Introduction
Lung cancer is a leading cause of cancer-related
mortality worldwide (1). Although
previous studies have demonstrated the molecular pathogenesis of
lung cancer, little progress has been made in terms of its
prevention, diagnosis and treatment (2). Thus, further investigation of its
molecular pathogenesis is required to reduce the mortality rate
caused by lung cancer.
MicroRNAs (miRs) are a group of non-coding small
RNAs mostly regulating their target mRNAs by post-transcriptional
repression (3). They bind to the
target mRNA transcript at the 3′-UTR, generating destabilization
that inhibits protein translation (4). Intensive studies have been carried
out investigating the involvement of miRs in human malignant tumors
due to their ability to regulate several genes involved in multiple
cancer pathways (5). miRs are
highly involved in various cell processes, including proliferation,
differentiation, apoptosis and development, by simultaneously
controlling the expression levels of hundreds of genes (6,7). In
cancer cells, both oncogenic and tumor suppressive miRs have been
reported to contribute to cancer development and progression
(7,8). Notably, certain miRs, such as
miR-200s, have a biphasic role as oncogenes or tumor suppressor
genes (TSGs) (9).
miR-9, including the family members miR-9-1, 9-2 and
9-3, is encoded by three chromosomal loci: miR-9-1 on chromosome
(Chr) 1q22, miR-9-2 on Chr 5q14.3 and miR-9-3 on Chr 15q26.1. miR-9
has been reported to have both oncogenic and tumor suppressive
roles. In its oncogenic function, miR-9 induces
epithelial-mesenchymal transition via the direct suppression of
E-cadherin mRNA (8), which
downregulates β-catenin to promote vascular endothelial growth
factor expression, leading to angiogenesis and tumor metastasis.
Moreover, miR-9 is able to repress the expression of TSGs such as
FOX1 (10) and CDX2 (11). By contrast, in its tumor
suppressive role, the loss of miR-9 expression through methylation
is associated with metastasis or poor prognosis in several types of
cancer (12–14).
In this study, we determined the miR-9s methylation
status in surgically resected non-small cell lung cancers (NSCLC)
cases to investigate its role and impact on NSCLC.
Materials and methods
Tissue samples
Between January, 2005 and August, 2009, 556 NSCLC
cases underwent complete pulmonary resection at the Department of
Cancer and Thoracic Surgery, in the Okayama University Hospital,
Japan. Of these, 293 specimens were made available for this study,
while 20 corresponding non-malignant lung tissues from these
patients were also used for this study. Institutional review board
permission and informed consent were obtained for the cases.
Detection of miR-9s methylation by
combined bisulfite restriction analysis (COBRA)
COBRA is a semiquantitative method used to measure
methylation at specific methylation-sensitive restriction sites
(15). Briefly, genomic DNA was
isolated from tissue samples by proteinase K digestion followed by
phenol-chloroform (1:1) extraction and ethanol precipitation
(16). Genomic DNA (200 ng) was
subjected to bisulfite treatment using the EZ DNA Methylation Gold™
kit (Zymo Research, Irvine, CA, USA). Primers designed for the
predicted bisulfite-modified sequences were based on the nucleotide
sequence submitted to GenBank (accession numbers: NR_029691 for
miR-9-1, NR_030741 for miR-9-2, and NR_029692 for miR-9-3) and were
as follows: miR-9-1, forward: GGGGTTTGGTTGTTTATTT and reverse:
TCCACTACCCTTCTCTAAAAA: miR-9-2, forward: GGA
ATAAATTTTGAAGGTAATAGATTT and reverse: CAC AAACCTCTATCTTCCTCTTACC;
miR-9-3, forward: GTTTGTTTATTTTTTTTGGTTTTT and reverse: AACCT
CCCTTAACCAATACC. PCR was carried out in 25 μl reaction
mixtures under the following conditions: an initial denaturation
step at 95°C for 12 min, followed by 45 cycles of 94°C for 30 sec,
annealing at 54°C (miR-9-1), 58°C (miR-9-2) or 55°C (miR-9-3) for
60 sec and extension at 72°C for 60 sec., with a final extension
step at 72°C for 7 min with the GeneAmp PCR System 9700 (Applied
Biosystems, Carlsbad, CA, USA). PCR products (5 μl) were
digested for 9 h with BstUI (New England Biolabs, Beverly,
MA, USA) that recognizes sequences unique to methylated and
bisulfite-converted alleles ( CG|CG site). The restricted products
were electrophoresed on 2.2% agarose gels [3:1 mixture of agarose
(Sigma-Aldrich Corp., St. Louis, MO, USA) and NUSieve®
GTG® agarose (Cambrex Bio Science Rockland, Inc.,
Rockland, ME, USA)] and stained with ethidium bromide.
In situ hybridization (ISH) of miR-9
Sections (10 μm) for ISH analysis and 5
μm sections stained with hematoxylin and eosin (H&E)
were generated from paraffin-embedded tissue blocks. The
pathological classification was determined from an H&E-stained
5 μm slide section based on the 7th World Health
Organization criteria of lung cancer (17). Sections (10 μm) were
deparaffinized in xylene, then dehydrated in an ethanol series
(100, 90, 70 and 50%). After blocking endogenous peroxidase with 3%
hydrogen peroxide, sections were digested with 1 μg/ml
proteinase K (Roche Diagnostics, Indianapolis, IN, USA) at 37°C for
15 min. The LNA-modified probe, complementary to miR-9 (100 nM;
Exiqon, Woburn, MA, USA), diluted with NTE buffer (ISHR 7; Nippon
Gene Co., Ltd., Tokyo, Japan) was hybridized with the sections for
16 h at 37°C. Post-hybridization washes were performed three times
for 20 min in 2X standard saline citrate (SSC)/25% formamide and
three times for 10 min in 0.1X SSC at 37°C. The probe signal was
amplified using Tyramide Signal Amplification Plus DNP kits
(PerkinElmer, Inc., Waltham, MA, USA) and color reactions were
developed in 5-bromo-4-chloro-3-indolyl phosphate/nitro blue
tetrazolium solution. Nuclear counterstaining was performed with
0.2% methylgreen.
Statistical analysis
Differences in statistical significance among the
categorized groups were compared using the Chi-square test. The
logistic regression analysis using the variables with the
univariate analysis was used to further explore statistically
significant differences observed and to identify baseline factors
that may independently correlate with miR-9s methylation. A
univariate analysis of overall survival (OS) was performed using
the Kaplan-Meier method with log-rank testing, while a multivariate
analysis was performed using the Cox proportional hazard model. The
data were analyzed using the JMP software version 9.0.0 (SAS
Institute, Inc., Cary, NC, USA). P<0.05 was considered to
indicate a statistically significant difference for each
analysis.
Results
Patient characteristics
Patient characteristics are provided in Table I. The median age of the patients
was 66 years (range, 29–87). Younger patients were defined as ≤66
(n=147) years old, while elderly patients were >66 years old
(n=146). The cases were divided into two groups of never (n=114)
and ever (n=179) smokers. Never smokers were defined as individuals
with a lifetime exposure to ≤100 cigarettes, while ever smokers had
a lifetime exposure to >100 cigarettes. The early pathological
stage of NSCLC was defined as stage I NSCLC (n=205), while the
advanced as stages II and III NSCLC (n=88).
 | Table IClinicopathological parameters and
miR-9s methylation (n=293). |
Table I
Clinicopathological parameters and
miR-9s methylation (n=293).
| Any miR-9s
methylation (n=76)
|
|---|
| Subsets | n | % | P-value |
|---|
| Agea (n) | | | |
| ≤66 (147) | 29 | 20 | 0.03 |
| >66 (146) | 47 | 32 | |
| Gender (n) | | | |
| Male (172) | 54 | 31 | 0.01 |
| Female (121) | 22 | 18 | |
| Smoking (n) | | | |
| Never (114) | 27 | 24 | NS |
| Ever (179) | 49 | 27 | |
| Histology (n) | | | |
| AD (221) | 54 | 24 | NS (AD vs. SQ and
others) |
| SQ (62) | 16 | 26 | |
| Others (10) | 6 | 60 | |
| Stage (n) | | | |
| I (205) | 60 | 29 | (I vs. 210
II–III) |
| II (35) | 6 | 17 | 0.047 |
| III (53) | 10 | 19 | |
| p-N (n) | | | |
| Negative (220) | 67 | 30 | 0.002 |
| Positive (73) | 9 | 12 | |
miR-9s methylation by COBRA and miR-9
expression
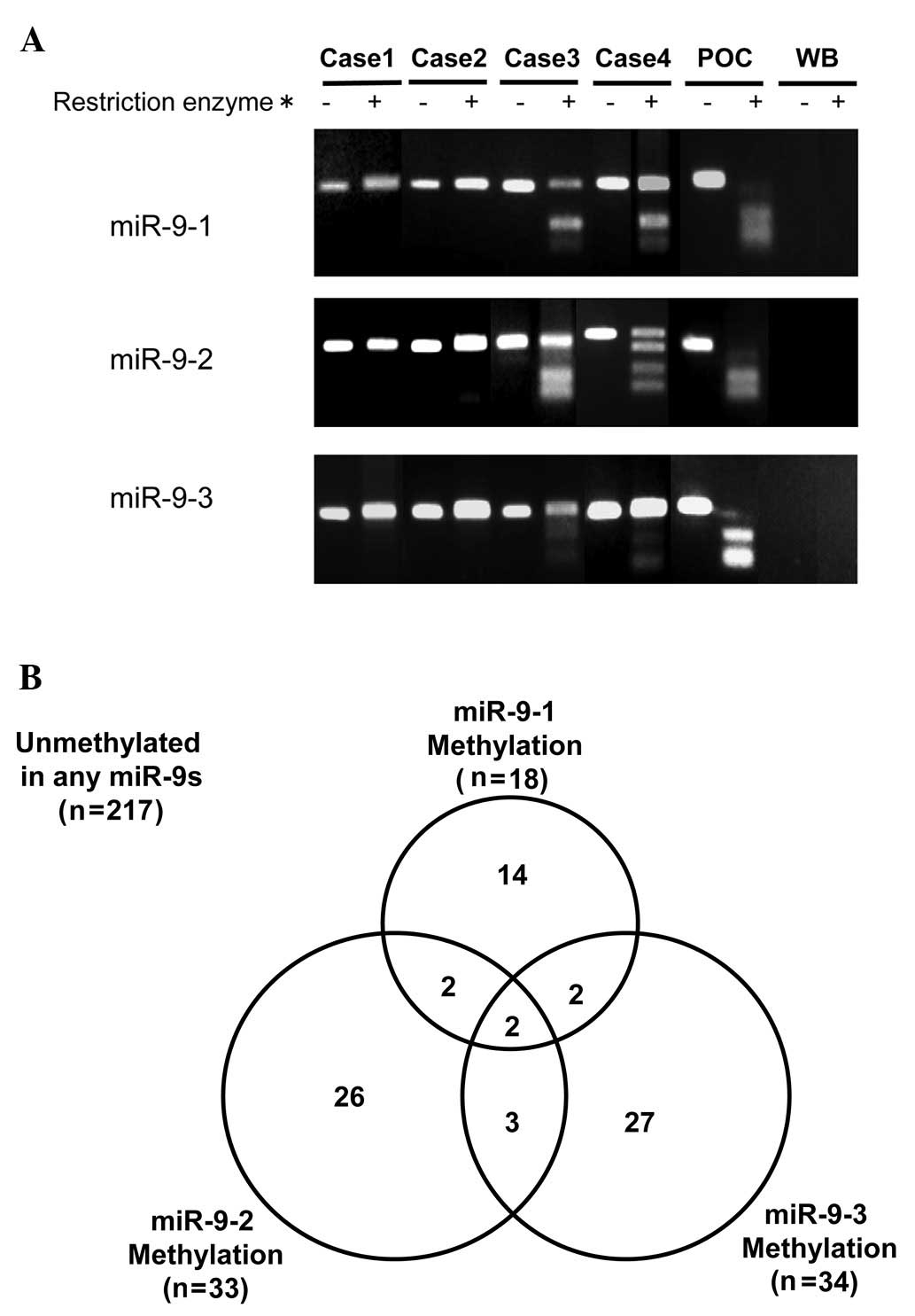
A representative example of COBRA is shown in
Fig. 1A. A malignant pleural
mesothelioma cell line, NCI-H290, was used as the positive control
for the methylation of miR-9-1, 9-2 and 9-3. The methylation of
miR-9-1, 9-2 and 9-3 was present in 20 (7%), 33 (11%) and 34 (12%)
of 293 NSCLC cases, respectively. The methylation of any miR-9s
(miR-9s methylation) was observed in 76 of 293 NSCLC cases (26%)
(Fig. 1B). Of these 76 cases, two
harbored methylation of the three (1%), seven harbored methylation
of two (2%) and 67 (23%) harbored methylation of one miR-9s. miR-9
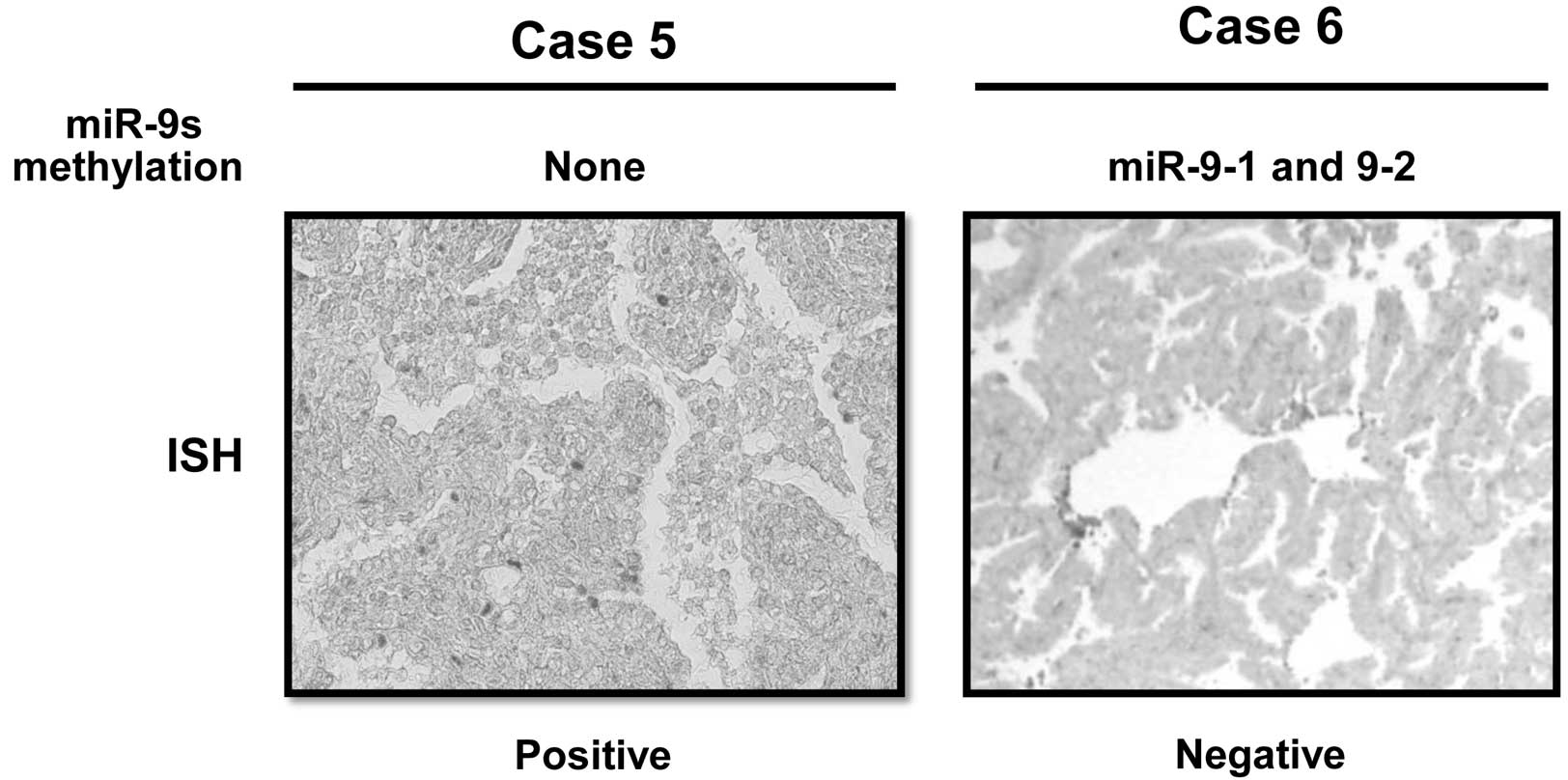
expression was determined by ISH assay in eight NSCLC cases.
Representative examples are shown in Fig. 3. The five miR-9-methylated cases,
including the two harboring methylation in miR-9-1 and 9-2, but not
in miR-9-3, and the three harboring methylation in either miR-9-1,
9-2 or 9-3, showed a negative miR-9 expression, whereas the other
three miR-9s unmethylated cases showed a positive miR-9 expression.
None of the 20 corresponding non-malignant lung tissues harbored
any miR-9s methylation.
miR-9s methylation and
clinicopathological factors
The associations between miR-9s methylation and
clinicopathological factors are shown in Table I. miR-9s methylation was
significantly more frequent in elderly [32 vs. 20% (younger
patients): P=0.03], male patients [31 vs. 18% (female): P=0.01], in
early pathological stage [29 vs. 18% (advanced stage): P=0.047] and
pathologically lymph node-negative patients [30 vs. 12% (lymph
nodal metastasis positive): P=0.002]. Logistic regression analysis
of the significant univariate analysis factors showed that male
gender [odds ratio (OR), 2.0; 95% confidence interval (95% CI),
1.1–3.6; P=0.01] and pathologically negative lymph node metastasis
(OR, 4.8; 95% CI, 1.4–17.2; P=0.002) were independent relative
factors for miR-9s methylation.
miR-9s methylation and clinical
outcome
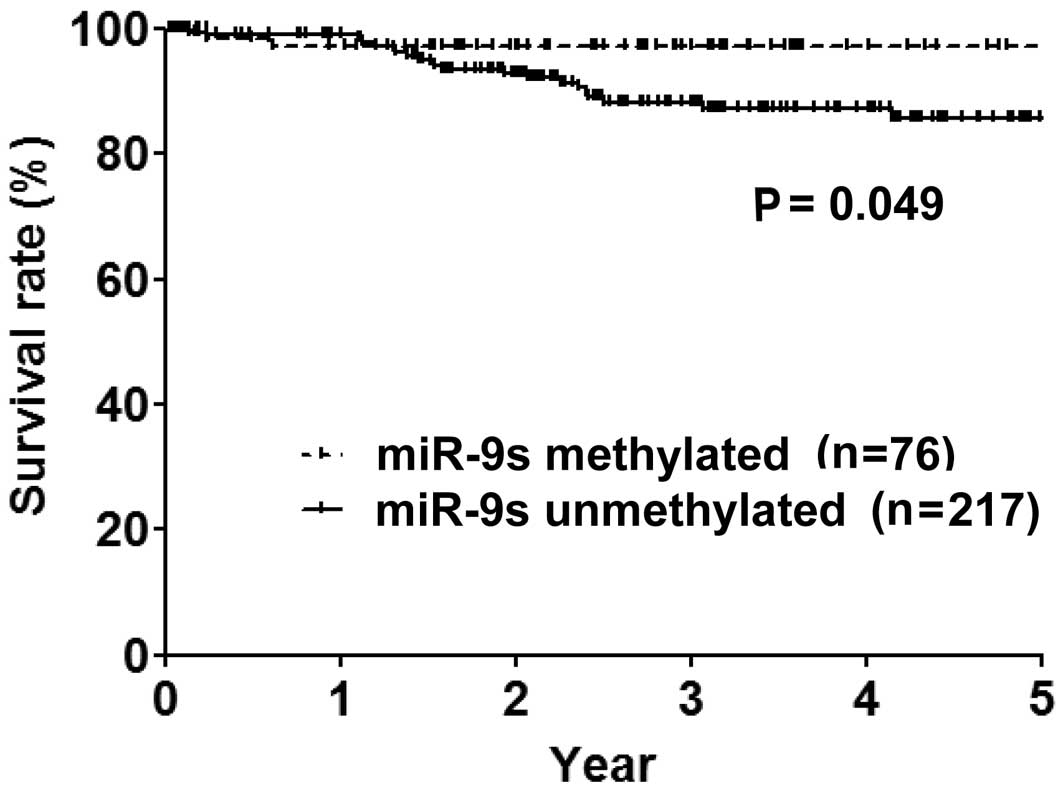
Over a median follow-up period of 32 months, the OS
rates of the cases (n=293), miR-9-methylated (n=76) and
unmethylated cases (n=217) were 91.8, 97.3 and 90%, respectively.
miR-9-methylated cases had a significantly more prolonged OS period
compared to miR-9-unmethylated cases (P=0.049) (Fig. 2). Female gender, never smoker,
adenocarcinoma histology, early pathological stage and
pathologically lymph node-negativity were additional significant
predictive factors for a prolonged OS period in the univariate
analysis (Table II). The
multivariate analysis of the significant univariate analysis
demonstrated that miR-9s methylation [hazard ratio (HR) 4.2, 95%
CI: 1.2–27.0, P=0.026] and early pathological stage (HR 8.3, 95%
CI: 2.1–28.6, P=0.004) were independent predictive factors for a
prolonged OS period.
| Table IIImpact of miR-9s methylation on
overall survival. |
Table II
Impact of miR-9s methylation on
overall survival.
| Factors | Median OS
(month) | P-value |
|---|
| miR-9s
methylation | | 0.049 |
| Positive | 33.3 | |
| Negative | 31.1 | |
| Age (years) | | 0.3 |
| ≤66 | 36.4 | |
| >66 | 28.3 | |
| Gender | | 0.013 |
| Male | 29.1 | |
| Female | 34.9 | |
| Smoking history | | 0.006 |
| Never | 35.5 | |
| Ever | 28.3 | |
| Histology | | 0.017 |
| AD | 33.1 | |
| Non-AD | 25.2 | |
| Stage | | <0.0001 |
| I | 33.0 | |
| II and III | 27.0 | |
| p-N | | 0.002 |
| Negative | 32.6 | |
| Positive | 29.9 | |
Discussion
miRs are capable of acting as oncogenes or TSGs and
the observed widespread alteration in miR expression patterns is
highly correlated with various human malignancies (18). Regarding NSCLC, miR alterations
also have clinical implications: reduced let-7 expression (19) and miR-155 over-expression (20) reportedly correlate with improved or
worse clinical outcomes, respectively. In the present study, miR-9
families were demonstrated to frequently methylate, especially in
lymph node-negative cases, and to silence miR-9 expression in cases
with miR-9s methylation. These findings suggest that miR-9 is
oncogenically involved in NSCLC.
Methylation in CpG islands of promoter regions is an
important epigenetic mechanism of gene silencing that is often
observed in cancer, leading to the inhibition of gene transcription
(21). Similar to protein coding
genes, miRs are downregulated through epigenetic mechanisms
(11,22,23).
Although methylation typically affects TSGs, it also occurs in
oncogenes, such as COX219, telomerase reverse transcriptase and
epidermal growth factor receptor, silencing their expression
(24–27). miR-9 is reportedly involved in an
oncogenic as well as a tumor-suppressing manner in human
malignancies.
In the former case, miR-9 expression levels were
elevated in primary tumors in breast cancer patients with
metastasis compared to metastasis-free patients (28), while other reports showed the
oncogenic involvement of miR-9 based on basic experiments (8) and clinical findings (29). In their study, Tsai et al
demonstrated that the tumor-suppressive miR-9 was silenced by
promoter methylation in gastric cancer (30), while Heller et al found that
patients with miR-9-3 methylation showed shorter OS duration among
101 NSCLC patients (31). In this
study, the methylation status of the miR-9 family members was
determined using COBRA in a larger patient set compared to previous
reports, and NSCLC patients with methylation of any miR-9 family
member were found to be an independent factor associated with a
prolonged OS period. Moreover, miR-9-3 methylation showed no
prognostic impact (data not shown).
In conclusion, miR-9s methylation, which silences
miR-9 expression, is associated with early-stage and pathologically
lymph node-negative cases of NSCLC, suggesting miR-9 to be
oncogenically involved in NSCLC in promoting metastasis and tumor
aggressiveness.
Acknowledgements
The authors would like to thank Ms.
Chikako Isobe for preparing the sections from paraffin-embedded
tissue blocks.
References
|
1.
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics. CA Cancer J Clin. 60:277–300. 2010.
|
|
2.
|
Toyooka S, Mitsudomi T, Soh J, et al:
Molecular oncology of lung cancer. Gen Thorac Cardiovasc Surg.
59:527–537. 2011.
|
|
3.
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
|
|
4.
|
Ambros V: MicroRNA pathways in flies and
worms: growth, death, fat, stress, and timing. Cell. 113:673–676.
2003.
|
|
5.
|
Lu J, Getz G, Miska EA, et al: MicroRNA
expression profiles classify human cancers. Nature. 435:834–838.
2005.
|
|
6.
|
Otaegi G, Pollock A, Hong J and Sun T:
MicroRNA miR-9 modifies motor neuron columns by a tuning regulation
of FoxP1 levels in developing spinal cords. J Neurosci. 31:809–818.
2011.
|
|
7.
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in Cancer. Annu Rev Med. 60:167–179. 2009.
|
|
8.
|
Ma L, Young J, Prabhala H, et al: miR-9, a
MYC/MYCN-activated microRNA, regulates E-cadherin and cancer
metastasis. Nat Cell Biol. 12:247–256. 2010.
|
|
9.
|
Korpal M, Ell BJ, Buffa FM, et al: Direct
targeting of Sec23a by miR-200s influences cancer cell secretome
and promotes metastatic colonization. Nat Med. 17:1101–1108.
2011.
|
|
10.
|
Myatt SS, Wang J, Monteiro LJ, et al:
Definition of microRNAs that repress expression of the tumor
suppressor gene FOXO1 in endometrial cancer. Cancer Res.
70:367–377. 2010.
|
|
11.
|
Saito Y, Liang G, Egger G, et al: Specific
activation of microRNA-127 with downregulation of the
proto-oncogene BCL6 by chromatin-modifying drugs in human cancer
cells. Cancer Cell. 9:435–443. 2006.
|
|
12.
|
Lehmann U, Hasemeier B, Christgen M, et
al: Epigenetic inactivation of microRNA gene hsa-mir-9-1 in human
breast cancer. J Pathol. 214:17–24. 2008.
|
|
13.
|
Hildebrandt MA, Gu J, Lin J, et al: Hsa-
miR-9s methylation status is associated with cancer development and
metastatic recurrence in patients with clear cell renal cell
carcinoma. Oncogene. 29:5724–5728. 2010.
|
|
14.
|
Lujambio A, Calin GA, Villanueva A, et al:
A microRNA DNA methylation signature for human cancer metastasis.
Proc Natl Acad Sci USA. 105:13556–13561. 2008.
|
|
15.
|
Xiong Z and Laird PW: COBRA: a sensitive
and quantitative DNA methylation assay. Nucleic Acids Res.
25:2532–2534. 1997.
|
|
16.
|
Herrmann BG and Frischauf AM: Isolation of
genomic DNA. Methods Enzymol. 152:180–183. 1987.
|
|
17.
|
Venneti S, Boateng LA, Friedman JR, et al:
MiR-9 and MiRNA-200a distinguish hemangioblastomas from metastatic
clear cell renal cell carcinomas in the CNS. Brain Pathol.
22:522–529. 2012.
|
|
18.
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006.
|
|
19.
|
Takamizawa J, Konishi H, Yanagisawa K, et
al: Reduced expression of the let-7 microRNAs in human lung cancers
in association with shortened postoperative survival. Cancer Res.
64:3753–3756. 2004.
|
|
20.
|
Yanaihara N, Caplen N, Bowman E, et al:
Unique microRNA molecular profiles in lung cancer diagnosis and
prognosis. Cancer Cell. 9:189–198. 2006.
|
|
21.
|
Toyooka S, Toyooka KO, Miyajima K, et al:
Epigenetic down-regulation of death-associated protein kinase in
lung cancers. Clin Cancer Res. 9:3034–3041. 2003.
|
|
22.
|
Lujambio A, Ropero S, Ballestar E, et al:
Genetic unmasking of an epigenetically silenced microRNA in human
cancer cells. Cancer Res. 67:1424–1429. 2007.
|
|
23.
|
Toyota M, Suzuki H, Sasaki Y, et al:
Epigenetic silencing of microRNA-34b/c and B-cell translocation
gene 4 is associated with CpG island methylation in colorectal
cancer. Cancer Res. 68:4123–4132. 2008.
|
|
24.
|
Devereux TR, Horikawa I, Anna CH, Annab
LA, Afshari CA and Barrett JC: DNA methylation analysis of the
promoter region of the human telomerase reverse transcriptase
(hTERT) gene. Cancer Res. 59:6087–6090. 1999.
|
|
25.
|
Santini V, Kantarjian HM and Issa JP:
Changes in DNA methylation in neoplasia: pathophysiology and
therapeutic implications. Ann Intern Med. 134:573–586. 2001.
|
|
26.
|
Jones PA and Baylin SB: The fundamental
role of epigenetic events in cancer. Nat Rev Genet. 3:415–428.
2002.
|
|
27.
|
Scartozzi M, Bearzi I, Mandolesi A, et al:
Epidermal growth factor receptor (EGFR) gene promoter methylation
and cetuximab treatment in colorectal cancer patients. Br J Cancer.
104:1786–1790. 2011.
|
|
28.
|
Khew-Goodall Y and Goodall GJ:
Myc-modulated miR-9 makes more metastases. Nat Cell Biol.
12:209–211. 2010.
|
|
29.
|
Tsai KW, Liao YL, Wu CW, et al: Aberrant
hypermethylation of miR-9 genes in gastric cancer. Epigenetics.
6:1189–1197. 2011.
|
|
30.
|
Gravgaard KH, Lyng MB, Laenkholm AV, et
al: The miRNA-200 family and miRNA-9 exhibit differential
expression in primary versus corresponding metastatic tissue in
breast cancer. Breast Cancer Res Treat. 2012.
|
|
31.
|
Heller G, Weinzierl M, Noll C, et al:
Genome-wide miRNA expression profiling identifies miR-9-3 and
miR-193a as targets for DNA methylation in non-small cell lung
cancers. Clin Cancer Res. 18:1619–1629. 2012.
|