are important for cell survival and motility of
human skin keratinocytes and play a role in wound healing. In:
Contemporary Topics about Phosphorus in Biology and Materials.
Churchill DG, Sikirić MD, Čolović B and Milhofer HF (eds).
IntechOpen, London, 2019.
|
|
42
|
Simbulan-Rosenthal CM, Gaur A, Sanabria
VA, Dussan LJ, Saxena R, Schmidt J, Kitani T, Chen YS, Rahim S,
Uren A, et al: Inorganic polyphosphates are important for cell
survival and motility of human skin keratinocytes. Exp Dermatol.
24:636–639. 2015.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Protti A, Fortunato F, Artoni A, Lecchi A,
Motta G, Mistraletti G, Novembrino C, Comi GP and Gattinoni L:
Platelet mitochondrial dysfunction in critically ill patients:
Comparison between sepsis and cardiogenic shock. Crit Care.
19(39)2015.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Ishiyama M, Miyazono Y, Sasamoto K, Ohkura
Y and Ueno K: A highly water-soluble disulfonated tetrazolium salt
as a chromogenic indicator for NADH as well as cell viability.
Talanta. 44:1299–1305. 1997.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Aibibula M, Naseem KM and Sturmey RG:
Glucose metabolism and metabolic flexibility in blood platelets. J
Thromb Haemost. 16:2300–2314. 2018.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Arepally GM and Ortel TL: Vaccine-induced
immune thrombotic thrombocytopenia: What we know and do not know.
Blood. 138:293–298. 2021.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Huynh A, Kelton JG, Arnold DM, Daka M and
Nazy I: Antibody epitopes in vaccine-induced immune thrombotic
thrombocytopaenia. Nature. 596:565–569. 2021.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Novak N, Tordesillas L and Cabanillas B:
Adverse rare events to vaccines for COVID-19: From hypersensitivity
reactions to thrombosis and thrombocytopenia. Int Rev Immunol: Jul
12, 2021 (Epub ahead-of print).
|
|
49
|
Arepally GM and Padmanabhan A:
Heparin-induced thrombocytopenia. Arterioscler Thromb Vasc Biol.
41:141–152. 2021.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Cines DB, Yarovoi SV, Zaitsev SV, Lebedeva
T, Rauova L, Poncz M, Arepally GM, Khandelwal S, Stepanova V, Rux
AH, et al: Polyphosphate/platelet factor 4 complexes can mediate
heparin-independent platelet activation in heparin-induced
thrombocytopenia. Blood Adv. 1:62–74. 2016.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Prechel MM and Walenga JM: Emphasis on the
Role of PF4 in the incidence, pathophysiology and treatment of
heparin induced thrombocytopenia. Thromb J. 11(7)2013.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Rauova L, Zhai L, Kowalska MA, Arepally
GM, Cines DB and Poncz M: Role of platelet surface PF4 antigenic
complexes in heparin-induced thrombocytopenia pathogenesis:
Diagnostic and therapeutic implications. Blood. 107:2346–2353.
2006.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Conway EM: Polyphosphates and complement
activation. Front Med (Lausanne). 6(67)2019.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Wat JM, Foley JH, Krisinger MJ, Ocariza
LM, Lei V, Wasney GA, Lameignere E, Strynadka NC, Smith SA,
Morrissey JH and Conway EM: Polyphosphate suppresses complement via
the terminal pathway. Blood. 123:768–776. 2014.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Kuffler DP: Variables affecting the
potential efficacy of PRP in providing chronic pain relief. J Pain
Res. 12:109–116. 2019.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Ubel PA: Vaccinating health care
employees-do they all deserve early access? N Engl J Med.
384(e39)2021.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Ciarambino T, Para O and Giordano M:
Immune system and COVID-19 by sex differences and age. Womens
Health (Lond). 17(17455065211022262)2021.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Lo Sasso B, Giglio RV, Vidali M, Scazzone
C, Bivona G, Gambino CM, Ciaccio AM, Agnello L and Ciaccio M:
Evaluation of Anti-SARS-Cov-2 S-RBD IgG antibodies after COVID-19
mRNA BNT162b2 vaccine. Diagnostics (Basel). 11(1135)2021.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Pellini R, Venuti A, Pimpinelli F, Abril
E, Blandino G, Campo F, Conti L, De Virgilio A, De Marco F, Di
Domenico EG, et al: Initial observations on age, gender, BMI and
hypertension in antibody responses to SARS-CoV-2 BNT162b2 vaccine.
EClinicalMedicine. 36(100928)2021.PubMed/NCBI View Article : Google Scholar
|