Introduction
Iron is one of the essential trace elements for
animal organisms, and is involved in numerous life processes, such
as oxygen transport (1), DNA
synthesis (2), the host defense
and inflammation (3). Disruption
of iron homeostasis caused by iron deficiency or overload is
strongly associated with some of the most common human diseases
(4,5). Iron can influence the clinical
course of several chronic metabolic diseases, such as type 2
diabetes (6), obesity (7), non-alcoholic fatty liver disease
(8) and atherosclerosis (9).
Iron overload has been linked to a variety of human
diseases, such as hereditary haemochromatosis (HH), thalassemias
and neurodegeneration (10,11). Iron overload diseases result in
excess body iron deposition, which can be caused by genetic or
secondary causes. HH is defined as a genetically derived systemic
iron overload caused by decreased concentrations of the iron
regulatory hormone hepcidin or decreased hepcidin-ferroportin
binding (12). Secondary iron
overload may result from frequent blood transfusions, exogenous
iron intake or certain haematological disorders, such as refractory
anemia or aplastic anemia (13).
Iron overload can cause toxic accumulation in the liver, heart,
joints or endocrine glands (14).
However, the clinical manifestations of iron overload syndrome are
complex and far from being understood.
The pancreas is commonly affected in iron overload
syndromes. Clinical data show that patients with HH are at risk of
developing diabetes due to β-cell dysfunction, but may display iron
overload in the pancreas (15).
Significant pancreatic iron overload has been reported in other
iron overload disorders, including thalassaemia, sickle cell
anaemia and Diamond-Blackfan anaemia (16-18). Iron overload is also significantly
associated with exocrine pancreatic dysfunction. The most common
disease of exocrine pancreatic dysfunction is pancreatitis, which
mainly includes acute pancreatitis (AP) and chronic pancreatitis
(CP). AP is an inflammatory disease of the pancreas that is
associated with high morbidity and mortality rates (19). AP is characterized by acinar cell
death and local and systemic inflammation (20), whereas CP, a progressive and
irreversible fibroinflammatory disease of the pancreas, consists of
inflammation and pancreatic fibrosis in individuals with genetic,
environmental and other risk factors (21). CP is characterized by pancreatic
atrophy, fibrosis, ductal stenosis and distortion, calcification,
exocrine insufficiency and diabetes mellitus (22). Pancreatitis causes alterations in
circulating markers of iron in patients. In a recent study, serum
iron, serum ferritin and transferrin saturation levels were all
increased in patients with pancreatitis compared with those in
healthy subjects, whereas these parameters were significantly
decreased in patients after treatment (23). Pancreatic iron deposition has been
observed in several genetic mutation models of iron overload, such
as hepcidin knockout mice (24),
bone morphogenetic protein-deficient mice (25) and ceruloplasmin mutants (26). However, to the best of our
knowledge, there are no studies on the effect of non-hereditary
iron overload on the pancreas.
Overall, iron plays a key role in life activities as
an essential nutrient for body growth. Iron homeostasis plays an
important role in the maintenance of pancreatic health and the
development of diseases, but the specific effects and mechanisms of
iron overload in the development of these pancreatic diseases are
still unclear. Considering the well-established role of iron
metabolism and pancreatic function in multiple transgenic mouse
models, the present study aimed to further explore the pathogenic
consequences of iron overload on pancreatic tissue in mice using a
model of non-hereditary iron overload.
Materials and methods
Animals
A total of 20, male, 8-week-old, C57BL/6 (20±2 g)
mice were purchased from Shanghai Slack Laboratory Animal Center.
The mice were housed in an environment with a 12-h light/dark cycle
at 24°C and a relative humidity of 50-70%. The mice were given free
access to water and food, and the bedding was changed every other
day. After 7 days of acclimation, the mice were randomly divided
into the following two groups with 10 mice each: Control group and
iron overload group. The iron overload mice were injected
intraperitoneally with 120 mg/kg body weight of iron dextran
(Pharmacosmos A/S) every other week for 12 weeks. Since infused
dextran is eliminated 70% by the kidney and 30% by the
gastrointestinal tract, the control mice were injected
intraperitoneally with saline as reported previously (27). Mice were sacrificed by cervical
dislocation at the end of the experiment. The blood and pancreas
were collected. Pancreas tissues were rapidly dissected and fixed
in 4% paraformaldehyde solution at room temperature for 24 h for
histological analysis, or frozen in liquid nitrogen and stored at
−80°C until further analysis. All animal experiments were approved
by the Committee of Experimental Animal Care of Zhejiang University
(Hangzhou, China; approval no. 20077).
Serum biochemical assays
Mouse serum was separated from the blood samples by
centrifugation at 3,000 × g for 15 min at 4°C. A serum iron assay
kit (cat. no. A039-1-1; Nanjing Jiancheng Bioengineering Institute)
and total iron binding capacity assay kit (cat. no. A040-1-1;
Nanjing Jiancheng Bioengineering Institute) were used to detect the
iron level of the mice according to the manufacturer's
instructions. For serum iron detection, iron chromogen was added to
the serum and incubated in boiling water for 5 min. After cooling
and centrifugation at 3,000 × g for 10 min at 4°C, the absorbance
at 520 nm was measured by a fluorescence spectrophotometer. For
measurement of total iron binding capacity, the serum was mixed
with 179.1 mmol/l iron standard so that all transferrin bound iron
in the serum. The excess iron in the serum was then adsorbed away
using an iron adsorbent, and the iron content was measured by
assessing the iron level in the serum. A lipase assay kit (cat. no.
A054-2-1) and an α-amylase assay kit (cat. no. C016-1-1) (both
Nanjing Jiancheng Bioengineering Institute) were used to detect
amylase and lipase levels, respectively, in the mouse serum.
α-amylase hydrolyzes the starch in the substrate. In the case of
known substrate concentration and excess, the added iodine solution
can combine with the unhydrolyzed starch in the substrate to form a
blue complex. The absorbance at 660 nm was measured by a
fluorescence spectrophotometer to obtain the amount of starch that
had been hydrolyzed, and thus the activity of α-amylase was
calculated. Latex made of triglyceride and water has a turbid
character due to the absorption and scattering of incident light by
its micelles. The triglycerides in the micelles are hydrolyzed by
the action of lipase, which causes the micelles to split and thus
the scattered light or turbidity is reduced. Lipase activity was
calculated by measuring the absorbance at 420 nm using fluorescence
spectrophotometer and measuring the rate of reduction of scattered
light or turbidity.
Histological analysis
Paraffin-embedded samples were cut into 5-µm
sections. For haematoxylin and eosin (H&E) staining, paraffin
sections were deparaffinized with xylene and rehydrated using
different concentrations of alcohol, and then stained with
haematoxylin for 5 min. The sections were differentiated in aqueous
hydrochloric acid for 2 sec and blunted in aqueous ammonia for
15-30 sec at room temperature. The sections were put into eosin
staining solution for 5-8 sec at room temperature. The sections
were mounted with neutral gum after dehydration using absolute
ethanol and xylene. The H&E staining then examined by light
microscope (Leica Microsystems GmbH) and image acquisition was
performed to analyze the histopathological features of the
pancreatic sections.
Prussian blue staining was used to detect iron
deposition in the pancreatic tissues. The 5-µm sections of
pancreas were deparaffinized with xylene and rehydrated using
different concentrations of alcohol. Pancreatic tissue was
incubated in a mixture (1:1) of 2% potassium ferrocyanide and 2%
hydrochloric acid for 30 min at room temperature. Sections were
then rinsed with PBS (cat. no. KGB5001; Nanjing KeyGen Biotech Co.,
Ltd.) and counterstained with eosin for 20 sec at room temperature.
The sections were mounted with neutral gum after dehydration using
absolute ethanol and xylene.
Masson staining and Sirius red staining were
employed to evaluate the collagen content of the pancreas. For
Masson staining, paraffin sections were deparaffinized with xylene
and rehydrated using different concentrations of alcohol. The
sections were then placed in potassium dichromate standards at room
temperature overnight (~17 h). Sections were treated with aqueous
phosphomolybdic acid for 1-2 min and counterstained with aniline
blue liquid for 5 sec at room temperature. Sections were
sequentially treated with 1% glacial acetic acid for 5-10 sec each
at room temperature. Sections were dehydrated and mounted using
neutral resin glue. For Sirius red staining, pancreatic tissue
sections were deparaffinized and stained with Wiegert's iron
haematoxylin stain for 15 min at room temperature. Next, pancreatic
sections were differentiated with an acidic differentiation
solution. After washing the sections in tap water for 10 min, the
pancreatic tissue was stained with Sirius red staining droplets for
1 h at room temperature. The sections were gently rinsed with a
flowing water stream to remove the surface dye solution. All
sections were examined using a DM3000 microscope (Leica
Microsystems GmbH). The inflammation, atrophy and fibrosis in the
pancreatic tissues were scored from 0 to 3 using a
histopathological scoring criteria system, as previously reported
(28).
Immunohistochemical analysis
The pancreatic sections were deparaffinized in
xylene and then sequentially dehydrated in different concentrations
of ethanol solution. Endogenous peroxidase activity was blocked
with 3% H2O2 for 10 min, and sections were
resuspended in antigen retrieval solution (pH 6.0) and boiled in a
microwave oven for 5 min. Pancreatic tissue was then blocked with
3% bovine albumin [cat. no. 36101ES25; Yeasen Biotechnology
(Shanghai) Co., Ltd.]. for 1 h at room temperature. Sections were
incubated overnight at 4°C with anti-CD11b antibody (1:1,000; cat.
no. ab133357), anti-F4/80 antibody (1:5,000; cat. no. ab300421) and
anti-CD3 antibody (1:150; cat. no. ab16669) antibodies (all Abcam),
then washed with Tris-buffered saline containing 0.1% Tween-20
(TBST). The pancreatic tissues were incubated with anti-rabbit IgG
H&L (conjugated with horseradish peroxidase; 1:5,000 cat. no.
ab205718; Abcam) for 1 h at room temperature, and then stained and
developed with horseradish peroxidase for 30 min. Finally, the
nuclei were counterstained using haematoxylin solution for 30 sec
at room temperature. All sections were examined using a DM3000
microscope (Leica Microsystems GmbH). In this experiment,
immunohistochemical quantification was performed using ImageJ
software (version 2.0; National Institutes of Health).
Determination of pancreatic
malondialdehyde (MDA) content, and superoxide dismutase (SOD) and
glutathione peroxidase (GSH-PX) activity
The levels of MDA, SOD and GSH-PX in the pancreatic
tissues were measured by MDA (cat. no. A003-1-1), SOD (cat. no.
A001-1-1) and GSH-PX (cat. no. A005-1-2) assay kits (Nanjing
Jiancheng Bioengineering Institute), respectively, according to the
manufacturer's instructions. MDA can combine with thiobarbituric
acid to form a red product with an absorption maximum at 532 nm
using a fluorescence spectrophotometer. SOD generates superoxide
anion radical (O2−) through xanthine and the
xanthine oxidase reaction system, which oxidizes hydroxylamine to
form nitrite, presenting a purple red color under the action of
chromogenic agents. Therefore, the activity of SOD can be
determined by measuring the absorbance at 550 nm using a
fluorescence spectrophotometer. GSH-PX can promote the reaction of
hydrogen peroxide (H2O2) with reduced GSH to
produce H2O and the oxidized GSH. The activity of GSH-PX
can be expressed as the rate of its enzymatic reaction. GSH and
dithiodinitrobenzoic acid act to generate 5-thiodinitro-benzoate
anions exhibiting a more stable yellow color, and the amount of GSH
can be calculated by measuring its absorbance at 412 nm using a
fluorescence spectrophotometer.
Analysis by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from the pancreas was extracted using
Total RNA Extraction Reagent (cat. no. BS259A; Biosharp Life
Sciences) and reverse-transcribed into cDNA using
Hifair® III 1st Strand cDNA Synthesis SuperMix for qPCR
[cat. no. 11141ES60; Yeasen Biotechnology (Shanghai) Co., Ltd.]
according to the manufacturer's protocols. The concentration of RNA
was determined using a Nanodrop 2000 spectrophotometer (Thermo
Fisher Scientific, Inc.). Real-time qPCR was performed using Hief
UNICON® qPCR SYBR Green Master Mix [cat. no. 11200ES08;
Yeasen Biotechnology (Shanghai) Co., Ltd.] and the ABI 7500
Real-Time PCR system (Thermo Fisher Scientific, Inc.). The
following thermocycling conditions were used for the qPCR: Initial
denaturation at 95°C for 1 min; followed by 40 cycles at 95°C for
15 sec and 60°C for 1 min. The fold difference in gene expression
was calculated using the 2−ΔΔCq method and presented
relative to endogenous β-actin mRNA (29). All reactions were performed at
least in triplicate. Primer sequences are listed in Table I.
 | Table IPrimers used in quantitative
polymerase chain reaction. |
Table I
Primers used in quantitative
polymerase chain reaction.
| Gene | Primers
(5′-3′) | Accession
number |
|---|
| IL-1β | F:
AGTTGACGGACCCCAAAAG | NM_008361.4 |
| R:
TTTGAAGCTGGATGCTCTCAT |
| IL-6 | F:
CCCCAATTTCCAATGCTCTCC | NM_031168.2 |
| R:
CGCACTAGGTTTGCCGAGT |
| iNOS | F:
CTCACCTACTTCCTGGACATTAC | NM_010927.4 |
| R:
CAATCTCTGCCTATCCGTCTC |
| IL-10 | F:
GCTCTTACTGACTGGCATGAG | NM_010548.2 |
| R:
CGCAGCTCTAGGAGCATGTG |
| FtH | F:
TGGAACTGCACAAACTGGCTACT | NM_010239.2 |
| R:
ATGGATTTCACCTGTTCACTCAGATAA |
| Fpn | F:
ATGGGAACTGTGGCCTTCAC | NM_016917.2 |
| R:
TCCAGGCATGAATACGGAGA |
| DMT1 | F:
TCTGGGCAGTGGGGATCCTG | NM_001146161.1 |
| R:
GACGAGCAGGGTGGGGATGA |
| TfR | F:
TCGTACAGCAGCGGAAGT | NM_001357298.1 |
| R:
TCTCCACGAGCGGAATACAG |
| COX2 | F:
CCTCGTCCAGATGCTA | NM_011198.4 |
| R:
CTCGGCTTCCAGTATTG |
| GPX4 | F:
CGCGATGATTGGCGCT | NM_001037741.4 |
| R:
CACACGAAACCCCTGTACTTATCC |
| SLC7A11 | F:
TGGCGGTGACCTTCTCTGA | NM_011990.2 |
| R:
ACAAAGATCGGGACTGCTAATGA |
| Sox9 | F:
CACACGTCAAGCGACCCATGAA | NM_011448.4 |
| R:
TCTTCTCGCTCTCGTTCAGCAG |
| Krt19 | F:
AATGGCGAGCTGGAGGTGAAGA | NM_001313963.1 |
| R:
CTTGGAGTTGTCAATGGTGGCAC |
| Vim | F:
CGGAAAGTGGAATCCTTGCAGG | NM_011701.4 |
| R:
AGCAGTGAGGTCAGGCTTGGAA |
| CD3e | F:
ATGCGGTGGAACACTTTCTGG | NM_007648.5 |
| R:
GCACGTCAACTCTACACTGGT |
| CD11b | F:
CCATGACCTTCCAAGAGAATGC | NM_001082960.1 |
| R:
ACCGGCTTGTGCTGTAGTC |
| Mrc1 | F:
CTCTGTTCAGCTATTGGACGC | NM_008625.2 |
| R:
TGGCACTCCCAAACATAATTTGA |
| α-SMA | F:
GTCCCAGACATCAGGGAGTAA | NM_007392.3 |
| R:
TCGGATACTTCAGCGTCAGGA |
| Col1a1 | F:
GCTCCTCTTAGGGGCCACT | NM_007742.4 |
| R:
CCACGTCTCACCATTGGGG |
| CTGF | F:
CAGAGTGGAGCGCCTGTT | NM_010217.2 |
| R:
GGATGCACTTTTTGCCCTTCT |
| Fibronectin-1 | F:
CGAAGAGCCCTTACAGTTCC | NM_001276408.1 |
| R:
CCGTGTAAGGGTCAAAGCAT |
| β-actin | F:
CCACCATGTACCCAGGCATT | NM_007393.5 |
| R:
AGGGTGTAAAACGCAGCTCA |
Western blot analysis
Total pancreatic protein was isolated using Cell
Lysis Buffer for Western and IP (cat. no. BL509A; Biosharp Life
Sciences). The total protein concentration was measured by BCA
Protein Assay kit (cat. no. KGP902; Nanjing KeyGen Biotech Co.,
Ltd.). Protein samples (25 µg/lane) were separated on 10%
gels using SDS-PAGE, and then transferred to PVDF membranes. At
room temperature, the membranes were blocked with 5% skimmed milk
for 1 h. The membranes were incubated overnight with the following
primary anti-bodies at 4°C: β-actin (1:10,000; cat. no. ET1702-52;
HuaBio); glutathione peroxidase 4 (GPX4; 1:1,000; cat. no.
ER1803-15; HuaBio), SLC7a11 (1:1,000; cat. no. HA600097; HuaBio)
and cytochrome c oxidase subunit II (COX2; 1:1,000; cat. no.
4842S; Cell Signaling Technology, Inc.). Next, the membranes were
incubated with HRP-linked goat anti-rabbit IgG (1:5,000; cat. no.
BL003A; Biosharp Life Sciences) at room temperature for 1 h. The
target proteins in the membranes were visualized by enhanced
chemiluminescence detection kit (cat. no. BL520A; Biosharp Life
Sciences). The band strength was analyzed using ImageJ software
(version 2.0; National Institutes of Health) and normalized to
β-actin protein intensity.
Statistical analysis
Results are shown as the mean ± standard error of
the mean. Statistical analysis was performed using GraphPad Prism
8.0 software (GraphPad Software, Inc.). Differences between the two
groups were compared using an unpaired two-tailed Student's t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Iron-overloaded mice have massive iron
deposition in the pancreas
The iron overload mouse model was constructed by
intraperitoneal injection of 120 mg/kg body weight of iron dextran
every other week for 12 weeks. Serum iron, transferrin saturation
and pancreatic tissue iron levels were all significantly
(P<0.01) elevated in the mice with iron dextran injection
(Fig. 1A-C). Prussian blue
staining revealed a large number of Prussian blue-positive spots,
hemosiderin, in the pancreas of the mice injected with iron dextran
(Fig. 1D). Massive iron
deposition was showed in the exocrine rather than the endocrine
pancreas. Moreover, the mRNA level of iron storing protein ferritin
H (FtH) was significantly (P<0.01) increased in the
pancreas of iron-treated mice, while those of iron transporting
membrane proteins divalent metal transporter 1 (DMT1),
ferroportin 1 (FPN) and transferrin receptor (TfR)
were all significantly (P<0.01) decreased (Fig. 1E-H). The protein level of iron
storing protein FtH was significantly (P<0.05) increased in the
pancreas of iron-treated mice, while those of iron transporting
membrane proteins DMT1, FPN, and iron binding receptor TfR were all
significantly (P<0.05) decreased (Fig. 1I). The results indicated that the
iron overload mouse model was successfully established and that
large amounts of iron were deposited in the pancreas.
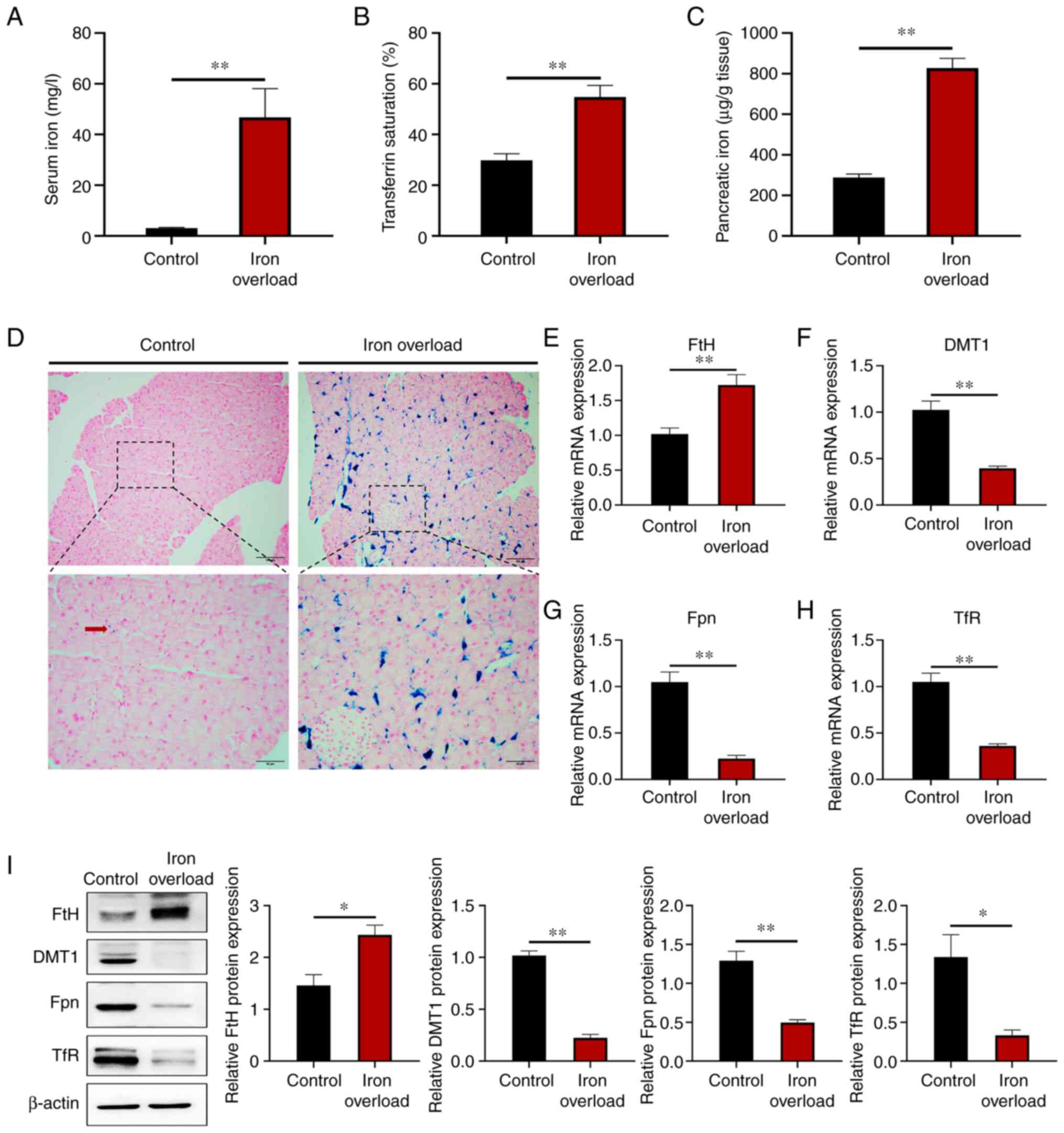 | Figure 1Iron-overloaded mice have massive
iron deposition in the pancreas. Male, 8-week-old, C57BL/6 mice
were injected intraperitoneally with 120 mg/kg body weight of iron
dextran every other week for 12 weeks. (A) Iron content in the
serum. (B) Transferrin saturation in the serum. (C) Pancreatic iron
content. (D) Prussian blue staining in the pancreas. Scale bars:
Upper, 100 µm; lower, 50 µm. (E-H) Relative gene
expression of (E) FtH, (F) DMT1, (G) FPN and
(H) TfR in pancreatic tissue. (I) FtH, DMT1, Fpn and TfR
protein expression levels were detected by western blotting in the
pancreas. Values are expressed as the mean ± standard error of the
mean. Differences between the two groups were compared using an
unpaired two-tailed Student's t-test. *P<0.05;
**P<0.01 (n=6 per group). FtH, ferritin H; DMT1,
divalent metal transporter 1; FPN, ferroportin 1; TfR, transferrin
receptor. |
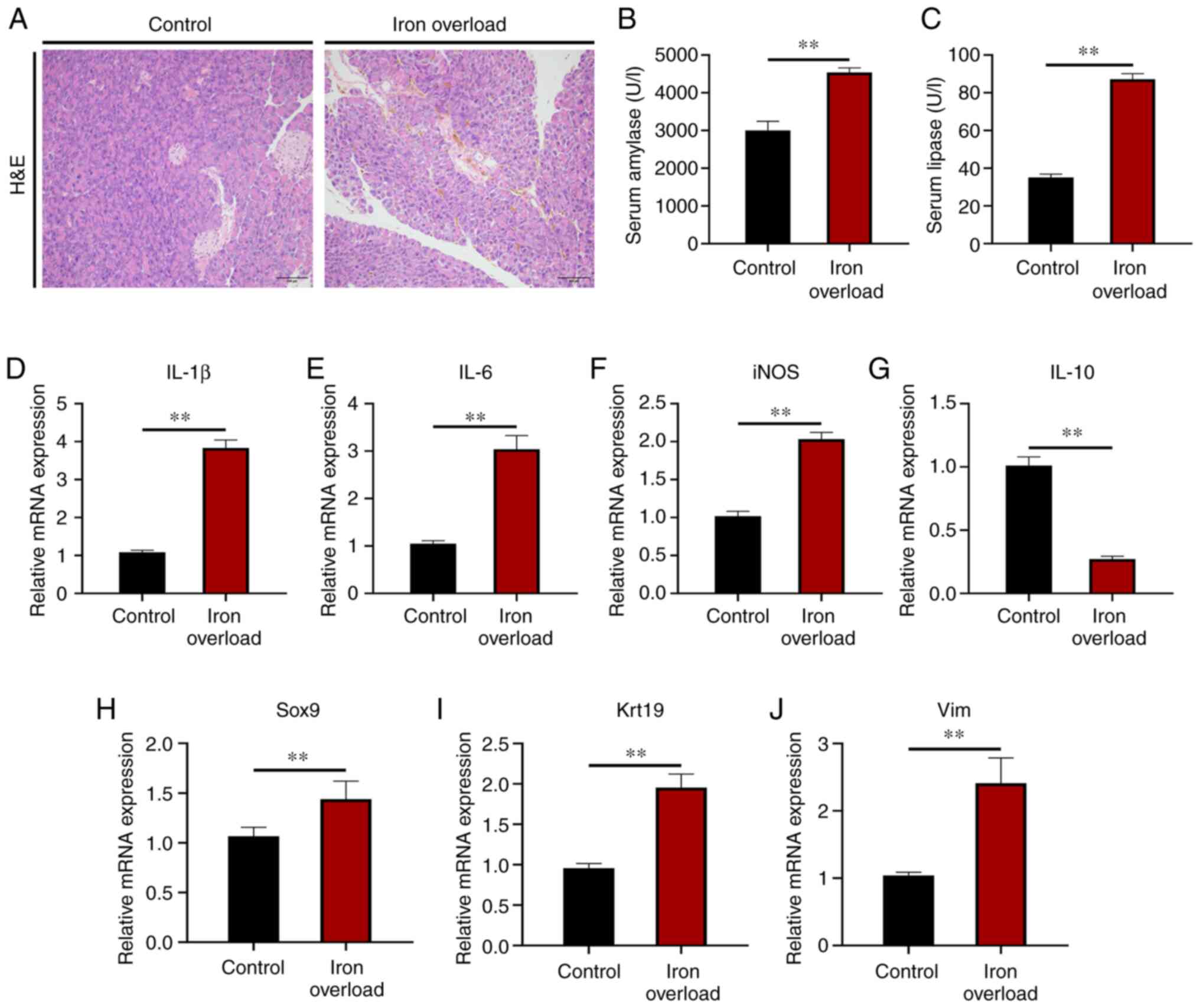
Iron-overloaded mice develop mild chronic
pancreatitis
In view of the obvious pancreatic iron deposition in
iron-overloaded mice, the effect of iron overload on the morphology
and function of the pancreas was then analyzed. H&E staining
showed that the acinar cells of the pancreas in iron-overloaded
mice were atrophied and the area of intercellular substance was
larger compared with those of control mice (Fig. 2A), which suggested mild pancreatic
injury existed in the iron-overloaded mice. Serum amylase and
lipase activities were both significantly (P<0.01) increased in
the iron-overloaded mice compared with those of the control mice
(Fig. 2B and C). Moreover,
compared with those of the control group, the mRNA expression
levels of pro-inflammatory cytokines, including interleukin
(IL)-1β, IL-6 and inducible nitric oxide
synthase, were significantly (P<0.01) increased in the pancreas
of iron-overloaded animals (Fig.
2D-F). Accordingly, the transcript of anti-inflammatory
cytokine IL-10 was significantly (P<0.01) decreased in
the pancreas of iron-overloaded mice (Fig. 2G). Furthermore, the mRNA levels of
SRY-related high-mobility-group-box gene 9 (a molecular marker of
acinar to ductal metaplasia), keratin 19 (a molecular marker of
ductal lesions) and vimentin (a molecular marker of stromal
response) were all significantly (P<0.01) elevated by iron
injection (Fig. 2H-J). The
results indicated that iron overload induced mild CP and resulted
in pancreatic injury.
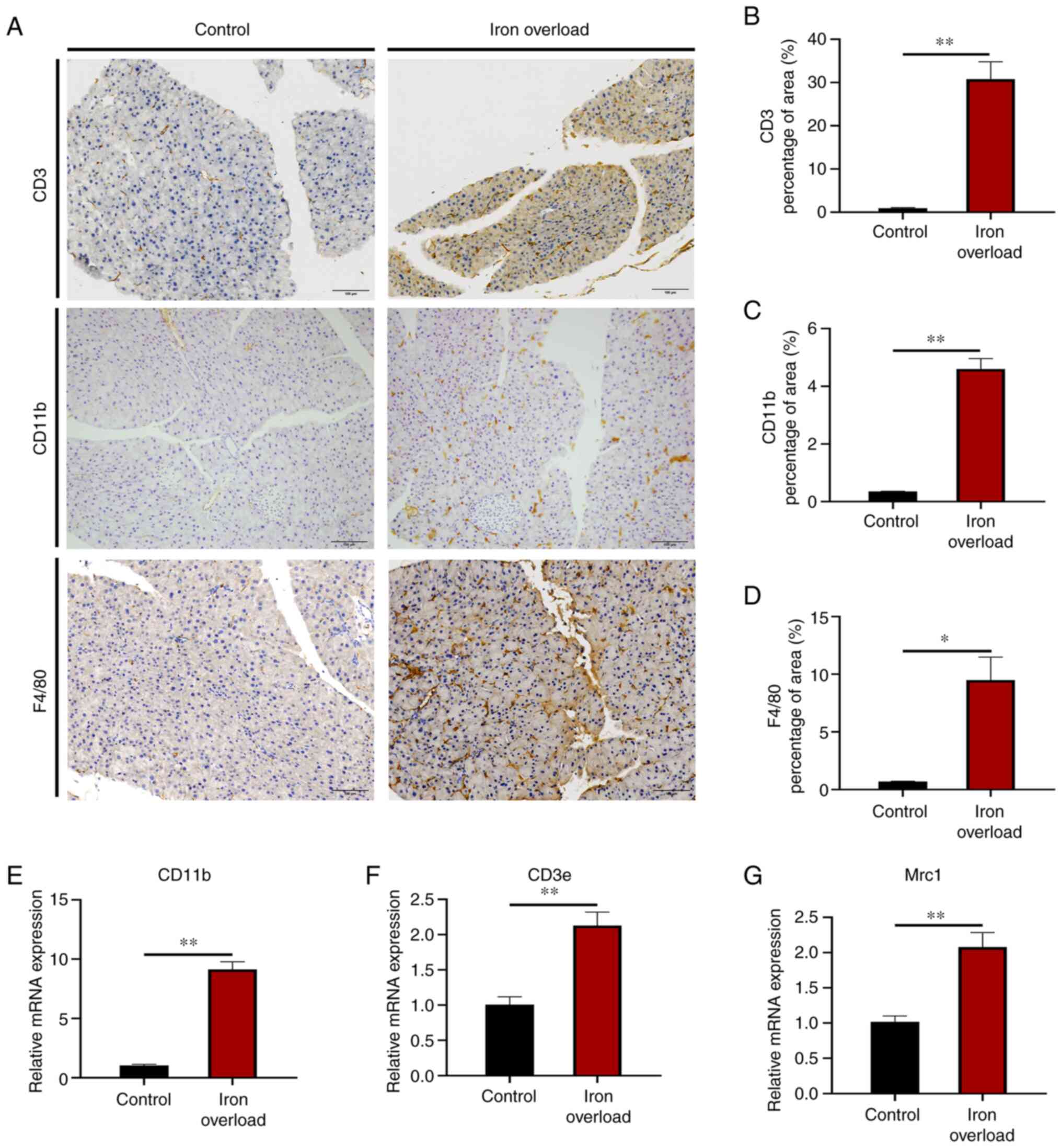
Iron-overloaded mice show increased
immunocyte infiltration in the pancreas
Histological evaluation of the pancreatic lesions
showed that increased levels of lymphocytes (anti-CD3), neutrophils
(anti-CD11b) and macrophages (anti-F4/80) were observed in the
pancreas of iron-overloaded mice compared with those of control
mice (Fig. 3A). The quantitative
positive areas of CD3 (P<0.01), CD11b (P<0.01) and F4/80
(P<0.05) cells showed a significant increase in the pancreas of
the iron-overloaded mice compared with those in the control mice
(Fig. 3B-D). Furthermore, the
mRNA expression of CD11b, CD3e (lymphocyte marker)
and mannose receptor C type 1 (macrophage marker) was significantly
(P<0.01) increased relative to the control group (Fig. 3E-G). The results indicated that
pancreatic accumulation of immunocytes, including lymphocytes,
neutrophils and macrophages, was present in iron-overloaded mice,
which was characteristic of CP.
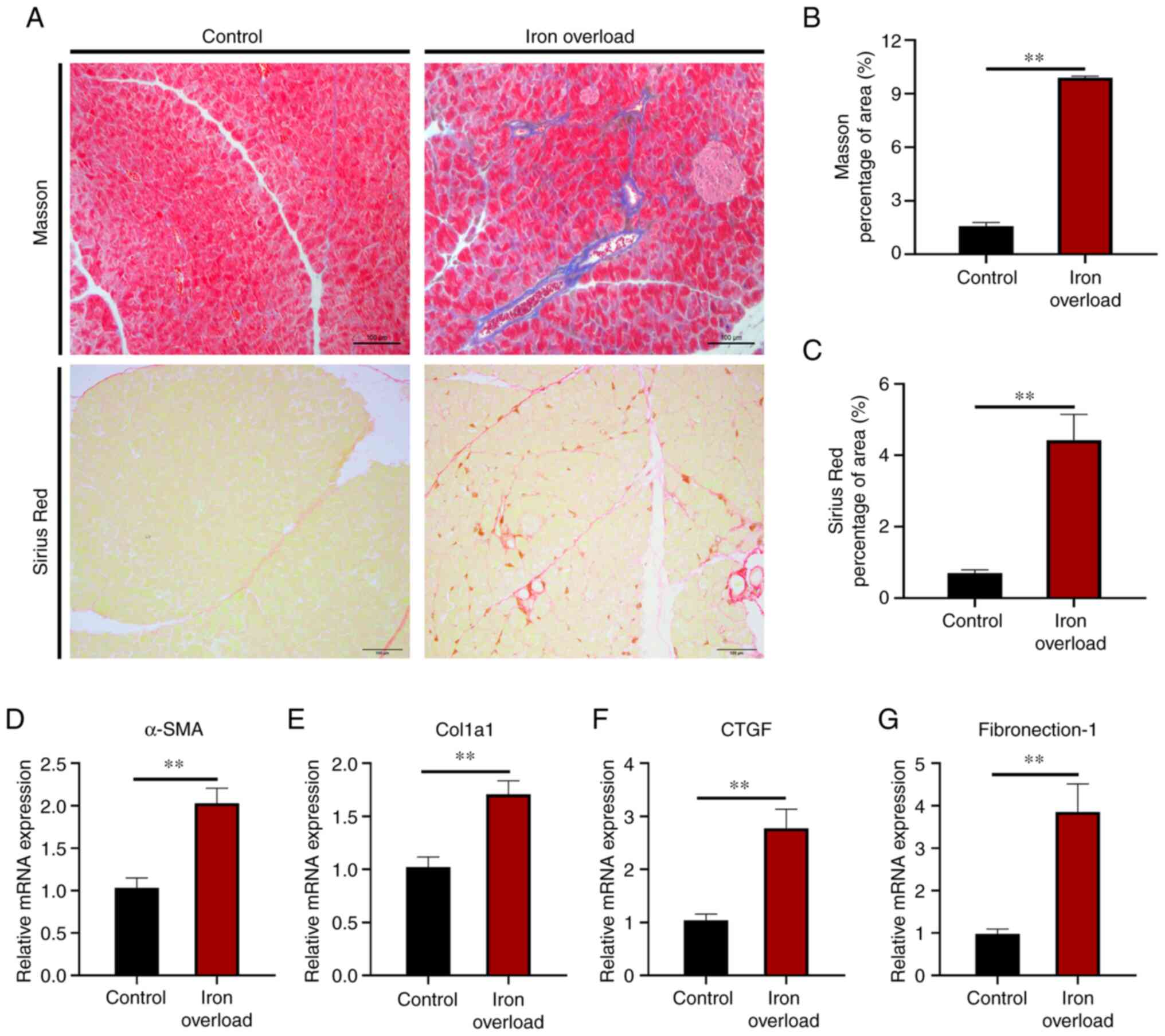
Iron-overloaded mice display pancreatic
fibrosis
Since pancreatitis symptoms were shown in the
iron-overloaded mice, Masson's trichrome and Sirius red staining
were next performed to assess collagen accumulation in the
pancreatic tissue sections to examine the degree of pancreatic
fibrosis. The analysis showed that iron overload induced
perivascular collagen accumulation (Fig. 4A). Semi-quantitative morphometric
analysis demonstrated that the collagen-positive area of tissue
sections was significantly (P<0.01) increased in the pancreas of
iron-overloaded mice compared with that of the control group
(Fig. 4B and C). Additionally,
analysis of transcripts of pancreatic fibrosis markers, such as
α-smooth muscle actin (α-SMA), collagen type I α1,
connective tissue growth factor and fibronectin-1, showed a
significant (P<0.01) increase in their levels in the pancreas
tissue of iron-overloaded mice (Fig.
4D-G). It was observed that the development of atrophy,
inflammatory cell infiltration and fibrosis in the pancreatic
tissue of iron-overloaded mice was significantly higher than that
of control mice. Statistical comparisons between groups are
summarized in Table II.
| Table IIComparison of mean histopathological
scores of the groups (n=5). |
Table II
Comparison of mean histopathological
scores of the groups (n=5).
| Histopathological
score | Groups |
|---|
| Control | Iron overload |
|---|
| 0, n (%) | 3 (60) | - |
| 1, n (%) | 2 (40) | - |
| 2, n (%) | - | 1 (20) |
| 3, n (%) | - | 1 (20) |
| 4, n (%) | - | 1 (20) |
| 5, n (%) | - | 1 (20) |
| 6, n (%) | - | 1 (20) |
| Mean ± SEM | 0.4±0.490 | 4±1.414 |
| P-value (iron
overload) | | 0.0013 |
Iron-overloaded mice exhibit increased
oxidative stress and ferroptosis in the pancreas
Oxidative stress is a common pathogenesis of a
number of chronic diseases, and it is well known that iron overload
affects the redox state. Compared with that of the control group,
the MDA level was increased by 1.12-fold (P<0.01) in the
pancreas of iron-overloaded mice (Fig. 5A). However, the injection of iron
dextran led to a reduction of SOD activity by 52% (P<0.01) and
GSH-PX activity by 37% (P<0.01) in the pancreas (Fig. 5B and C). This suggested that
excess iron could elevate the pancreatic oxidative stress of
pancreatic acinar cells in mice. Moreover, the injection of iron
dextran significantly (P<0.05) promoted COX2 (a putative
molecular marker of ferroptosis) protein expression, but inhibited
GPX4 and SLC7A11 protein expression in the pancreas of
iron-overloaded mice compared with the control group (Fig. 5D). The mRNA level of COX2
was also significantly (P<0.01) increased, and GPX4 and
SLC7A11 were significantly (P<0.01) decreased in the
pancreas of iron-overloaded mice compared with the levels in
control mice (Fig. 5E-G). The
results indicated that iron overload induced ferroptosis.
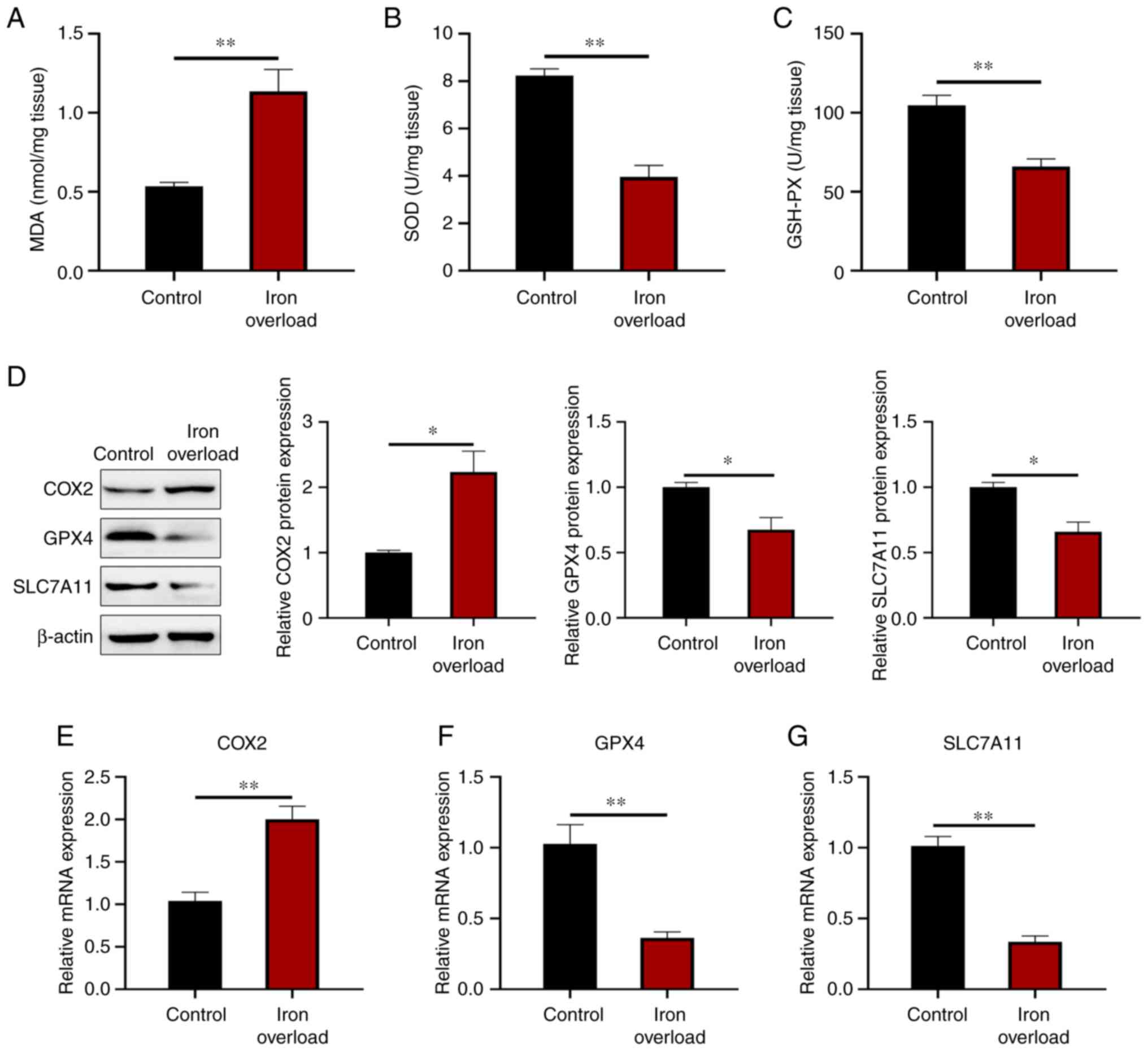 | Figure 5Iron-overloaded mice exhibit
increased oxidative stress and ferroptosis in the pancreas. (A) MDA
content, and (B) SOD and (C) GSH-PX activity in the pancreas. (D)
COX2, GPX4 and SLC7A11 protein expression levels were detected by
western blotting in the pancreas. (E-G) Relative gene expression of
pancreatic fibrosis markers, including (E) COX2, (F)
GPX4 and (G) SLC7A11 in the pancreas. Values are
expressed as the mean ± standard error of the mean. Differences
between the two groups were compared by an unpaired two-tailed
Student's t-test. *P<0.05; **P<0.01
(n=6 per group). MDA, malondialdehyde; SOD, superoxide dismutase;
GSH-PX, glutathione peroxidase; COX2, cytochrome c oxidase
subunit II; GPX4, glutathione peroxidase 4; SCL7A11, solute carrier
family 7 member 11. |
Discussion
Genetic mutant mouse models, such as hepcidin
knockout mice and BMP6 knockout mice, were previously found to have
a large amount of iron deposition in the pancreas, while
diet-induced iron overload did not lead to iron accumulation in the
pancreas (24,25). The present study identified that
the long-term injection of iron dextran caused iron overload in the
mouse pancreas and induced CP. The injection animal model was
adopted instead of the knockout model and the oral animal model.
Although it could not better reveal the pathogenesis of iron
overload syndrome, it could simulate the phenomenon of iron
overload caused by disease-dependent blood transfusion and explore
the influence of acquired iron overload on the development of
pancreatic diseases.
Iron dextran, currently one of the most widely used
iron preparations in livestock production, has previously been used
to establish an iron overload mouse model by intraperitoneal
injection with concentrations of 100-300 mg/kg (30,31). We previously established an iron
overload mouse model by injecting 120 mg/kg of iron dextran
intraperitoneally every other week for 12 weeks. A large amount of
iron deposition was found, as well as obvious lipid peroxidation
and ferroptosis in the liver of iron overload mice, and the iron
metabolism of the body was seriously unbalanced (30). In the present study,
intraperitoneal injection of iron dextran resulted in circulatory
iron overload with significantly increased serum iron and
transferrin saturation, and iron accumulation in the pancreatic
tissues of the mice. Gene expression of iron transporting proteins
such as DMT1, fpn1 and TfR1 was significantly reduced, whereas
levels of iron storing proteins such as FtH were significantly
elevated in the pancreas, which was similar to the results for
intestinal iron overload (32).
Previous studies demonstrated that iron accumulation was mainly
observed in exocrine pancreatic acini in hepcidin knockout mice and
Bmp6 knockout mice, but not in pancreatic islets. Thus, iron
deposition in both congenital and acquired iron overload occurs at
the same location, in the acini of the exocrine glands of the
pancreas (24,25). These results suggested that the
present non-hereditary iron overload model of mice was successfully
established, and massive iron deposition was observed in the
pancreas.
Previous studies have found that iron overload can
lead to the atrophy and dysfunction of organs such as the liver,
heart, muscles or brain (33,34). In the present study, iron
accumulation in the pancreas may be associated with pancreatic
dysfunction. The most common disease of the pancreatic exocrine
disorders is pancreatitis, which mainly includes CP and AP.
Clinically, CP continuously damages pancreatic endocrine and
exocrine tissues due to repeated episodes of AP and chronic
inflammation (35). In the later
stage of CP, pain, sclerosis, calcification, diabetes and/or
lipo-dysentery are manifested (36). The pathological features of CP are
diverse, and the most common include acinar atrophy, immune
infiltration, fibrosis, ductal irregularity, stenosis and
dilatation (37). Hepcidin
knockout mice at 6 and 12 months of age were found to exhibit a
significantly increased serum lipase level, a reduced pancreatic
acinar cell content and a large amount of macrophage infiltration
compared with control mice and iron-rich diet mice (24). The present study demonstrated that
iron overload damaged pancreatic acinar cells, enlarged
intercellular spaces and caused pancreatic ductal lesions. The
pancreases of iron-overloaded mice secreted large amounts of
amylase and lipase into the blood, and produced a large number of
pro-inflammatory factors. Therefore, mild CP was present in the
pancreas of iron-overloaded mice. Since mild CP was found in the
mice with iron overload during the late sample detection, a
positive control group was not set up in the early experimental
design. In addition, CP is regulated by the release of a variety of
proinflammatory and anti-inflammatory cytokines and chemokines
(38). At the same time, these
inflammatory signals recruit granulocytes (neutrophils and
eosinophils), monocytes, macrophages and lymphocytes to regulate
the development of CP (39).
Neutrophils are traditionally considered as the
first line of defense against foreign microorganisms in the innate
immune system, with limited proinflammatory functions (40). Although there is less infiltration
of neutrophils than macrophages in the development of CP, they are
associated with disease progression and disease symptoms in CP
(41). Neutrophils can activate
the secretion of inflammatory cytokines by various immune cells and
stromal cells, leading to the aggravation of inflammation (42). Unlike classical activation of
macrophages (M1) during AP, it has been shown that M2-like
macrophages predominate in CP (39). GPX4 knockout (a mouse model of
ferroptosis) promoted macrophage infiltration and activation
(43). The present results
confirmed that iron overload promoted the infiltration of
neutrophils in the pancreas of mice, and then increased the
inflammatory response of the pancreas. Although neutrophils and
monocytes/macrophages have been recognized as the main acting
leukocyte populations of the inflamed pancreas, a local imbalance
of T cells at the site of inflammation and in the circulation has
also been observed in pancreatitis (39). A study showed that massive
infiltration of mouse pancreatic neutrophils and macrophages in
hepcidin knockout mice induced pancreatitis, whereas
CD3+ T cells showed little change (24). In contrast with this, in the
present study, iron-overloaded mice showed increased infiltration
of CD3+ T cells, macrophages and neutrophils.
Cytokines and chemokines induced by immune cells can
activate pancreatic stellate cells and accelerate disease
progression (38). Pancreatic
stellate cells are the major contributing cells in the progression
of pancreatic fibrosis (44).
Pancreatic fibrosis is one of the important hallmarks of
pancreatitis and pancreatic cancer (45). One study demonstrated that aging
Bmp6 knockout mice developed pancreatic fibrosis with collagen
distributed in the interlobular, periacinar and peripancreatic
ducts (25). The present results
showed that iron overload promoted the progression of pancreatic
fibrosis in mice. Studies have confirmed that activated pancreatic
stellate cells (α-SMA-positive cells) can promote pancreatic
fibrosis by secreting extracellular matrix components, such as
collagen and fibronectin, during the progression of pancreatic
fibrosis (46). In the present
study, the expression of α-SMA, collagen and fibronectin was
increased in the pancreases of iron-overloaded mice. These results
indicated that iron overload could activate pancreatic stellate
cells and promote the progression of pancreatitis.
Iron overload can generate reactive oxygen species,
leading to dysfunction of mitochondria and other organelles, lipid
peroxidation, cell damage and death (47). Ferroptosis, an iron-dependent
non-apoptotic regulated form of cell death, exhibits unique
features that distinguish it from other types of cell death such as
apoptosis, autophagy and necrosis (48). Ferroptosis has two major typical
features, namely, the accumulation of Fe2+ and an
increase in lipid peroxidation (49). Increased peroxides are usually
characterized by increased MDA content, accompanied by changes in
markers of ferroptosis such as SLC7A11, GPX4 and COX2. When iron
overload causes oxidative stress, it will lead to changes in MDA
content, and SOD and GSH-PX enzyme activity (50). MDA is one of the end products of
the lipid peroxidation of polyunsaturated fatty acids (51), which can be produced by either
enzymatic pathways or non-enzymatic processes (52). MDA is not only a biomarker of
oxidative stress, but also a bioactive compound with multiple
biological effects (53). SOD is
an antioxidant defense enzyme that plays a crucial role in the
balance between theoxidation and antioxidation of the body
(54). SOD destroys superoxide
radicals by dismutase to produce hydrogen peroxide, which is
continuously reduced by catalase or GSH-PX activity (55). Thus, SOD and GSH-PX are able to
protect cells from injury. When ferroptosis occurs in the organism,
SLC7A11 expression on the cell membrane is suppressed (56), and cells then take up less
cysteine. GSH is continuously consumed and synthesis cannot
continue. Consequently, the synthesis of GPX4 is blocked, which
disrupts the ability of the cell to scavenge reactive oxygen
species (ROS), leading to ROS accumulation and triggering
ferroptosis (57). It has been
found that persistent inflammation and oxidative stress are
important mediators of cancer development and progression (58). In the present study, the
accumulation of excess lipid peroxides induced by iron overload
activated ferroptosis in the pancreas. Ferroptosis has been
reported to be involved in numerous pathological processes,
including neurotoxicity (59),
acute renal failure (60),
hepatotoxicity (61) and
pancreatic cancer (43). Studies
have confirmed a key role for ferroptosis in AP, and addition of
the ferroptosis inhibitor liproxstatin-1 or upregulation of GPX4
slowed AP and acute kidney injury in rats (62,63). These findings illustrate the
possibility that the iron overload-induced ferroptosis in this
experimental model leads to spontaneous formation of CP in mice,
resulting in pancreatic acinar cell death and dysfunction. However,
further validation is required to reach this conclusion. Meanwhile,
ferroptosis may become a new target for the study of pancreatitis,
and provide a new idea for the clinical research of pancreatitis.
However, little is known about the exact mechanism of how iron
overload affects the progression of pancreatitis, and further
studies of the interaction between iron and pancreatic function are
also needed to clarify the role of iron in the pancreas.
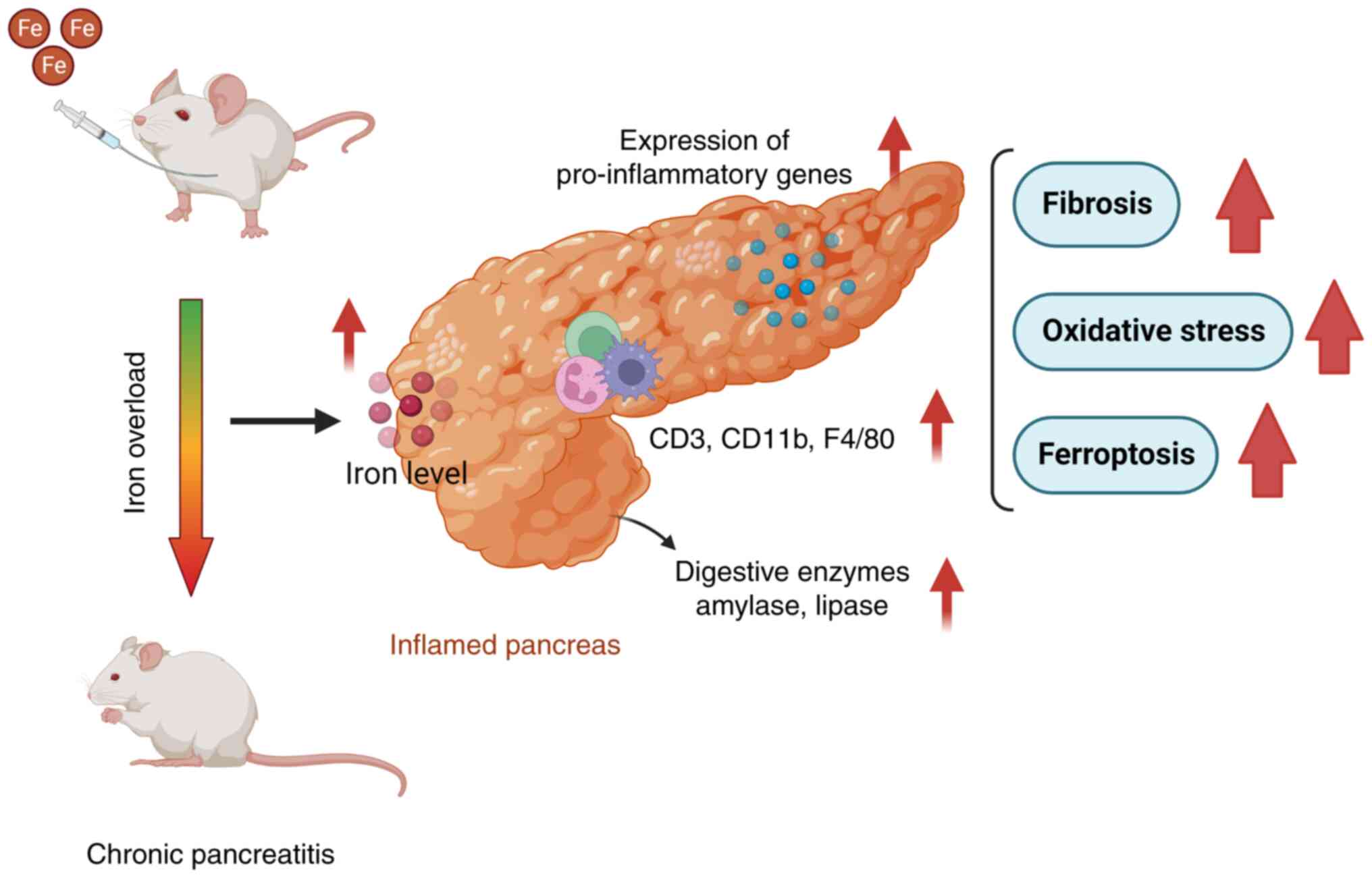
In summary, the present study provides evidence that
secondary iron overload induced by multiple intraperitoneal
injections of iron dextran resulted in massive iron deposition in
the pancreas of mice. Iron-overloaded mice developed CP with
elevated levels of serum amylase and lipase, upregulated gene
expression of pro-inflammatory factors and increased infiltration
of immune cells (Fig. 6).
Moreover, iron overload also led to pancreatic fibrosis, oxidative
stress and ferroptosis. This study suggests that secondary iron
overload is a risk factor for pancreatitis and highlights the
importance of iron in maintaining the normal functions of the
pancreas.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CT, JZ and QX performed the experiments. CT, HY and
HD analyzed the data. JZ conducted the literature search and
analyzed the data. HY and HD designed the study and confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
All experiments were reviewed and approved by the
Committee of Laboratory Animal Care and Animal Research Ethics
Committee of Zhejiang University (approval no. 20077).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
HH
|
hereditary haemochromatosis
|
|
AP
|
acute pancreatitis
|
|
CP
|
chronic pancreatitis
|
|
MDA
|
malondialdehyde
|
|
SOD
|
superoxide dismutase
|
|
GSH-PX
|
glutathione peroxidase
|
|
FtH
|
ferritin H
|
|
DMT1
|
divalent metal transporter 1
|
|
FPN
|
ferroportin 1
|
|
TfR
|
transferrin receptor
|
|
IL
|
interleukin
|
|
α-SMA
|
α-smooth muscle actin
|
|
COX2
|
cytochrome c oxidase subunit
II
|
|
GPX4
|
glutathione peroxidase 4
|
Acknowledgments
Not applicable.
Funding
This study was supported by the Natural Science Foundation of
Zhejiang Province of China (grant no. LZ20C170004), the National
Natural Science Foundation of China (grant no. 31872363), the 'Ten
Thousand Plan' Innovation Leader of Zhejiang Province of China
(grant no. 2020R52007) and the Fundamental Research Funds for the
Central Universities (grant no. 226-2022-00023).
References
|
1
|
Sarkar J, Potdar AA and Saidel GM:
Whole-body iron transport and metabolism: Mechanistic, multi-scale
model to improve treatment of anemia in chronic kidney disease.
PLoS Comput Biol. 14:e10060602018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Puig S, Ramos-Alonso L, Romero AM and
Martinez-Pastor MT: The elemental role of iron in DNA synthesis and
repair. Metallomics. 9:1483–1500. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ganz T and Nemeth E: Iron homeostasis in
host defence and inflammation. Nat Rev Immunol. 15:500–510. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hentze MW, Muckenthaler MU, Galy B and
Camaschella C: Two to tango: Regulation of mammalian iron
metabolism. Cell. 142:24–38. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ganz T: Systemic iron homeostasis. Physiol
Rev. 93:1721–1741. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fernandez RJM, Moreno-Navarrete JM and
Manco M: Iron influences on the Gut-Brain axis and development of
type 2 diabetes. Crit Rev Food Sci Nutr. 59:443–449. 2019.
View Article : Google Scholar
|
|
7
|
Zafon C, Lecube A and Simo R: Iron in
obesity. An ancient micronutrient for a modern disease. Obes Rev.
11:322–328. 2010. View Article : Google Scholar
|
|
8
|
Mayneris-Perxachs J, Cardellini M, Hoyles
L, Latorre J, Davato F, Moreno-Navarrete JM, Arnoriaga-Rodriguez M,
Serino M, Abbott J, Barton RH, et al: Iron status influences
non-alcoholic fatty liver disease in obesity through the gut
microbiome. Microbiome. 9:1042021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu S: Iron and atherosclerosis: The link
revisited. Trends Mol Med. 25:659–661. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fleming RE and Ponka P: Iron overload in
human disease. N Engl J Med. 366:348–359. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fernandez-Real JM and Manco M: Effects of
iron overload on chronic metabolic diseases. Lancet Diabetes
Endocrinol. 2:513–526. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Brissot P, Pietrangelo A, Adams PC, de
Graaff B, McLaren CE and Loréal O: Haemochromatosis. Nat Rev Dis
Primers. 4:180162018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bottomley SS: Secondary iron overload
disorders. Semin Hematol. 35:77–86. 1998.PubMed/NCBI
|
|
14
|
Che J, Yang J, Zhao B, Zhang G, Wang L,
Peng S and Shang P: The effect of abnormal iron metabolism on
osteoporosis. Biol Trace Elem Res. 195:353–365. 2020. View Article : Google Scholar
|
|
15
|
Hatunic M, Finucane FM, Brennan AM, Norris
S, Pacini G and Nolan JJ: Effect of iron overload on glucose
metabolism in patients with hereditary hemochromatosis. Metabolism.
59:380–384. 2010. View Article : Google Scholar
|
|
16
|
Roggero S, Quarello P, Vinciguerra T,
Longo F, Piga A and Ramenghi U: Severe iron overload in
blackfan-diamond anemia: A case-control study. Am J Hematol.
84:729–732. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vogiatzi MG, Macklin EA, Trachtenberg FL,
Fung EB, Cheung AM, Vichinsky E, Olivieri N, Kirby M, Kwiatkowski
JL, Cunningham M, et al: Differences in the prevalence of growth,
endocrine and vitamin D abnormalities among the various
thalassaemia syndromes in North America. Br J Haematol.
146:546–556. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Berdoukas V, Nord A, Carson S, Puliyel M,
Hofstra T, Wood J and Coates TD: Tissue iron evaluation in
chronically transfused children shows significant levels of iron
loading at a very young age. Am J Hematol. 88:E283–E285. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee PJ and Papachristou GI: New insights
into acute pancreatitis. Nat Rev Gastroenterol Hepatol. 16:479–496.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lugea A, Waldron RT, Mareninova OA,
Shalbueva N, Deng N, Su HY, Thomas DD, Jones EK, Messenger SW, Yang
J, et al: Human pancreatic acinar cells: Proteomic
characterization, physiologic responses, and organellar disorders
in ex vivo pancreatitis. Am J Pathol. 187:2726–2743. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Singh VK, Yadav D and Garg PK: Diagnosis
and management of chronic pancreatitis: A review:. JAMA.
322:2422–2434. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Whitcomb DC, Frulloni L, Garg P, Greer JB,
Schneider A, Yadav D and Shimosegawa T: Chronic pancreatitis: An
international draft consensus proposal for a new mechanistic
definition. Pancreatology. 16:218–224. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kimita W and Petrov MS: Iron metabolism
and the exocrine pancreas. Clin Chim Acta. 511:167–176. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lunova M, Schwarz P, Nuraldeen R, Levada
K, Kuscuoglu D, Stützle M, Spasić MV, Haybaeck J, Ruchala P, Jirsa
M, et al: Hepcidin knockout mice spontaneously develop chronic
pancreatitis owing to cytoplasmic iron overload in acinar cells. J
Pathol. 241:104–114. 2017. View Article : Google Scholar
|
|
25
|
Pauk M, Kufner V, Rumenovic V, Dumic-Cule
I, Farkas V, Milosevic M, Bordukalo-Niksic T and Vukicevic S: Iron
overload in aging Bmp6(/) mice induces exocrine pancreatic injury
and fibrosis due to acinar cell loss. Int J Mol Med. 47:602021.
View Article : Google Scholar
|
|
26
|
Pelucchi S, Ravasi G and Piperno A:
Ceruloplasmin variants might have different effects in different
iron overload disorders. J Hepatol. 75:1003–1004. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ganz T: Hepcidin and iron regulation, 10
years later. Blood. 117:4425–4433. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yildirim M, Kaplan M, Duzenli T, Tanoglu
A, Kucukodaci Z, Tastan YO, Guney BC and Serindag Z: Pentoxifylline
has favorable preventive effects on experimental chronic
pancreatitis model. Scand J Gastroenterol. 55:236–241. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
30
|
Ma W, Jia L, Xiong Q and Du H: Iron
overload protects from obesity by ferroptosis. Foods. 10:17872021.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xiong H, Zhang C, Han L, Xu T, Saeed K,
Han J, Liu J, Klaassen CD, Gonzalez FJ, Lu Y and Zhang Y:
Suppressed farnesoid X receptor by iron overload in mice and humans
potentiates iron-induced hepatotoxicity. Hepatology. 76:387–403.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang DL, Hughes RM, Ollivierre-Wilson H,
Ghosh MC and Rouault TA: A ferroportin transcript that lacks an
iron-responsive element enables duodenal and erythroid precursor
cells to evade translational repression. Cell Metab. 9:461–473.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Petrillo S, Manco M, Altruda F, Fagoonee S
and Tolosano E: Liver sinusoidal endothelial cells at the crossroad
of iron overload and liver fibrosis. Antioxid Redox Signal.
35:474–486. 2021. View Article : Google Scholar
|
|
34
|
Martin D, Nay K, Robin F, Rebillard A,
Orfila L, Martin B, Leroyer P, Guggenbuhl P, Dufresne S, Noirez P,
et al: Oxidative and glycolytic skeletal muscles deploy protective
mechanisms to avoid atrophy under pathophysiological iron overload.
J Cachexia Sarcopenia Muscle. 13:1250–1261. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lohr JM, Dominguez-Munoz E, Rosendahl J,
Besselink M, Mayerle J, Lerch MM, Haas S, Akisik F, Kartalis N,
Iglesias-Garcia J, et al: United European gastroenterology
evidence-based guidelines for the diagnosis and therapy of chronic
pancreatitis (HaPanEU). United European Gastroenterol J. 5:153–199.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Beyer G, Habtezion A, Werner J, Lerch MM
and Mayerle J: Chronic pancreatitis. Lancet. 396:499–512. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kleeff J, Whitcomb DC, Shimosegawa T,
Esposito I, Lerch MM, Gress T, Mayerle J, Drewes AM, Rebours V,
Akisik F, et al: Chronic pancreatitis. Nat Rev Dis Primers.
3:170602017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Manohar M, Verma AK, Venkateshaiah SU,
Sanders NL and Mishra A: Pathogenic mechanisms of pancreatitis.
World J Gastrointest Pharmacol Ther. 8:10–25. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xue J, Sharma V and Habtezion A: Immune
cells and immune-based therapy in pancreatitis. Immunol Res.
58:378–386. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kolaczkowska E and Kubes P: Neutrophil
recruitment and function in health and inflammation. Nat Rev
Immunol. 13:159–175. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Grunwald B, Harant V, Schaten S,
Fruhschutz M, Spallek R, Hochst B, Stutzer K, Berchtold S, Erkan M,
Prokopchuk O, et al: Pancreatic premalignant lesions secrete tissue
inhibitor of metalloproteinases-1, which activates hepatic stellate
cells via CD63 signaling to create a premetastatic niche in the
liver. Gastroenterology. 151:1011–1024.e1017. 2016. View Article : Google Scholar
|
|
42
|
Chin AC and Parkos CA: Neutrophil
transepithelial migration and epithelial barrier function in IBD:
potential targets for inhibiting neutrophil trafficking. Ann N Y
Acad Sci. 1072:276–287. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dai E, Han L, Liu J, Xie Y, Zeh HJ, Kang
R, Bai L and Tang D: Ferroptotic damage promotes pancreatic
tumorigenesis through a TMEM173/STING -dependent DNA sensor
pathway. Nat Commun. 11:63392020. View Article : Google Scholar
|
|
44
|
Han L, Ma J, Duan W, Zhang L, Yu S, Xu Q,
Lei J, Li X, Wang Z, Wu Z, et al: Pancreatic stellate cells
contribute pancreatic cancer pain via activation of sHH signaling
pathway. Oncotarget. 7:18146–18158. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kirkegård J, Mortensen FV and
Cronin-Fenton D: Chronic pancreatitis and pancreatic cancer risk: A
systematic review and meta-analysis. Am J Gastroenterol.
112:1366–1372. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen B, Li J, Fellows GF, Sun Z and Wang
R: Maintaining human fetal pancreatic stellate cell function and
proliferation require beta1 integrin and collagen I matrix
interactions. Oncotarget. 6:14045–14059. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tang D and Kroemer G: Ferroptosis. Curr
Biol. 30:R1292–R1297. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xie Y, Hou W, Song X, Yu Y, Huang J, Sun
X, Kang R and Tang D: Ferroptosis: Process and function. Cell Death
Differ. 23:369–379. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang C, Zheng L, Liu S, Guo X, Qu Y, Gao
M, Cui X and Yang Y: A novel acidic polysaccharide from the residue
of Panax notoginseng and its hepatoprotective effect on alcoholic
liver damage in mice. Int J Biol Macromol. 149:1084–1097. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ayala A, Muñoz MF and Argüelles S: Lipid
peroxidation: Production, metabolism, and signaling mechanisms of
malondialdehyde and 4-hydroxy-2-nonenal. Oxid Med Cell Longev.
2014:3604382014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jové M, Mota-Martorell N, Pradas I,
Martín-Gari M, Ayala V and Pamplona R: The advanced lipoxidation
end-product malondialdehyde-lysine in aging and longevity.
Antioxidants (Basel). 9:11322020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tsikas D: Assessment of lipid peroxidation
by measuring malondialdehyde (MDA) and relatives in biological
samples: Analytical and biological challenges. Anal Biochem.
524:13–30. 2017. View Article : Google Scholar
|
|
54
|
Olsvik PA, Kristensen T, Waagbø R,
Rosseland BO, Tollefsen KE, Baeverfjord G and Berntssen MHG: MRNA
expression of antioxidant enzymes (SOD, CAT and GSH-Px) and lipid
peroxidative stress in liver of Atlantic salmon (Salmo salar)
exposed to hyperoxic water during smoltification. Comp Biochem
Physiol C Toxicol Pharmaco. 141:314–323. 2005. View Article : Google Scholar
|
|
55
|
Sefi M, Fetoui H, Lachkar N, Tahraoui A,
Lyoussi B, Boudawara T and Zeghal N: Centaurium erythrea
(Gentianaceae) leaf extract alleviates streptozotocin-induced
oxidative stress and β-cell damage in rat pancreas. J
Ethnopharmacol. 135:243–250. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Dolma S, Lessnick SL, Hahn WC and
Stockwell BR: Identification of genotype-selective antitumor agents
using synthetic lethal chemical screening in engineered human tumor
cells. Cancer Cell. 3:285–296. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Forcina GC and Dixon SJ: GPX4 at the
crossroads of lipid homeostasis and ferroptosis. Proteomics.
19:e18003112019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yao W, Qiu HM, Cheong KL and Zhong S:
Advances in anti-cancer effects and underlying mechanisms of marine
algae polysaccharides. Int J Biol Macromol. 221:472–485. 2002.
View Article : Google Scholar
|
|
59
|
Xia Y, Sun X, Luo Y and Stary CM:
Ferroptosis contributes to isoflurane neurotoxicity. Front Mol
Neurosci. 11:4862018. View Article : Google Scholar
|
|
60
|
Belavgeni A, Meyer C, Stumpf J, Hugo C and
Linkermann A: Ferroptosis and necroptosis in the kidney. Cell Chem
Biol. 27:448–462. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yamada N, Karasawa T, Kimura H, Watanabe
S, Komada T, Kamata R, Sampilvanjil A, Ito J, Nakagawa K, Kuwata H,
et al: Ferroptosis driven by radical oxidation of n-6
polyunsaturated fatty acids mediates acetaminophen-induced acute
liver failure. Cell Death Dis. 11:1442020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Fan R, Sui J, Dong X, Jing B and Gao Z:
Wedelolactone alleviates acute pancreatitis and associated lung
injury via GPX4 mediated suppression of pyroptosis and ferroptosis.
Free Radic Biol Med. 173:29–40. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ma D, Li C, Jiang P, Jiang Y, Wang J and
Zhang D: Inhibition of ferroptosis attenuates acute kidney injury
in rats with severe acute pancreatitis. Dig Dis Sci. 66:483–492.
2021. View Article : Google Scholar
|