Introduction
Glioblastoma (GBM) is a highly aggressive, invasive
and poorly responsive brain tumor. At present, the median survival
time for children that have received chemotherapy and radiotherapy
is reported between 11–24 months; the five-year survival rate is
below 20% (1). Adult (aGBM) and
primary pediatric GBM (pGBM) show distinct molecular pathways of
tumorigenesis and genetic profiling (1–6); pGBM
appears to be more similar to secondary aGBM that has evolved from
diffuse grade II or grade III gliomas. Indeed, similarly to
secondary aGBM, pGBM exhibits transcriptional regulator ATRX,
histone H3.3 and cellular tumor antigen p53 mutations and only
rarely shows epidermal growth factor receptor
amplification/overexpression (7,8) or
phosphatase and tensin homolog mutations (9).
Chemoresistance is the main obstacle to successful
chemotherapy for brain tumors. Drug resistance is the main cause of
tumor recurrence and patient relapse; this phenomenon is associated
with several biological mechanisms, including apoptosis, DNA damage
and repair, epigenetic regulation, alteration in ATP-binding
cassette transporter family and dysregulation of microRNAs
(miRNAs/miRs). A limited number of studies have investigated the
underlying mechanisms of pGBM chemoresistance, although in recent
decades, increasing amounts of data report the involvement of
miRNAs in drug sensitivity and chemoresistance (10–17).
miRNAs represent a novel class of gene regulators
that are involved in several physiological processes, including
cell differentiation, proliferation, stress response and anti-viral
defense, as well as pathological conditions such as cancer
(18). They are a large family of
evolutionary conserved, short (19–24 nucleotides), single-stranded,
non-coding RNAs that also exhibit strong tissue and cellular
specificity to developmental stages. They are able to regulate gene
expression at the post-transcriptional level, leading to mRNA
degradation, with consequential downregulation of encoded protein
by translational repression. miRNAs regulate 3% of the human genome
and up to 30% of protein-coding genes (19,20).
Many miRNAs could regulate multiple mRNAs and a single miRNA could
regulate multiple mRNA targets (21).
The etiological role of miRNAs in tumor development
is supported by the observation that half of miRNA genes are
localized in cancer-associated genes, fragile genome sites
(22) or regions that are often
amplified (23), acting as tumor
suppressors or as oncogenes, depending on which genes/pathways they
control (24). Aberrant microRNA
expression profiles have been identified in aGBM (25–32), but
few studies have investigated the role of miRNAs in pGBM (33,34).
In order to better understand chemoresistance
mechanism and regulation in high-grade glioma (HGG), the present
study generated a microRNA profile of pGBMs, through a
TaqMan® Human MicroRNA Array v2.0 approach. A set of
differentially expressed miRNAs in pGBMs and in GBM cell lines
(A172, U87MG and resistant-T98G) was identified, in comparison to
non-tumor pediatric cerebral cortex samples.
The present preliminary study may contribute the
biological understanding of pGBM chemoresistance, which represents
the most common causes of relapse (35), and may provide biomarkers for
therapeutic strategies.
Materials and methods
Patients and samples
Patients with pGBM seen between April 2008 and May
2013 at the Meyer Children's University Hospital (Florence, Italy)
were eligible for the present study. Histological diagnosis and
tumor grading was performed based on the 2007 World Health
Organization criteria (36). In
total, five non-tumor pediatric cerebral cortex samples
(non-tumoral pool) and seven pGBMs were obtained at the
Neuro-Surgery Unit of the Meyer Children's University Hospital. The
present study was approved by the institutional Ethical Committee.
Informed consent was obtained from the parents or legal guardians
in all cases. Diagnosis was confirmed by the review of the CNS
national panel of pathologists (Umberto I, Policlinico General
Hospital Sapienza University, Rome, Italy). The median age at the
time of diagnosis was 8±4.6 years (age range, 1–15 years; 4 females
and 3 males). All patients had been treated with chemotherapy
and/or radiotherapy according to consolidated pediatric treatments
(37–39) and underwent surgery for resection of
disease. The median follow-up was 10±6.1 months (range, 3–24
months).
Cell lines
Three human GBM cell lines, A172
(CRL-1620™; ATTC, Manassas, VA, USA), U87MG (HTB-14™;
ATTC) and resistant-T98G (CRL-1690™; ATCC) were
obtained. U87MG and resistant-T98G were cultured in Eagle's minimum
essential medium (Thermo Fisher Scientific, Inc., Waltham, MA,
USA), while A172 cells were grown in Dulbecco's modified Eagle's
medium (Thermo Fisher Scientific, Inc.). Each medium was
supplemented with 10% fetal bovine serum (cat. no. ECS0180L;
EuroClone SpA, Via Figino, Milan, Italy) and 1%
penicillin-streptomycin (Penicillin/Streptomicin 100X; cat. no.
ECB3001D; EuroClone SpA). All cell lines were maintained in a
humidified atmosphere of 5% CO2/95% air at 37°C. Cells
from exponentially growing cultures were used for all
experiments.
Expression study
miRNA and mRNA were extracted using the mirVana™
miRNA Isolation kit (cat. no. AM1560; Thermo Fisher Scientific,
Inc.) from tumor (pGBM 1–7) and non-tumor pediatric cerebral cortex
samples (pool of 5 samples), as well as pellets of the three cell
lines (A172, U87MG and resistant-T98G). Cells were trypsinized
(Trypsin-EDTA 1X, cat. no. ECB3052D, Euroclone SpA) from the
culture surface (6-well Primo multiwell plate; Euroclone SpA) and
transferred to 15 ml conical tubes (TC Tube, 15 ml; SARSTEDT AG
& Co. KG, Nümbrecht, Germany). The tubes containing cells and
media were centrifuged at 800 × g for 5 min at 4°C to pellet cells
and decant culture media. Subsequently, cells were washed in PBS
(cat. no. ECB4004L; EuroClone SpA) and further centrifuged at 800 ×
g for 5 min at 4°C for pelleting. Finally, PBS was decanted and
cell pellets were stored at −80°C.
MicroRNA expression profiles of three pGBMs and
non-tumoral pool of 5 samples were generated using
TaqMan® Human MicroRNA A Cards v2.0 (cat. no. 4398977;
Thermo Fisher Scientific, Inc.) according to manufacturers'
protocol, using the 7900HT Fast Real-Time PCR system (Thermo Fisher
Scientific, Inc.).
Bioinformatic analysis
Raw data were analyzed with the R computational
environment by using the HTqPCR package version 1.0 (40). The package HTqPCR is designed for the
analysis of cycle threshold (Ct) values from quantitative PCR
(qPCR) data. The heatmap was generated using the R version 3.5.1
(41) and the HTqPCR package. Raw
data were first normalized by using the quantile normalization
approach and then analyzed for differential expression with the
two-tailed t-test. miRNAs with statistically significant
differential expression were analyzed with the miRanda algorithm,
(www.microrna.org; version 3.3a) to search for
miRNA gene targets (42).
Finally, gene targets were analyzed for enrichment
in Gene Ontology (www.geneontology.org) (43,44) and
Kyoto Encyclopedia of Genes and Genomes database (www.genome.jp/kegg/kegg1.html) (45–47) with
a Fisher's exact test.
Validation of miRNAs by quantitative
polymerase chain reaction (qPCR)
miRNA was extracted using the mirVana™ miRNA
Isolation kit (cat. no. AM1560; Thermo Fisher Scientific, Inc.)
from tumors (pGBM 1–7) and non-tumor pediatric cerebral cortex
samples (pool of 5 samples), as well as pellets from three cell
lines (A172, U87MG and resistant-T98G). The expression of
previously identified dysregulated miRNAs were determined using
commercial assays (miR-137, cat. no. 001129; miR-216a, cat. no.
477976; miR-490, cat. no. 001037; miR-501-3p, cat. no. 002435;
miR-521, cat. no. Hs99999903_m1; miR-525-3p, cat. no. 478995_mir;
miR-873, cat. no. 478204_mir; miR-876-3p, cat. no. 002225; miR-448,
cat. no. 001029; all, Thermo Fisher Scientific, Inc.; included
forward and reverse primers).
cDNA was synthesized using the TaqMan®
MicroRNA Reverse Transcription kit (cat. no. 4366596; Thermo Fisher
Scientific, Inc.) according to the manufacturers' protocol (30 min
at 16°C, 30 min at 42°C and 5 min at 85°C) with the
GeneAmp® PCR System 9700-Applied Biosystems (Thermo
Fisher Scientific, Inc.).
qPCR was performed using the aforementioned
commercial, ready to use assays according to the protocol
instructions (10 min at 95°C, 40 cycles at 15 sec at 95°C, 60 sec
at 60°C) of the 7900HT Fast Real-Time PCR system (Thermo Fisher
Scientific, Inc.). All assays were performed in triplicate. For
each miRNA, the expression was normalized to that of RNU48 (cat.
no. 001006; Thermo Fisher Scientific, Inc.) and calculated using
the 2−∆∆Cq method (48).
GBMs were subsequently normalized and compared with the non-tumoral
pool.
Validation of target genes by
SYBR-Green PCR
mRNAs were extracted using the mirVana™ miRNA
Isolation kit (cat. no. AM1560; Thermo Fisher Scientific, Inc.)
from tumor (pGBM 1–7) and non-tumor pediatric cerebral cortex (pool
of 5 samples) samples, as well as pellets (obtained as
aforementioned) of the three cell lines (A172, U87MG and
resistant-T98G). cDNA was synthesized using the High Capacity
RNA-to-Cdna kit (cat. no. 4387406; Thermo Fisher Scientific, Inc.)
according to the manufacturers' protocol (60 min at 37°C and 5 min
at 95°C) with the GeneAmp® PCR System 9700-Applied
Biosystems (Thermo Fisher Scientific, Inc.). Primers were designed
using Primer3web version 4.1.0; (http://primer3.ut.ee/). The primer sequences utilized
are presented in Table I. Validation
of the expression of hypothetical target genes was performed using
the LightCycler® 480 SYBR-Green I Master mix (Roche
Diagnostics, Basel, Switzerland) on a LightCycler® 480
II (Roche Diagnostics) according to the manufacturers' protocol (10
min at 95°C, 40 cycles at 15 sec at 95°C, 45 sec at 60°C, 60 sec at
72°C; melting curve at 10 min at 95°C, 60 sec at 65°C) and
quantification was obtained using 2−ΔΔCq method
(48).
 | Table I.Primer sequences for validation of
target genes by SYBR-Green. |
Table I.
Primer sequences for validation of
target genes by SYBR-Green.
| Primer | Forward sequence
(5′-3′) | Reverse sequence
(5′-3′) |
|---|
| GRIA1-EXON13 |
AGTCAGCAGAGGCATCAGTT |
TGGGTGTTGCAATGCCATAG |
| GRIA1-EXON10 |
CGTTACGAGGGCTACTGTGT |
TCCATAGACCAGCTCTCCC |
| SORL1-EXON46 |
TCACAGCTTACCTTGGGAATACT |
GACCCCAGCTCATCGTACAG |
| SORL1-EXON35 |
TGGTTGGAGAGAGCATATGGA |
GGTCCTCAGGGTCACAAAGT |
| NUCKS1-EXON5 |
AAAATGTGCGCCAACAACG |
AATGGTGCCTCATCCTCCTC |
| NUCKS1-EXON7 |
GTCCAGTGAAAGGCAAAGG |
TCAGACCCTTCATCCCCAG |
| SOX11-EXON1 |
AATTTCTCTCAAAGCGCGCA |
GTGCAGTAGTCGGGGAACT |
| SOX11-EXON1.2 |
ACATCAAGCGGCCGATGAA |
GGATGAACGGGATCTTCTC |
| SAP30L-EXON1 |
GCTTCAGCACGGAGGAGGA |
CTTCTGGACCCTCTTGCTGA |
| SAP30L-EXON4 |
CGACACTTCAGGAACATACCTG |
CCCTCCGATTTCTGGTCCAG |
| HTT-EXON63 |
TGTGGGGTGATGCTGTCTG |
GTTCACTCTGTCCACACTCA |
| HTT-EXON48 |
GTTCAACCTAAGCCTGCTAGC |
GGGCTGGAAGACATGATGGA |
| PXMP4-EXON3 |
TGCAGGCCACATATATCCACT |
CGTGTGCTGGGTAGGTCTT |
| PXMP4-EXON4 |
CTGGCTGTAGAGAAGGGCTA |
TGTCGTGCCATACATTGCTG |
| THRB-EXON8 |
GAGAAAAGACGGCGGGAAGA |
CATGGGCTTCGGTGACAGTT |
| THRB-EXON10 |
GCGCTATGACCCAGAAAGTG |
GGAGGGCTACTTCAGTGTCA |
| PSD3-EXON5 |
TCTGAAATGGGGAGCACTGA |
TTCTTGCCAAGGTGTTTTGC |
| PSD3-EXON11 |
ACTGAGGAGAAAGCTAACGGA |
TCTTTCCATCCATATCTGCATGA |
| SPN-EXON2 |
CCCTACCTCCCTCAACTTCC |
CTGGTTGCATGAGGGGTTTC |
| SPN-EXON2.1F |
GTGACAGTGACCGTGGGAG |
GACCCAGACTTCAGCTCCTC |
| AGPAT4-EXON2 |
ACCTGGTCTTCTGCTACGTC |
AGGACAGTCTGCAGTTGATCT |
| AGPAT4-EXON4 |
AAGGTCCTGGCCAAGAAAGA |
AAAATACTTCTCGGGGTAGTCC |
| USP31-EXON1 |
CTTCATGAACGCCACGCTG |
AGCTGCTCAGTGACCTCG |
| USP31-EXON13 |
AGACAGGCGCATGAAACTTC |
ATGTAGTCCTCAGGGTCCCT |
| GRIK3-EXON3 |
CAATGCCGTCCAGTCCATCT |
CTGACCGCCACTTGAGGTA |
| GRIK3-EXON14 |
TTCGAGAAGATGTGGGCCTT |
ATCTGGGTGAGGTTGCAGTT |
| TNRC6B-EXON11 |
CCAAATCAAGATGGGTGCCTT |
CTAGCAGCGAAGTTTTGGGG |
| TNRC6B-EXON20 |
TGGTCCCCAGATCCCATAGG |
GATCGGGGTGCTGTGCTG |
| SNX29-EXON5 |
CCGTGTTCTGGTACTACGTG |
GGAGTGTTCGTTGAGGGCA |
| SNX29-EXON8 |
CCAATGGAAGTGAGAGCAGC |
CCCTGTGCTTCCTTCCTGAT |
| HIPK2-EXON2 |
CGTGCTTGGTCTTCGAGATG |
GCGTGGATAAGACCTAGGCT |
| HIPK2-EXON13 |
CCCTACTCCGACTCCTCCA |
ACCAATACTTCGCTGGCCT |
| RIMKLA-EXON1 |
CAGCTCTGGTTCCTGACGG |
GCGATCTGGTCCATAAGCAC |
| RIMKLA-EXON5 |
TGACAGAACAAGGCAAGCAG |
GCAATGATCCCACCCACATC |
Statistical analysis
Statistical analysis of nuclear casein kinase and
cyclin dependent kinase substrate 1 (NUCKS1) expression was
performed using one-way analysis of variance followed by a
Newman-Keuls post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Bioinformatics analysis
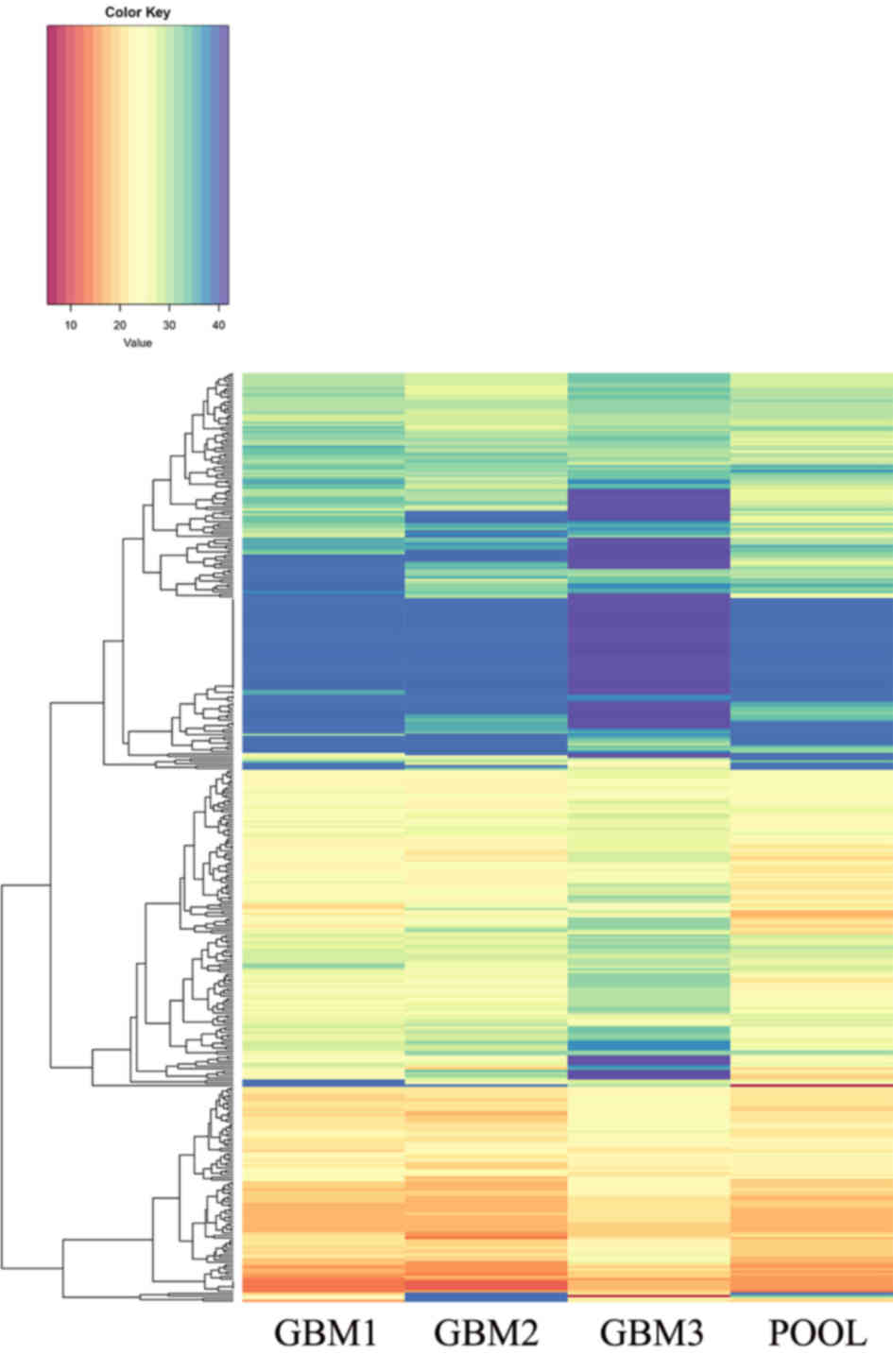
Bioinformatics analysis of TaqMan® Human
MicroRNA array version 2.0 revealed a set of miRNAs (miR-137,
miR-216a, miR-490, miR-501-3p, miR-521, miR-525-3p, miR-672,
miR-873, miR-876-3p and miR-448) that exhibited a statistically
significant differential expression in three pGBMs (GBM 1–3) when
compared with the non-tumoral pool (Fig.
1; significance cut-off level, two-tailed t-test with
P<0.01).
Validation of miRNAs and target
genes
In all tumors (pGBMs 1–7) and GBM cell lines (U87MG,
A172 and resistant-T98G), the downregulation of miR-137, miR-490,
miR-876-3p, miR-876-5p and miR-448 was confirmed, and the
upregulation of miR-501-3p was demonstrated.
Concerning the expression of the other dysregulated
miRNAs (miR-216a, −521, −525-3p, −672 and −873), the interpretation
of these results were unsuccessful due to poor reaction efficiency
of the commercial assays.
The validation of dysregulated miRNA expression was
obtained via commercial assays. Moreover, the expression of all
following predicted target genes was validated: Glutamate
ionotropic receptor AMPA type subunit 1 (GRIA1), sortilin
related receptor 1 (SORL1), NUCKS1, SRY-box 11
(SOX11), SAP30 like (SAP30L), huntingtin
(HTT), peroxisomal membrane protein 4 (PXMP4),
thyroid hormone receptor beta (THRB), pleckstrin and Sec7
domain containing 3 (PSD3), sialophorin (SPN),
1-acylglycerol-3-phosphate O-acyltransferase 4 (AGPAT4),
ubiquitin specific peptidase 31 (USP31), glutamate
ionotropic receptor kainate type subunit 3 (GRIK3),
trinucleotide repeat containing 6B (TNRC6B), sorting nexin
29 (SNX29), homeodomain interacting protein kinase 2
(HIPK2) and ribosomal modification protein rimK like family
member A (RIMKLA).
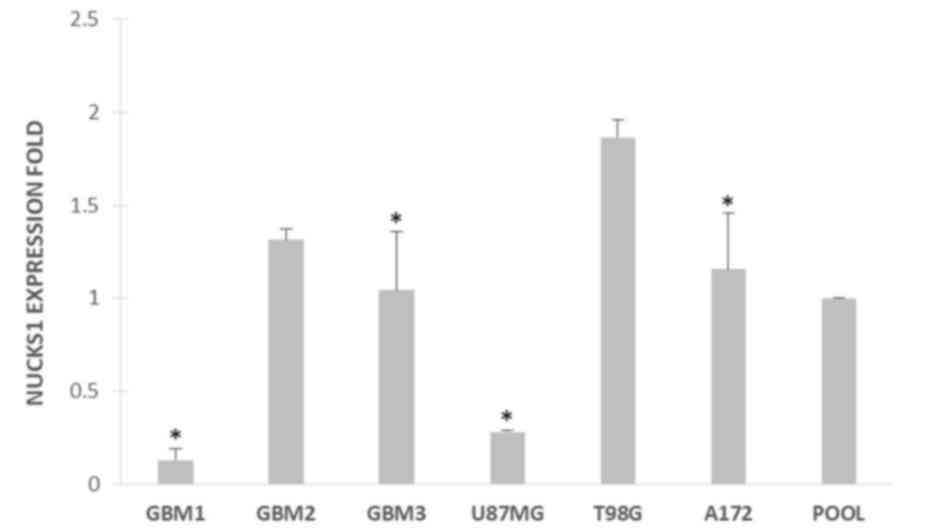
Finally, NUCKS1 was expressed in the tumor
tissues and cell lines of the current study. Furthermore, using
one-way analysis of variance followed by a Newman-Keuls post hoc
test, NUCKS1 expression was compared in T98G vs. GBM1, GBM2, GBM3,
U87MG and A172 cell lines. Statistically significant differences
were observed P<0.05; Fig. 2) in
T98G vs. GBM1, vs. GBM3, vs. U87MG and vs. A172 (all P<0.05;
Fig. 2). In particular, NUCKS1 was
overexpressed in T98G cells.
Discussion
In the present study, a microRNA expression profile
of pGBM was generated using TaqMan® Human MicroRNA Array
v2.0. in GBM tumors (7 pGBMs) and cell lines (U87MG, A172,
resistant-T98G). The results demonstrated that miR-137, miR-490,
miR-876-3p, miR-876-5p and miR-448 were downregulated, and
miR-501-3p was upregulated. Concerning the expression of the other
dysregulated miRNAs (miR-216a, −521, −525-3p, −672 and −873), the
interpretation of these results was unsuccessful due to the poor
reaction efficiency of the commercial assays used.
Furthermore, it was determined that the
aforementioned miRNAs were involved in the regulation of the
following target genes: GRIA1, SORL1, NUCKS1, SOX11, SAP30L,
HTT, PXMP4, THRB, PSD3, SPN, AGPAT4, USP31, GRIK3, POM121L8P,
TNRC6B, SNX29, HIPK2 and RIMKLA. All hypothetical target
genes were identified in tumors and cell lines and the
overexpression of NUCKS1 was detected in drug resistant T98G
cells.
NUCKS1 is a highly phosphorylated nuclear
DNA-binding protein that is involved in cell cycle progression and
proliferation (49). It serves as a
substrate for casein kinase 2 and cyclin-dependent kinase (CDK) −1,
−2, −4 and −6 (49–53). NUCKS1 also serves a role in the
response to DNA damage, homologous recombination and DNA repair
mechanisms that are critical for tumor suppression (54). The increased expression of NUCKS1 has
been reported in several different types of cancer, including
breast, colorectal, cervical and hepatocellular carcinoma (50,55–57).
However, its exact role in cancer development remains unclear. The
NUCKS1 gene is located on chromosome 1q32.1 (chr 1,
205,712,819-205,750,276), which undergoes recurrent
duplication/amplification in several different types of tumor
(58,59), including that of the brain (60–63). It
is well established that genes amplified in specific copy number
variants are associated with tumor progression and poor prognoses
(58–63).
Recently, Shen et al (64) demonstrated that NUCKS1 was a
target of miR-137 in human lung cancer tissue and resistant lung
cell lines. They also revealed that the tumor suppressive role of
miR-137 is mediated via the negative regulation of NUCKS1
protein expression. miR-137 is a tumor suppressor and a number of
its target genes, including cell division control protein 42, CDK6,
cyclooxygenase-2, paxillin, AKT2 and induced myeloid leukemia cell
differentiation protein are involved in cancer pathogenesis
(64). The loss of miR-137
expression has been determined in several different types of tumor
(65–69), including GBMs (28,30,32,69–72). In
addition, the restoration of miR-137 expression has been
demonstrated to be associated with the inhibition of tumorigenesis
(64). In glioma cell lines that
overexpress miR-137, cell cycle arrest in the G1 phase is promoted
via CDK6 suppression and retinoblastoma-associated protein-1
phosphorylation (26).
miR-137 expression increases during the glioma
stem-like cell differentiation in neurosphere cultures (70). The low expression of miR-137 observed
in GBM may reflect the loss of tumor cell differentiation, which
may contribute to an increased cell proliferation, whilst
maintaining an undifferentiated state (70).
At present, few data assess the differential
expression of the remaining miRNAs that were determined in the
present study. miR-490 is involved in the development and invasion
of different types of tumor (73–76) and
in the drug resistance of ovarian cancer (77). miR-448 functions as a tumor
suppressor gene in osteosarcoma, where it is downregulated in
tissues and in vitro models (78). miR-448 is also downregulated in
hepatocarcinoma and is associated with tumorigenesis (79). This association has also been
reported in ovarian cancer tissues and cell lines (80), breast cancer (81) and in T-cell acute lymphoblastic
leukemia (82). Conversely, miR-488
is overexpressed in lung cancer (83).
miR-501-3p has been determined to be a potential
biomarker associated with the progression of Alzheimer's disease
(84). In cancer however, it may
serve as a potential biomarker for pancreatic ductal adenocarcinoma
(85) and lymph node metastasis in
gastric cancer (86). Additionally,
miR-876-3p/5p has been associated with papillary thyroid carcinoma
(87), Hodgkin's lymphoma (88) and lung cancer (89). The miR-876 gene is located on the 9p
chromosome, which is deleted in various types of cancer (90,91).
Furthermore, pGBMs in particular exhibit a recurrent homozygous or
heterozygous 9p21.3 deletion, including the MIR876 gene
(60). Thus, the association between
each of the dysregulated miRNAs, NUCKS1 overexpression and
chemoresistance mechanisms in pGBMs requires further investigation.
Furthermore, it may be important to evaluate the role of NUCKS1
protein expression in pGMB tumor progression in a larger and
independent group of pediatric high grade glioma samples.
NUCKS1 overexpression in the resistant-T98G cell line, in
comparison with other non-resistant cell lines, U87MG and A172,
indicate its potential involvement in drug sensitivity and pGBM
response. The T98G cell line represents a useful in vitro
model, which may be utilized to determine the mechanism of acquired
chemoresistance in patients with pGBM. In a previous study, it was
demonstrated that T98G cells exhibit a different biological
response to antineoplastic treatments (doxorubicin) compared with
other GBM cell lines (92).
Chemoresistance represents an important challenge in
pGBM treatment and overcoming this phenomenon may improve patient
prognosis and increase survival rate. miRNAs are promising clinical
biomarkers, which may produce a greater understanding of the
biological processes associated with the development and
progression of pGBMs.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Meyer Children's
Hospital Anna Meyer Foundation, Florence, Italy.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LaG designed the current study and analyzed the
data. LaG, BM, SL, VDG and MDR performed machine learning analysis.
AM interpreted the data. LaG revised the work critically for
important intellectual content. LaG and VDG made substantial
contributions to drafting the manuscript. AMB histologically
diagnosed the tumors. IS, LoG and SG supervised the study and were
involved in patient recruitment, selection and treatment.
Ethics approval and consent to
participate
The present study was approved by the institutional
Ethical Committee of Meyer Children's Hospital, Florence, Italy.
Informed consent was obtained from the parents or legal guardians
in all cases.
Patient consent for publication
Written informed consent was obtained all the
patients.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
GBM
|
glioblastoma multiforme
|
|
pGBM
|
pediatric glioblastoma multiforme
|
|
aGBM
|
adult glioblastoma multiforme
|
|
miRNAs
|
microRNAs
|
References
|
1
|
Pollack IF, Hamilton RL, James CD,
Finkelstein SD, Burnham J, Yates AJ, Holmes EJ, Zhou T and Finlay
JL: Children's Oncology Group: Rarity of PTEN deletions and EGFR
amplification in malignant gliomas of childhood: Results from the
Children's Cancer Group 945 cohort. J Neurosurg. 105 Suppl
5:S418–S424. 2006.
|
|
2
|
Pollack IF, Finkelstein SD, Woods J,
Burnham J, Holmes EJ, Hamilton RL, Yates AJ, Boyett JM, Finlay JL
and Sposto R: Children's Cancer Group: Expression of p53 and
prognosis in children with malignant gliomas. N Engl J Med.
346:420–427. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pollack IF, Boyett JM, Yates AJ, Burger
PC, Gilles FH, Davis RL and Finlay JL: Children's Cancer Group: The
influence of central review on outcome associations in childhood
malignant gliomas: Results from the CCG-945 experience. Neuro
Oncol. 5:197–207. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ganigi PM, Santosh V, Anandh B,
Chandramouli BA and Sastry Kolluri VR: Expression of p53, EGFR, pRb
and bcl-2 proteins in pediatric glioblastoma multiforme: A study of
54 patients. Pediatr Neurosurg. 41:292–299. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nakamura M, Shimada K, Ishida E, Higuchi
T, Nakase H, Sakaki T and Konishi N: Molecular pathogenesis of
pediatric astrocytic tumors. Neuro Oncol. 9:113–123. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ohgaki H, Dessen P, Jourde B, Horstmann S,
Nishikawa T, Di Patre PL, Burkhard C, Schüler D, Probst-Hensch NM,
Maiorka PC, et al: Genetic pathways to glioblastoma: A
population-based study. Cancer Res. 64:6892–6899. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Di Sapio A, Morra I, Pradotto L, Guido M,
Schiffer D and Mauro A: Molecular genetic changes in a series of
neuroepithelial tumors of childhood. J Neurooncol. 59:117–122.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bredel M, Pollack IF, Hamilton RL and
James CD: Epidermal growth factor receptor expression and gene
amplification in high-grade non-brainstem gliomas of childhood.
Clin Cancer Res. 5:1786–1792. 1999.PubMed/NCBI
|
|
9
|
Kraus JA, Felsberg J, Tonn JC,
Reifenberger G and Pietsch T: Molecular genetic analysis of the
TP53, PTEN, CDKN2A, EGFR, CDK4 and MDM2 tumour-associated genes in
supratentorial primitive neuroectodermal tumours and glioblastomas
of childhood. Neuropathol Appl Neurobiol. 28:325–333. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Visone R and Croce CM: MiRNAs and cancer.
Am J Pathol. 174:1131–1138. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Munoz JL, Walker ND, Scotto KW and
Rameshwar P: Temozolomide competes for P-glycoprotein and
contributes to chemoresistance in glioblastoma cells. Cancer Lett.
367:69–75. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun C, Li N, Yang Z, Zhou B, He Y, Weng D,
Fang Y, Wu P, Chen P, Yang X, et al: miR-9 regulation of BRCA1 and
ovarian cancer sensitivity to cisplatin and PARP inhibition. J Natl
Cancer Inst. 105:1750–1758. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shen R, Wang Y, Wang CX, Yin M, Liu HL,
Chen JP, Han JQ and Wang WB: MiRNA-155 mediates TAM resistance by
modulating SOCS6-STAT3 signalling pathway in breast cancer. Am J
Transl Res. 7:2115–2126. 2015.PubMed/NCBI
|
|
16
|
Dong Z, Ren L, Lin L and Li J, Huang Y and
Li J: Effect of microRNA-21 on multidrug resistance reversal in
A549/DDP human lung cancer cells. Mol Med Rep. 11:682–690. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Blower PE, Chung JH, Verducci JS, Lin S,
Park JK, Dai Z, Liu CG, Schmittgen TD, Reinhold WC, Croce CM, et
al: MicroRNAs modulate the chemosensitivity of tumor cells. Mol
Cancer Ther. 7:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
González-Gómez P, Sánchez P and Mira H:
MicroRNAs as regulators of neural stem cell-related pathways in
glioblastoma multiforme. Mol Neurobiol. 44:235–249. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Berindan-Neagoe I, Monroig Pdel C,
Pasculli B and Calin GA: MicroRNAome genome: A treasure for cancer
diagnosis and therapy. CA Cancer J Clin. 64:311–336. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rajewsky N and Socci ND: Computational
identification of microRNA targets. Dev Biol. 267:529–535. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hummel R, Maurer J and Haier J: MicroRNAs
in brain tumors: A new diagnostic and therapeutic perspective? Mol
Neurobiol. 44:223–234. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M
and Croce CM: Human microRNA genes are frequently located at
fragile sites and genomic regions involved in cancers. Proc Natl
Acad Sci USA. 101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fabbri M, Ivan M, Cimmino A, Negrini M and
Calin GA: Regulatory mechanisms of microRNAs involvement in cancer.
Expert Opin Biol Ther. 7:1009–1019. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Møller HG, Rasmussen AP, Andersen HH,
Johnsen KB, Henriksen M and Duroux M: A systematic review of
microRNA in glioblastoma multiforme: Micro-modulators in the
mesenchymal mode of migration and invasion. Mol Neurobiol.
47:131–144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shea A, Harish V, Afzal Z, Chijioke J,
Kedir H, Dusmatova S, Roy A, Ramalinga M, Harris B, Blancato J, et
al: MicroRNAs in glioblastoma multiforme pathogenesis and
therapeutics. Cancer Med. 5:1917–1946. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ahir BK, Ozer H, Engelhard HH and Lakka
SS: MicroRNAs in glioblastoma pathogenesis and therapy: A
comprehensive review. Crit Rev Oncol Hematol. 120:22–33. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ciafrè SA, Galardi S, Mangiola A, Ferracin
M, Liu CG, Sabatino G, Negrini M, Maira G, Croce CM and Farace MG:
Extensive modulation of a set of microRNAs in primary glioblastoma.
Biochem Biophys Res Commun. 334:1351–1358. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chan JA, Krichevsky AM and Kosik KS:
MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells.
Cancer Res. 65:6029–6033. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Silber J, Lim DA, Petritsch C, Persson AI,
Maunakea AK, Yu M, Vandenberg SR, Ginzinger DG, James CD, Costello
JF, et al: miR-124 and miR-137 inhibit proliferation of
glioblastoma multiforme cells and induce differentiation of brain
tumor stem cells. BMC Med. 6:142008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Godlewski J, Nowicki MO, Bronisz A,
Williams S, Otsuki A, Nuovo G, Raychaudhury A, Newton HB, Chiocca
EA and Lawler S: Targeting of the Bmi-1 oncogene/stem cell renewal
factor by microRNA-128 inhibits glioma proliferation and
self-renewal. Cancer Res. 68:9125–9130. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lawler S and Chiocca EA: Emerging
functions of microRNAs in glioblastoma. J Neurooncol. 92:297–306.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Birks DK, Barton VN, Donson AM, Handler
MH, Vibhakar R and Foreman NK: Survey of MicroRNA expression in
pediatric brain tumors. Pediatr Blood Cancer. 56:211–216. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Miele E, Buttarelli FR, Arcella A, Begalli
F, Garg N, Silvano M, Po A, Baldi C, Carissimo G, Antonelli M, et
al: High-throughput microRNA profiling of pediatric high-grade
gliomas. Neuro Oncol. 16:228–240. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Braunstein S, Raleigh D, Bindra R, Mueller
S and Haas-Kogan D: Pediatric high-grade glioma: Current molecular
landscape and therapeutic approaches. J Neurooncol. 134:541–549.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Massimino M, Gandola L, Luksch R,
Spreafico F, Riva D, Solero C, Giangaspero F, Locatelli F, Podda M,
Bozzi F, et al: Sequential chemotherapy, high-dose thiotepa,
circulating progenitor cell rescue and radiotherapy for childhood
high-grade glioma. Neuro Oncol. 7:41–48. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Biassoni V, Casanova M, Spreafico F,
Gandola L and Massimino M: A case of relapsing glioblastoma
multiforme responding to vinorelbine. J Neurooncol. 80:195–201.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dvinge H and Bertone P: HTqPCR:
High-throughput analysis and visualization of quantitative
real-time PCR data in R. Bioinformatics. 25:3325–3326. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
R Development Core Team R: A language and
environment for statistical computing. R Foundation for Statistical
Computing; Vienna, Austria: ISBN 3-900051-07-0. http://www.R-project.org2008
|
|
42
|
Enright AJ, John B, Gaul U, Tuschl T,
Sander C and Marks DS: MicroRNA targets in drosophila. Genome Biol.
5:R12003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
The Gene Ontology Consortium: Expansion of
the gene ontology knowledgebase and resources. Nucleic Acids Res.
45:D331–D338. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kanehisa M, Furumichi M, Tanabe M, Sato Y
and Morishima K: KEGG: New perspectives on genomes, pathways,
diseases and drugs. Nucleic Acids Res. 45:D353–D361. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kanehisa M, Sato Y, Kawashima M, Furumichi
M and Tanabe M: KEGG as a reference resource for gene and protein
annotation. Nucleic Acids Res. 44:D457–D462. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Whitfield ML, Sherlock G, Saldanha AJ,
Murray JI, Ball CA, Alexander KE, Matese JC, Perou CM, Hurt MM,
Brown PO and Botstein D: Identification of genes periodically
expressed in the human cell cycle and their expression in tumors.
Mol Biol Cell. 13:1977–2000. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Drosos Y, Kouloukoussa M, Østvold AC,
Grundt K, Goutas N, Vlachodimitropoulos D, Havaki S, Kollia P,
Kittas C, Marinos E and Aleporou-Marinou V: NUCKS overexpression in
breast cancer. Cancer Cell Int. 9:192009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Walaas SI, Ostvold AC and Laland SG:
Phosphorylation of P1, a high mobility group-like protein,
catalyzed by casein kinase II protein kinase C, cyclic
AMP-dependent protein kinase and calcium/calmodulin-dependent
protein kinase II. FEBS Lett. 258:106–108. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ostvold AC, Norum JH, Mathiesen S, Wanvik
B, Sefland I and Grundt K: Molecular cloning of a mammalian nuclear
phosphoprotein NUCKS, which serves as a substrate for Cdk1 in vivo.
Eur J Biochem. 268:2430–2440. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Meijer L, Ostvold AC, Walass SI, Lund T
and Laland SG: High-mobility-group proteins P1, I and Y as
substrates of the M-phase-specific p34cdc2/cyclincdc13 kinase. Eur
J Biochem. 196:557–567. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Parplys AC, Zhao W, Sharma N, Groesser T,
Liang F, Maranon DG, Leung SG, Grundt K, Dray E, Idate R, et al:
NUCKS1 is a novel RAD51AP1 paralog important for homologous
recombination and genome stability. Nucleic Acids Res.
43:9817–9834. 2015.PubMed/NCBI
|
|
55
|
Kikuchi A, Ishikawa T, Mogushi K, Ishiguro
M, Iida S, Mizushima H, Uetake H, Tanaka H and Sugihara K:
Identification of NUCKS1 as a colorectal cancer prognostic marker
through integrated expression and copy number analysis. Int J
Cancer. 132:2295–2302. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Gu L, Xia B, Zhong L, Ma Y, Liu L, Yang L
and Lou G: NUCKS1 overexpression is a novel biomarker for
recurrence-free survival in cervical squamous cell carcinoma.
Tumour Biol. 35:7831–7836. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Cheong JY, Kim YB, Woo JH, Kim DK, Yeo M,
Yang SJ, Yang KS, Soon SK, Wang HJ, Kim BW, et al: Identification
of NUCKS1 as a putative oncogene and immunodiagnostic marker of
hepatocellular carcinoma. Gene. 584:47–53. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Balcárková J, Urbánková H, Scudla V,
Holzerová M, Bacovský J, Indrák K and Jarosová M: Gain of
chromosome arm 1q in patients in relapse and progression of
multiple myeloma. Cancer Genet Cytogenet. 192:68–72. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Szponar A, Zubakov D, Pawlak J, Jauch A
and Kovacs G: Three genetic developmental stages of papillary renal
cell tumors: Duplication of chromosome 1q marks fatal progression.
Int J Cancer. 124:2071–2076. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Giunti L, Pantaleo M, Sardi I, Provenzano
A, Magi A, Cardellicchio S, Castiglione F, Tattini L, Novara F,
Buccoliero AM, et al: Genome-wide copy number analysis in pediatric
glioblastoma multiforme. Am J Cancer Res. 4:293–303.
2014.PubMed/NCBI
|
|
61
|
Faria C, Miguéns J, Antunes JL, Salgado D,
Nunes S, Barroso C, Martins Mdo C, Nunes VM and Roque L: Pediatric
brain tumors: Genetics and clinical outcome. J Neurosurg Pediatr.
5:263–270. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hirose Y, Aldape K, Bollen A, James CD,
Brat D, Lamborn K, Berger M and Feuerstein BG: Chromosomal
abnormalities subdivide ependymal tumors into clinically relevant
groups. Am J Pathol. 158:1137–1143. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Lo KC, Ma C, Bundy BN, Pomeroy SL,
Eberhart CG and Cowell JK: Gain of 1q is a potential univariate
negative prognostic marker for survival in medulloblastoma. Clin
Cancer Res. 13:7022–7028. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Shen H, Wang L, Ge X, Jiang CF, Shi ZM, Li
DM, Liu WT, Yu X and Shu YQ: MicroRNA-137 inhibits tumor growth and
sensitizes chemosensitivity to paclitaxel and cisplatin in lung
cancer. Oncotarget. 7:20728–20742. 2016.PubMed/NCBI
|
|
65
|
Yang Y, Li F, Saha MN, Abdi J, Qiu L and
Chang H: miR-137 and miR-197 induce apoptosis and suppress
tumorigenicity by targeting MCL-1 in multiple myeloma. Clin Cancer
Res. 21:2399–2411. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Liu LL, Lu SX, Li M, Li LZ, Fu J, Hu W,
Yang YZ, Luo RZ, Zhang CZ and Yun JP: FoxD3-regulated microRNA-137
suppresses tumour growth and metastasis in human hepatocellular
carcinoma by targeting AKT2. Oncotarget. 5:5113–5124. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Chen DL, Wang DS, Wu WJ, Zeng ZL, Luo HY,
Qiu MZ, Ren C, Zhang DS, Wang ZQ, Wang FH, et al: Overexpression of
paxillin induced by miR-137 suppression promotes tumor progression
and metastasis in colorectal cancer. Carcinogenesis. 34:803–811.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhu X, Li Y, Shen H, Li H, Long L, Hui L
and Xu W: miR-137 inhibits the proliferation of lung cancer cells
by targeting Cdc42 and Cdk6. FEBS Lett. 587:73–81. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Chen L, Wang X, Wang H, Li Y, Yan W, Han
L, Zhang K, Zhang J, Wang Y, Feng Y, et al: miR-137 is frequently
down-regulated in glioblastoma and is a negative regulator of
Cox-2. Eur J Cancer. 48:3104–3111. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Bier A, Giladi N, Kronfeld N, Lee HK,
Cazacu S, Finniss S, Xiang C, Poisson L, deCarvalho AC, Slavin S,
et al: MicroRNA-137 is downregulated in glioblastoma and inhibits
the stemness of glioma stem cells by targeting RTVP-1. Oncotarget.
4:665–676. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Giunti L, da Ros M, Vinci S, Gelmini S,
Iorio AL, Buccoliero AM, Cardellicchio S, Castiglione F, Genitori
L, de Martino M, et al: Anti-miR21 oligonucleotide enhances
chemosensitivity of T98G cell line to doxorubicin by inducing
apoptosis. Am J Cancer Res. 5:231–242. 2014.PubMed/NCBI
|
|
72
|
Li D, Shan W, Fang Y, Wang P and Li J:
miR-137 acts as a tumor suppressor via inhibiting CXCL12 in human
glioblastoma. Oncotarget. 8:101262–101270. 2017.PubMed/NCBI
|
|
73
|
Chen W, Ye L, Wen D and Chen F: miR-490-5p
inhibits hepatocellular carcinoma cell proliferation, migration and
invasion by directly regulating ROBO1. Pathol Oncol Res. Sep
19–2017.(Epub ahead of print).
|
|
74
|
Xu B, Xu T, Liu H, Min Q, Wang S and Song
Q: miR-490-5p suppresses cell proliferation and invasion by
targeting BUB1 in hepatocellular carcinoma cells. Pharmacology.
100:269–282. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Tang B, Liu C, Zhang QM and Ni M:
Decreased expression of miR-490-3p in osteosarcoma and its clinical
significance. Eur Rev Med Pharmacol Sci. 21:246–251.
2017.PubMed/NCBI
|
|
76
|
Li J, Feng Q, Wei X and Yu Y: MicroRNA-490
regulates lung cancer metastasis by targeting poly r(C)-binding
protein 1. Tumour Biol. 37:15221–15228. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Tian J, Xu YY, Li L and Hao Q: miR-490-3p
sensitizes ovarian cancer cells to cisplatin by directly targeting
ABCC2. Am J Transl Res. 9:1127–1138. 2017.PubMed/NCBI
|
|
78
|
Wu X, Yan L, Liu Y, Xian W, Wang L and
Ding X: MicroRNA-448 suppresses osteosarcoma cell proliferation and
invasion through targeting EPHA7. PLoS One. 12:e01755532017.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhu H, Zhou X, Ma C, Chang H, Li H, Liu F
and Lu J: Low expression of miR-448 induces EMT and promotes
invasion by regulating ROCK2 in hepatocellular carcinoma. Cell
Physiol Biochem. 36:487–498. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Lv Y, Lei Y, Hu Y, Ding W, Zhang C and
Fang C: miR-448 negatively regulates ovarian cancer cell growth and
metastasis by targeting CXCL12. Clin Transl Oncol. 17:903–909.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Bamodu OA, Huang WC, Lee WH, Wu A, Wang
LS, Hsiao M, Yeh CT and Chao TY: Aberrant KDM5B expression promotes
aggressive breast cancer through MALAT1 overexpression and
downregulation of hsa-miR-448. BMC Cancer. 16:1602016. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Correia NC, Melão A, Póvoa V, Sarmento L,
Gómez de Cedrón M, Malumbres M, Enguita FJ and Barata JT: microRNAs
regulate TAL1 expression in T-cell acute lymphoblastic leukemia.
Oncotarget. 7:8268–8281. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Powrózek T, Krawczyk P, Kowalski DM,
Kuźnar-Kamińska B, Winiarczyk K, Olszyna-Serementa M,
Batura-Gabryel H and Milanowski J: Application of plasma
circulating microRNA-448, 506, 4316, and 4478 analysis for
non-invasive diagnosis of lung cancer. Tumour Biol. 37:2049–2055.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Hara N, Kikuchi M, Miyashita A, Hatsuta H,
Saito Y, Kasuga K, Murayama S, Ikeuchi T and Kuwano R: Serum
microRNA miR-501-3p as a potential biomarker related to the
progression of Alzheimer's disease. Acta Neuropathol Commun.
5:102017. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Ling Q, Xu X, Ye P, Xie H, Gao F, Hu Q,
Liu Z, Wei X, Röder C, Trauzold A, et al: The prognostic relevance
of primary tumor location in patients undergoing resection for
pancreatic ductal adenocarcinoma. Oncotarget. 8:15159–15167. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Jiang X, Wang W, Yang Y, Du L, Yang X,
Wang L, Zheng G, Duan W, Wang R, Zhang X, et al: Identification of
circulating microRNA signatures as potential noninvasive biomarkers
for prediction and prognosis of lymph node metastasis in gastric
cancer. Oncotarget. 8:65132–65142. 2017.PubMed/NCBI
|
|
87
|
Huang Y, Liao D, Pan L, Ye R, Li X, Wang
S, Ye C and Chen L: Expressions of miRNAs in papillary thyroid
carcinoma and their associations with the BRAFV600E mutation. Eur J
Endocrinol. 168:675–681. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Paydas S, Acikalin A, Ergin M, Celik H,
Yavuz B and Tanriverdi K: Micro-RNA (miRNA) profile in Hodgkin
lymphoma: Association between clinical and pathological variables.
Med Oncol. 33:342016. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Bao L, Lv L, Feng J, Chen Y, Wang X, Han S
and Zhao H: miR-876-5p suppresses epithelial-mesenchymal transition
of lung cancer by directly down-regulating bone morphogenetic
protein 4. J Biosci. 42:671–681. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Riemenschneider MJ, Jeuken JW, Wesseling P
and Reifenberger G: Molecular diagnostics of gliomas: State of the
art. Acta Neuropathol. 120:567–584. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Conway C, Beswick S, Elliott F, Chang YM,
Randerson-Moor J, Harland M, Affleck P, Marsden J, Sanders DS, Boon
A, et al: Deletion at chromosome arm 9p in relation to BRAF/NRAS
mutations and prognostic significance for primary melanoma. Genes
Chromosomes Cancer. 49:425–438. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Giunti L, da Ros M, Vinci S, Gelmini S,
Iorio AL, Buccoliero AM, Cardellicchio S, Castiglione F, Genitori
L, de Martino M, et al: Anti-miR21 oligonucleotide enhances
chemosensitivity of T98G cell line to doxorubicin by inducing
apoptosis. Am J Cancer Res. 5:231–242. 2014.PubMed/NCBI
|