Introduction
Estrogen elicits many different responses in female
reproductive tissues, including the ovary and uterus, as well as in
extra-reproductive tissues, such as the brain, adipose tissue and
the liver (1,2). It is well known that numerous, but not
all, of the concerted actions of estrogen are mediated through
binding to two nuclear estrogen receptors (ERs), ERα and ERβ
(1), both of which belong to a family
of hormone-activated transcription factors and share common
structural and functional domains (3). Although there is only ~60% homology in
the ligand binding domain between ERα and ERβ, the receptors
exhibit a similar binding affinity to endogenous 17β-estradiol (E2)
(3). While ERα and ERβ can homo- or
heterodimerize in vivo, they are not functionally
equivalent, and in vitro experiments show that ERβ functions
as a transcriptional inhibitor of ERα when ERα and ERβ are
co-expressed (4). Although ERα and
ERβ are often co-expressed in estrogen target cells under
physiological conditions and although they can act together to
regulate gene transcription (1,5), the
cellular localization and abundance of the two receptors show
distinct patterns in human endometrial epithelial and stromal cells
(5). For example, ERα represents the
most prominent receptor type in the endometrial epithelial and
stromal cells during the menstrual cycle, whereas ERβ is found
predominantly in the endometrial stromal cells in the late
secretory phase (5). Direct evidence
for essential roles of the estrogen signaling pathway in uterine
physiology and disease is provided by different ER knockout and
mutation studies in mice (1,3) and rats (6). It has been reported that female
ERαβ-/- and ERα-/- mice and female
ERα-/- rats are insensitive to E2 stimulation and they
exhibit uterine hypoplasia and infertility, which is in contrast to
loss of ERβ (ERβ-/-) in female mice that leads to
subfertility. Moreover, changes in ERα expression levels and the
ERα: ERβ ratio are considered to be the main factors behind several
gynecological disorders, including impaired fertility and
endometrial hyperplasia and carcinoma (5,7).
Polycystic ovary syndrome (PCOS), like numerous
complex diseases, has a multifaceted etiology and pathophysiology,
and it is associated with hormonal and metabolic impairments,
ovarian dysfunction, menstrual irregularity and infertility
(8,9).
Due to chronic anovulation, patients with PCOS experience sustained
and persistent estrogen stimulation but minimal or completely
absent progesterone stimulation (10,11), and
patients with PCOS with endometrial hyperplasia have a four-fold
greater risk of developing endometrial carcinoma than non-PCOS
controls (12). Preclinical and
clinical studies have provided evidence that the endometrium from
PCOS-like rodents and patients with PCOS displays morphologically
normal, but structurally and biochemically abnormal responses to
hormone stimulation (10,13-17).
Although few PCOS endometrial samples have been analyzed, there is
some controversial evidence that levels of endometrial ERα and ERβ
mRNA and/or protein are higher in patients with PCOS compared with
phase-matched non-PCOS controls, regardless of whether endometrial
hyperplasia is present or not (18,19).
Moreover, studies have previously shown that ERα and ERβ mRNAs are
increased in PCOS-like rodent uteri (20,21). These
preclinical and clinical findings suggest that altered expression
and function of both ERs contribute to endometrial dysfunction in
patients with PCOS.
Glycolysis is an energy-producing mechanism that is
regulated by different levels and activities of enzymes, such as
hexokinase (HK), phosphofructokinase (PFK) and pyruvate kinase (PK)
(22). E2 is a master regulator of
endometrial cell proliferation (23)
and has been shown to increase HK1/2 and PK isozyme M2 (PKM2)
activities, as well as glycolytic flux in the rat uterus in
vivo (24-26)
and in human endometrial stromal cells in vitro (27). Importantly, functional experiments
demonstrated that de novo synthesis of E2 in stromal cells
facilitates the decidualization process in the mouse uterus, which
is a prerequisite for successful implantation and establishment of
pregnancy (28). It was reported that
the regulation and localization of uterine ERα but not ERβ mRNA was
associated with the onset of early implantation in mice (29), and the acceleration of glycolysis is
required for endometrial decidualization in humans and mice
(30,31). Moreover, suppression of HK2 levels
inhibited the proliferation and differentiation of human
endometrial stromal cells in vitro (32). Taken together, these in vivo
and in vitro studies suggest that it is possible that
uterine E2-regulated glycolysis via ERα activation contributes to
successful implantation and the establishment of pregnancy.
However, whether uterine glycolysis is regulated by E2 in a
specific ERα- and/or ERβ-dependent manner remains unclear.
In this study, the localization and regulation of
ERα and ERβ in human and mouse endometria was assessed and
ER-specific knockout mice that lack ERα and/or ERβ
(ERαβ-/-, ERα-/- and ERβ-/-) were
used to determine whether the selective contribution of ERα and ERβ
results in the differential expression of key glycolytic enzymes in
the mouse uterus.
Materials and methods
Animals and tissue collection
Two distinct experiments were performed with the
animals. In the first experiment, intact prepubertal female
C57BL/6J mice (Taconic Biosciences) at 26 days of age with a body
weight (BW) of 13-15 g were used to avoid the complexity of ovarian
functions associated with estrous cycles and endogenous surges of
gonadotropins (33,34). Animals (n=5/group) were given a
subcutaneous injection of 0.5 µg E2/g BW (in 100 µl sesame oil) or
vehicle (100 µl sesame oil; Sigma-Aldrich; Merck KGaA) alone for 4
days (35). In the second experiment,
homozygous mutant female mice lacking the genes for ERαβ, ERα and
ERβ were utilized (age, 60-65 days; weight, 20-25 g); the
generation of female ERαβ-/-, ERα-/- and
ERβ-/- mice has been previously described (36-38).
Scanbur AB bred and provided the different ER knockout mice;
animals were inbred on a C57BL/6J background and littermate
controls were used in all groups. All adult ER knockout mice were
compared to isogenic wild-type (WT) age/weight-matched littermates
at the same diestrus stage of the estrous cycle (n=4/group)
(39).
Under anesthesia, the uteri were removed and
stripped of fat and connective tissue. One side of the uterus in
each animal was fixed in 4% formaldehyde neutral-buffered solution
for 24 h at 4˚C and then embedded in paraffin for
immunohistochemical analysis. The other side was immediately frozen
in liquid nitrogen and stored at -70˚C for subsequent western blot
analysis. All mice were housed in polycarbonate plastic cages with
free access to food pellets and water at the infection-free animal
facility of University of Gothenburg under a controlled temperature
of 22±2˚C at 55-65% humidity with a 12-h light/dark cycles.
Human endometrial tissue
collection
Endometrial tissues were obtained from
reproductive-aged women (range, 25-45 years) during the
proliferative phase of the menstrual cycle who were undergoing
routine gynecological investigation. Tissues were collected in
Obstetrics and Gynecology Hospital of Fudan University between
March and October 2014. None of selected patients had been exposed
to any hormonal or steroidal therapies within three months prior to
tissue sampling. Each endometrial sample was diagnosed and staged
by routine pathology analysis using standard histological criteria
(40). All tissues were fixed in 10%
neutral formalin solution for 24 h at 4˚C and embedded in paraffin
for immunohistochemical analysis.
The animal study was approved by the Animal Care and
Use Committee of the local Ethics Committee of the University of
Gothenburg (Sweden) and all animal experiments and care procedures
were performed in compliance with the institutional guidelines for
the care and use of animals in research (170-2008 and 236-2012).
The human study protocol conformed to the principles outlined in
the Declaration of Helsinki under approval from the institutional
Ethics Review Committee of the Obstetrics and Gynecology Hospital
of Fudan University (approval no. OGHFU 2013-23). Appropriate
written informed consent was obtained from all patients.
Total protein extraction and western
blot analysis
Protein lysates were prepared from mouse uterine
tissues using ice-cold RIPA buffer (Sigma-Aldrich; Merck KGaA)
supplemented with cOmplete Mini protease inhibitor cocktail tablets
(Roche Diagnostics) and PhosSTOP phosphatase inhibitor cocktail
tablets (Roche Diagnostics). Protein concentration determination
and a western blot analysis protocol were previously described
(13,41,42). After
determining the total protein concentration by Bradford protein
assay (Thermo Fisher Scientific, Inc.), 30-µg protein was resolved
on 4-12% Bis-Tris gradient gels (Novex; Thermo Fisher Scientific,
Inc.) and transferred to PVDF membranes. The membranes were blocked
with 0.01 M Tris-buffered saline supplemented with 0.1% (v/v)
Triton X-100 (TBST) containing 5% non-fat dry milk for 1 h at room
temperature (RT) and then probed with different primary antibodies
in the blocking buffer overnight at 4˚C. The primary antibody
details are as follows: HK1 (1:100; cat. no. 2024), HK2 (1:100;
cat. no. 2867), PFK (1:100; cat. no. 8164), GAPDH (1:200; cat. no.
5174), PKM2 (1:100; cat. no. 4053), pyruvate dehydrogenase (PDH;
1:100; cat. no. 3205) (all from Cell Signaling Technology, Inc.,)
ERα (1:300; cat. no. 6F11; Novocastra Laboratories Ltd.; Leica
Biosystems), ERβ (1:1,000; cat. no. 06-629; Upstate Biotechnology,
Inc.), progesterone receptor (PR; 1:100; cat. no. sc-538),
proliferating cell nuclear antigen (PCNA; 1:100; cat. no. sc-25280)
(both from Santa Cruz Biotechnologies, Inc.), total caspase-3
(1:500; cat. no. C92-605; BD Biosciences) and β-actin (1:500; cat.
no. A1978; Sigma-Aldrich; Merck KGaA). On day 2, the membranes were
washed with TBST followed by either anti-rabbit IgG horseradish
peroxidase (HRP)-conjugated goat (1:1,000; cat. no. A0545) or
anti-mouse IgG HRP-conjugated goat (1:1,000; cat. no. A2304)
secondary antibody (both from Sigma-Aldrich; Merck KGaA) for 1 h at
RT. Chemiluminescence signals were detected using SuperSignal West
Dura substrate following the manufacturer's instructions (Thermo
Fisher Scientific, Inc.). Band densitometry and quantification was
performed using Image Laboratory (v5.0; Bio-Rad Laboratories, Inc.)
and the protein band densities were normalized to β-actin. To
reprobe the membrane with another antibody, the blot was washed
with TBST 3x for 10 min at RT and incubated with stripping buffer
(65 mM Tris-HCl, 2% SDS and 100 mM β-mercaptoethanol, pH 6.8) at RT
for 15 min. Then the steps regarding the washing, blocking and
probing of the membrane were repeated.
Immunohistochemical analyses and
microscopy
Immunohistochemistry and dual-immunofluorescence
were performed according to previously described methods (20,42,43). Human
endometria and mouse uterine and ovarian tissues were fixed in 4%
formaldehyde neutral-buffered solution for 24 h at 4˚C,
paraffin-embedded and 5 µm sections were obtained. Two sections per
sample were stained using standard hematoxylin and eosin methods
(13). After deparaffinization
(xylene, 10 min at RT) and rehydration (100, 90 and 70% ethanol,
each 10 min at RT), the sections were immersed in epitope retrieval
buffer (10 mM sodium citrate buffer, pH 6.0) and heated in a 700 W
microwave for 15 min. Sections were subsequently rinsed twice with
deionized H2O and once with TBST, each 5 min at RT.
Endogenous peroxidase was removed and non-specific binding was
blocked by incubation with 3% H2O2 for 10 min
at RT and then with 10% normal goat serum for 1 h at RT. After
incubation with primary antibody overnight at 4˚C in a humidified
chamber, same sections were incubated with secondary antibodies (30
min; RT) and stain from the avidin-biotinylated-peroxidase ABC kit
according to the manufacturer's instructions (Vector Laboratories,
Inc.; Maravai LifeSciences) followed by a 5-min treatment with
3,3'-diaminobenzidine (SK-4100; Vector Laboratories, Inc.; Maravai
LifeSciences) at RT. All sections were incubated with DAB for the
same length of time so that comparisons could be made between
individual samples and all slides were stained in a single run to
eliminate inter-experiment variations in staining intensity.
Digital images of stained sections were obtained with a Nikon
E-1000 microscope (Nikon Corporation) using bright-field optics
(magnification, x2, x10 and x40) and photomicrographed using Easy
Image 1 (Bergström Instrument AB). Primary antibodies for
immunohistochemistry included: ERα (1:50; cat. no. MC-20; Santa
Cruz Biotechnologies Inc.), ERβ (1:300; cat. no. 06-629; Upstate
Biotechnology, Inc.), ERβ1 (1:100; cat. no. PPG5/10), ERβ2 (1:100;
cat. no. 57/3) (both from AbD Serotec; Bio-Rad Laboratories, Inc.),
cytokeratin 8 (1:200; cat. no. C5301; Sigma-Aldrich; Merck KGaA)
and Ki-67 (1:100; cat. no. 9027; Cell Signaling Technology,
Inc.).
Human and mouse endometrial tissue sections were
blocked in PBS containing 1% BSA and 3% fat-free milk for 1 h at
room temperature. Sections were incubated with the anti-ERα (1:50;
cat. no. MC-20; Santa Cruz Biotechnologies for human tissues; and
1:100; cat. no. 6F11; Novocastra Laboratories for mouse tissues),
anti-ERβ1 (1:100; cat. no. PPG5/10; AbD Serotec for human tissues),
anti-ERβ2 (1:100; cat. no. 57/3; AbD Serotec for human tissues) or
anti-ERβ (1:300; cat. no. 06-629; Upstate Biotechnology for mouse
tissues) antibody in PBS supplemented with 0.1% (v/v) Triton X-100
(PBST) containing 1% BSA and 3% fat-free milk overnight at 4˚C.
After washing with PBST three times for 5 min each, sections were
incubated with Alexa Fluor 594-conjugated goat polyclonal
anti-rabbit IgG (1:250; cat. no. A11037), Alexa Fluor
488-conjugated goat polyclonal anti-rabbit IgG (1:250; cat. no.
A11008) or Alexa Fluor 488-conjugated goat polyclonal anti-mouse
IgG (1:250; cat. no. A11039) (all from Invitrogen; Thermo Fisher
Scientific, Inc.) for 1 h at RT. After the sections were washed
with PBST, they were examined under an Axiovert 200 confocal
microscope (magnification, x20 and x60; Zeiss GmbH) equipped with a
laser-scanning confocal imaging LSM 510 META system (Carl Zeiss AG)
and were photomicrographed. Background settings were adjusted from
the examination of negative control specimen; different controls
for non-specific staining have been described previously (43).
Statistical analysis
For all experiments, n represents the numbers of
individual animals. Data are presented as the mean ± SEM
(n=4/group). Statistical analyses were performed using the SPSS
version 24.0 (IMB Corp.). The normal distribution of the data was
tested by Shapiro-Wilk test. Differences between groups were
analyzed by one-way ANOVA followed by Bonferroni's post hoc test
for normally distributed data or by the Kruskal-Wallis test
followed by Mann-Whitney U test for skewed data. P<0.05 was
considered to indicate a statistically significant difference.
Results and Discussion
The endometrium is composed of a lining of surface
epithelium and associated glands, and a stroma composed of
connective tissue (5). During each
reproductive cycle, the endometrial epithelial and stromal cells
display distinct and well-defined patterns of functional
differentiation under the cyclic influence of estrogen and
progesterone (23). Increasing
evidence suggests that the differential effects of estrogen on
endometrial cells likely depend on the total amount of cellular ERs
and/or the ratio of ERα to ERβ (5,7). In this
study, it was found that the level of ERα was higher than ERβ in
human (Fig. 1) and mouse (Fig. 2A and B)
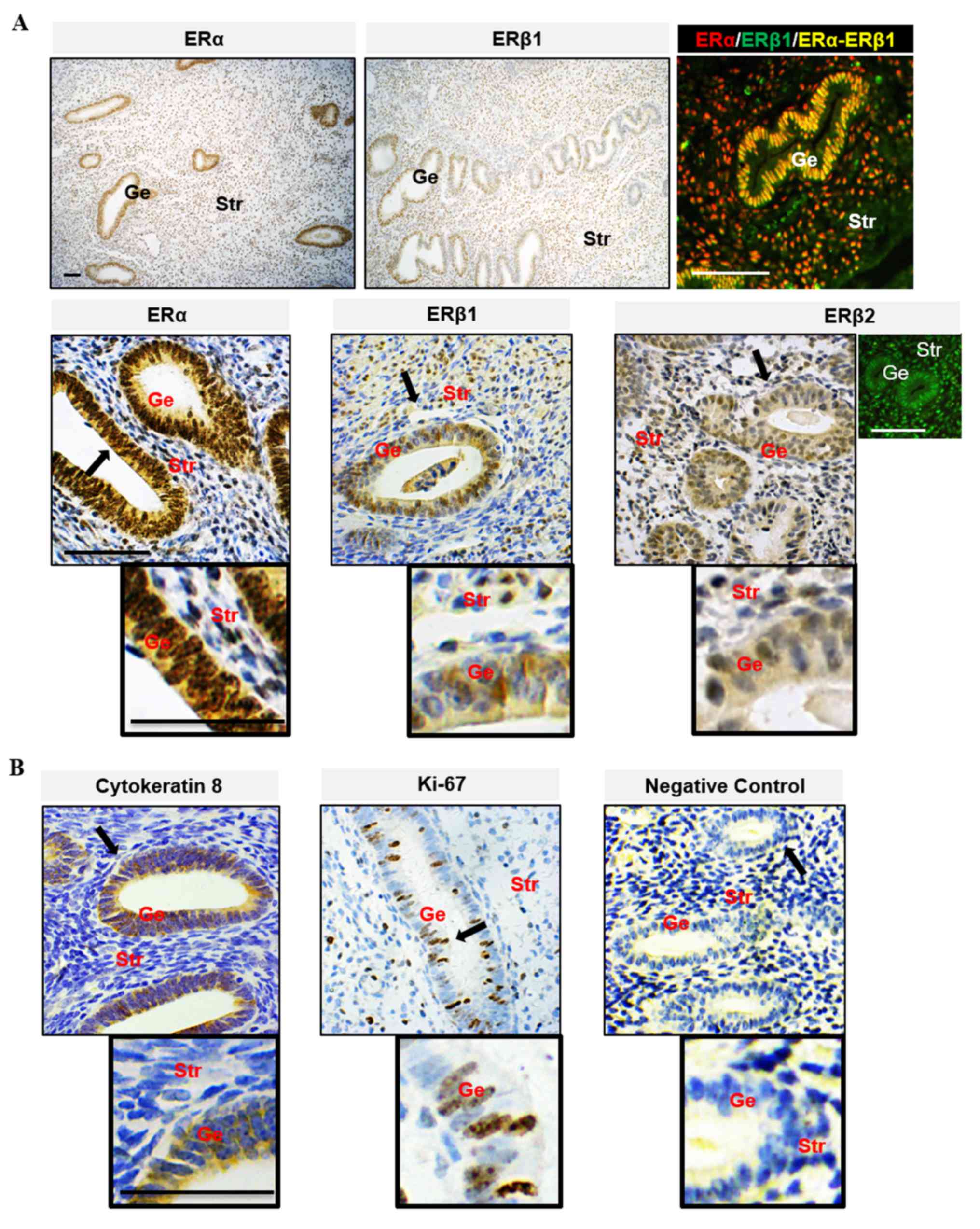
endometrial epithelial and stromal cells. This suggested that ERα
was the predominant ER expressed in the uterus. Immunohistochemical
analysis of tissues from women during the estrogen-dominant
proliferative phase showed strong positive nuclear staining for ERα
in epithelial and stromal cells but weak to moderate positive
nuclear staining for ERβ1 and ERβ2, and ERα immunoreactivity was
more abundant in the nucleus than in the cytoplasm in epithelial
cells. ERα and ERβ1 were heterogeneously co-localized in the
nucleus of epithelial and stromal cells. Compared with ERβ1, the
ERβ2 immunoreactivity was evenly detected in the nuclei and
cytoplasm of epithelial and stromal cells (Fig. 1A). These observations of endometrial
cellular ERα and ERβ localization were broadly in agreement with
previous human studies (5). As shown
in Fig. 2B, ERα immunoreactivity was
detected in the nuclei of epithelial and stromal cells, whereas ERβ
immunoreactivity was detected mainly in the nuclei of stromal cells
only. It was found that prepubescent mice treated chronically with
E2 had decreased ERα and ERβ protein expression and increased PR
isoform protein expression compared with the vehicle-treated
controls (Fig. 2C). In the same
experimental mouse uterus, increased PCNA, a cellular marker for
proliferation, was associated with decreased activated caspase-3, a
marker for cell apoptosis (Fig. 2C).
These finding confirmed that E2 contributed to normal endometrial
growth through the direct regulation of uterine ERα and ERβ in
vivo.
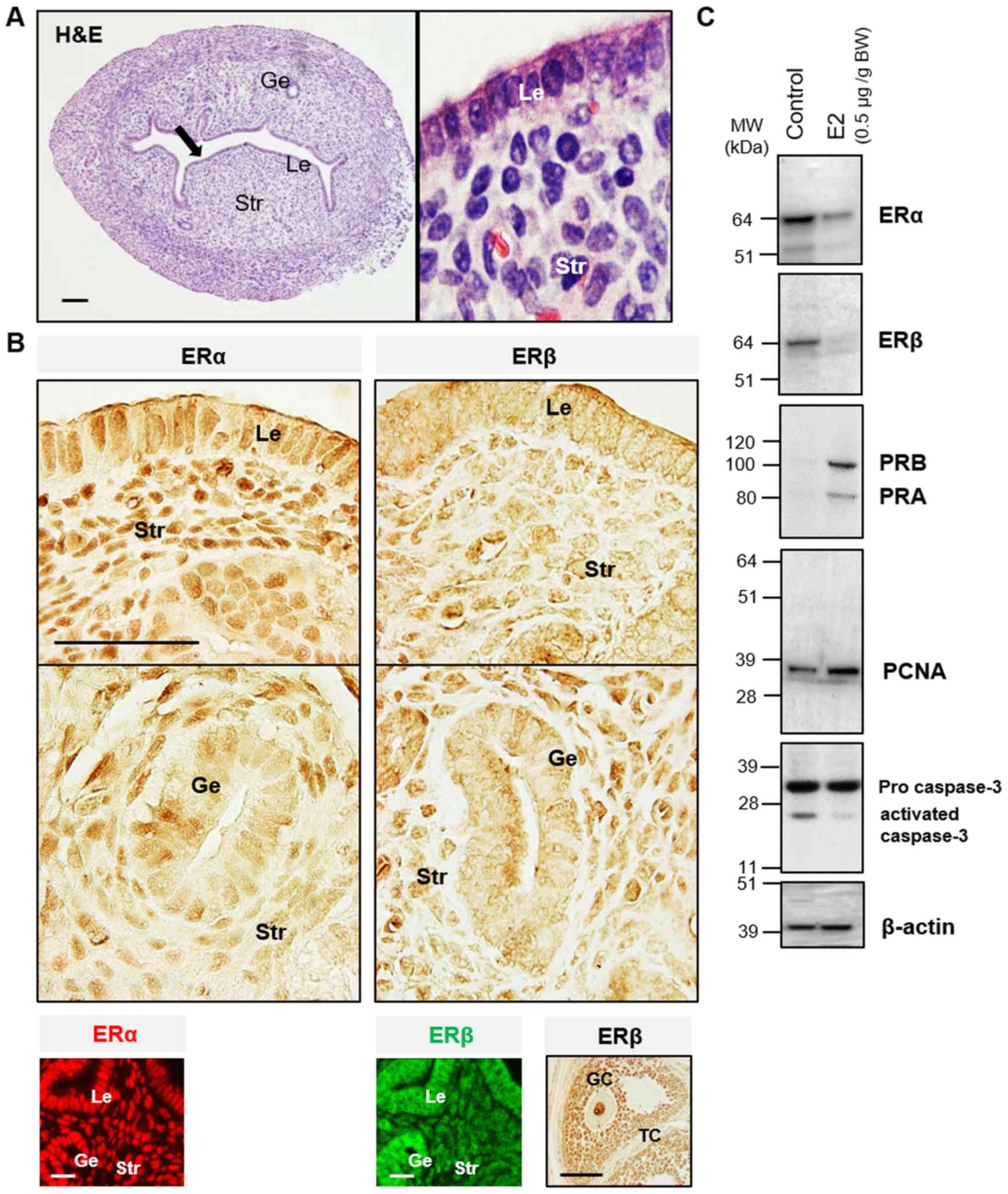 | Figure 2Localization of ER subtypes in the
mouse uterus. (A) Uterine tissue sections stained with hematoxylin
and eosin staining. (B) Immunohistological localization of ERα and
ERβ in the mouse uterus. A mouse ovarian tissue section was used as
the positive control for the anti-ERβ antibody specificity.
Immunohistochemistry was performed using 3,3'-diaminobenzidine
(brown). Immunofluorescence detection shows ERα (red) and ERβ
(green). Black arrows indicate areas shown at higher magnification;
scale bar, 100 µm. (C) Western blot analysis of ER subtypes,
proliferation and apoptosis markers in the prepubescent mouse
uterus. E2, 17β-estradiol; BW, body weight; MW, molecular weight;
Le, luminal epithelial cells; Ge, glandular epithelial cells; Str,
stromal cells; GC, granulosa cells; TC, thecal cells; ER, estrogen
receptor; PR, progesterone receptor; proliferating cell nuclear
antigen. |
There is increasing clinical and experimental
evidence suggesting that aberrant regulation of ERα and ERβ
expression is involved in the development and progression of
several reproductive and metabolic diseases (1,2). For
example, one female patient with a homozygous ERα mutation and
female mice and rats lacking ERα display similar polycystic ovary
phenotypes and infertility (6,44,45) as observed in patients with PCOS.
Moreover, an ERβ polymorphism (+1730 G/A) has been implicated in
susceptibility to the development of PCOS in humans (46). While the exact mechanisms of the
pathogenesis of PCOS remains unknown, increasing evidence suggests
that PCOS is a clinically heterogeneous and multifactorial disorder
(8,9).
Taken together, these findings indicate that multiple cellular and
molecular signaling pathways are likely to be involved in its
pathogenesis.
Glycolysis is the splitting apart of a glucose
molecule in the cytosol by a sequence of enzymatic reactions, and
its efficient operation requires adequate glucose uptake mediated
by a number of glucose transporters (GLUTs) (47). Among the GLUTs, GLUT1 has been
identified as the most prominent in endometrial tissues in
vivo (48). Thus, it is thought
that GLUT1 is responsible for the basal level of glucose uptake
needed for normal glucose utilization in the uterus. It has been
reported that E2 decreases glucose uptake in association with
decreased GLUT1 expression in human and mouse endometrial stromal
cells in vitro (49). Although
there is no cyclical fluctuation of insulin-sensitive GLUT4
expression observed in human endometrium (50), it was previously shown that GLUT4 mRNA
and protein expression are decreased in patients with PCOS compared
with non-PCOS controls (50-52)
and a similar observation has been made in the PCOS-like rat uterus
(13,51). An analysis of gene expression in
endometrial tissues found significantly reduced levels of key
glycolytic genes in patients with PCOS compared with non-PCOS
controls (53). Reproductive
dysfunction and infertility are common in patients with PCOS
(11,54), who often display E2-mediated
endometrial hyperproliferation (55).
Further studies have demonstrated that several proteins involved in
cytosolic glycolysis, such as PKM2, are impaired in the endometrium
of patients with PCOS and in PCOS-like animals with an endometrial
hypoplasia phenotype (13,56). These findings support the notion that
dysregulation of E2-mediated glycolysis is, at least partially,
involved in the endometrial dysfunction in patients with PCOS with
endometrial hyperplasia.
Changes in the glucose metabolism are a fundamental
part of many biological processes (22). However, at present there is limited
knowledge as to whether the estrogenic regulation of the uterine
glucose metabolism depends on different ER subtypes. In this study,
the expression pattern of uterine glycolytic enzymes in
ERαβ-/-, ERα-/- and ERβ-/- mice
was compared with WT controls (Fig.
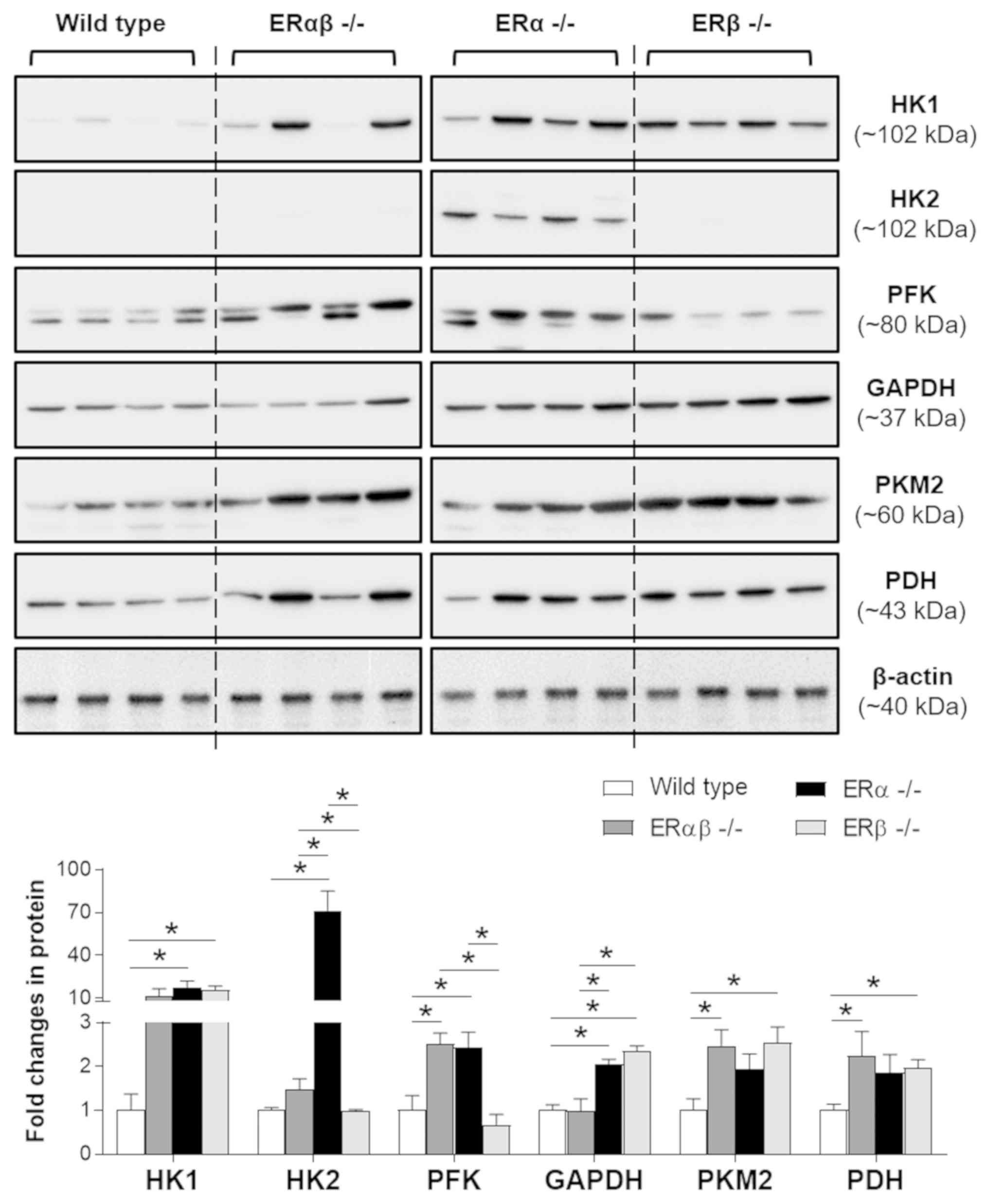
3). Using western blot analysis, it was found that HK1 and
GAPDH expression was significantly increased in ERα-/-
and ERβ-/- mice compared with the WT controls. Moreover,
significantly increased HK2 and PFK expression was observed in
ERα-/- mice compared with the WT controls, whereas
significantly increased PKM2 and PDH expression was observed for
ERβ-/- mice compared with the WT controls. As indicated
in the expression pattern of glycolytic enzymes, disruption of ERα
and ERβ (ERαβ-/-) resulted in significantly increased
PFK, PKM2 and PDH expression compared with the WT controls. This
suggested that although ERα is the predominant ER expressed in the
uterus, ERβ may partially compensate for the loss of ERα by
increasing the expression of certain glycolytic enzymes in the
uterus. Furthermore, the significantly increased uterine PDH
expression in ERβ-/- but not ERα-/- animals
compared with the WT controls suggested that the cell's
mitochondria contained primarily ERβ and not ERα (5). It is noteworthy that the estrogen
responsiveness becomes more complex because human and rodent
reproductive tissues contain splice variants of ERα and ERβ
(5,35,57), and
the two subtypes form heterodimers with in vivo (3). It was previously shown that
ERα-/- mouse uteri, similar to ERαβ-/- mouse
uteri, remain to have one ERα splice form (35) and the levels of estrogen-regulated ERα
protein are positively associated with endometrial hyperplasia in
patients with PCOS (56). To better
understand the role of estrogen-regulated glycolysis in the
endometria of patients with PCOS, further studies are needed to
determine whether ERα splice variants are differentially regulated
by E2 using well-controlled endometrial tissue samples collected
from patients with PCOS with various phenotypes.
Furthermore, the western blot analysis demonstrated
that two distinct forms of PFK were present in the mouse uterus
(Fig. 3). PFK is synthesized as an
unstable inactive monomer, which associates rapidly to form
minimally active dimers essential for maintaining the tertiary
structure of the enzyme (58).
Several studies have shown that PFK has three isoforms (M, P and L)
and differentially expresses in various mammalian tissues in
vivo (59,60). It is hypothesized that the PFK
antibody used in this study was able to detect two different
isoforms of PFK; however, which PFK isoform is expressed in the
mouse uterus remains to be determined. As the varying ratio of PFK
isoforms may determine the glycolytic rate in a tissue-specific
manner (61), further work is needed
to determine which PFK isoforms contribute to the uterine
glycolytic rates in mice.
The role of aberrant glucose metabolism in the
development of hormone-related diseases has become a topic of great
interest. In addition to hyperandrogenism, numerous patients with
PCOS also exhibit core metabolic manifestations, including
peripheral insulin resistance (8,9). Of
interest, female ERα-/- but not ERβ-/- mice
develop obesity and insulin resistance (2). Hulchiy et al (62) have reported that endometrial ERα but
not ERβ mRNA is decreased in overweight/obese patients with PCOS
compared with controls, which is in contrast to the increased
endometrial ERα and ERβ mRNA and/or protein expression observed in
patients with hyperandrogenic PCOS (18,19). Thus,
it remains unclear how insulin resistance and hyperandrogenism
differentially affect ER subtype-mediated regulation of glycolysis
in the endometrium in patients with PCOS. Based on a growing number
of preclinical and clinical studies (13,18,41,42,63,64),
it is hypothesized that abnormal steroid hormone responsiveness,
such as hyperandrogenism, metabolic dysfunction, such as insulin
resistance, molecular aberrations in the endometrium, such as
glycolysis (Fig. 4), oxidative
stress, immune factors and inflammatory uterine environments are
all potential to be involved in the endometrial dysfunction
observed in patients with PCOS (42).
Further investigations are required to elucidate the crosstalk
between these possible mechanisms in the uterus under both
physiological and pathological conditions.
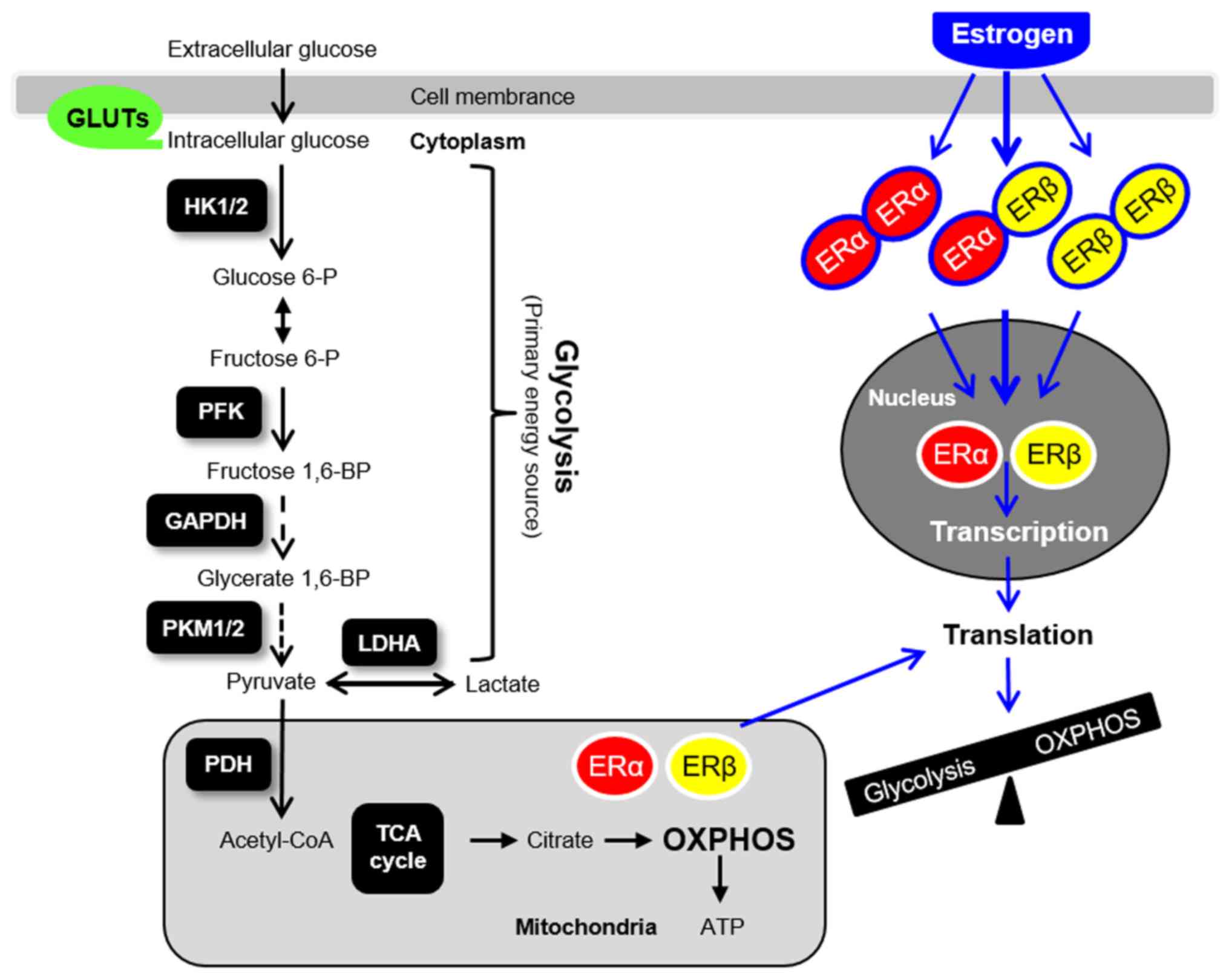 | Figure 4Glycolysis, mitochondria-mediated
energy metabolism and ER-mediated genomic actions in the uterus.
GLUT, glucose transporter; HK, hexokinase; PFK,
phosphofructokinase; PKM, pyruvate kinase isoform; LDHA, lactate
dehydrogenase A; PDH, pyruvate dehydrogenase; OXPHOS, oxidative
phosphorylation; TCA, tricarboxylic acid; ER, estrogen receptor; P,
phosphate; BP, bisphosphate. |
Acknowledgements
Not applicable.
Funding
This study was funded by grants from the Swedish
Medical Research Council (grant no. 10380), the Swedish state under
the ALF agreement between the Swedish government and the county
councils (grant no. ALFGBG-147791), the Jane and Dan Olsson's
Foundation, the Knut and Alice Wallenbergs Foundation, and the
Adlerbert Research Foundation to HB and LRS, as well as the
Guangzhou Medical University High-level University Construction
Talents Fund (grant no. B185006010046) and the National Natural
Science Foundation of China (grant no. 81774136).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LRS conceptualized the experiments, supervised the
study and provided key research direction. MH, YZ, EE, XL and LRS
performed the experiments. MH and LRS took responsibility for the
integrity of the data analysis. LRS wrote and revised the
manuscript. HB interpreted data and provided the critical comments
on the manuscript. LRS and HB provided scientific oversight and
guidance. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The animal study was approved by the Animal Care and
Use Committee of the local Ethics Committee of the University of
Gothenburg (Sweden) and all animal experiments and care procedures
were performed in compliance with the institutional guidelines for
the care and use of animals in research (170-2008 and 236-2012).
The human study protocol conformed to the principles outlined in
the Declaration of Helsinki under approval from the institutional
Ethics Review Committee of the Obstetrics and Gynecology Hospital
of Fudan University (approval no. OGHFU 2013-23). Appropriate
written informed consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hamilton KJ, Hewitt SC, Arao Y and Korach
KS: Estrogen hormone biology. Curr Top Dev Biol. 125:109–146.
2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Barros RP and Gustafsson JÅ: Estrogen
receptors and the metabolic network. Cell Metab. 14:289–299.
2011.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Hewitt SC, Winuthayanon W and Korach KS:
What's new in estrogen receptor action in the female reproductive
tract. J Mol Endocrinol. 56:R55–R71. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Matthews J, Wihlén B, Tujague M, Wan J,
Strom A and Gustafsson JA: Estrogen receptor (ER) beta modulates
ERalpha-mediated transcriptional activation by altering the
recruitment of c-Fos and c-Jun to estrogen-responsive promoters.
Mol Endocrinol. 20:534–543. 2006.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Hapangama DK, Kamal AM and Bulmer JN:
Estrogen receptor β: The guardian of the endometrium. Hum Reprod
Update. 21:174–193. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Rumi MA, Dhakal P, Kubota K, Chakraborty
D, Lei T, Larson MA, Wolfe MW, Roby KF, Vivian JL and Soares MJ:
Generation of Esr1-knockout rats using zinc finger
nuclease-mediated genome editing. Endocrinology. 155:1991–1999.
2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Rodriguez AC, Blanchard Z, Maurer KA and
Gertz J: Estrogen signaling in endometrial cancer: A key oncogenic
pathway with several open questions. Horm Cancer. 10:51–63.
2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Azziz R, Carmina E, Chen Z, Dunaif A,
Laven JS, Legro RS, Lizneva D, Natterson-Horowtiz B, Teede HJ and
Yildiz BO: Polycystic ovary syndrome. Nat Rev Dis Primers.
2(16057)2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Goodarzi MO, Dumesic DA, Chazenbalk G and
Azziz R: Polycystic ovary syndrome: Etiology, pathogenesis and
diagnosis. Nat Rev Endocrinol. 7:219–231. 2011.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Li X, Feng Y, Lin JF, Billig H and Shao R:
Endometrial progesterone resistance and PCOS. J Biomed Sci.
21(2)2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Li X and Shao R: PCOS and obesity: Insulin
resistance might be a common etiology for the development of type I
endometrial carcinoma. Am J Cancer Res. 4:73–79. 2014.PubMed/NCBI
|
|
12
|
Fearnley EJ, Marquart L, Spurdle AB,
Weinstein P and Webb PM: Australian Ovarian Cancer Study Group and
Australian National Endometrial Cancer Study Group: Polycystic
ovary syndrome increases the risk of endometrial cancer in women
aged less than 50 years: An Australian case-control study. Cancer
Causes Control. 21:2303–2308. 2010.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhang Y, Sun X, Sun X, Meng F, Hu M, Li X,
Li W, Wu XK, Brännström M, Shao R and Billig H: Molecular
characterization of insulin resistance and glycolytic metabolism in
the rat uterus. Sci Rep. 6(30679)2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Kuyucu Y, Çelik LS, Kendirlinan Ö, Tap Ö
and Mete UÖ: Investigation of the uterine structural changes in the
experimental model with polycystic ovary syndrome and effects of
vitamin D treatment: An ultrastructural and immunohistochemical
study. Rep Biol. 18:53–59. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Brosens I and Benagiano G: Menstrual
preconditioning for the prevention of major obstetrical syndromes
in polycystic ovary syndrome. Am J Obstet Gynecol. 213:488–493.
2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Leonhardt H, Gull B, Kishimoto K, Kataoka
M, Nilsson L, Janson PO, Stener-Victorin E and Hellström M: Uterine
morphology and peristalsis in women with polycystic ovary syndrome.
Acta Radiol. 53:1195–1201. 2012.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Villavicencio A, Bacallao K, Gabler F,
Fuentes A, Albornoz J, Casals A and Vega M: Deregulation of tissue
homeostasis in endometria from patients with polycystic ovarian
syndrome with and without endometrial hyperplasia. Gynecol Oncol.
104:290–295. 2007.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Piltonen TT: Polycystic ovary syndrome:
Endometrial markers. Best Pract Res Clin Obstet Gynaecol. 37:66–79.
2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Baracat MC, Serafini PC, Simões Rdos S,
Maciel GA, Soares JM Jr and Baracat EC: Systematic review of cell
adhesion molecules and estrogen receptor expression in the
endometrium of patients with polycystic ovary syndrome. Int J
Gynaecol Obstet. 129:1–4. 2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hu M, Zhang Y, Feng J, Xu X, Zhang J, Zhao
W, Guo X, Li J, Vestin E, Cui P, et al: Uterine progesterone
signaling is a target for metformin therapy in PCOS-like rats. J
Endocrinol. 237:123–137. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Li SY, Song Z, Song MJ, Qin JW, Zhao ML
and Yang ZM: Impaired receptivity and decidualization in
DHEA-induced PCOS mice. Sci Rep. 6(38134)2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Burns JS and Manda G: Metabolic pathways
of the warburg effect in health and disease: Perspectives of
choice, Chain or chance. Int J Mol Sci. 18pii.
(E2755)2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Gellersen B and Brosens JJ: Cyclic
decidualization of the human endometrium in reproductive health and
failure. Endocr Rev. 35:851–905. 2014.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Baquer NZ and McLean P: The effect of
oestradiol on the profile of constant and specific proportion
groups of enzymes in rat uterus. Biochem Biophys Res Commun.
48:729–734. 1972.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Baquer NZ, Sochor M, Kunjara S and McLean
P: Effect of oestradiol on the carbohydrate metabolism of immature
rat uterus: The role of fructose-2, 6-bis-phosphate and of
phosphoribosyl pyrophosphate. Biochem Mol Biol Int. 31:509–519.
1993.PubMed/NCBI
|
|
26
|
Reiss NA: Ontogeny and estrogen
responsiveness of creatine kinase and glycolytic enzymes in brain
and uterus of rat. Neurosci Lett. 84:197–202. 1988.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Singhal RL and Valadares JR: Estrogenic
regulation of uterine pyruvate kinase. Am J Physiol. 218:321–327.
1970.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Das A, Mantena SR, Kannan A, Evans DB,
Bagchi MK and Bagchi IC: De novo synthesis of estrogen in pregnant
uterus is critical for stromal decidualization and angiogenesis.
Proc Natl Acad Sci USA. 106:12542–12547. 2009.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Tan J, Paria BC, Dey SK and Das SK:
Differential uterine expression of estrogen and progesterone
receptors correlates with uterine preparation for implantation and
decidualization in the mouse. Endocrinology. 140:5310–5321.
1999.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Zuo RJ, Gu XW, Qi QR, Wang TS, Zhao XY,
Liu JL and Yang ZM: Warburg-like glycolysis and lactate shuttle in
mouse decidua during early pregnancy. J Biol Chem. 290:21280–21291.
2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kommagani R, Szwarc MM, Kovanci E, Gibbons
WE, Putluri N, Maity S, Creighton CJ, Sreekumar A, DeMayo FJ, Lydon
JP and O'Malley BW: Acceleration of the glycolytic flux by steroid
receptor coactivator-2 is essential for endometrial
decidualization. PLoS Genet. 9(e1003900)2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Lv H, Tong J, Yang J, Lv S, Li WP, Zhang C
and Chen ZJ: Dysregulated pseudogene HK2P1 may contribute to
preeclampsia as a competing endogenous RNA for hexokinase 2 by
impairing decidualization. Hypertension. 71:648–658.
2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Shao R, Markström E, Friberg PA, Johansson
M and Billig H: Expression of progesterone receptor (PR) A and B
isoforms in mouse granulosa cells: Stage-dependent PR-mediated
regulation of apoptosis and cell proliferation. Biol Reprod.
68:914–921. 2003.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Shao R, Zhang FP, Rung E, Palvimo JJ,
Huhtaniemi I and Billig H: Inhibition of small ubiquitin-related
modifier-1 expression by luteinizing hormone receptor stimulation
is linked to induction of progesterone receptor during ovulation in
mouse granulosa cells. Endocrinology. 145:384–392. 2004.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Shao R, Egecioglu E, Weijdegård B,
Kopchick JJ, Fernandez-Rodriguez J, Andersson N and Billig H:
Dynamic regulation of estrogen receptor-alpha isoform expression in
the mouse fallopian tube: Mechanistic insight into
estrogen-dependent production and secretion of insulin-like growth
factors. Am J Physiol Endocrinol Metab. 293:E1430–E1442.
2007.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Couse JF, Hewitt SC, Bunch DO, Sar M,
Walker VR, Davis BJ and Korach KS: Postnatal sex reversal of the
ovaries in mice lacking estrogen receptors alpha and beta. Science.
286:2328–2331. 1999.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Lubahn DB, Moyer JS, Golding TS, Couse JF,
Korach KS and Smithies O: Alteration of reproductive function but
not prenatal sexual development after insertional disruption of the
mouse estrogen receptor gene. Proc Natl Acad Sci USA.
90:11162–11166. 1993.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Krege JH, Hodgin JB, Couse JF, Enmark E,
Warner M, Mahler JF, Sar M, Korach KS, Gustafsson JA and Smithies
O: Generation and reproductive phenotypes of mice lacking estrogen
receptor beta. Proc Natl Acad Sci USA. 95:15677–15682.
1998.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Feng Y, Johansson J, Shao R, Mannerås L,
Fernandez-Rodriguez J, Billig H and Stener-Victorin E: Hypothalamic
neuroendocrine functions in rats with dihydrotestosterone-induced
polycystic ovary syndrome: Effects of low-frequency
electro-acupuncture. PLoS One. 4(e6638)2009.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Noyes RW, Hertig AT and Rock J: Dating the
endometrial biopsy. Am J Obstet Gynecol. 122:262–263.
1975.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zhang Y, Hu M, Meng F, Sun X, Xu H, Zhang
J, Cui P, Morina N, Li X, Li W, et al: Metformin ameliorates
uterine defects in a rat model of polycystic ovary syndrome.
EBioMedicine. 18:157–170. 2017.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Hu M, Zhang Y, Guo X, Jia W, Liu G, Zhang
J, Li J, Cui P, Sferruzzi-Perri AN, Han Y, et al: Hyperandrogenism
and insulin resistance induce gravid uterine defects in association
with mitochondrial dysfunction and aberrant reactive oxygen species
production. Am J Physiol Endocrinol Metab. 316:E794–E809.
2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Li X, Pishdari B, Cui P, Hu M, Yang HP,
Guo YR, Jiang HY, Feng Y, Billig H and Shao R: Regulation of
androgen receptor expression alters AMPK phosphorylation in the
endometrium: In vivo and in vitro studies in women with polycystic
ovary syndrome. Int J Bio Sci. 11:1376–1389. 2015.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Quaynor SD, Stradtman EW Jr, Kim HG, Shen
Y, Chorich LP, Schreihofer DA and Layman LC: Delayed puberty and
estrogen resistance in a woman with estrogen receptor alpha
variant. N Engl J Med. 369:164–171. 2013.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Hamilton KJ, Arao Y and Korach KS:
Estrogen hormone physiology: Reproductive findings from estrogen
receptor mutant mice. Reprod Biol. 14:3–8. 2014.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Kim JJ, Choi YM, Choung SH, Yoon SH, Lee
GH and Moon SY: Estrogen receptor beta gene +1730 G/A polymorphism
in women with polycystic ovary syndrome. Fertil Steril.
93:1942–1947. 2010.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Frolova AI and Moley KH: Glucose
transporters in the uterus: An analysis of tissue distribution and
proposed physiological roles. Reproduction. 142:211–220.
2011.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Frolova AI and Moley KH: Quantitative
analysis of glucose transporter mRNAs in endometrial stromal cells
reveals critical role of GLUT1 in uterine receptivity.
Endocrinology. 152:2123–2128. 2011.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Frolova A, Flessner L, Chi M, Kim ST,
Foyouzi-Yousefi N and Moley KH: Facilitative glucose transporter
type 1 is differentially regulated by progesterone and estrogen in
murine and human endometrial stromal cells. Endocrinology.
150:1512–1520. 2009.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Cui P, Li X, Wang X, Feng Y, Lin JF,
Billig H and Shao R: Lack of cyclical fluctuations of endometrial
GLUT4 expression in women with polycystic ovary syndrome: Evidence
for direct regulation of GLUT4 by steroid hormones. BBA Clin.
4:85–91. 2015.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Li X, Cui P, Jiang HY, Guo YR, Pishdari B,
Hu M, Feng Y, Billig H and Shao R: Reversing the reduced level of
endometrial GLUT4 expression in polycystic ovary syndrome: A
mechanistic study of metformin action. Am J Transl Res. 7:574–586.
2015.PubMed/NCBI
|
|
52
|
Orostica L, Astorga I, Plaza-Parrochia F,
Vera C, Garcia V, Carvajal R, Gabler F, Romero C and Vega M:
Proinflammatory environment and role of TNF-α in endometrial
function of obese women having polycystic ovarian syndrome. Int J
Obes (Lond). 40:1715–1722. 2016.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Kim JY, Song H, Kim H, Kang HJ, Jun JH,
Hong SR, Koong MK and Kim IS: Transcriptional profiling with a
pathway-oriented analysis identifies dysregulated molecular
phenotypes in the endometrium of patients with polycystic ovary
syndrome. J Clin Endocrinol Metab. 94:1416–1426. 2009.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Evans J, Salamonsen LA, Winship A,
Menkhorst E, Nie G, Gargett CE and Dimitriadis E: Fertile ground:
Human endometrial programming and lessons in health and disease.
Nat Rev Endocrinol. 12:654–667. 2016.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Hardiman P, Pillay OC and Atiomo W:
Polycystic ovary syndrome and endometrial carcinoma. Lancet.
361:1810–1812. 2003.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Wang T, Zhang J, Hu M, Zhang Y, Cui P, Li
X, Li J, Vestin E, Brännström M, Shao LR and Billig H: Differential
expression patterns of glycolytic enzymes and
mitochondria-dependent apoptosis in PCOS patients with endometrial
hyperplasia, an early hallmark of endometrial cancer, in vivo and
the impact of metformin in vitro. Int J Biol Sci. 15:714–725.
2019.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Couse JF, Curtis SW, Washburn TF, Lindzey
J, Golding TS, Lubahn DB, Smithies O and Korach KS: Analysis of
transcription and estrogen insensitivity in the female mouse after
targeted disruption of the estrogen receptor gene. Mol Endocrinol.
9:1441–1454. 1995.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Schoneberg T, Kloos M, Brüser A,
Kirchberger J and Sträter N: Structure and allosteric regulation of
eukaryotic 6-phosphofructokinases. Biol Chem. 394:977–993.
2013.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Eto K, Sakura H, Yasuda K, Hayakawa T,
Kawasaki E, Moriuchi R, Nagataki S, Yazaki Y and Kadowaki T:
Cloning of a complete protein-coding sequence of human
platelet-type phosphofructokinase isozyme from pancreatic islet.
Biochem Biophys Res Commun. 198:990–998. 1994.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Hannemann A, Jandrig B, Gaunitz F,
Eschrich K and Bigl M: Characterization of the human P-type
6-phosphofructo-1-kinase gene promoter in neural cell lines. Gene.
345:237–247. 2005.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Al Hasawi N, Alkandari MF and Luqmani YA:
Phosphofructokinase: A mediator of glycolytic flux in cancer
progression. Crit Rev Oncol Hematol. 92:312–321. 2014.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Hulchiy M, Nybacka Å, Sahlin L and
Hirschberg AL: Endometrial expression of estrogen receptors and the
androgen receptor in women with polycystic ovary syndrome: A
lifestyle intervention study. J Clin Endocrinol Metab. 101:561–571.
2016.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Shang K, Jia X, Qiao J, Kang J and Guan Y:
Endometrial abnormality in women with polycystic ovary syndrome.
Reprod Sci. 19:674–683. 2012.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Hu M, Li J, Zhang Y, Li X, Brännström M,
Shao LR and Billig H: Endometrial progesterone receptor isoforms in
women with polycystic ovary syndrome. Am J Transl Res.
10:2696–2705. 2018.PubMed/NCBI
|