Introduction
Malignant tumors are amongst the leading causes of
human mortality (1). The current
methods of treating tumors are predominantly radiotherapy and
chemotherapy, which improve the therapeutic outcomes of patients
with malignant tumors; however, the side effects are a major
obstacle in some patients (2).
Numerous patients who do not respond to traditional
therapies frequently seek adjuvant and alternative medical
treatments in the hope of reducing tumor size and enhancing
immunity (3). In Western high-income
countries, the use of complementary and alternative medicine (CAM)
in patients with cancer is as high as 91% (4,5).
Immunostimulants can enhance the body's immunity, and one of the
research directions of CAM is to find safe and effective immune
stimulants that can be used in clinical practice to prevent
diseases (6,7).
Flor·Essence (FE) Tonics is a complex mixture of
commercially available herbal extracts, and is sold as a dietary
supplement and used by patients with cancer due to reports that it
may aid in treating or preventing disease (8). Large quantities of supplements are
shipped to the US and Canada annually, primarily for use in cancer
treatment (9-13).
FE contains eight different types of herb, which
predominantly contain phenolics. The FE formula is proprietary;
however, the composition in descending order of magnitude is as
follows: i) Burdock root; ii) sheep sorrel herb; iii) slippery elm
bark; iv) watercress herb; v) kelp; vi) blessed thistle herb; vii)
red clover herb; and viii) Turkish rhubarb root (14). It has been reported that kelp may
reduce the incidence of cancer by affecting oxidative activity,
specifically in liver cancer and lung cancer (15,16).
Arctigenin inhibits mitogen-activated protein kinases, including
ERK1/2, p38 kinase and JNK, resulting in an increase in the
activity of NAPDH oxidase 2, tumor necrosis factor-α (TNF-α) and
transforming growth factor-β1(17).
Dietary isoflavones inhibit the progression of prostate cancer by
inducing apoptosis in tumor cells (18). It has been reported FE has various
positive functions, including antioxidant, antiestrogen,
immunostimulatory and antitumor effects (8,19,20).
Cheung et al (20) reported
that FE inhibited nitric oxide (NO) production in a
concentration-dependent manner in lipopolysaccharide
(LPS)-stimulated RAW 264.7 cells. Reverse
transcription-quantitative PCR assays revealed that FE induced the
expression of IL-1β, inducible nitric oxide synthase and
cyclooxygenase-2 in RAW 264.7 cells.
In the present study, an immunosuppressive mouse
model was established using cyclophosphamide (CP) to explore the
effects of FE on the side effects of chemotherapy.
Materials and methods
Sample preparation
FE was obtained from Shenzhen Fulan Trading Co.,
Ltd. (Flora Manufacturing & Distributing, Ltd.). FE was boiled
with distilled water overnight in a small bowl. Samples were
filtered with a 200-µm nylon mesh. Then, it was cooled and
refrigerated at 4˚C. Concentration of 10, 20 and 40 mg/kg FE were
prepared for oral administration in mice.
Animals and supplementation with
FE
Specific pathogen-free Kunming mice (male, 6-8 weeks
old, 18-22 g) were supplied by the Experimental Animal Center of
Dalian Medical University. The mice were housed under standard
laboratory conditions (25˚C, 40-60% relative humidity, under a
12:12 h light/dark cycle) and were fed with standard rodent food
and water ad libitum. Mice were monitored and weighed daily.
A total of 80 mice were randomly divided (20/group) into control
group (saline), low-dose FE group (10 mg/kg), medium-dose group (20
mg/kg) and high-dose group (40 mg/kg). FE was administered via
gavage twice daily for 5 weeks.
Measurement of immune organs and
weight
Mice were sacrificed by CO2 asphyxiation
after a final weight measurement (3 mice/group). The spleen and
thymus were removed aseptically, dried and weighed. The organ index
was calculated as follows: Thymus index = thymic weight (mg)/animal
weight (g) x100%; spleen index = spleen weight (mg)/animal weight
(g) x100%.
Spleen lymphocyte isolation
Spleens were aseptically removed and homogenized
through 200-µm nylon mesh (three mice per group). To prepare a
single cell suspension, red blood cell lysate (Beijing Solarbio
Science & Biotechnology Co., Ltd.) was added, allowed to stand
for 8 min, and centrifuged at 800 x g for 30 min at room
temperature. The lymphocyte layer was aspirated and resuspended in
RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc.) for density
gradient centrifugation at 250 x g for 10 min at room temperature.
The cells were cultured in RPMI-1640 supplemented with 10% FBS
(cat. no. 10099141; Gibco, Thermo Fisher Scientific, Inc.). All
cells were maintained in a humidified atmosphere at 37˚C in 5%
CO2.
MTT assay for cell viability
Each group of mouse spleen cells were seeded at
4x104/well in 96-well plates and cultured in RPMI-1640
supplemented with 10% FBS at 37˚C with 5% CO2. LPS (100
µg/ml; cat. no. L2630; Sigma-Aldrich; Merck KGaA) was added to the
culture medium for 24 h, following which MTT reagent (5 mg/ml; cat.
no. V13154; Gibco; Thermo Fisher Scientific, Inc.) was added to the
cell medium and cells were incubated at 37˚C for an additional 4 h.
The reaction was terminated with 150 µl/well DMSO and the cells
were lysed for 15 min, with the plates agitated every 5 min.
Absorbance values were determined using a microplate reader at 492
nm.
Peritoneal cell isolation
Mice were fasted for >8 h before surgery, and
mice were sacrificed by CO2 asphyxiation (3 mice/group).
The mice were soaked for 3 min, the skin was cut open and the
peritoneum was fully exposed. Pre-cooled 1640 medium (5 ml) was
intraperitoneally injected, and the abdomen of the mouse was
incubated for 5 min. The peritoneal lavage fluid was withdrawn by a
syringe and collected in a 15-ml centrifuge tube; this was repeated
twice. The supernatant was centrifuged at 4˚C and 450 x g for 8
min, and red blood cell lysate was added until a white cell pellet
was observed. The cells were cultured in RPMI-1640 supplemented
with 10% FBS, and then seeded into 96-well culture plates. Cells
were maintained in a humidified atmosphere at 37˚C in 5%
CO2.
Phagocytosis neutral red method
Peritoneal macrophages were obtained, 4% trypan blue
was mixed with cells at a concentration of 9:1 at room temperature
(final concentration, 0.04%), and cells counted after ~3 min and
incubated at 37˚C for 2 h to allow cells to adhere. The cells were
washed twice with D-Hanks solution (Beijing Solarbio Science &
Biotechnology Co., Ltd.) to remove unattached cells. Neutral red
(100 µl) was added to each well, and after 4 h at 37˚C, wells were
washed 3 times with D-Hanks solution. After adding 100 µl of cell
lysate, after 30 min, the optical density (OD) value of each well
was measured at 490 nm by a microplate reader.
Semi-quantitative PCR
TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.) reagent was used to extract total RNA from spleen
lymphocytes. Then, a ProtoScript II RT-PCR kit (Takara
Biotechnology Co., Ltd.) (21) was
used to reverse transcribe RNA. qPCR analysis of interferon
(IFN)-α, IFN-β and GAPDH was conducted using the following
gene-specific primers: IFN-α, forward, 5'-ATGAGCACTGAAAGCATGATC-3'
and reverse, 5'-TCACAGGGCAATGATCCCAAAGTAGACCTG CCC-3'; IFN-β,
forward, 5'-CTTGAATTCACTGCTCTC CTG-3' and reverse,
5'-CGGCTCGAGTCAGTTAGGGAG-3'; GAPDH, forward,
5'-CTAGCACCCCTGGCCAAG-3' and reverse, 5'-GATGTTCTGGAGAGCCCCG-3'.
PCR was conducted as follows: An initial denaturation step at 94˚C
for 5 min, followed by 30 cycles of annealing for 30 sec, 53˚C for
30 sec and 72˚C for 20 sec, and a final extension step at 72˚C for
10 min. PCR products were resolved via 1% agarose electrophoresis
and visualized using ethidium bromide. Quantitative analysis was
performed using ImageQuant TL 7.0 software (GE Healthcare Life
Sciences).
Lactate dehydrogenase (LDH) assay of
the viability of natural killer (NK) cells
Mice were sacrificed by CO2 asphyxiation
and soaked in 75% ethanol, and the spleens of the mice were removed
(3 mice/group). Mouse 1X lymphocyte separation solution (4-5 ml;
cat. no. P8620; Beijing Solarbio Science & Technology Co.,
Ltd.) was added to the culture dish for grinding. Immediately, the
lymphocyte supernatant containing the spleen cells was transferred
to a 15-ml centrifuge tube, 200-500 µl of RPMI-1640 medium was
added (maintaining a clear liquid boundary), the lymphocyte layer
was aspirated and a further 10 ml of RPMI-1640 medium was added.
The solution was centrifuged at 800 x g for 30 min at room
temperature. The cells were collected via further centrifugation at
250 x g for 10 min at room temperature. The supernatant was
decanted and the cells were cultured in RPMI-1640 supplemented with
10% FBS. Cells were maintained in a humidified atmosphere at 37˚C
in 5% CO2.
The cytotoxicity of NK cells towards K562 cells
(cat. no. BNCC339825; BeNa Culture Collection) was evaluated via
co-culture (50:1) under standard conditions for 24 h at 37˚C in 5%
CO2, followed by using a CytoTox 96® LDH
assay kit according to the manufacturer's protocol (cat. no.
A020-2-2; Nanjing Jiancheng Bio-Engineering Institute Co.,
Ltd.).
The percentage of cytotoxicity was calculated using
the formula below:
% Cytotoxicity =
(ODsample-ODcontrast)/(ODstandard-ODblank)x100
Measurement of cytokines
The expression of IL-12, IFN-γ, IL-2 and TNF-α was
measured using double antibody sandwich ELISA kits (IL-12, cat. no.
BMS6004; IFN-γ, cat. no. BMS228; IL-2, cat. no. BMS601; TNF-α, cat.
no. BMS607-3; all from Invitrogen; Thermo Fisher Scientific, Inc.).
During euthanasia, blood (0.5 ml) was collected from each mouse (3
mice/group) via retro-orbital sampling, allowed to stand at room
temperature for 2 h and at 4˚C for 3-4 h, centrifuged at 800 x g
for 5 min at room temperature and stored at -20˚C. The supernatant
from each group was collected, and their levels were quantitated by
ELISA according to the manufacturer's protocols.
Western blot analysis
Spleens were aseptically removed and homogenized
through 200-µm nylon mesh. Equal quantities (100 µg/lane) of spleen
protein were analyzed via western blot analysis as previously
described (22). Membranes were
incubated with primary antibodies at 4˚C overnight, and secondary
antibodies [horseradish peroxidase-conjugated goat anti-rabbit and
anti-mouse IgG (1:8,000; cat. nos. TA140003 and TA130003,
respectively; OriGene Technologies, Inc.)] at room temperature for
1 h. The following primary antibodies were used: Anti-caveolin-1
(Cav-1; mouse monoclonal; 1:1,000; cat. no. 03-6000; Invitrogen;
Thermo Fisher Scientific, Inc.); anti-NF-κB (rabbit monoclonal;
1:1,000; cat. no. 8242T; Cell Signaling Technology, Inc.);
anti-proliferating cell nuclear antigen (PCNA; mouse monoclonal;
1:500; cat. no. BM0104; Wuhan Boster Biological Technology, Ltd.);
and anti-β-actin (mouse monoclonal; 1:1,000; cat. no. BM0627; Wuhan
Boster Biological Technology, Ltd.). Bands were visualized using
the BeyoECL Plus reagent (cat. no. P0018S; Beyotime Institute of
Biotechnology) according to the manufacturer's protocols.
Immunoblot data were quantified using ImageJ software (version
1.46; National Institutes of Health).
CP preparation of mouse model of
immunosuppression
Specific pathogen-free Kunming mice (male, 6-8 weeks
old, 18-22 g) were provided by the Experimental Animal Center of
Dalian Medical University. A total of 90 mice were randomly divided
(45/group) into a control group and CP group. The 45 mice in each
group were randomized into 3 groups (15/group): Control group
(saline), low-dose FE group (10 mg/kg) and high-dose FE group (40
mg/kg). Mice were intraperitoneally injected with physiological
saline or CP (75 mg/mg; Beijing Solarbio Science &
Biotechnology Co., Ltd.) once every other day with CP for 8 days.
At 8 days later, immunological indicators of the control and CP
groups were measured to verify whether the immunocompromised mouse
model was successfully constructed. Then, all mice were
intragastrically administered with saline, or 10 or 40 mg/kg FE
twice a day for 5 weeks, and the tissues were extracted on the last
day.
Determination of the number of white
blood cells
During euthanasia, blood (0.5 ml) was collected from
each mouse (3 mice/group) via retro-orbital sampling, allowed to
stand at room temperature for 2 h and at 4˚C for 3-4 h, centrifuged
at 800 x g for 5 min at room temperature and stored at -20˚C.
Leukocyte diluent (2.0 ml glacial acetic acid, 1 ml 1% gentian
violet, 100 ml distilled water) was prepared and filtered, and 0.19
ml of this solution was added to 20 µl peripheral blood using a
hemoglobin pipette. The pipette was used to mix the blood and
diluent thoroughly; the micropipette quickly absorbs the mixed
suspensions, which was added to a counting cell. After standing for
2-3 min, the cell was microscopically examined; the total number of
white blood cells in the four corner squares was counted using an
optical inverted microscope (magnification, x40).
Statistical analyses
All data were expressed as the mean ± SE. Unpaired
Student's t-tests were used to analyze significant differences
between the control and CP groups. Comparisons of multiple groups
were analyzed using one- or two-way ANOVA followed by post hoc
Tukey's test. P<0.05 was considered to indicate a statistically
significant difference.
Results
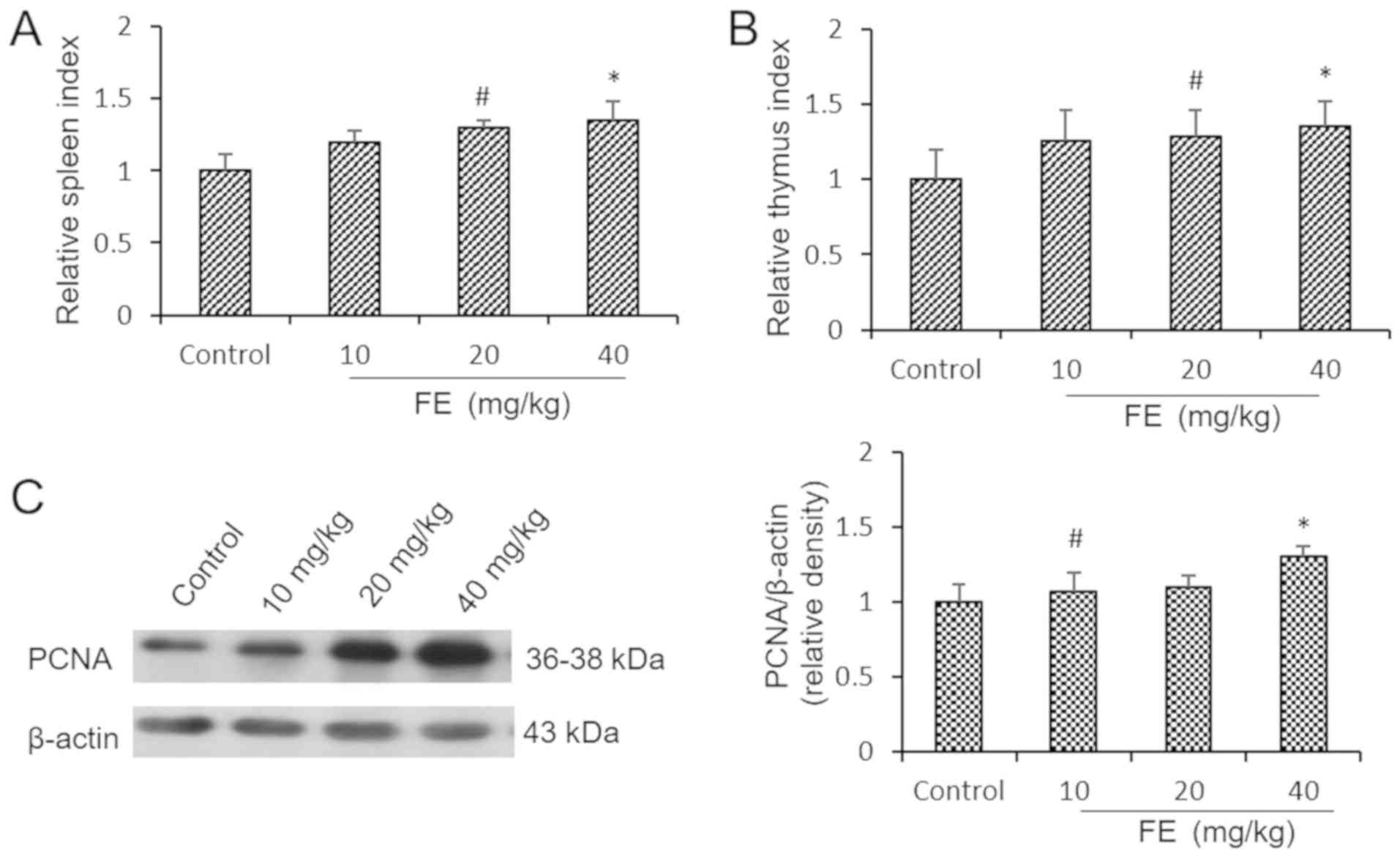
Effects of FE on immune organs,
splenocytes and PCNA
The spleen and thymic indexes of the mice
supplemented with FE were significantly higher than the control
group following 5 weeks of supplementation (Fig. 1A and B). In almost all rapidly proliferating
cells, PCNA is important for DNA replication and cell cycle
regulation, and is a key factor for measuring cell proliferation
ability (23,24). The effects of FE on proliferation in
the immune organs were investigated by analyzing PCNA expression in
splenocytes. The levels of PCNA in the mice administered with 10 or
40 mg/kg FE were significantly higher than in the control group
(Fig. 1C). These results suggested
that FE can enhance the viability of immune cells.
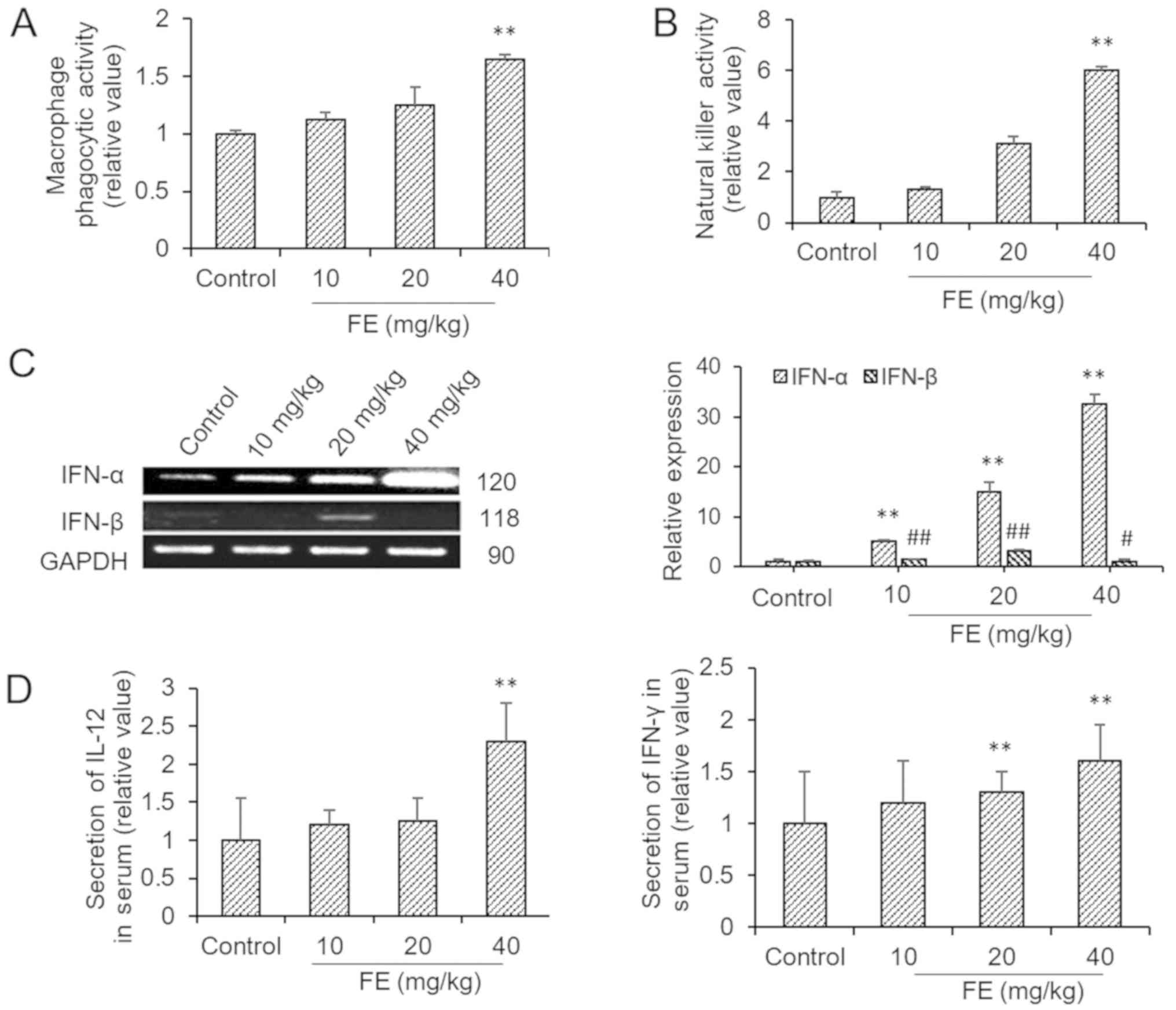
Effects of FE on immune cell activity
and cytokine production
The macrophages obtained from mice which had been
orally administered with FE had significantly increased phagocytic
activity against neutral red (Fig.
2A). Mice supplemented with FE exhibited increased NK
cytotoxicity compared with the control (Fig. 2B). In addition, FE also induced the
production of IFN-α and IFN-β, which were evaluated via PCR. The
data demonstrated that spleen lymphocytes from FE-supplemented mice
expressed higher levels of IFN-α and IFN-β than those from the
controls (Fig. 2C). It was also
shown that FE supplementation significantly increased the secretion
of IL-12 and IFN-γ into the blood (Fig.
2D).
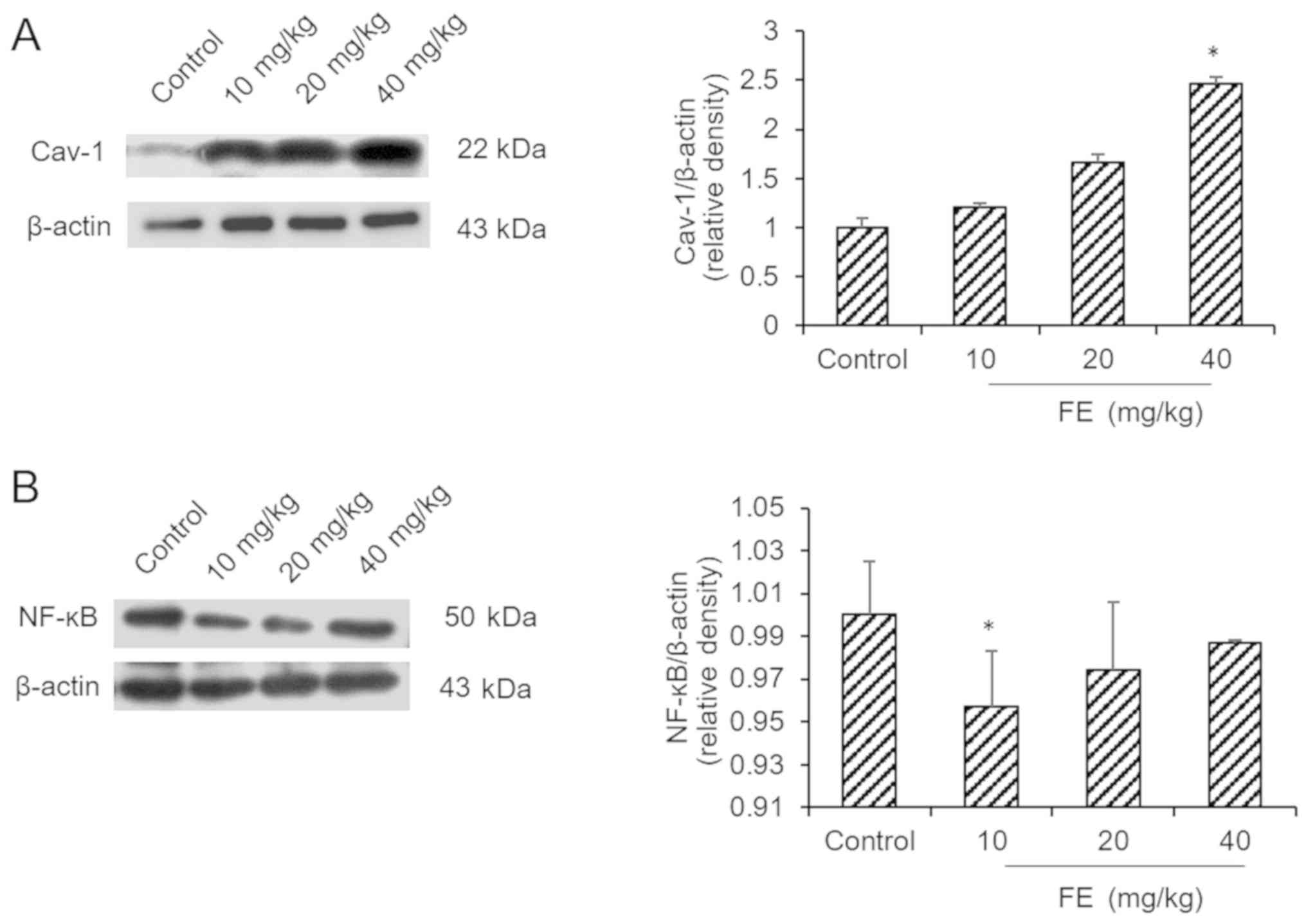
Effects of FE on the expression of
NF-κB and Cav-1
The results indicated that the mice supplemented
with FE expressed more Cav-1 than the control mice (Fig. 3A). Additionally, it was demonstrated
that the expression of NF-κB was downregulated in 10 mg/kg FE group
compared with the control (Fig.
3B).
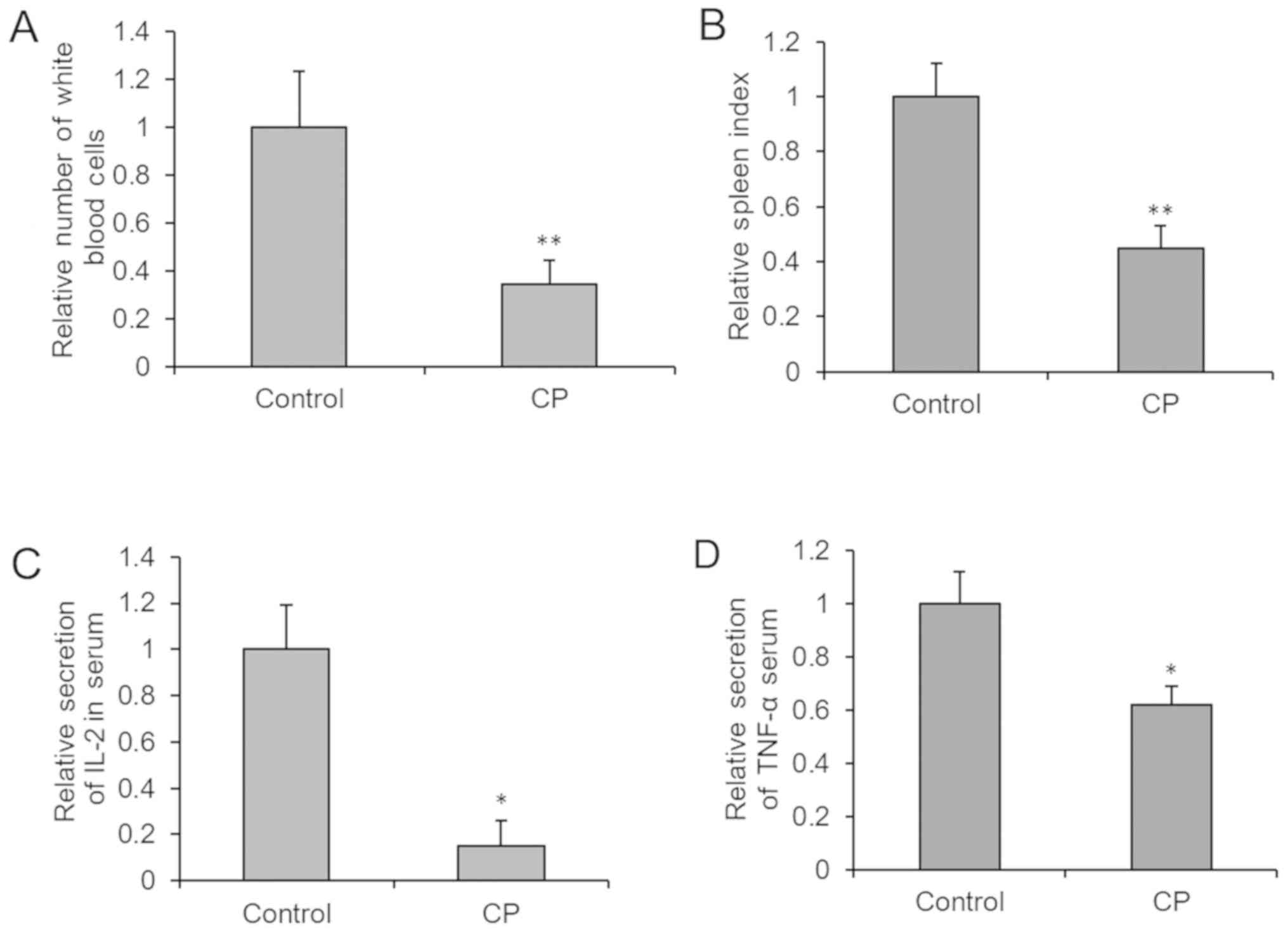
Developing a mouse model of
immunosuppression
CP was administered to mice via intraperitoneal
injection every other day for 8 days (Fig. 4). Immunological parameters were
measured, and immune damage and immunosuppression was observed.
Compared with the control group, the white blood cell count, spleen
index, and IL-2 and TNF-α levels in serum were decreased
significantly. This indicated that the immune function of CP group
was compromised.
Effects of FE on immune cells in the
mouse model of immunosuppression
The previous results obtained during the present
study indicated that FE can increase the relative spleen size of
mice, the phagocytic ability of peritoneal macrophages and the
antitumor cytotoxicity of NK cells against K562 cells. To further
investigate the effects of FE, a CP-induced model of
immunosuppression was established, and the effects of treatment of
these mice with FE on immune cell properties were analyzed.
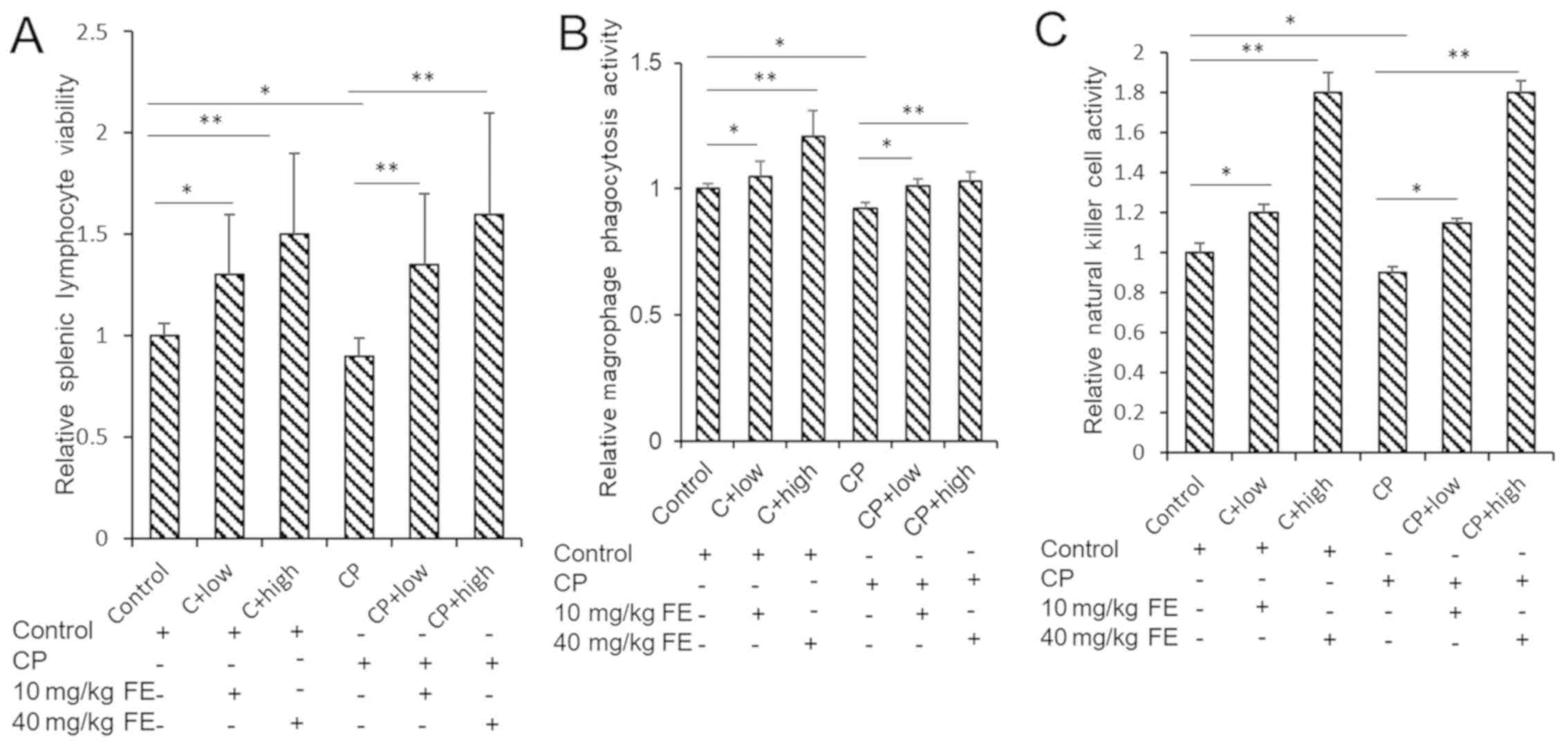
The viability of spleen lymphocytes in control- or
FE-treated normal mice and mice immunized with CP was detected. The
results showed that the activity of spleen lymphocytes in the CP
group was significantly decreased, and low (10 mg/kg) and high (40
mg/kg) concentrations of FE attenuated the effects of CP on cell
viability. It was shown that FE could significantly increase the
activity of spleen lymphocytes from immunosuppressed mice (Fig. 5A).
The phagocytic activity of peritoneal macrophages in
normal mice and CP mice was detected. FE (10 and 40 mg/kg) was used
to promote the immune activity of mice. The results showed that the
phagocytic ability of macrophages was significantly increased after
oral FE administration (Fig.
5B).
The killing activity of NK cells in normal mice and
CP mice was detected. NK cell killing ability of mice after oral
administration of FE was detected. The results showed that the
ability of NK cells to kill cancer cells was significantly
increased after FE administration in normal and immunosuppressed
mice (Fig. 5C).
Discussion
The effects of FE on immune cell activity and
cytokine production were investigated. Neutral red is recognized as
a foreign substance by macrophages, and the present findings showed
that FE treatment increases the phagocytic activity of macrophages
isolated from the peritoneum of mice. NK cells are lymphocytes
associated with autoimmune conditions that form an early immune
defense during innate immunity (25). It was previously reported that the
number of NK cells was reduced in patients with non-small cell lung
cancer and melanoma (26). Active
hexose correlated compound (AHCC) is an edible fungal extract that
can affect immune responses when ingested (27). In vivo and in vitro
studies have reported that the immunomodulatory effects of AHCC
enhance the activity of NK, CD8+ T and γδT cells
(27). The present study showed that
FE can enhance the killing ability of mouse NK cells, indicating
that FE promotes innate immune responses. There is a strong
association between NK activity and changes in spleen cells
(28). In addition, FE also induced
the production of IFN-α and IFN-β as determined via
semi-quantitative PCR analysis. The data demonstrated that
FE-supplemented mice exhibited greater IFN-α and IFN-β mRNA
expression than control mice. IFN-α and IFN-β can activate NK
cells, induce antiviral states and inhibit viral republication,
which are important for host defenses against viruses (29). IL-12 is a stimulating factor of NK
cells, inducing the production of IFN and TNF, and activating NK
cells to stimulate their differentiation, enabling NK cells to
produce more TNF-α; therefore, IL-12 and TNF-α are associated with
adaptive immunity (30-32).
IFN-γ is an important component of the body's immune system, and is
produced by activated T cells and NK cells (33). It can effectively resist viral
infection and tumor formation, and promote the body's immune
response (34). Therefore, IL-12 and
IFN-γ levels were evaluated in peripheral blood via ELISA. Results
showed that FE-supplemented mice exhibited increased secretion of
IFN-γ and IL-12.
FE has previously been reported to enhance cell
inflammation. Cheung et al (20) reported that FE inhibits NO production
in a concentration-dependent manner in LPS-stimulated RAW 264.7
cells. Long et al (35)
reported that ovarian cancer stem cells upregulate chemokine ligand
5 and activate NF-κB signaling through autocrine signaling, and
induce tumor metastasis. Inhibiting the key inflammatory
transcription factor NF-κB may suppress tumor invasion (36). NF-κB is associated with cancer, and
is located downstream of the oncogenes Ras, Myc and Ret (37). In addition, NF-κB plays a key role in
connecting inflammation with cancer (38). In the present study, it was
demonstrated that expression of NF-κB was downregulated in animals
supplemented with FE, suggesting that FE may dampen inflammation
through NF-κB.
Cav-1, a structural protein of the cell membrane, is
involved in tissue homeostasis, inflammation, oxidative stress,
microbial clearance, and fibrosis (39). Cav-1 serves an important role in
regulating inflammatory cell signaling in leukocytes (40). A previous study reported interactions
between Cav-1 and Toll-like receptor 4, as detected by
immunofluorescence in MCF-10A and MCF-10ACE cells (40). This interaction may influence the
downstream regulation of inflammation-associated gene expression
(41). Therefore, the expression of
inflammation-related protein Cav-1 was analyzed. The results
indicated that the mice supplemented with FE expressed more Cav-1
than the mice without FE, suggesting that FE may suppress
inflammation via enhanced Cav-1 expression.
In the present study, increases in the proliferation
of splenocytes, phagocytosis of macrophages and cytotoxicity of NK
cells towards K562 cells were observed following FE treatment in a
mouse model of CP-induced immunosuppression. The mechanism of
CP-induced immunosuppression is similar to that of nitrogen mustard
(42). CP is crosslinked with DNA to
inhibit the synthesis of DNA and interfere with RNA function,
inhibit tumor growth; however, when killing tumor cells, CP can
damage the immune organs and inhibit humoral immunity (43). The enhancement effects of FE against
immunosuppression suggested that FE may be effective as an adjuvant
to chemotherapy or for use in patients with reduced immune
function.
Acknowledgements
Not applicable.
Funding
This study was supported by Scientific and
Technological Developing Scheme of Ji Lin Province, Liaoning
Provincial Natural Science Foundation (grant no. 2015020568) and
Liaoning Provincial Department of Education Research Project (grant
no. L201783647)
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JW, JL and CQ were involved in conceiving the study,
acquiring and analyzing the data, and writing the original draft.
YW, YZhu, YZha and HL were involved in designing the study and
analyzing the data. BZ and YS analyzed data. WZ designed the
project, and acquired funding and resources. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Liaoning Normal University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yuan P, Chen TH, Chen ZW and Lin XQ:
Calculation of life-time death probability due malignant tumors
based on a sampling survey area in China. Asian Pac J Cancer Prev.
15:4307–4309. 2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Tai J, Cheung S, Wong S and Lowe C: In
vitro comparison of Essiac and Flor-Essence on human tumor cell
lines. Oncol Rep. 11:471–476. 2004.PubMed/NCBI
|
|
3
|
Nejatian M, Alami A, Tehrani H,
Lael-Monfared E and Jafari A: Perceptions and personal use of
complementary and alternative medicine (CAM) by Iranian health care
providers. Complement Ther Clin Pract. 32:145–150. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kuo YH, Tsay SL, Chang CC, Liao YC and
Tung HH: Cancer impact, complementary/alternative medicine beliefs,
and quality of life in cancer patients. J Altern Complement Med.
24:276–281. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Farooqui M, Hassali MA, Shatar AK,
Farooqui MA, Saleem F, Haq NU and Othman CN: Use of complementary
and alternative medicines among Malaysian cancer patients: A
descriptive study. J Tradit Complement Med. 6:321–326.
2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Hunault J, Diswall M, Frison JC, Blot V,
Rocher J, Marionneau-Lambot S, Oullier T, Douillard JY, Guillarme
S, Saluzzo C, et al: 3-Fluoro- and 3,3-difluoro-3,4-dideoxy-KRN7000
analogues as new potent immunostimulator agents: Total synthesis
and biological evaluation in human invariant natural killer T cells
and mice. J Med Chem. 55:1227–1241. 2012.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Yeap SK, Omar AR, Ho WY, Beh BK, Ali AM
and Alitheen NB: Rhaphidophora korthalsii modulates
peripheral blood natural killer cell proliferation, cytokine
secretion and cytotoxicity. BMC complement Altern Med.
13(145)2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kulp KS, Montgomery JL, Nelson DO, Cutter
B, Latham ER, Shattuck DL, Klotz DM and Bennett LM: Essiac and
Flor-Essence herbal tonics stimulate the in vitro growth of human
breast cancer cells. Breast Cancer Res Treat. 98:249–259.
2006.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Richardson MA, Sanders T, Palmer JL,
Greisinger A and Singletary SE: Complementary/alternative medicine
use in a comprehensive cancer center and the implications for
oncology. J Clin Oncol. 18:2505–2514. 2000.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Seely D, Kennedy DA, Myers SP, Cheras PA,
Lin D, Li R, Cattley T, Brent PA, Mills E and Leonard BJ: In vitro
analysis of the herbal compound Essiac. Anticancer Res.
27:3875–3882. 2007.PubMed/NCBI
|
|
11
|
Song J, Li N, Xia Y, Gao Z, Zou SF, Yan
YH, Li SH, Wang Y, Meng YK, Yang JX, et al: Arctigenin confers
neuroprotection against mechanical trauma injury in human
neuroblastoma SH-SY5Y cells by regulating miRNA-16 and miRNA-199a
expression to alleviate inflammation. J Mol Neurosci. 60:115–129.
2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Moy KA, Yuan JM, Chung FL, Wang XL, Van
Den Berg D, Wang R, Gao YT and Yu MC: Isothiocyanates, glutathione
S-transferase M1 and T1 polymorphisms and gastric cancer risk: A
prospective study of men in Shanghai, China. Int J Cancer.
125:2652–2659. 2009.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Solanky KS, Bailey NJ, Beckwith-Hall BM,
Bingham S, Davis A, Holmes E, Nicholson JK and Cassidy A: Biofluid
1H NMR-based metabonomic techniques in nutrition research -
metabolic effects of dietary isoflavones in humans. J Nutr Biochem.
16:236–244. 2005.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lin H, He YH, Xu R and Zou W: Effect of
Flor-Essence on serum levels of IL-6, IL-12, TNF-α and NK cells in
exercise rats. Sheng Li Xue Bao. 67:618–622. 2015.PubMed/NCBI(In Chinese).
|
|
15
|
Zhu Q, Chen J, Li Q, Wang T and Li H:
Antitumor activity of polysaccharide from Laminaria japonica on
mice bearing H22 liver cancer. Int J Biol Macromol. 92:156–158.
2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Mei C, Zhou S, Zhu L, Ming J, Zeng F and
Xu R: Antitumor effects of Laminaria extract fucoxanthin on lung
cancer. Mar Drugs. 15(E39)2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Lu Z, Chang L, Du Q, Huang Y, Zhang X, Wu
X, Zhang J, Li R, Zhang Z, Zhang W, et al: Arctigenin induces an
activation response in porcine alveolar macrophage through
TLR6-NOX2-MAPKs signaling pathway. Front Pharmacol.
9(475)2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Sivoňová MK, Kaplán P, Tatarková Z,
Lichardusová L, Dušenka R and Jurečeková J: Androgen receptor and
soy isoflavones in prostate cancer. Mol Clin Oncol. 10:191–204.
2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Tamayo C, Richardson MA, Diamond S and
Skoda I: The chemistry and biological activity of herbs used in
Flor-Essence herbal tonic and Essiac. Phytother Res. 14:1–14.
2000.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Cheung S, Lim KT and Tai J: Antioxidant
and anti-inflammatory properties of ESSIAC and Flor-Essence. Oncol
Rep. 14:1345–1350. 2005.PubMed/NCBI
|
|
21
|
Qu C, Ma J, Zhang Y, Han C, Huang L, Shen
L, Li H, Wang X, Liu J and Zou W: Estrogen receptor variant ER-α36
promotes tamoxifen agonist activity in glioblastoma cells. Cancer
Sci. 110:221–234. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Qu C, Sun J, Liu Y, Wang X, Wang L, Han C,
Chen Q, Guan T, Li H, Zhang Y, et al: Caveolin-1 facilitated KCNA5
expression, promoting breast cancer viability. Oncol Lett.
16:4829–4838. 2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Strzalka W and Ziemienowicz A:
Proliferating cell nuclear antigen (PCNA): A key factor in DNA
replication and cell cycle regulation. Ann Bot. 107:1127–1140.
2011.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zhao H, Lo YH, Ma L, Waltz SE, Gray JK,
Hung MC and Wang SC: Targeting tyrosine phosphorylation of PCNA
inhibits prostate cancer growth. Mol Cancer Ther. 10:29–36.
2011.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Göbel TW, Kaspers B and Stangassinger M:
NK and T cells constitute two major, functionally distinct
intestinal epithelial lymphocyte subsets in the chicken. Int
Immunol. 13:757–762. 2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Al Omar SY, Marshall E, Middleton D and
Christmas SE: Increased killer immunoglobulin-like receptor
expression and functional defects in natural killer cells in lung
cancer. Immunology. 133:94–104. 2011.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Lee WW, Lee N, Fujii H and Kang I: Active
hexose correlated compound promotes T helper (Th) 17 and 1 cell
responses via inducing IL-1β production from monocytes in humans.
Cell Immunol. 275:19–23. 2012.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chen L, Zhu SL and Cao M: Protective
effect of orgnoselenium from Se-enriched lactobacillus on lipid
peroxidation reaction and NK cell activity in spleen of liver
injury mice. Shi Yan Sheng Wu Xue Bao. 38:1–6. 2005.PubMed/NCBI(In Chinese).
|
|
29
|
Nguyen KB, Salazar-Mather TP, Dalod MY,
Van Deusen JB, Wei XQ, Liew FY, Caligiuri MA, Durbin JE and Biron
CA: Coordinated and distinct roles for IFN-alpha beta, IL-12, and
IL-15 regulation of NK cell responses to viral infection. J
Immunol. 169:4279–4287. 2002.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Ritz BW, Aktan I, Nogusa S and Gardner EM:
Energy restriction impairs natural killer cell function and
increases the severity of influenza infection in young adult male
C57BL/6 mice. J Nutr. 138:2269–2275. 2008.PubMed/NCBI View Article : Google Scholar
|
|
31
|
García-Sastre A and Biron CA: Type 1
interferons and the virus-host relationship: A lesson in détente.
Science. 312:879–882. 2006.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Trinchieri G: Interleukin-12 and the
regulation of innate resistance and adaptive immunity. Nat Rev
Immunol. 3:133–146. 2003.PubMed/NCBI View
Article : Google Scholar
|
|
33
|
Zhao X, Wei Y and Kariya Y: Induction of
antitumor immune response by NK-cell-sensitive target cells
transfected by B7-1 gene. Zhonghua Yi Xue Yi Chuan Xue Za Zhi.
15:210–213. 1998.PubMed/NCBI(In Chinese).
|
|
34
|
Fuchs A, Vermi W, Lee JS, Lonardi S,
Gilfillan S, Newberry RD, Cella M and Colonna M: Intraepithelial
type 1 innate lymphoid cells are a unique subset of IL-12- and
IL-15-responsive IFN-γ-producing cells. Immunity. 38:769–781.
2013.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Long H, Xie R, Xiang T, Zhao Z, Lin S,
Liang Z, Chen Z and Zhu B: Autocrine CCL5 signaling promotes
invasion and migration of CD133+ ovarian cancer
stem-like cells via NF-κB-mediated MMP-9 upregulation. Stem Cells.
30:2309–2319. 2012.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Elinav E, Nowarski R, Thaiss CA, Hu B, Jin
C and Flavell RA: Inflammation-induced cancer: Crosstalk between
tumours, immune cells and microorganisms. Nat Rev Cancer.
13:759–771. 2013.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Grivennikov SI and Karin M: Inflammation
and oncogenesis: A vicious connection. Curr Opin Genet Dev.
20:65–71. 2010.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Naugler WE and Karin M: NF-kappaB and
cancer-identifying targets and mechanisms. Curr Opin Genet Dev.
18:19–26. 2008.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Vandermeulen E, Ruttens D, Verleden SE,
Vos R, Van Raemdonck DE, Kastelijn EA, Wauters E, Lambrechts D,
Nawrot TS, Cox B, et al: Genetic variation in caveolin-1 affects
survival after lung transplantation. Transplantation. 98:354–359.
2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Hu G, Ye RD, Dinauer MC, Malik AB and
Minshall RD: Neutrophil caveolin-1 expression contributes to
mechanism of lung inflammation and injury. Am J Physiol Lung Cell
Mol Physiol. 294:L178–L186. 2008.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Wang XX, Wu Z, Huang HF, Han C, Zou W and
Liu J: Caveolin-1, through its ability to negatively regulate TLR4,
is a crucial determinant of MAPK activation in LPS-challenged
mammary epithelial cells. Asian Pac J Cancer Prev. 14:2295–2299.
2013.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Figgins B, Primeaux B, Shank BR, Chen SE,
Weber DM and Lu H: Cyclophosphamide desensitization in patients
with severe hypersensitivity reactions to bendamustine. J Oncol
Pharm Pract: Aug 21, 2019 (Epub ahead of print). doi:
10.1177/1078155219867127.
|
|
43
|
Liu M, Hales BF and Robaire B: Effects of
four chemotherapeutic agents, bleomycin, etoposide, cisplatin, and
cyclophosphamide, on DNA damage and telomeres in a mouse
spermatogonial cell line. Biol Reprod. 90(72)2014.PubMed/NCBI View Article : Google Scholar
|