Introduction
Regucalcin gene promoter region-related protein-p117
(RGPR-p117; gene symbol, rgpr-p117) was originally
discovered as a novel transcription factor for regucalcin
(rgn) gene expression in 2001 (1,2). The
RGPR-p117 gene consists of 26 exons spanning ~4.1 kbp and is
localized on human chromosome 1q25.2 (2). RGPR-p117 conserves a leucine zipper motif
(2). Thus, RGPR-p117 was termed as the
leucine zipper transcription regulator 2 (Lztr2).
Phosphorylated RGPR-p117 in the nucleus specifically binds to a
nuclear factor I consensus motif, TTGGC(N)6CC that is
located on the 5′-flanking region of the regucalcin (3–8). Regucalcin
has been shown to be pivotal as a suppressor in various cell
signaling pathways and in transcription activity for maintaining
cell homeostasis and disorder in various cell and tissue types
(9–11).
In 2007 it was proposed that RGPR-p117 be renamed to
SEC16 homolog B, endoplasmic reticulum export factor (SEC16B)
(12). RGPR-p117 has a putative role
as an endoplasmic reticulum (ER) export factor implicated in the
transports of proteins and lipids in the ER (13). In recent years, growing evidence
indicates that the RGPR-p117/SEC16B gene is involved in human
obesity and type 2 diabetes (14).
However, to the best of our knowledge, the role of RGPR-p117/SEC16B
as an ER export factor implicated in lipid metabolism has not been
investigated. Thus, the aim of the current review was to discuss
the findings regarding the role of RGPR-p117/SEC16B as an ER export
factor implicated in lipid metabolism and its involvement in human
obesity and type 2 diabetes.
Pivotal role of RGPR-p117 as a transcription
factor
RGPR-p117 enhances rgn gene
expression
The RGPR-p117 gene, rgpr-p117, is expressed
in various species, including humans, rats, mice, dogs, cows, pigs,
rabbits, chickens, frogs (Xenopus), fish (Zebrafish),
C. elegans and yeast (8,15–17). The human rgpr-p117 is located in
human chromosome 1q25.2 and is organized with a minimum of 26 exons
spanning ~4.1 kbp (2,8). RGPR-p117 in the human liver consists of
1,060 amino acid residues with a calculated molecular mass of 117
kDa and an estimated isoelectric point (pI) of 5.71
(2,15).
RGPR-p117 is highly homologous in various types of mammalian
species, indicating a functional significance in vertebrate species
(15–17). Mammalian RGPR-p117 was confirmed to
highly conserve a leucine zipper motif, which is a characteristic
structure of nuclear transcription factors (2,17–21). The leucine zipper pattern is conserved
in numerous gene regulatory proteins, such as CCATT-box/enhancer
binding protein (22), nuclear
oncogenes, fos and jun (23), cAMP response element (CRE) binding
proteins (CREB, CRE-BP1 and ATFs) (24), c-Myc, L-Myc, and
N-Myc oncogenes (25), and
octamer-binding transcription factor 2 (Oct-2/OTF-2) (26). This finding may support the hypothesis
that RGPR-p117 is a transcription factor.
RGPR-p117 was demonstrated to enhance the rgn
promoter activity (7,8). RGPR-p117 possesses various motifs
associated with post-translational modification, such as casein
kinase II phosphorylation, protein kinase C (PKC) phosphorylation,
N-myristoylation, N-glycosylation and amidation
(2). Modified native RGPR-p117 may
bind to the TTGGC motif in the promoter region of the rgn in
the nucleus (5,8). A computer analysis of subcellular
localization of RGPR-p117 from six vertebrates demonstrated a
higher probability of nuclear localization, particularly in rats
and mice (78.3%) (2,17). RGPR-p117 was demonstrated to localize
in the cytoplasm and nucleus of modeled, normal rat kidney proximal
tubular epithelial NRK-52E cells with immunocytochemical analysis
using transient HA-RGPR-p117/phCMV2-transfected cells (5). This nuclear localization was enhanced by
a culture with fetal bovine serum, parathyroid hormone or phorbol
12-myristate 13-acetate which activates various protein kinases,
including PKC (5,27–29).
Overexpression of RGPR-p117 was found to enhance
regucalcin mRNA and protein expression levels in NRK52E cells
(6). Notably, the stimulatory effects
of RGPR-p117 overexpression on the rgn promoter activity in
NRK52E cells transfected with the −710/+18 LUC construct (which
contains the TTGGC sequence) disappeared in the cells that were
transfected with the LUC construct with the deleted TTGGC motif
(7). Furthermore, the stimulatory
effects of overexpressed RGPR-p117 on regucalcin mRNA expression
were blocked by culture with staurosporine, an inhibitor of PKC
(30), while this stimulatory effect
was not suppressed in the presence of dibucaine, an inhibitor of
Ca2+/calmodulin-dependent protein kinase (29) or PD98059, an inhibitor of MAP kinase
(31). The stimulatory effects of
RGPR-p117 on regucalcin mRNA expression were mediated through
PKC-dependent signaling process that stimulates protein
phosphorylation in the nucleus of NRK52E cells (6). Furthermore, regucalcin mRNA expression
has been shown to be stimulated through Ca2+ signaling
factors, which have been associated with PKC in modeled rat
hepatoma H4-II-E cells (29) and
NRK52E cells (32). In addition,
stimulatory effects of overexpressed RGPR-p117 on the rgn
gene promoter activity were clearly blocked by culture with
vanadate, an inhibitor of tyrosine kinase (6,33). Such an
effect was not revealed in the case of okadaic acid, an inhibitor
of protein serine/threonine phospharase (6,33). The
inhibition of protein tyrosine phosphatases contributed to
demonstrating the stimulatory effects of RGPR-p117 on the
rgn promoter activity (6,33).
As described above, phosphorylated RGPR-p117 in the
nucleus may directly bind to the TTGGC motif of the rgn
promoter region and enhance the transcription activity of
rgn expression (5,7,8).
RGPR-p117 regulates the gene
expression implicated in apoptotic cell death
Overexpression of RGPR-p117 did not exhibit a
significant effect on proliferation in NRK52E cells (34). Notably, overexpressed RGPR-p117
demonstrated protective effects on apoptotic cell death induced by
tumor necrosis factor-α, lipopolysaccharides or Bay K8644 in
NRK-52E cells (34). These
factor-induced cell deaths were clearly prevented by culture with
inhibitor of caspase-3, which activates endonuclease to mediate
nuclear DNA cleavage into oligonucleosome fragments (35,36) in
NRK52E cells (37). Mechanically,
overexpression of RGPR-p117 was demonstrated to decrease the mRNA
expression levels of Fas-associating death domain (FADD) protein,
caspase-8, caspase-9, and caspase-3 that are implicated in the
stimulation of apoptotic cell death in NRK52E cells (37). RGPR-p117 may exert protective effects
on apoptotic cell death due to suppressing the gene expression of
various proteins implicated in the stimulation of apoptosis
(37). Notably, the TTGGC motif, which
is present in the rgn promoter region (1,2), is also
located in the promoter region of the genes of caspase-3, caspase-8
or FADD (2,37). RGPR-p117, which binds to the TTGGC
motif in the promoter region of caspase-3, caspase-8 or FADD, may
repress the gene expression levels of caspase-3, caspase-8 or FADD
in the nucleus of NRK52E cells.
Thus, RGPR-p117 may be critical in cell regulation
as a transcription factor in the gene expression of various
proteins. As summarized in Fig. 1,
cytoplasm RGPR-p117 is translocated to the nucleus via a signaling
mechanism associated with Ca2+-dependent PKC in cells.
Nuclear RGPR-p117, which is phosphorylated by various protein
kinases, including PKC, specifically binds to the TTGGC motif in
the promoter region of the various target genes in the nucleus,
leading to regulation of the gene expression in cells. RGPR-p117
possesses a leucine zipper motif, which is conserved in various
species (1,2). Thus, RGPR-p117 may be crucial role as a
transcription factor for gene expression in various cell and tissue
types.
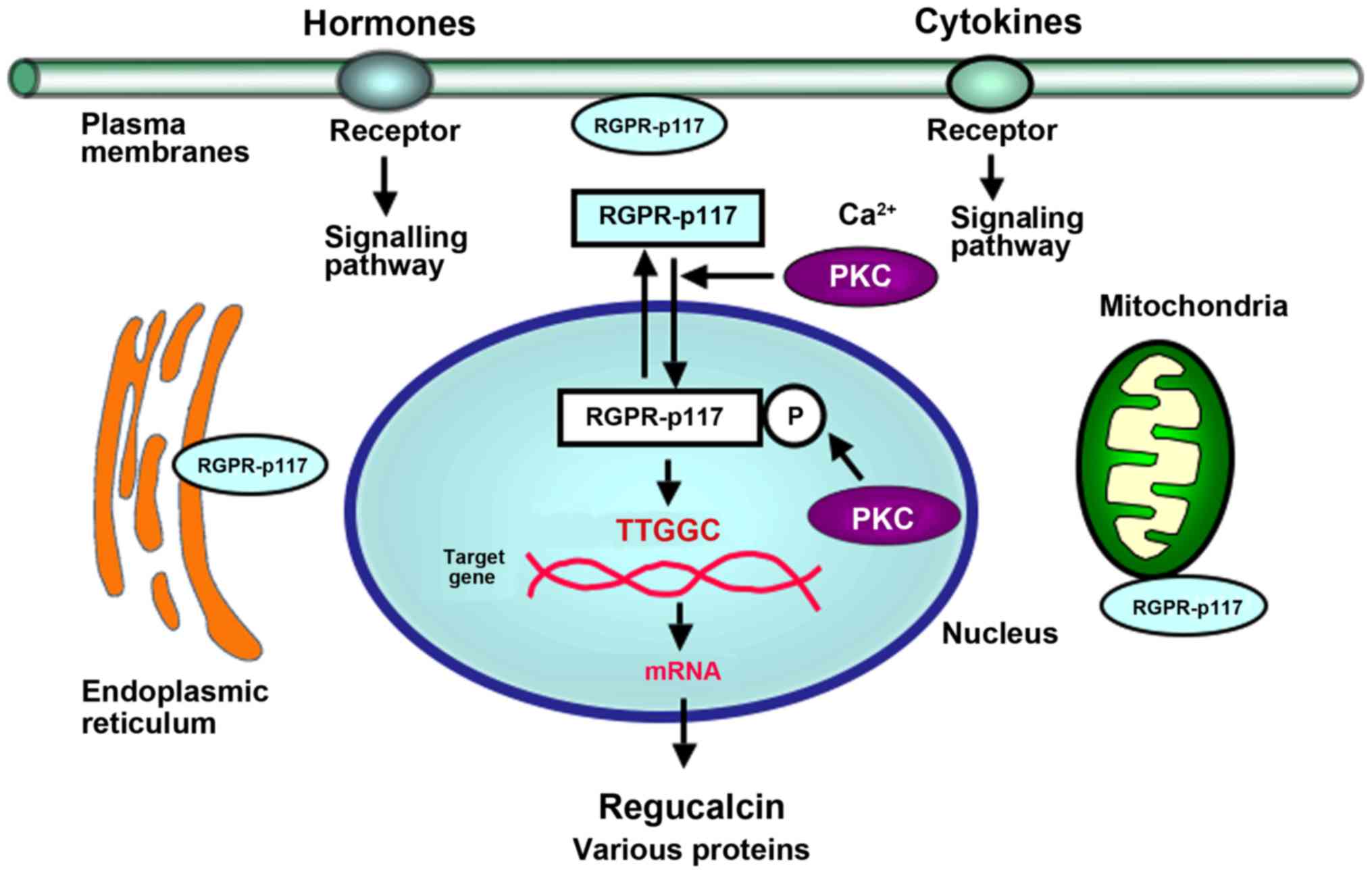 | Figure 1.RGPR-p117 has a crucial role as a
transcription factor. Cytoplasmic RGPR-p117 is translocated to the
nucleus, which is enhanced via activation of Ca2+
signaling-dependent PKC. Nuclear RGPR-p117 is phosphorylated by
PKC. Phosphorylated RGPR-p117 in the nucleus binds to the TTGGC
motif in the promoter region of the regucalcin gene, as well as
other genes, and it regulates the transcription activity of target
genes. RGPR-p117 exerts suppressive effects on DNA and protein
synthesis, and protective effects on apoptotic cell death.
Furthermore, RGPR-p117 is localized in the plasma membranes,
mitochondria and ER in cells. RGPR-p117 may have a role as an ER
export factor for lipids that are implicated in obesity. RGPR-p117,
regucalcin gene promoter region-related protein; PKC, protein
kinase C; ER, endoplasmic reticulum; P, phosphorylated. |
Role of RGPR-p117/SEC16B as an ER export
factor
The organelle distribution of RGPR-p117 has been
analyzed in NRK-52E cells transfected with HA-RGPR-p117/phCMV2
(34). Results of western blot
analysis for HA-RGPR-p117 demonstrated that this protein localizes
in the plasma membranes, cytoplasm, mitochondria, ER and nucleus of
the cells (34). RGPR-p117 may have a
regulatory role in these organelles. Of note, overexpression of
RGPR-p117 was shown to decrease the protein and DNA contents in
NRK52E cells (34).
RGPR-p117 was shown to possess higher homology to
SEC16B (2,12). Furthermore, SEC16B has been reported to
be an ER export factor (13).
RGPR-p117 was demonstrated to bind to the ER of normal rat kidney
NRK52E cells (35). RGPR-p117 has been
suggested to perform a functional role in ER export within
cells.
The ER represents the entry point into the secretory
pathway, and from here newly synthesized proteins and lipids are
delivered to the Golgi (13,38,39). The
selective cargo export from the ER is mediated by coat protein II
(COPII)-assembly at specific sites of the ER, the transitional ER
(tER) (13,38). The peripheral membrane protein, SEC16
localizes to tER and may be key to the organization of these sites,
and this protein defines the tER and is hypothesized to act as a
scaffold for COPII-assembly (13,38). SEC16
may be crucial in the formation of COPII vesicles, which mediate
transport of proteins and lipids from the ER to the Golgi apparatus
(13,38). Two isoforms of SEC16 have been reported
as follows: The larger isoform, SEC16A, and the smaller isoform,
SEC16B (13,38). However, the functional differences
between the two isoforms are ill-defined (13,38). SEC16B
was shown as not functionally redundant in ER export and
transitional ER organization (13,38).
Characterization of the localization and dynamics of SEC16B
relative to Sec16A provide evidence that SEC16B may be a minor, or
perhaps specialized, form of SEC16 and that it is not functionally
redundant with SEC16A (38–40).
SEC16B, as well as SEC16A, defines ER exit sites,
where COPII vesicles are formed in mammalian cells (38,39). SEC16A
and SEC16B, mammalian orthologs of yeast SEC16, are scaffold
proteins that organize ER exit sites by interacting with COPII
components (39). Overexpressed SEC16B
targeted the entire ER, whereas SEC16A was predominantly cytosolic
(39). Concomitant with the
overexpression of SEC16B, peroxisomal biogenesis factor 3 (PEX3)
and PEX16 were redistributed from peroxisomes to SEC16B-positive ER
membranes (39). Knockdown of SEC16B,
but not SEC16A, by RNAi affected the peroxisome morphology,
inhibited the transport of PEX16 from the ER to the peroxisomes,
and suppressed the expression of PEX3 (39). These phenotypes were reversed by the
expression of RNAi-resistant SEC16B (39). SEC16B, but not SEC16A, was demonstrated
to regulate the transport of peroxisomal biogenesis factors from
the ER to peroxisomes in mammalian cells (39). Furthermore, the C-terminal region of
SEC16B, which is not conserved in SEC16A, is required for this
function (38,39). SEC16B, in ER areas other than ER exit
sites, was demonstrated to perform this role, providing an
explanation of how secretory and peroxisomal trafficking from the
ER are distinguished (38).
As described above, RGPR-p117 may be involved in ER
export process in the cells. However, more studies are required to
further elucidate this process.
Involvement of RGPR-p117/SEC16B in human
obesity
There is growing evidence to indicate that
RGPR-p117/SEC16B may be involved in human obesity (41–46). Obesity
is a major health issue worldwide, with increasing prevalence in
adults and children from developed and developing countries
(41). Obesity causes various chronic
diseases, including cardiovascular and respiratory disease,
osteoarthritis, hypertension, stroke, type II diabetes, obstructive
sleep apnea, and different types of cancer (41). At least 52 genetic loci have been shown
to associate with obesity-associated traits (42). Genome-wide association studies (GWAS)
effectively reveal an association between single nucleotide
polymorphisms and different measures of obesity, and have
identified a number of genes associated with human body weight
(41–46). Previous GWAS have identified numerous
genes of proteins associated with obesity, including leptin (LEP),
LEP receptor, proopiomelanocortin, proprotein convertase
subtilisin/kexin type 1, α-ketoglutarate dependent dioxygenase
(FTO), melanocortin 3 receptor (MC3R), MC4R,
glucosamine-6-phosphate deaminase 2 (GNPDA2), transmembrane protein
18, glutaminyl-peptide cyclotransferase like/gastric inhibitory
polypeptide receptor, brain derived neurotrophic factor (BDNF), ETS
variant 5 (ETV5), mitogen-activated protein kinase kinase 5/SKI
family transcriptional corepressor 1, SEC16B, single-minded family
bHLH transcription factor 1, and tankyrase/methionine sulfoxide
reductase A (45). RGPR-p117/SEC16B
may be a kind of gene implicated in human obesity.
The involvement of SEC16B in obesity has been
investigated in model animals using obese diabetic rats (47). Schmid et al (47) performed an expression analysis of 14
genes, namely BDNF, ETV5, Fas apoptotic inhibitory molecule 2
(FAIM2), FTO, GNPDA2, potassium channel tetramerization domain
containing 15, lysophospholipase like 1 (LYPLAL1), MC4R,
mitochondrial carrier 2, neuronal growth regulator 1, neurexin 3,
transmembrane protein 18, SEC16B, and transcription factor AP-2 β
(TFAP2B), via reverse transcription-quantitative polymerase chain
reaction in the adipose tissue of the kidney capsule, the
mesenterium and subcutaneum, as well as the hypothalamus of obese
Zucker diabetic fatty (ZDF) and Zucker lean (ZL) rats aged 22
weeks. All of the target genes, not including SEC16B, exhibited the
highest expression levels in the hypothalamus, indicating a
critical role of these obesity-associated genes in the central
regulation of energy balance (47).
Notably, the expression patterns observed in the hypothalamus were
not different between the obese ZDF and lean ZL rats (47). However, LYPLAL1, TFAP2B, SEC16B and
FAIM2 were expressed significantly less in the kidney fat of the
ZDF rats compared with the ZL rats (47). The expression pattern of the
investigated obesity genes may imply a role in the central
regulation of energy balance, but, for certain genes, an additional
role within the adipose tissue (47).
However, the role of SEC16B in lipid metabolism in animal models
remains to be elucidated.
RGPR-p117, which is a transcription factor (1,2,8), has been shown to enhance the gene
expression of regucalcin in kidney cells (7,8). Regucalcin
has been found to stimulate adipogenesis in bone marrow mesenchymal
stem cells (48), and overexpressed
regucalcin was demonstrated to cause diabetes and lipid metabolic
disorders with hyperlipidemia in vivo (49,50).
RGPR-p117 is speculated to partly participate in lipid metabolic
disorder, which is mediated through regucalcin. As RGPR-p117/SEC16B
may regulate ER export implicated in lipids (13), RGPR-p117/SEC16B is speculated to
contribute to lipid metabolic disorders.
Further studies are required to elucidate the direct
evidence regarding the role of RGPR-p117/SEC16B in the regulation
of lipid and glucose metabolism, which are implicated in obesity
and diabetes. It is possible that the target genes of RGPR-p117 may
be involved in human obesity.
Conclusion
RGPR-p117 was originally discovered as a novel
protein that binds to a nuclear factor I consensus motif,
TTGGC(N)6CC in the promoter region of the rgn
gene (1,2). Phosphorylated RGPR-p117, which possesses
a leucine zipper motif, was demonstrated to be crucial as a
transcription factor in the expression of various target genes
(7,9).
Furthermore, RGPR-p117 localizes in the plasma membranes,
mitochondria and ER of modeled kidney cells (34), indicating that this protein has a
multifunctional role in the regulation of cellular functions.
Notably, RGPR-p117 exhibited a protective role in apoptotic cell
death evoked by various stimulatory factors in vitro
(37). Additionally, incomplete
splicing for the human RGPR-p117 gene is suggested to be involved
in carcinogenesis in human placenta (2,9). Thus,
RGPR-p117 may have a multifunctional role in cell regulation.
RGPR-p117/SEC16B was proposed to be involved as an
ER export factor implicated in the transport of proteins and lipids
in the ER (12). As a result of this
finding, it was proposed that RGPR-p117 is renamed to SEC16B
(12). There is currently growing
evidence that RGPR-p117/SEC16B is involved in human obesity and
diabetes. However, to the best of our knowledge, the existing
studies do not provide direct evidence that RGPR-p117/SEC16B has a
regulatory role in lipid metabolism in vivo and in
vitro. It is possible that RGPR-p117, a transcription factor,
may target the gene expression for molecules associated with lipid
metabolism. However, further studies are required to elucidate this
fully.
References
|
1
|
Misawa H and Yamaguchi M: Identification
of transcription factor in the promoter region of rat regucalcin
gene: Binding of nuclear factor I-A1 to TTGGC motif. J Cell
Biochem. 84:795–802. 2000. View Article : Google Scholar
|
|
2
|
Misawa H and Yamaguchi M: Molecular
cloning and sequencing of the cDNA coding for a novel regucalcin
gene promoter region-related protein in rat, mouse and human liver.
Int J Mol Med. 8:513–520. 2001.PubMed/NCBI
|
|
3
|
Miasawa H and Yamaguchi M: Involvement of
hepatic nuclear factor I binding motif in transcriptional
regulation of Ca2+-binding protein regucalcin gene. Biochem Biophys
Res Commun. 269:270–278. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Misawa H and Yamaguchi M: Intracellular
signaling factors-enhanced hepatic nuclear protein binding to TTGGC
sequence in the rat regucalcin gene promoter: Involvement of
protein phosphorylation. Biochem Biophys Res Commun. 279:275–281.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sawada N, Nakagawa T, Murata T and
Yamaguchi M: Nuclear localization of a novel protein, RGPR-p117, in
cloned normal rat kidney proximal tubular epithelial cells. Int J
Mol Med. 16:809–814. 2005.PubMed/NCBI
|
|
6
|
Sawada N and Yamaguchi M: Overexpression
of RGPR-p117 enhances regucalcin gene expression in cloned normal
rat kidney proximal tubular epithelial cells. Int J Mol Med.
16:1049–1055. 2005.PubMed/NCBI
|
|
7
|
Sawada N and Yamaguchi M: Overexpression
of RGPR-p117 enhances regucalcin gene promoter activity in cloned
normal rat kidney proximal tubular epithelial cells: Involvement of
TTGGC motif. J Cell Biochem. 99:589–597. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yamaguchi M: Novel protein RGPR-p117: Its
role as the regucalcin gene transcription factor. Mol Cell Biochem.
327:53–63. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yamaguchi M: Role of regucalcin in calcium
signaling. Life Sci. 66:1769–1780. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yamaguchi M: Role of regucalcin in
maintaining cell homeostasis and function (Review). Int J Mol Med.
15:372–389. 2005.
|
|
11
|
Yamaguchi M: Regucalcin and cell
regulation: Role as a suppressor in cell signaling. Mol Cell
Biochem. 353:101–137. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bhattacharyya D and Glick BS: Two
mammalian Sec16 homologues have nonredundant functions in
endoplasmic reticulum (ER) export and transitional ER organization.
Mol Biol Cell. 18:839–849. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Budnik A and Stephens DJ: ER exit
sites-Localization and control of COPII vesicle formation. FEBS
Lett. 583:3796–3803. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ng MCY, Tam CHT, So WY, Ho JSK, Chan AW,
Lee HM, Wang Y, Lam VKL, Chan JCN and Ma RCW: Implication of
genetic variants near NEGR1, SEC16B, TMEM18, ETV5/DGKG, GNPDA2,
LIN7C/BDNF, MTCH2, BCDIN3D/FAIM2, SHsB1, FTO, MC4R, and KCTD15 with
obesity and type 2 diabetes in 7705 Chinese. J Clin Endocrinol
Metab. 95:2418–2425. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Misawa H and Yamaguchi M: Gene expression
for a novel protein RGPR-p117 in various species: The stimulation
by intracellular signaling factors. J Cell Biochem. 87:188–193.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamaguchi M, Misawa H and Ma ZJ: Novel
protein RGPR-p117: The gene expression in physiologic state and the
binding activity to regucalcin gene promoter region in rat liver. J
Cell Biochem. 88:1092–1100. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sawada N and Yamaguchi M: A novel
regucalcin gene promoter region-related protein: Comparison of
nucleotide and amino acid sequences in vertebrate species. Int J
Mol Med. 15:97–104. 2005.PubMed/NCBI
|
|
18
|
Bairch A, Bucher P and Hofmann K: The
PROSITE database, its status in 1997. Nucleic Acids Res.
25:217–221. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Landschulz WH, Johnson PF and McKnight SL:
The leucine zipper: A hypothetical structure common to a new class
of DNA binding proteins. Science. 240:1759–1764. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Busch SJ and Sassone-Corsi P: Dimers,
leucine zippers and DNA-binding domains. Trends Genet. 6:36–40.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
O'Shea EK, Rutkowski R and Kim PS:
Evidence that the leucine zipper is a coiled coil. Science.
243:538–542. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vinson CR, Sigler PB and McKnight SL:
Scissors-grip model for DNA recognition by a family of leucine
zipper proteins. Science. 246:911–916. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
O'Shea EK, Rutkowski R, Stafford WF III
and Kim PS: Preferential heterodimer formation by isolated leucine
zippers from fos and jun. Science. 245:646–648. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maekawa T, Sakura H, Kanei-Ishii C, Sudo
T, Yoshimura T, Fujisawa J, Yoshida M and Ishii S: Leucine zipper
structure of the protein CRE-BP1 binding to the cyclic AMP response
element in brain. EMBO J. 8:2023–2028. 1989.PubMed/NCBI
|
|
25
|
Collum RG and Alt FW: Are myc proteins
transcription factors? Cancer Cells. 2:69–75. 1990.PubMed/NCBI
|
|
26
|
Clerc RG, Corcoran LM, LeBowitz JH,
Baltimore D and Sharp PA: The B-cell-specific Oct-2 protein
contains POU box- and homeo box-type domains. Genes Dev.
2:1570–1581. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Verheijen MHG and Defize LHK: Parathyroid
hormone activates mitogen-activated protein kinase via a
cAMP-mediated pathway independent of rats. J Biol Chem.
272:3423–3429. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nishizuka Y: Studies and perspectives of
protein kinase C. Science. 233:305–312. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamaguchi M and Nakajima M: Involvement of
intracellular signaling factors in the serum-enhanced Ca2+-binding
protein regucalcin mRNA expression in the cloned rat hepatoma cells
(H4-II-E). J Cell Biochem. 74:81–89. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tamaoki T, Nomoto H, Takahashi I, Kato Y,
Morimoto M and Tomita E: Staurosporine, a potent inhibitor of
phospholipid/Ca2+ dependent protein kinase. Biochem Biophys Res
Commun. 135:397–402. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Peleck SL, Charest DL, Mordret GP, Siow
YL, Palaty C, Campbell D, Chaslton L, Samiei M and Sanghera JS:
Networking with mitogen-activated protein kinases. Mol Cell
Biochem. 127:157–169. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nakagawa T and Yamaguchi M: Hormonal
regulation on regucalcin mRNA expression in cloned normal rat
kidney proximal tubular epithelial NRK52E cells. J Cell Biochem.
95:589–597. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hunter T: Protein kinases and
phosphatases: The Yin and Yang of protein phosphorylation and
signaling. Cell. 80:225–236. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tomono S, Sawada N and Yamaguchi M:
Overexpression of RGPR-p117 induces the decrease in protein and DNA
contents in cloned normal rat kidney proximal tubular epithelial
NRK52E cells. Int J Mol Med. 20:79–83. 2007.PubMed/NCBI
|
|
35
|
Ribeiro JM and Carson DA:
Ca2+/Mg2+-dependent endonuclease from human spleen: Purification,
properties, and role in apoptosis. Biochemistry. 32:9129–9136.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yamaguchi M and Sakurai K: Inhibitory
effect of calcium-binding protein regucalcin on Ca2+-activated DNA
fragmentation in rat liver nuclei. FEBS Lett. 279:281–284. 2002.
View Article : Google Scholar
|
|
37
|
Yamaguchi M, Tomono S and Nakagawa T:
Overexpression of RGPR-p117 suppresses apoptotic cell death and its
related gene expression in cloned normal rat kidney proximal
tubular epithelial NRK52E cells. Int J Mol Med. 20:565–571.
2007.PubMed/NCBI
|
|
38
|
Tani K, Tagaya M, Yonekawa S and Baba T:
Dual function of Sec16B: Endoplasmic reticulum-derived protein
secretion and peroxisome biogenesis in mammalian cells. Cell
Logist. 1:164–167. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yonekawa S, Furuno A, Baba T, Fujiki Y,
Ogawasara Y, Yamamoto A, Tagaya M and Tani K: Sec16B is involved in
the endoplasmic reticulum export of the peroxisomal membrane
biogenesis factor peroxin 16 (Pex16) in mammalian cells. Proc Natl
Acad Sci USA. 108:12746–12751. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Budnik A, Heesom KJ and Stephens DJ:
Characterization of human Sec16B: Indications of specialized,
non-redundant functions. Sci Rep. 1:772011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hotta K, Kitamoto T, Kitamoto A, Mizusawa
S, Matsuo T, Nakata Y, Kamohara S, Miyatake N, Kotani K, Komatsu R,
et al: Association of variations in the FTO, SCG3 and MTMR9 genes
with metabolic syndrome in a Japanese population. J Hum Genet.
56:647–651. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Albuquerque D, Nóbrega C, Rodríguez-López
R and Manco L: Association study of common polymorphisms in MSRA,
TFAP2B, MC4R, NRXN3, PPARGC1A, TMEM18, SEC16B, HOXB5 and OLFM4
genes with obesity-related traits among Portuguese children. J Hum
Genet. 59:307–313. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Williams MJ, Almén MS, Fredriksson R and
Schiöth HB: What model organisms and interactomics can reveal about
the genetics of human obesity. Cell Mol Life Sci. 69:3819–3834.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lv D, Zhang DD, Wang H, Zhang Y, Liang L,
Fu JF, Xiong F, Liu GL, Gong CX, Luo FH, et al: Genetic variations
in SEC16B, MC4R, MAP2K5 and KCTD15 were associated with childhood
obesity and interacted with dietary behaviors in Chinese school-age
population. Gene. 560:149–155. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kaewsutthi S, Santiprabhob J, Phonrat B,
Tungtrongchitr A, Lertrit P and Tungtrongchitr R: Exome sequencing
in Thai patients with familial obesity. Genet Mol Res. 15:83112016.
View Article : Google Scholar
|
|
46
|
Ng MC, Tam CH, So WY, Ho JS, Chan AW, Lee
HM, Wang Y, Lam VK, Chan JC and Ma RC: Implication of genetic
variation near NEGR1, SEC16B, TMEM18, ETV5/DGKG, GNPDA2,
LIN7C/BDNF, MTCH2, BCDIN3D/FAIM2, SH2B1, FTO, MC4R and KCTD15 with
obesity and type 2 duabetes in 7705 Chinese. J Clin Endocrinol
Metab. 95:2418–2425. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Schmid PM, Heid I, Buechler C, Steege A,
Resch M, Birner C, Endemann DH, Riegger GA and Luchner A:
Expression of fourteen novel obesity-related genes in Zucker
diabetic fatty rats. Cardiovas Diabetol. 11:482012. View Article : Google Scholar
|
|
48
|
Yamaguchi M, Weitzmann MN, Baile CA and
Murata T: Exogenous regucalcin suppresses osteoblastogenesis and
stimulates adipogenesis in mouse bone marrow culture. Integr Biol
(Camb). 4:1215–1222. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yamaguchi M: Regucalcin and metabolic
disorders: Osteoporosis and hyperlipidemia are induced in
regucalcin transgenic rats. Mol Cell Biochem. 341:119–133. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yamaguchi M and Murata T: Involvement of
regucalcin in lipid metabolism and diabetes. Metabolism.
62:1045–1051. 2013. View Article : Google Scholar : PubMed/NCBI
|














