1. Introduction
Endometriosis is a common estrogen-dependent
inflammatory disorder, which affects ~10% of reproductive-age women
(1). Endometriosis is defined by
the presence of endometrial-like benign lesions located outside of
the uterus (1). This disease is a
heterogeneous disease that can present in different forms,
including superficial peritoneal disease, ovarian endometrioma and
deep infiltrating endometriosis (2). Endometriosis has been reported as a
complex and multifactorial disease, which is characterized by
estrogen dominance, increased cytokine levels, and innate and
adaptive immune system activation (2,3).
Endometriosis is caused by endometrial cell
engraftment outside of the uterus and is characterized by periodic
hemorrhage from ectopic endometriotic lesions (3). Epidemiologic studies have reported
that the risk of endometriosis is associated with increased
exposure to menstruation (e.g., early age at menarche, shorter
menstrual cycle length, longer menstrual flow duration, long
menstrual period or reduced parity) (4). Typically, modern females marry later
in life, have ≤2 children, and experience much shorter
breastfeeding periods, which results in more frequent menstrual
cycles (5). Females today
menstruate approximately eight times more often than females in
hunter-gatherer societies (i.e., ~400 vs. 50 menstrual cycles in
their lifetime) (6). Menstruation
is considered an inflammatory response with cyclic fluctuations in
female steroid hormone levels and results in the production and
release of inflammatory mediators, such as cytokines and
prostaglandins (7,8). A surge in inflammatory mediators
causes immune cell activation and recruitment, which results in
further stressor production (e.g., cytokines, chemokines,
angiogenesis factors and growth factors) (8). Multiple stressors are produced by
different immune cells, mainly macrophages, and the regurgitated
endometrial tissue (9,10) is eventually terminated by the host's
immune system and immunoregulatory mechanisms. Therefore, a higher
frequency of menstruation may increase the risk of developing
endometriosis.
Certain previous studies have suggested a role for
microbes in the female reproductive tract in endometriosis
(8,11). The female reproductive tract
microbiota constitutes a defensive barrier to prevent infections,
serves an important role in each of the structural, inflammatory,
immune, metabolic, and endocrine systems, and may act as a key link
between inflammation and immunity (12). Previous studies have suggested that
the microbiota may be important for the prevention of
non-communicable diseases (7,13).
Furthermore, the abundance and composition of the gut and
reproductive tract microbiota differ between females with and
without endometriosis (8,14). Therefore, quantitative and
qualitative imbalances between pathogenic bacteria and
non-pathogenic environmental microbes (i.e., dysbiosis) may be
involved in endometriosis pathogenesis (8). Dysfunction of the inflammatory and
immune systems is a crucial aspect of endometriosis
pathogenesis.
Thereby, both deleterious inflammations due to
frequent menstrual cycles (an incessant attack from intrinsic
stimuli) and pathogens or pathogenic microbes (an attack from
extrinsic stimuli) are controlled by the host defense system. This
review summarizes current knowledge about dysbiosis in
endometriosis and discusses how this imbalance causes inappropriate
immune responses, which can lead to the development of disease.
2. Search strategy and selection
criteria
A literature search was performed to identify
relevant studies. The PubMed and Google Scholar electronic
databases were searched for literature published up to the 31st
March 2022, combining the following keywords: ‘Endometriosis’,
‘microbiome’, ‘dysbiosis’, ‘retrograde menstruation’,
‘inflammation’ and ‘immunity’. The search strategy combined these
keywords with the Boolean operators ‘and’ and ‘or’, as described in
Table I. Inclusion criteria were
studies which specifically focused on the microbiome in
endometriosis and included the publication of peer-reviewed
original articles and reference lists in review articles. The
exclusion criteria were duplicated studies, non-English language
publications, letters to the editor, poster presentations, and
unrelated literature. The identified articles were initially
assessed for eligibility and then subsequently, the full-text of
the articles was assessed. The first identification phase included
an electric database search, hand search (manually searching for
necessary documents), and the generation of a reference list of
collected articles and review articles to identify additional
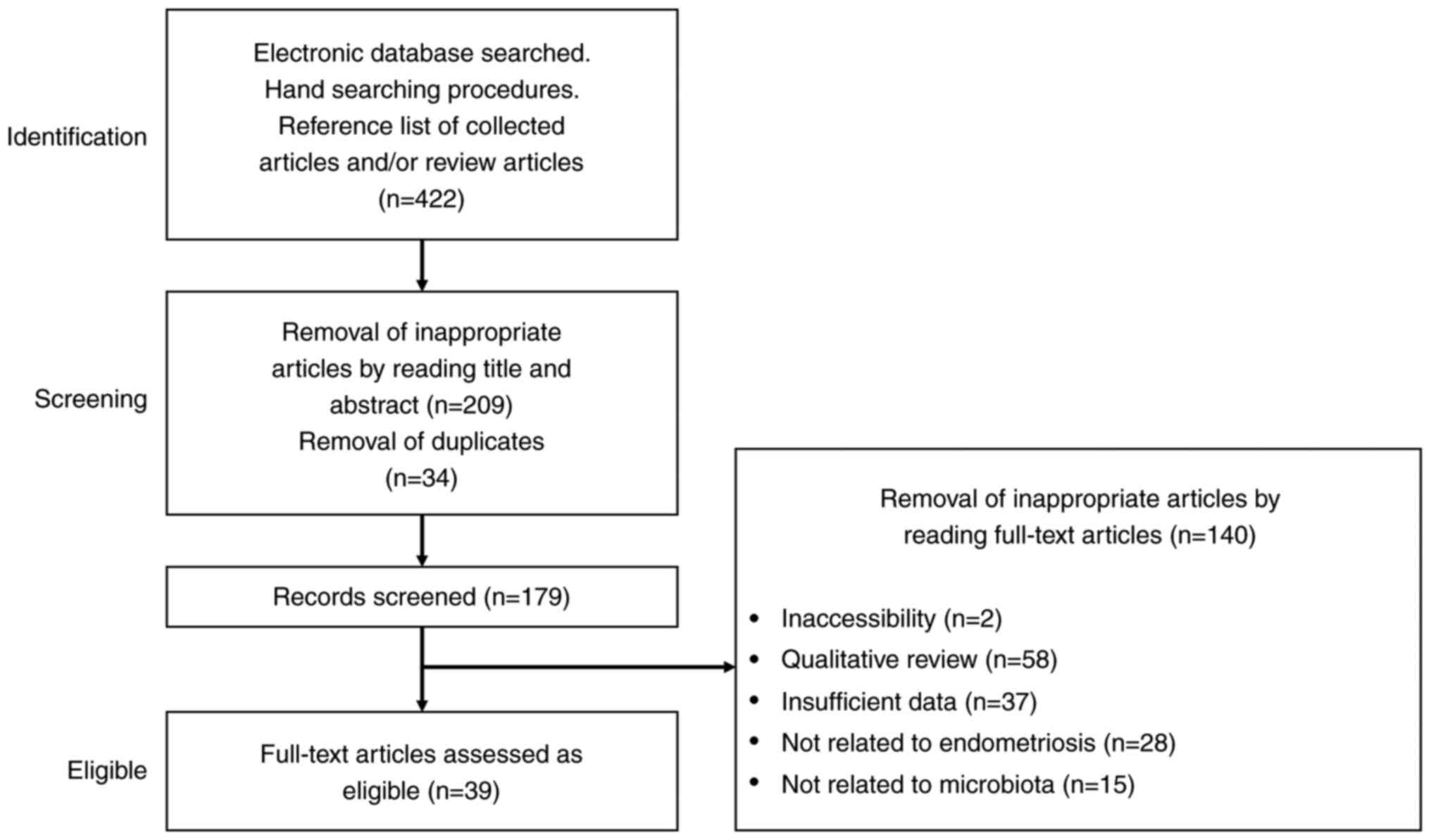
relevant articles (Fig. 1).
Duplicates were removed during the second, screening, phase. Titles
and abstracts were read to remove inappropriate papers. The final
eligible phase included full-text articles assessed as
eligible.
 | Table IThe search strategy. |
Table I
The search strategy.
| Search | Keyword and search
term combinations |
|---|
| 1 | Endometriosis |
| 2 | Microbiome OR
Microbiota OR |
| | Pathogenic OR
Probiotics |
| 3 | Dysbiosis AND
(Imbalance OR Impairment) |
| 4 | Retrograde
menstruation OR Menses |
| 5 | Inflammation OR
Inflammatory |
| 6 | Immunity OR
Immune |
| 7 | Search 1 AND search
2 |
| 8 | Search 1 AND search
3 |
| 9 | Search 1 AND search
2 AND search 3 |
| 10 | Search 1 AND search
4 AND search 5 |
| 11 | Search 1 AND search
4 AND search 6 |
3. Results
Identification of studies
Searches of the PubMed and Google Scholar electronic
databases yielded 422 literature citations (Fig. 1). After removing duplicates, 179
records were identified, of which 140 were excluded and 39 met the
eligibility criteria.
Role of the microbiota in the gut and
female reproductive tract
This subsection summarizes the mechanisms underlying
the effect of microbiome dysbiosis with a focus on endometriosis.
Mucosal barriers serve crucial roles in the defense against
pathogens or pathogenic microbes (12). The gut, skin, lungs, mouth, vagina,
etc., contain certain non-pathogenic commensal microbes (12). The human microbiota positively
affects physical health by maintaining structural, immunological,
metabolic and neurological homeostasis (12). Mammals, including humans, have
co-evolved with host-resident microorganisms in a symbiotic
relationship (15). Microbiota
co-evolving with humans supported the development of a symbiotic
relationship and accelerated immune system function (16). Furthermore, the microbiota has an
essential role in the endocrine systems of the gut and reproductive
systems by interacting with estrogen and other hormones via a
healthy estrogen-gut axis (17,18).
Role of the gut microbiota in
non-communicable disease development
The commensal microbiome sustains the
host-microbiota symbiotic relationship and contributes to numerous
interactions which affect human health, including the synthesis and
absorption of essential nutrients to maintain energy homeostasis;
maintenance of intestinal mucosal function and structural
integrity; protection against pathogenic microbes or harmful
agents; production of antimicrobial peptides that affect the
epigenome (15); maturation,
development and functions of the innate and adaptive immune system;
modulation of host brain function and cognitive behavior;
regulation of estrogen modulated by estrogen-metabolizing enzyme
production; and dynamic modulation of the metabolome profile
(19,20). The microbiota is recognized by
innate immune receptors, in particular, toll-like receptors (TLRs)
(21,22). Gut epithelial cell TLRs are the
signaling pathways which can implement host defense mechanisms
against invading microbes, and they not only eliminate pathogens
via increased immunosurveillance (21), but also establish host-microbial
symbiosis to maintain microorganism-induced homeostasis (22). Therefore, the microbial
contamination theory or hygiene hypothesis, which claims the
positive effect of exposure to non-pathogenic environmental
microbes and pathogenic bacteria in early-life on human health, has
attracted attention (7,8,13,23).
The exposure of humans to non-pathogenic environmental microbes or
pathogenic bacteria begins prenatally, and continues during
parturition and postnatally (20).
Generally, early-life exposures to microbial diversity may be
associated with a reduced risk of developing allergies, including
atopy and asthma (24). In
contrast, abundant specific bacterial taxa which affect the gut
microbiota composition are associated with disease susceptibility
in adult life, including inflammatory bowel diseases (25,26),
metabolic disorders (e.g., obesity, gestational diabetes mellitus,
type II diabetes) (25),
neurological diseases (e.g., autism) (26), and reproductive and endocrine
disorders (e.g., pregnancy complications, adverse pregnancy
outcomes, polycystic ovary syndrome) (27). For example, gut microbiome
alteration increases inflammatory bowel disease susceptibility, as
the relative abundance of beneficial organisms (e.g.,
Lachnospiraceae, Bifidobacterium species, Roseburia,
Sutterella and Faecalibacterium prausnitzii)
decreases whereas the relative abundance of pathogens (e.g.,
Proteobacteria, Fusobacteria species and Ruminococcus
gnavus) increase (28).
Furthermore, imbalances in the composition of the gut microbiota
(i.e., dysbiosis) favor inflammation via specific inflammatory
immune cell recruitment, proinflammatory cytokine production and
compromised immunosurveillance (8,29).
Diseases affecting the immune and inflammatory system, such as
inflammatory bowel disease and metabolic disorders, are believed to
be related to a decreased microbial community diversity. Similarly,
a lifelong risk of non-communicable inflammatory diseases,
including reproductive pathologies, may result from early-life
microbial exposures (26).
Role of the reproductive tract microbiota in
obstetric, gynecological, and reproductive diseases. The
healthy human vagina is colonized by numerous types of
microorganisms and is characterized by the beneficial microbiota,
Lactobacillus. The presence of a vaginal microbiota
dominated by Lactobacillus maintains an acidic vaginal
environment with a pH of <4.5, which prevents the growth of
pathogenic bacteria (30). This can
protect the host from pathogenic microbial infections such as
Gardnerella species (31)
and Neisseria gonorrhoeae (32). Furthermore, the upper female
reproductive tract, consisting of the uterus, fallopian tubes,
ovaries and peritoneum, is not a sterile environment and
demonstrates a highly diverse and unique microbiota. Therefore, the
microbial community composition differs between the uterus and the
vagina (33-35).
Commensal microbes produce biological resources and other microbes
can use valuable metabolic resources to persist in the ecosystem
via resource-sharing/competition and cross-feeding, which limits
the availability of resources to neighboring pathogenic bacteria
and inhibits pathogenic microbial growth with symbiotic microbes
(36). This indicates that
pathogenic bacteria can survive and even grow with a decreased
commensal microbial diversity and an abundance of microbial
taxa.
Imbalances in the composition of the reproductive
tract microbiota can cause certain obstetric, gynecological and
reproductive diseases. For example, a decrease in the abundance of
the typical Lactobacillus and an abnormal increase in
opportunistic pathogenic bacterial diversity cause pathologies,
such as bacterial vaginosis, which is the most common cause of
vaginal inflammation and infection (i.e., vaginitis) (37,38).
Bacteria associated with bacterial vaginosis, such as
Gardnerella, Prevotella and Bacteroides,
induce increased levels of proinflammatory cytokine and mucosal
epithelial barrier disruption (8,37).
Moreover, elevated levels of inflammatory cytokine confer an
increased risk for multiple gynecologic diseases, including
endometritis, pelvic inflammatory disease and infertility (8,37).
Additionally, patients with pelvic inflammatory disease are
generally at increased risk of developing endometriosis (39). Furthermore, the lack of
Lactobacillus-dominated microbiota species has been reported
to be associated with poor fertility treatment outcomes (i.e., poor
in vitro fertilization outcomes, with low fertilization and
pregnancy rates) (35). Changes in
microbiota distribution from eubiosis to dysbiosis can affect human
health; however, the timing (i.e., pre-natal, intra-natal,
post-natal or over time to the adult life) of these changes remains
unknown. Research in this field is only beginning to apply
microbiota analysis in the clinical setting, and its physiological
and pathological roles in human reproduction and disease remain to
be fully understood (34).
The clinicaltrials.gov electronic database was searched
using the keywords ‘endometriosis’ and ‘microbiome’. The electronic
search yielded a total of six clinical trials, in which recruitment
was completed, currently being performed, and not yet started in
two, one and three studies, respectively. Currently, clinical
studies in ‘Microbiome and immunologic analysis in females with
endometriosis (NCT04159740)’ and ‘Establishment of the human
intestinal and salivary microbiota biobank in gynecological
diseases (NCT04698109)’ are currently being performed.
The role of the immune system is
mediated by the microbiota in endometriosis
Immune cells, which are essential reproductive tract
microenvironment components, secrete numerous cytokines and
chemokines (40). Both innate
[e.g., macrophages, neutrophils, dendritic cells and natural killer
(NK) cells] and adaptive immune cells (e.g., T cells and B cells)
contribute to regulation of tissue inflammation, immune cell
recruitment and resolution, and the host defense response. Pelvic
endometriosis is thought to arise from endometrial cell
implantation through retrograde menstruation (2). Escherichia coli bacteria in
menstrual blood and endotoxins in the peritoneal fluid are
bioaccumulative contaminants which have been previously reported in
females with endometriosis (41).
Endometrial cell fragments that are shed during menstruation
produce a damage-associated molecular pattern (DAMP) through the
specialized pattern recognition receptors (e.g., TLRs) and activate
inflammatory cytokine production, such as interleukin-1β (IL-1β),
to trigger inflammation (9).
Furthermore, DAMPs activate innate and adaptive immune cells, such
as neutrophils, mast cells and Th17 cells, initiate immune cell
response, and promote endometrial cell adhesion and angiogenesis
(9). Particular TLR ligands (e.g.,
lipopolysaccharide, nucleic acids, flagellin, or zymosan of both
commensal microbiota and pathogenic microbiota) are sensed by the
immune system via the pattern recognition receptors (42). TLR-4-mediated nuclear factor κβ
activation may be the main factor which affects endometriosis
development (41). Over time,
persistent immune stimuli may drive a vicious cycle of
inflammation, adhesion and angiogenesis, which in turn facilitates
ectopic endometrium implantation and growth (8). The immune system fights against
bacterial infection but triggers an inflammatory response that
damages reproductive tract tissue. The excessive inflammatory
response may lead to endometriosis through host immune dysfunction.
Therefore, the immune system mediated by the
microbiota-gut-reproductive tract axis is thought to serve an
important role in developing endometriosis.
Role of the gastrointestinal tract
microbiome
This subsection summarizes the role of the
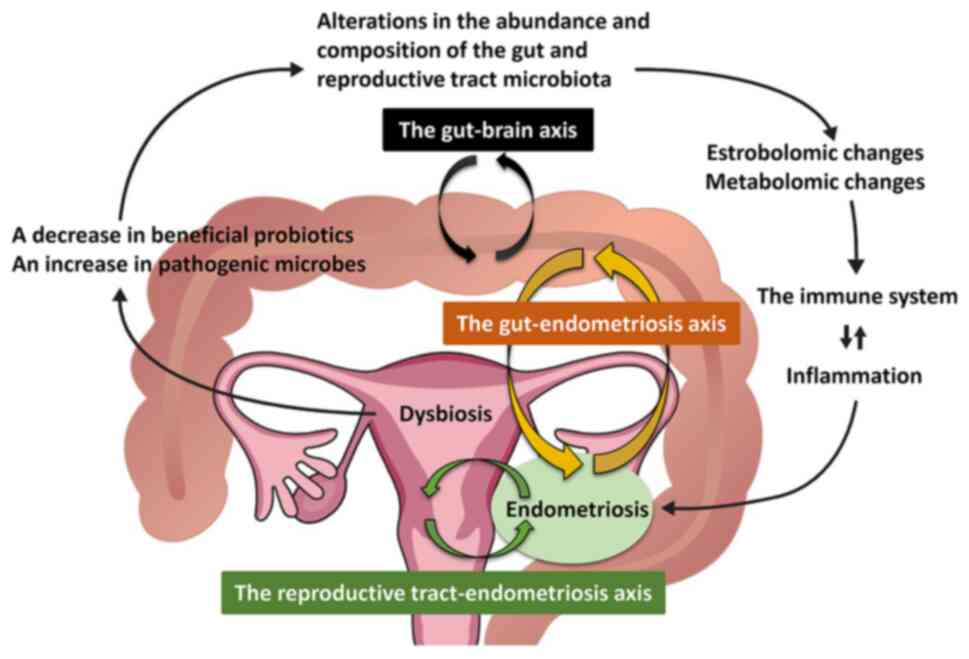
gastrointestinal tract microbiome in endometriosis (Fig. 2). Data from mice and nonhuman
primates are included, as microbiome research in females with
endometriosis is still in its early stages (Table II). Previous in vivo studies
have reported the influence of gut microbiota on animals with
endometriosis. Mouse models of endometriosis demonstrated an
increased ratio of Firmicutes to Bacteroidetes and an increased
number and proportion of certain gut microbes (e.g.,
Ruminococcaceae, Bifidobacterium and
Parasutterella generae) after endometrial fragment
transimplantation (43). Moreover,
a study which used rhesus macaques reported lower numbers of
Lactobacillus bacteria and higher Gram-negative microbial loads in
the endometriosis group compared with that in the non-endometriosis
group (44). Furthermore, rhesus
macaque with endometriosis were reported to have an increased risk
of gastrointestinal inflammation (44). This finding was consistent with
clinical observations that endometriosis is associated with an
increased risk of gastrointestinal disorders, such as irritable
bowel syndrome (45). Gut
microbiota diversity and abundance decreased in animals with
endometriosis compared with healthy controls (46). Further, previous studies have
reported an increased absolute number of Gardnerella,
Streptococcus, Enterococci and Escherichia
coli in the vagina of females with endometriosis compared with
healthy females, which indicated an increased proportion of
potentially pathogenic microbes and a reduced microbiome diversity
(47,48). Therefore, the type and abundance of
gut microbiota differ between females with and without
endometriosis (49).
| Table IISummary of alterations in the
microbiota in endometriosis. |
Table II
Summary of alterations in the
microbiota in endometriosis.
| First author/s,
year | Study design | Aims | Results and
summary | (Refs.) |
|---|
| Bailey et
al, 2002 | In vitro
experimental analysis | To identify an
altered profile of intestinal microflora in female rhesus monkeys
with endometriosis. | A significant
decrease in Lactobacilli and an increase in gram-negative bacteria
in rhesus monkeys with endometriosis. A higher prevalence of
intestinal inflammation in monkeys with endometriosis in comparison
to healthy controls. The microflora was affected by
endometriosis-associated intestinal inflammation. | (44) |
| Seaman et
al, 2008 | A case-control
study | To determine
whether women with endometriosis were more likely to receive a
diagnosis of pelvic inflammatory disease or irritable bowel
syndrome than women without endometriosis. | Women with
endometriosis were more likely to develop pelvic inflammatory
disease or irritable bowel syndrome than women without
endometriosis (Odds ratio 3.5). | (45) |
| Khan et al,
2010 | A retrospective
empirical research | To test the
hypothesis that bacterial contamination of menstrual blood could be
a local biologic event in the development of endometriosis | Elevated
Escherichia coli contamination and endotoxin levels were identified
in menstrual fluid or peritoneal fluid in women with endometriosis
compared with control women. Endotoxin levels in menstrual fluid
and peritoneal fluid in women with endometriosis may promote
Toll-like receptor 4-mediated growth of endometriosis. | (41) |
| Attaman et
al, 2014 | In vitro
experimental analysis | To elucidate the
anti- inflammatory impact of polyunsaturated fatty acids in mouse
endometriosis models. | Elevated systemic
polyunsaturated fatty acid levels suppressed immune, angiogenic,
and growth factors in endometriosis animal models. Omega-3
polyunsaturated fatty acids may suppress the establishment of
endometriosis-like lesions through anti-inflammatory effects. | (69) |
| Hopeman et
al, 2015 | A cross-sectional
study | To elucidate the
relationship between serum polyunsaturated fatty acids and the risk
of developing endometriosis. | A cross-sectional
study of serum polyunsaturated fatty acids and clinical data from
205 women undergoing in vitro fertilization suggested that
low eicosapentaenoic acid levels may be a risk factor for
endometriosis. | (68) |
| Khan et al,
2016 | A case-controlled
biological study | To investigate
microbial colonization in intrauterine environment and cystic fluid
of women with and without endometriosis by molecular approach. | Molecular detection
of intrauterine microbial colonization by 16S metagenome assay
detected an increase in Streptococcaceae and Staphylococaceae in
the cystic fluid of ovarian endometrioma comparing to that in
cystic fluid of non-endometrioma cysts. Sub-clinical infection was
identified in intrauterine environment and in the cystic fluid of
ovarian endometrioma. | (54) |
| Dols et al,
2016 | A case-controlled
biological study | To identify
microbiota composition in the vagina of women with and without
bacterial vaginosis. | Analysis of the
bacterial communities by 16S rRNA amplicon sequencing revealed an
increase in Gardnerella vaginalis, Lachnospiraceae, and
Leptotrichiaceae in women with bacterial vaginosis. A simple
molecular test may serve as a practical diagnostic method for the
assessment of female reproductive tract dysbiosis. | (57) |
| Shi et al,
2017 | A review
article | To outline the
interaction between the gut microbiota and the immune system and
provide a reference for future studies. | Inflammation caused
by abnormal immune responses may modulate the gut microbiome
composition, resulting in intestinal diseases. This review
summarizes the interaction between the gut microbiota and the
immune system. | (60) |
| Khan et al,
2018 | A retrospective
empirical research | To measure the
endotoxin levels in the menstrual fluid and peritoneal fluid in
women with or without endometriosis. | The menstrual blood
of women with endometriosis was contaminated with pathogenic
bacteria including Escherichia coli. Lipopolysaccharide stimulates
the growth of endometriosis via the LPS/TLR4 cascade- mediated
pro-inflammatory response. A new concept called the ‘bacterial
contamination hypothesis’ in endometriosis was proposed. | (42) |
| Yuan et al,
2018 | In vitro
experimental analysis | To investigate
changes that occur in the gut microbiota during the development of
murine endometriosis. | After
transplantation of endometrial tissue, gain of pathogenic microbes
and loss of probiotics were identified in mouse models of
endometriosis. This study showed that endometriosis induces gut
microbiota alterations in mice. | (43) |
| Ervin et al,
2019 | In vitro
experimental analysis | To provide the
in vitro analysis of the ability of some human gut microbial
gut microbial β-glucuronidase enzymes to reactivate estrogen
glucuronides to estrone and estradiol. | This research
showed that gut microbial β-glucuronidase enzymes participate in
estrogen metabolism within the mammalian gastrointestinal tract.
β-glucuronidase can reactivate estrogens from their inactive
glucuronides, suggesting that this enzyme is involved in the
estrobolome. | (18) |
| Ata et al,
2019 | A prospective
study | To compare vaginal,
cervical and gut microbiota between women with stage 3/4
endometriosis and healthy controls. | The study compared
vaginal, cervical and gut microbiota between women with stage 3/4
endometriosis and healthy controls. An increase in Gardnerella,
Streptococcus, Escherichia, Shigella, and Ureoplasma was identified
in the cervical microbiota of women with stage 3/4 endometriosis.
Dysbiosis in the genital tract microbiome was identified in the
affected women. | (48) |
| Hernandes et
al, 2020 | A case-controlled
biological study | To identify
microbiome profiles in vaginal fluid, endometrium, and lesion of
women with deep endometriosis. | An increase in
Lactobacillus, Gardnerella, Streptococcus, and Prevotella in
vaginal fluid, eutopic endometrium and endometriotic lesion. An
increase in Alishewanella, Enterococcus, and Pseudomonas in women
with deep endometriotic lesions. | (55) |
| Wei et al,
2020 | A case-controlled
biological study | To identify
microbiota composition and distribution along the female
reproductive tract of women with endometriosis. | The specific
microbiota composition in upper reproductive tract may be an
indicator for the risk of endometriosis. | (56) |
| Jiang et al,
2021 | A review
article | To investigate and
summarize intricate connections between the gut and reproductive
tract microbiota and endometriosis. | 16S rRNA amplicon
analysis revealed altered microbiota composition and distribution
along the female reproductive tract in women with endometriosis.
Dysbiosis disrupts normal immune function, impairs
immunosurveillance, alters immune cell profiles, and leads to the
elevation of proinflammatory cytokines, which may contribute to the
pathogenesis of endometriosis. | (8) |
| Kovács et
al, 2021 | A review
article | To discuss the
identification and diagnostic potential of novel disease biomarkers
of microbial origin. | Altered profile of
gut microbial composition and function may be a potent source of
biomarkers of endometriosis. Endometriosis-specific microbial
profiles may also represent targets for novel diagnostic options
for endometriosis. | (47) |
Experimental induction of endometriosis by fecal
microbiota transplantation has been previously reported; the
transplantation of fecal microbiota obtained from a surgically
induced endometriosis mouse donor into the gastrointestinal tract
of other endometriotic mice promoted lesion growth and associated
inflammation (50). Furthermore,
in vivo experiments demonstrated that broad-spectrum
antibiotic treatment in mice with endometriosis reduced the
endometriotic lesion size (43).
The mouse model demonstrated that endometriosis-specific gut
microbiota can alter disease progression by modulating inflammation
(50). Non-pathogenic and
pathogenic fecal bacterial contamination, followed by a process of
proinflammatory cytokine- and immune-activation-mediated gut
dysfunction, provides a plausible explanation of gut microbial
dysbiosis in endometriosis. Furthermore, preliminary studies on the
efficacy of probiotics for women with endometriosis have revealed
positive effects in pain relief (8,51,52)
(see Treatment of endometriosis by modulating the microbiome).
Conversely, studies with endometriosis models which
used mice and rhesus macaques have reported that endometriosis
induced gut microbiota alterations or dysbiosis (43,44).
Previous animal studies have reported that gut microbiota
composition alterations promote the development of endometriosis,
and that experimental endometriosis affects the composition of the
gut microbiota (43,44). However, changes in the gut
microbiota in the same female patient before and after the onset of
endometriosis are unknown. In contrast, other in vivo mouse
studies did not report changes in the gut microbiome composition
profiles (53). Therefore, a gut
microbiota composition imbalance could cause endometriosis
exacerbation; however, there are conflicting results that
endometriosis itself may or may not change the gut microbiota.
Role of the reproductive tract
microbiome
This subsection summarizes the role of the
reproductive tract microbiome in endometriosis (Fig. 2). Numerous studies have analyzed the
microbiome composition of the reproductive tract in females with
endometriosis and reported an increased abundance of opportunistic
pathogens associated with bacterial vaginosis and diminished
Lactobacillus dominance along the reproductive tract (from
the vagina up into the cervix, endometrium, fallopian tube and
peritoneal fluid) of females with endometriosis (8,47,48,54-57).
Opportunistic pathogens, including Streptococcaceae and
Staphylococaceae, were reported to be enriched in the cystic
fluid of females with ovarian endometrioma (54). Moreover, Alishewanella,
Enterococcus, Ureaplasma and Pseudomonas are
often detected in endometriotic lesions (9), whereas Atopobium is completely
absent in the vagina of patients with endometriosis (48). Additionally, greater numbers of
bacteria such as Gardnerella, Streptococcus,
Enterococci and Escherichia coli are present in the
endometrium of females with endometriosis compared with controls
(14). Furthermore, an increased
absolute number of bacterial pathogens, including
Gardnerella, Streptococcus, Escherichia,
Shigella and Ureaplasma, was reported in the cervical
microbiota of females with stage III-IV endometriosis (48). Endometriosis may affect the
composition of the reproductive tract microbiota, which in turn
increases the risk of lower genital tract infection, endometritis,
pelvic inflammatory disease and surgical site infections after
hysterectomy (14). Additionally,
increased numbers of Firmicutes and decreased
Actinobacteria and Bacteroidetes in the cervical
microbiota have been reported to increase the risk of developing
endometriosis (58). Increasing
disease severity (e.g., advanced stage, the coexistence of deep
infiltrating endometriosis, disease burden and symptom severity)
have been reported to be associated with decreasing abundance of
Dialister and an increasing abundance of
Lactobacillus and Streptococcus (58). Endometriosis is characterized by the
loss of beneficial microorganisms (e.g., Lactobacilli) and
the gain of pathogenic microorganisms. However, the specific
bacteria identified as the causative microbiota remains unknown.
Furthermore, evidence of the effects of the host site on the
reproductive tract microbiota composition is currently scarce.
Possible mechanisms of endometriosis
development by alterations of microbiota abundance and
composition
This section summarizes the mechanisms by which
alterations in gut and reproductive tract microbiota composition
and function cause endometriosis. The growth and maintenance of
endometriosis are influenced by estrogen metabolism (59). The gut microbiome is an important
regulator of circulating estrogens (17). The gut microbiota is involved in
regulation of estrogen levels via the β-glucuronidase enzyme, which
is responsible for the deconjugation of conjugated estrogens into
their active forms (18). Thus,
estrogen metabolism is modulated by the estrobolome (i.e., the
aggregate of genes capable of metabolizing estrogens in the gut
microbiome) (17). Gut microbiome
dysbiosis caused by the imbalance between the commensal and
pathogenic microbiomes or reduced microbial diversity dysregulates
the bidirectional crosstalk between the gut and uterus, which
results in not only immune dysfunction but also altered estrogen
signaling (8,60). Microbial diversity and dysbiosis
reduction lower β-glucuronidase activity (17). Conversely, the estrobolome, through
the increased abundance of β-glucuronidase-producing bacteria,
increases circulating estrogens, thereby developing endometriosis
(8,17). Investigating the number of
β-glucuronidase-producing bacteria in the gut microbiome of
patients with endometriosis is of great interest.
The gut microbiome communicates bidirectionally with
distant tissues via the gut-brain axis (61). The gut microbiota modulates
neurophysiological processes by altering immune, endocrine,
reproductive and neural signaling pathways via the gut-brain axis
(62). Estrogen has been reported
to stimulate neural growth and differentiation both in vitro
(63) and in animal studies
(64). The gut-brain axis promotes
the central sensitization of chronic pain (65). Therefore, endometriosis-associated
pain may be affected by regulation of microglia and astrocytes via
the dysbiotic gut-brain axis (65).
Severe pain and infertility associated with endometriosis have been
reported to be associated with decreased cervical microbial
diversity and abundance (66).
Finally, changes in the microbiome abundance and
composition can affect the metabolic profile, and vice versa
(17). A metabolomic study in
animals has reported that bile acid biosynthesis and α-linolenic
acid metabolism were characteristic pathways in the feces of
endometriosis mice (46). Gut
dysbiosis in mice with endometriosis appears to cause fecal
metabolomic alterations, but its significance in the progression of
endometriosis is unclear.
Collectively, these results suggest that the gut
microbiota initiates inflammation and affects the pathogenesis and
progression of endometriosis via dysregulation of the estrobolome,
metabolome and gut-brain axis (58,66).
However, whether the composition, abundance and functional
alteration of the gut microbiota causes disease progression is
unknown.
Treatment of endometriosis by
modulating the microbiome
This section first discusses the beneficial effect
of probiotics on endometriosis. A recent review summarized dietary
supplements (i.e., anti-inflammatory, anti-oxidant,
anti-proliferative and immune modulators) used for treating
endometriosis (67). Targeting the
gut microbiota with probiotics, prebiotics and symbiotics may be
beneficial dietary interventions for certain patients, including
those with endometriosis, osteoporosis and obesity (67-70).
Data on the oral administration of Lactobacillus and omega-3
polyunsaturated fatty acids (PUFAs) are well documented and were
presented as examples. See reference (67) for the remaining drugs. The previous
study reviewed the up-to-date evidence on dietary supplements used
as a complementary treatment, including vitamin C, vitamin D,
vitamin E, zinc, magnesium, selenium, omega 3, propolis, quercetin,
curcumin, N-acetylcysteine, probiotics, resveratrol, alpha lipoic
acid and epigallocatechin-3-gallate (67). Oral Lactobacillus administration
reduced the growth of mouse endometriotic lesions by increasing
IL-12 concentration and NK cell activity (51,71,72).
Randomized, placebo-controlled trials reported that oral
Lactobacillus administration ameliorated
endometriosis-associated pain in females (52,73).
Furthermore, Lactobacillus probiotic treatment has been
reported to prevent endometriosis proliferation in rats (71). Additionally, a high intake of
omega-3 PUFAs may reduce the risk of developing endometriosis by
modulating inflammatory and oxidative stress (68). A mouse model of endometriosis
demonstrated that a high-dose PUFA treatment inhibited lesion
growth (69). Therefore, available
data suggest the important role of dietary supplement-induced
beneficial changes in the gut microbiota in human health and reduce
the risk of inflammatory diseases, including endometriosis.
There are numerous available therapeutic approaches,
such as modulating the microbiome, to treat endometriosis.
Preclinical studies have evaluated the possibility of antibiotic
administration, fecal microbial transplantation or administration
of bacterial components for the treatment of endometriosis in
addition to probiotic treatment. Chadchan et al (50) reported that antibiotic therapy with
metronidazole reduced endometriotic lesion progression in mice
through gut microbiota modulation. Moreover, fecal microbiota
transplantation is a promising therapeutic option to target
Clostridium difficile infections and holds promise in
developing novel therapeutics for multiple sclerosis, autism,
obesity and other systemic diseases (74). Additionally, the gut microbiota
could induce inflammatory pain in endometriosis by increasing
glutamate and decreasing gamma-aminobutyric acid levels through
modulation of microglia, astrocytes and immune cells (65). Fecal microbiota transplantation has
been reported to be a potential treatment option for endometriosis
due to its beneficial effects in reducing pain despite the absence
of no clinical data (74).
Furthermore, a preclinical mouse model demonstrated that gut
microbiota-derived short-chain fatty acids (e.g., n-butyrate)
protected against endometriosis progression (75).
4. Discussion
Microbial profile abnormalities (e.g., the
composition and abundance) of the gut and reproductive tract
microbiome have been reported in numerous diseases, such as
inflammatory bowel disease, allergies, autoimmunity, psoriasis,
arthritis, reproductive disorders, cancer and endometriosis
(6,7,23-28,30-37,76,77).
Both animal and human studies have reported that pathogenic
bacteria are enriched in the endometriosis group compared with
controls (8,9,14,41-48,54-57,73).
Additionally, abnormal gut and cervical microbiomes may be
associated with endometriosis severity, including pain and
infertility, possibly through the impact of the estrobolome and
metabolome (58,76). Reproductive tract microbiota
alterations were also identified in mice, nonhuman primates and
females with endometriosis, which has been universally observed
across species (8,43,44,50-52).
Microbiota dysbiosis and endometriosis development and progression
are thought to be supported by a bidirectional relationship
(76); however, no evidence
suggests a cause-effect relationship. Additionally, altered
specific microbiota composition in endometriosis has no clear
consensus. The microbiota has been reported to be a major regulator
of such physiological processes, not only within the gut and
reproductive tract but also at distant sites, e.g.,
microbiota-brain crosstalk (78).
As mentioned above, microbial profiles in the gut and endometriosis
and in the genital tract and endometriosis have been reported, but
the crosstalk between the gut and genital tract environments
remains unclear. Collectively, microbiome composition and abundance
alterations may cause endometriosis and its associated symptoms,
including infertility and pelvic pain, possibly through the
gut-brain axis.
The microbiota is not merely composed of
microorganisms, such as commensals and symbionts, and they are
increasingly recognized as serving beneficial roles in human health
and reproduction beyond infection (34). The immune system is unable to
eliminate the microbial symbiont population once the commensal
microbiomes (e.g., non-pathogenic environmental bacteria, certain
commensals and probiotics) are established in the host (79). Females infected with commensals or
non-pathogenic microbiota early in life maintain a symbiotic
relationship, thereby avoiding excessive tissue damage, preserving
immune homeostasis and suppressing the inflammatory response
(7,23). However, modern females often face a
rapidly changing microbial environment, with serious biodiversity
loss due to industrialized Western lifestyles. Environmental
changes (e.g., diet and lifestyle), which lead to reduced
microorganism exposure can affect the microbiota composition and
diversity. A microbiota imbalance or impairment (e.g., a
combination of increased pathogenic microbes and loss of
probiotics) in the gut and female reproductive tract alters immune
cell profiles, disrupts normal immune function and compromises
immunosurveillance, which leads to chronic states of aberrant
immune activation and persistent inflammatory responses (8). This concept may be supported by
epidemiological data that the prevalence of autoimmunity, allergy,
inflammatory bowel disease and reproductive disease, such as
endometriosis, is increasing in high-income countries (7,25,76,80).
The so-called hygiene hypothesis initially focused on allergic
diseases, but a reduced non-pathogenic commensal microbial
diversity may cause numerous types of disease, including autoimmune
diseases, through impaired immunoregulatory mechanisms (23). Microbiotas, inherited at birth,
serve a role in an individual's predisposition to developing
certain diseases (2). Moreover,
endometriosis is generally more common in females with irritable
bowel syndrome because both diseases are characterized by chronic
inflammation (45,47). Interestingly, decreased NK activity
(81) and increased risk of
pregnancy complications (82,83)
remain unchanged after surgery for endometriosis.
Microbiota dysbiosis and frequent retrograde
menstruation can contribute to the development of endometriosis.
Alterations of eubiosis to dysbiosis (the first event) may induce
immune, metabolomic and estrobolomic disturbances, which can
trigger systemic inflammation and contribute to the development and
progression of endometriosis (7,8).
Furthermore, frequent retrograde menstruation (the second event)
causes repeated inflammation and serves a critical role in the
development of endometriosis. The current retrograde menstrual
theory does not provide an adequate explanation for why
endometriosis occurs only in certain females while almost all
females have retrograde menstruation. It can be hypothesized that
only females who experience the first and second events, not just
one of the two events, are more likely to develop endometriosis.
The increased risk of developing endometriosis in daughters of
mothers with endometriosis may be related to the effects of
microbiota exposure (84). This
evidence is at least partly explained by the hygiene hypothesis,
which suggests that infections by microorganisms, such as
commensals and symbionts, early in life prevent non-communicable
inflammatory diseases (7,13).
Finally, preclinical and clinical studies have
demonstrated no commonality in gut microbiota composition profiles
across species. Several studies have reported that the microbial
composition of the gut and reproductive tract is altered in females
with endometriosis; however, its clinical relevance needs to be
evaluated. Future studies will focus on the immune and molecular
mechanisms which underlie the host-microbiota relationship to
identify effective strategies to diagnose, manage and prevent
endometriosis. In conclusion, this review provides an updated
overview of the relationship between the gut and reproductive tract
microbiome and endometriosis, and discusses how dysbiosis increases
the risk of disease.
5. Summary and future perspective
Endometriosis has become a significant public health
problem in Japan because of its higher prevalence in Asian females
compared with that in Caucasian females (85). In recent years, the impact of
symbiotic and pathogenic microorganisms on human health has
attracted considerable attention. This review summarized the latest
findings on the role of microbiomes in the study of endometriosis.
The gut and reproductive tract microbiota components of females
with endometriosis differ from those in healthy females, which may
help identify potential biomarker candidates. Therefore, an
in-depth analysis of microbiota compositions should be performed to
assess the most appropriate prevention, diagnosis and treatment in
patients with endometriosis. Further research is required to
identify specific microbiota compositions that are altered in
females with endometriosis and to explore the causal
relationship.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
HK performed conception and design, acquisition of
data, analysis and interpretation of data and wrote the manuscript.
Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Author ORCID
ORCID: 0000-0002-8124-6269.
Competing interests
The author declares that they have no competing
interests.
References
|
1
|
Sampson JA: Perforating hemorrhagic
(chocolate) cysts of the ovary. Their importance and especially
their relation to pelvic adenomas of the endometrial type
(‘adenomyoma’ of the uterus, rectovaginal septum, sigmoid, etc.).
Arch Surg. 3:245–323. 1921.
|
|
2
|
Amro B, Ramirez Aristondo ME, Alsuwaidi S,
Almaamari B, Hakim Z, Tahlak M, Wattiez A and Koninckx PR: New
understanding of diagnosis, treatment and prevention of
endometriosis. Int J Environ Res Public Health.
19(6725)2022.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Shafrir AL, Farland LV, Shah DK, Harris
HR, Kvaskoff M, Zondervan K and Missmer SA: Risk for and
consequences of endometriosis: A critical epidemiologic review.
Best Pract Res Clin Obstet Gynaecol. 51:1–15. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Eskenazi B and Warner ML: Epidemiology of
endometriosis. Obstet Gynecol Clin North Am. 24:235–258.
1997.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Emera D, Romero R and Wagner G: The
evolution of menstruation: A new model for genetic assimilation:
Explaining molecular origins of maternal responses to fetal
invasiveness. Bioessays. 34:26–35. 2012.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Mignemi G, Facchini C, Raimondo D,
Montanari G, Ferrini G and Seracchioli R: A case report of nasal
endometriosis in a patient affected by Behcet's disease. J Minim
Invasive Gynecol. 19:514–516. 2012.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Rook GA, Lowry CA and Raison CL: Microbial
‘old friends’, immunoregulation and stress resilience. Evol Med
Public Health. 2013:46–64. 2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Jiang I, Yong PJ, Allaire C and Bedaiwy
MA: Intricate connections between the microbiota and endometriosis.
Int J Mol Sci. 22(5644)2021.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Symons LK, Miller JE, Kay VR, Marks RM,
Liblik K, Koti M and Tayade C: The immunopathophysiology of
endometriosis. Trends Mol Med. 24:748–762. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Shigesi N, Kvaskoff M, Kirtley S, Feng Q,
Fang H, Knight JC, Missmer SA, Rahmioglu N, Zondervan KT and Becker
CM: The association between endometriosis and autoimmune diseases:
A systematic review and meta-analysis. Hum Reprod Update.
25:486–503. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Kobayashi H, Higashiura Y, Shigetomi H and
Kajihara H: Pathogenesis of endometriosis: The role of initial
infection and subsequent sterile inflammation (review). Mol Med
Rep. 9:9–15. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Agostinis C, Mangogna A, Bossi F, Ricci G,
Kishore U and Bulla R: Uterine immunity and microbiota: A shifting
paradigm. Front Immunol. 10(2387)2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Ege MJ: The hygiene hypothesis in the age
of the microbiome. Ann Am Thorac Soc. 14 (Suppl 5):S348–S353.
2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Koninckx PR, Ussia A, Tahlak M, Adamyan L,
Wattiez A, Martin DC and Gomel V: Infection as a potential cofactor
in the genetic-epigenetic pathophysiology of endometriosis: A
systematic review. Facts Views Vis Obgyn. 11:209–216.
2019.PubMed/NCBI
|
|
15
|
Hooper LV, Littman DR and Macpherson AJ:
Interactions between the microbiota and the immune system. Science.
336:1268–1273. 2012.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Pickard JM, Zeng MY, Caruso R and Núñez G:
Gut microbiota: Role in pathogen colonization, immune responses,
and inflammatory disease. Immunol Rev. 279:70–89. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Baker JM, Al-Nakkash L and
Herbst-Kralovetz MM: Estrogen-gut microbiome axis: Physiological
and clinical implications. Maturitas. 103:45–53. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ervin SM, Li H, Lim L, Roberts LR, Liang
X, Mani S and Redinbo MR: Gut microbial β-glucuronidases reactivate
estrogens as components of the estrobolome that reactivate
estrogens. J Biol Chem. 294:18586–18599. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Laschke MW and Menger MD: The gut
microbiota: A puppet master in the pathogenesis of endometriosis?
Am J Obstet Gynecol. 215:68.e1–e4. 2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Collado MC, Cernada M, Baüerl C, Vento M
and Pérez-Martínez G: Microbial ecology and host-microbiota
interactions during early life stages. Gut Microbes. 3:352–365.
2012.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Schnupf P, Gaboriau-Routhiau V and
Cerf-Bensussan N: Modulation of the gut microbiota to improve
innate resistance. Curr Opin Immuno. 54:137–144. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Benner M, Ferwerda G, Joosten I and van
der Molen RG: How uterine microbiota might be responsible for a
receptive, fertile endometrium. Hum Reprod Update. 24:393–415.
2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Lambrecht BN and Hammad H: The immunology
of the allergy epidemic and the hygiene hypothesis. Nat Immunol.
18:1076–1083. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Gaffin JM, Kanchongkittiphon W and
Phipatanakul W: Perinatal and early childhood environmental factors
influencing allergic asthma immunopathogenesis. Int
Immunopharmacol. 22:21–30. 2014.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Crusell MKW, Hansen TH, Nielsen T, Allin
KH, Rühlemann MC, Damm P, Vestergaard H, Rørbye C, Jørgensen NR,
Christiansen OB, et al: Comparative studies of the gut microbiota
in the offspring of mothers with and without gestational diabetes.
Front Cell Infect Microbiol. 10(536282)2020.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Jain N: The early life education of the
immune system: Moms, microbes and (missed) opportunities. Gut
Microbes. 12(1824564)2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Qi X, Yun C, Pang Y and Qiao J: The impact
of the gut microbiota on the reproductive and metabolic endocrine
system. Gut Microbes. 13:1–21. 2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Colquhoun C, Duncan M and Grant G:
Inflammatory bowel diseases: Host-microbial-environmental
interactions in dysbiosis. Diseases. 8(13)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Weiss GA and Hennet T: Mechanisms and
consequences of intestinal dysbiosis. Cell Mol Life Sci.
74:2959–2977. 2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Ling Z, Liu X, Luo Y, Wu X, Yuan L, Tong
X, Li L and Xiang C: Associations between vaginal pathogenic
community and bacterial vaginosis in Chinese reproductive-age
women. PLoS One. 8(e76589)2013.PubMed/NCBI View Article : Google Scholar
|
|
31
|
He Y, Na R, Niu X, Xiao B and Yang H:
Lactobacillus rhamnosus and Lactobacillus casei
affect various stages of Gardnerella species biofilm
formation. Front Cell Infect Microbiol. 11(568178)2021.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Spurbeck RR and Arvidson CG: Inhibition of
Neisseria gonorrhoeae epithelial cell interactions by
vaginal Lactobacillus species. Infect Immun.
80(3742)2012.
|
|
33
|
Chen C, Song X, Wei W, Zhong H, Dai J, Lan
Z, Li F, Yu X, Feng Q, Wang Z, et al: The microbiota continuum
along the female reproductive tract and its relation to
uterine-related diseases. Nat Commun. 8(875)2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Peric A, Weiss J, Vulliemoz N, Baud D and
Stojanov M: Bacterial colonization of the female upper genital
tract. Int J Mol Sci. 20(3405)2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Moreno I, Codoñer FM, Vilella F, Valbuena
D, Martinez-Blanch JF, Jimenez-Almazán J, Alonso R, Alamá P, Remohí
J, Pellicer A, et al: Evidence that the endometrial microbiota has
an effect on implantation success or failure. Am J Obstet Gynecol.
215:684–703. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Kallus Y, Miller JH and Libby E: Paradoxes
in leaky microbial trade. Nat Commun. 8(1361)2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Muzny CA, Łaniewski P, Schwebke JR and
Herbst-Kralovetz MM: Host-vaginal microbiota interactions in the
pathogenesis of bacterial vaginosis. Curr Opin Infect Dis.
33:59–65. 2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Onderdonk AB, Delaney ML and Fichorova RN:
The human microbiome during bacterial vaginosis. Clin Microbiol
Rev. 29:223–238. 2016.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Tai FW, Chang CYY, Chiang JH, Lin WC and
Wan L: Association of pelvic inflammatory disease with risk of
endometriosis: A nationwide cohort study involving 141,460
individuals. J Clin Med. 7(379)2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Moghaddam MZ, Ansariniya H, Seifati SM,
Zare F and Fesahat F: Immunopathogenesis of endometriosis: An
overview of the role of innate and adaptive immune cells and their
mediators. Am J Reprod Immunol. 87(e13537)2022.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Khan KN, Kitajima M, Hiraki K, Yamaguchi
N, Katamine S, Matsuyama T, Nakashima M, Fujishita A, Ishimaru T
and Masuzaki H: Escherichia coli contamination of menstrual
blood and effect of bacterial endotoxin on endometriosis. Fertil
Steril. 94:2860–2863.e1-e3. 2010.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Khan KN, Fujishita A, Hiraki K, Kitajima
M, Nakashima M, Fushiki S and Kitawaki J: Bacterial contamination
hypothesis: A new concept in endometriosis. Reprod Med Biol.
17:125–133. 2018.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Yuan M, Li D, Zhang Z, Sun H, An M and
Wang G: Endometriosis induces gut microbiota alterations in mice.
Hum Reprod. 33:607–616. 2018.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Bailey MT and Coe CL: Endometriosis is
associated with an altered profile of intestinal microflora in
female rhesus monkeys. Hum Reprod. 17:1704–1708. 2002.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Seaman HE, Ballard KD, Wright JT and de
Vries CS: Endometriosis and its coexistence with irritable bowel
syndrome and pelvic inflammatory disease: Findings from a national
case-control study-part 2. BJOG. 115:1392–1396. 2008.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Ni Z, Sun S, Bi Y, Ding J, Cheng W, Yu J,
Zhou L, Li M and Yu C: Correlation of fecal metabolomics and gut
microbiota in mice with endometriosis. Am J Reprod Immunol.
84(e13307)2020.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Kovács Z, Glover L, Reidy F, MacSharry J
and Saldova R: Novel diagnostic options for endometriosis-based on
the glycome and microbiome. J Adv Res. 33:167–181. 2021.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Ata B, Yildiz S, Turkgeldi E, Brocal VP,
Dinleyici EC, Moya A and Urman B: The endobiota study: Comparison
of vaginal, cervical and gut microbiota between women with stage
3/4 endometriosis and healthy controls. Sci Rep.
9(2204)2019.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Shan J, Ni Z, Cheng W, Zhou L, Zhai D, Sun
S and Yu C: Gut microbiota imbalance and its correlations with
hormone and inflammatory factors in patients with stage 3/4
endometriosis. Arch Gynecol Obstet. 304:1363–1373. 2021.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Chadchan SB, Cheng M, Parnell LA, Yin Y,
Schriefer A, Mysorekar IU and Kommagani R: Antibiotic therapy with
metronidazole reduces endometriosis disease progression in mice: A
potential role for gut microbiota. Hum Reprod. 34:1106–1116.
2019.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Molina NM, Sola-Leyva A, Saez-Lara MJ,
Plaza-Diaz J, Tubić-Pavlović A, Romero B, Clavero A, Mozas-Moreno
J, Fontes J and Altmäe S: New opportunities for endometrial health
by modifying uterine microbial composition: Present or future?
Biomolecules. 10(593)2020.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Itoh H, Uchida M, Sashihara T, Ji ZS, Li
J, Tang Q, Ni S, Song L and Kaminogawa S: Lactobacillus
gasseri OLL2809 is effective especially on the menstrual pain and
dysmenorrhea in endometriosis patients: Randomized, double-blind,
placebo-controlled study. Cytotechnology. 63:153–161.
2011.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Hantschel J, Weis S, Schäfer KH, Menger
MD, Kohl M, Egert M and Laschke MW: Effect of endometriosis on the
fecal bacteriota composition of mice during the acute phase of
lesion formation. PLoS One. 14(e0226835)2019.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Khan KN, Fujishita A, Masumoto H, Muto H,
Kitajima M, Masuzaki H and Kitawaki J: Molecular detection of
intrauterine microbial colonization in women with endometriosis.
Eur J Obstet Gynecol Reprod Biol. 199:69–75. 2016.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Hernandes C, Silveira P, Rodrigues Sereia
AF, Christoff AP, Mendes H, Valter de Oliveira LF and Podgaec S:
Microbiome profile of deep endometriosis patients: Comparison of
vaginal fluid, endometrium and lesion. Diagnostics (Basel).
10(163)2020.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Wei W, Zhang X, Tang H, Zeng L and Wu R:
Microbiota composition and distribution along the female
reproductive tract of women with endometriosis. Ann Clin Microbiol
Antimicrob. 19(15)2020.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Dols JA, Molenaar D, van der Helm JJ,
Caspers MP, de Kat Angelino-Bart A, Schuren FH, Speksnijder AG,
Westerhoff HV, Richardus JH, Boon ME, et al: Molecular assessment
of bacterial vaginosis by Lactobacillus abundance and
species diversity. BMC Infect Dis. 16(180)2016.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Chang CY, Chiang AJ, Lai MT, Yan MJ, Tseng
CC, Lo LC, Wan L, Li CJ, Tsui KH, Chen CM, et al: A more diverse
cervical microbiome associates with better clinical outcomes in
patients with endometriosis: A pilot study. Biomedicines.
10(174)2022.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Vallvé-Juanico J, Houshdaran S and Giudice
LC: The endometrial immune environment of women with endometriosis.
Hum Reprod Update. 25:564–591. 2019.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Shi N, Li N, Duan X and Niu H: Interaction
between the gut microbiome and mucosal immune system. Mil Med Res.
4(14)2017.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Chadchan SB, Singh V and Kommagani R:
Female reproductive dysfunctions and the gut microbiota. J Mol
Endocrinol. 69:R81–R94. 2022.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Collins SM, Surette M and Bercik P: The
interplay between the intestinal microbiota and the brain. Nat Rev
Microbiol. 10:735–742. 2012.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Ferreira A and Caceres A:
Estrogen-enhanced neurite growth: Evidence for a selective
induction of Tau and stable microtubules. J Neurosci. 11:392–400.
1991.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Toran-Allerand CD, Miranda RC, Bentham WD,
Sohrabji F, Brown TJ, Hochberg RB and MacLusky NJ: Estrogen
receptors colocalize with low-affinity nerve growth factor
receptors in cholinergic neurons of the basal forebrain. Proc Natl
Acad Sci USA. 89:4668–4672. 1992.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Ustianowska K, Ustianowski Ł, Machaj F,
Gorący A, Rosik J, Szostak B, Szostak J and Pawlik A: The role of
the human microbiome in the pathogenesis of pain. Int J Mol Sci.
23(13267)2022.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Talwar C, Singh V and Kommagani R: The gut
microbiota: A double-edged sword in endometriosis†. Biol Reprod.
107:881–901. 2022.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Yalçın Bahat P, Ayhan I, Üreyen Özdemir E,
İnceboz Ü and Oral E: Dietary supplements for treatment of
endometriosis: A review. Acta Biomed. 93(e2022159)2022.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Hopeman MM, Riley JK, Frolova AI, Jiang H
and Jungheim ES: Serum polyunsaturated fatty acids and
endometriosis. Reprod Sci. 22:1083–1087. 2015.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Attaman JA, Stanic AK, Kim M, Lynch MP,
Rueda BR and Styer AK: The anti-inflammatory impact of omega-3
polyunsaturated Fatty acids during the establishment of
endometriosis-like lesions. Am J Reprod Immunol. 72:392–402.
2014.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Kelly OJ, Gilman JC, Kim Y and Ilich JZ:
Long-chain polyunsaturated fatty acids may mutually benefit both
obesity and osteoporosis. Nutr Res. 33:521–533. 2013.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Uchida M and Kobayashi O: Effects of
Lactobacillus gasseri OLL2809 on the induced endometriosis
in rats. Biosci Biotechnol Biochem. 77:1879–1881. 2013.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Itoh H, Sashihara T, Hosono A, Kaminogawa
S and Uchida M: Lactobacillus gasseri OLL2809 inhibits
development of ectopic endometrial cell in peritoneal cavity via
activation of NK cells in a murine endometriosis model.
Cytotechnology. 63:205–210. 2011.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Khodaverdi S, Mohammadbeigi R, Khaledi M,
Mesdaghinia L, Sharifzadeh F, Nasiripour S and Gorginzadeh M:
Beneficial effects of oral Lactobacillus on pain severity in
women suffering from endometriosis: A pilot placebo-controlled
randomized clinical trial. Int J Fertil Steril. 13:178–183.
2019.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Quaranta G, Sanguinetti M and Masucci L:
Fecal microbiota transplantation: A potential tool for treatment of
human female reproductive tract diseases. Front Immunol.
10(2653)2019.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Chadchan SB, Popli P, Ambati CR, Tycksen
E, Han SJ, Bulun SE, Putluri N, Biest SW and Kommagani R: Gut
microbiota-derived short-chain fatty acids protect against the
progression of endometriosis. Life Sci Alliance.
4(e202101224)2021.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Salliss ME, Farland LV, Mahnert ND and
Herbst-Kralovetz MM: The role of gut and genital microbiota and the
estrobolome in endometriosis, infertility and chronic pelvic pain.
Hum Reprod Update. 28:92–131. 2021.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Cho I and Blaser MJ: The human microbiome:
At the interface of health and disease. Nat Rev Genet. 13:260–270.
2012.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Zhong SR, Kuang Q, Zhang F, Chen B and
Zhong ZG: Functional roles of the microbiota-gut-brain axis in
Alzheimer's disease: Implications of gut microbiota-targeted
therapy. Transl Neurosci. 12:581–600. 2021.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Zhou JZ, Way SS and Chen K: Immunology of
uterine and vaginal mucosae. Trends Immunol. 39:302–314.
2018.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Petraglia F, Serour GI and Chapron C: The
changing prevalence of infertility. Int J Gynaecol Obstet. 123
(Suppl 2):S4–S8. 2013.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Vercellini P, Sergenti G, Buggio L,
Frattaruolo MP, Dridi D and Berlanda N: Advances in the medical
management of bowel endometriosis. Best Pract Res Clin Obstet
Gynaecol. 71:78–99. 2020.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Nirgianakis K, Ma L, McKinnon B and
Mueller MD: Recurrence patterns after surgery in patients with
different endometriosis subtypes: A long-term hospital-based cohort
study. J Clin Med. 9(496)2020.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Nirgianakis K, Gasparri ML, Radan AP,
Villiger A, McKinnon B, Mosimann B, Papadia A and Mueller MD:
Obstetric complications after laparoscopic excision of posterior
deep infiltrating endometriosis: A case-control study. Fertil
Steril. 110:459–466. 2018.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Dalsgaard T, Hjordt Hansen MV, Hartwell D
and Lidegaard O: Reproductive prognosis in daughters of women with
and without endometriosis. Hum Reprod. 28:2284–2288.
2013.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Yamamoto A, Johnstone EB, Bloom MS,
Huddleston HG and Fujimoto VY: A higher prevalence of endometriosis
among Asian women does not contribute to poorer IVF outcomes. J
Assist Reprod Genet. 34:765–774. 2017.PubMed/NCBI View Article : Google Scholar
|