Introduction
Melanoma is the most aggressive form of skin cancer
and it is derived from activated or genetically altered epidermal
melanocytes (1). Human malignant
melanoma is a highly metastatic cancer that is markedly resistant
to chemotherapy with dacarbazine or temozolomide. The best
single-agent activity achieves a response rate of 15–30% and a
median duration of response of a few months (2). In the fields of veterinary medicine,
canine melanoma is the most common oral malignant tumor (3).
Lupeol is a triterpene contained in olives, fruits
such as mangos, strawberries, grapes and figs, numerous vegetables
and several medicinal plants (4).
Previous studies reported that lupeol possesses several
bioactivities, such as anti-inflammatory (5), antioxidant (6) and antitumor effects (7,8). In
particular, lupeol has been described as being beneficial for
inhibiting the proliferation of melanoma cells in vitro via
several mechanisms of action, including differentiation- (9,10) and
apoptosis-inducing activities (11), activation of p38 mitogen-activated
protein kinases (MAPK) (12),
anti-angiogenic activities (13),
remodeling of the actin cytoskeleton (14) and melanosome maturation (15).
A previous study by Saleem et al (11) reported that lupeol inhibits the
growth of highly aggressive human metastatic melanoma cells in
vivo by inducing apoptosis. They demonstrated the beneficial
effects of lupeol in a melanoma mouse model using intraperitoneal
injection of the compound. In that experiment, however, the
injections of lupeol were initiated at the time of tumor cell
transplantation. To the best of our knowledge, there is currently
no report of the differences between lupeol injection routes and
the effects of lupeol on tuberous tumor tissue in vivo. In
order to evaluate the use of lupeol in the clinical treatment of
melanoma patients, the effects of lupeol must be assessed using an
easy administration route. In this study, we investigated the
effects of lupeol via systemic and local administration in a
tuberous melanoma-bearing mouse model.
Materials and methods
Reagent
In this study, we used lupeol extracted from Indian
lettuce (Lactuca indica), [Hata et al (16)]. Olive oil was purchased from Wako
Pure Chemical Industries, Ltd. (Osaka, Japan). Lupeol was dissolved
in olive oil using heat (37°C) and sonification (3 h). The
concentration of dissolved lupeol was diluted to 5 mg/ml.
Preparation of the melanoma-bearing mouse
model
Forty female, 6-week-old C57BL/6 mice were purchased
from CLEA Japan, Inc. (Osaka, Japan). The animals were maintained
under conventional conditions. The use of these animals and the
procedures they underwent were approved by the Animal Research
Committee of Tottori University.
B16 2F2 melanoma cells were maintained in Dulbecco’s
modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine
serum (FBS), 100 μg/ml streptomycin and 100 U/ml penicillin in an
incubator at 37°C under a humidified atmosphere of 5%
CO2. The mice were anesthetized with inhalation of 3–5%
isoflurane (Intervet, Inc., Tokyo, Japan). A total of
1×106 B16 2F2 cells (1×107 cells/ml) were
subcutaneously injected into the dorsal regions of the mice. Mice
whose tumors grew to 5 mm in size were used in this study.
Study design
The mice (n=40) were randomized into five groups:
the non-treatment (NT) group, the subcutaneous injection of olive
oil (solvent control) into the dorsal region (C-si) group, the
subcutaneous injection of lupeol into the dorsal region (L-si)
group, the local injection of olive oil into the tumor tissue
(C-li) group and the local injection of lupeol into the tumor
tissue (L-li) group (n=8 per group). Single injections of lupeol
were systemically or locally administered to the mice (day 0). A
total of 0.1 ml of lupeol (20 mg/kg) was injected into each mouse.
On day 7 after the injections the mice were euthanized with
inhalation of 5% isoflurane followed by cervical dislocation. The
tumor growth rates were calculated according to the tumor volumes
(mm3/day). On days 0 and 7, the volumes of the tumor
tissues were calculated by measuring the mediastinum, transverse
lie and depth of each tumor. The tumors were removed and fixed in
10% buffered formalin.
Ki-67 staining
Tissue sections (3 μm) were placed on glass slides
and were deparaffinized, washed with ethanol and water and soaked
in phosphate-buffered saline (PBS). The sections were autoclaved
with 0.01 M citrate buffer (pH 6.0) for 15 min at 121°C, then
washed with PBS and incubated with rabbit polyclonal anti-Ki-67
antibodies (1:50, code no. E0468, Dako, Glostrup, Denmark) for 30
min at room temperature. After being washed with PBS, the sections
were incubated with rat anti-IgG antibodies (1:100, sc-372; Vector
Laboratories, Inc., Burlingame, CA, USA) for 30 min at room
temperature. The slides were washed with PBS and stained with the
VECTASTAIN ABC kit (PK-4000; Vector Laboratories, Inc.) for 30 min.
The tissue sections were counterstained with HistoGreen (Nichirei
Bioscience, Inc., Osaka, Japan) and then stained with nuclear fast
red.
Proliferating cell nuclear antigen (PCNA)
staining
Tissue sections (3-μm) were placed on glass slides
and were deparaffinized, washed with ethanol and water and soaked
in PBS. The sections were treated using microwaves with distilled
water for 5 min, then washed with PBS and incubated with 1%
hydrogen peroxide methanol for 30 min at room temperature. After
being washed with PBS, the sections were incubated with Histofine
MOUSESTAIN kit blocking reagent A (Nichirei Biosciences, Inc.) for
60 min at room temperature. The sections were then washed with PBS
and incubated with rabbit polyclonal anti-PCNA antibodies (1:200,
code no. M0879, Dako) for 60 min at room temperature. The slides
were then incubated with Histofine MOUSESTAIN kit blocking reagent
B (Nichirei Biosciences, Inc.) for 10 min at room temperature and
were then washed with PBS and incubated with Histofine MOUSESTAIN
kit-labeled polymer Max PO (Nichirei Biosciences, Inc.) for 10 min
at room temperature. The tissue sections were counterstained with
HistoGreen and then stained with nuclear fast red (Nichirei
Biosciences, Inc.).
Image analysis of Ki-67 and PCNA
staining
A quantitative digital morphometric analysis of the
Ki-67- and PCNA-positive tumor areas was performed. In brief, 10
randomly selected high-power fields (×200 magnification) per cross
section were photographed with a digital camera attached to an
Olympus microscope system (Olympus Corporation, Tokyo, Japan). The
color wavelengths of the copied images were transformed into
digital readings using the Lumina Vision software program (Mitani
Corporation, Tokyo, Japan), allowing for quantification of the
various color wavelengths with pixels as the unit of measurement.
The percentage of positive areas in the tumor tissues was
calculated by dividing the total pixel area of the positive areas
by the total pixel area corresponding to the total tumor tissue in
the field of view. The tumor tissues of three mice were analyzed
per group. The mean scores for 30 fields were used as the
percentages of positive areas per group.
RNA extraction and complementary DNA
(cDNA) synthesis
A total of 2.5×105 B16 2F2 melanoma cells
were seeded and precultured in 1 ml DMEM supplemented with 10% FBS
in 12-well plates for 12 h. The cells were washed twice with PBS
and incubated in the same medium for 2 days with or without 5 μM of
lupeol. Total RNA was isolated using the QuickGene RNA cultured
cell kit S (Fujifilm Co., Tokyo, Japan). Template cDNA synthesis
was performed with 5 μg of total RNA using the PrimeScript RT
reagent kit (Takara Bio Inc., Shiga, Japan).
Real-time reverse transcription
polymerase chain reaction (qRT-PCR)
In a Chromo4 fluorescent temperature cycler
(Bio-Rad, Hercules, CA, USA), 2.5% of each RT reaction solution was
amplified in 25 μl of 1X SYBR Premix Ex Taq (Takara Bio Inc.)
containing 0.2 μM of each primer. The samples were incubated in the
thermal cycler for an initial denaturation at 95°C for 10 sec
followed by 40 PCR cycles. Each cycle consisted of 95°C for 5 sec
and 60°C for 30 sec. The oligonucleotide primers used in the
experiment are listed in Table I.
To confirm the amplification of specific transcripts, melting curve
profiles (cooling the sample to 60°C and reheating slowly to 95°C
with continuous measurement of fluorescence) were produced at the
end of each PCR cycle. The relative expression levels of the two
mRNAs were normalized according to the amount of glyceraldehyde
3-phosphate dehydrogenase (GAPDH) mRNA (forward primer,
5′-TGTGTCCGTCGTGGATCTGA-3′; reverse primer,
5′-TTGCTGTTGAAGTCGCAGGAG-3′).
 | Table I.Effects of lupeol on gene expression
of B16 2F2 melanoma cells. |
Table I.
Effects of lupeol on gene expression
of B16 2F2 melanoma cells.
| Sequence
| RI/GAPDH (%)
|
|---|
| Gene | Forward | Reverse | Control | Lupeol |
|---|
| GUS |
5′-CTGTGACCGATACGGGATTGTG-3′ |
5′-ACCTCTAGGTGGTGCCGAAGTG-3′ | 100.0±2.8 | 108.4±8.0 |
| Tyrosinase |
5′-CAAGTACAGGCATCGGCCAAC-3′ |
5′-GGTGCATTGGCTTCTGGGTAA-3′ | 100.0±6.4 | 184.1±8.2a |
| Rab27a |
5′-TAGCACTGCAGGGACGCAAC-3′ |
5′-CAAGGCCAAGAACTTGATGAGGTAA-3′ | 100.0±4.8 | 123.9±14.5b |
| PCNA |
5′-GAGAGCTTGGCAATGGGAACA-3′ |
5′-GGGCACATCTGCAGACATACTGA-3′ | 100.0±5.3 | 36.5±3.1a |
| Ki-67 |
5′-CGTGTCAAACAAACTTGAATCTGTG-3′ |
5′-TCTGCAGATGCATCAAACTTGG-3′ | 100.0±4.0 | 39.9±4.0a |
Statistical analysis
Data were expressed as the mean ± standard error or
standard deviation. The statistical analyses were performed using
the Steel-Dwass test or the Student’s t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of lupeol via systemic and local
administration on tumor growth and histopathological changes in the
melanoma-bearing mouse model
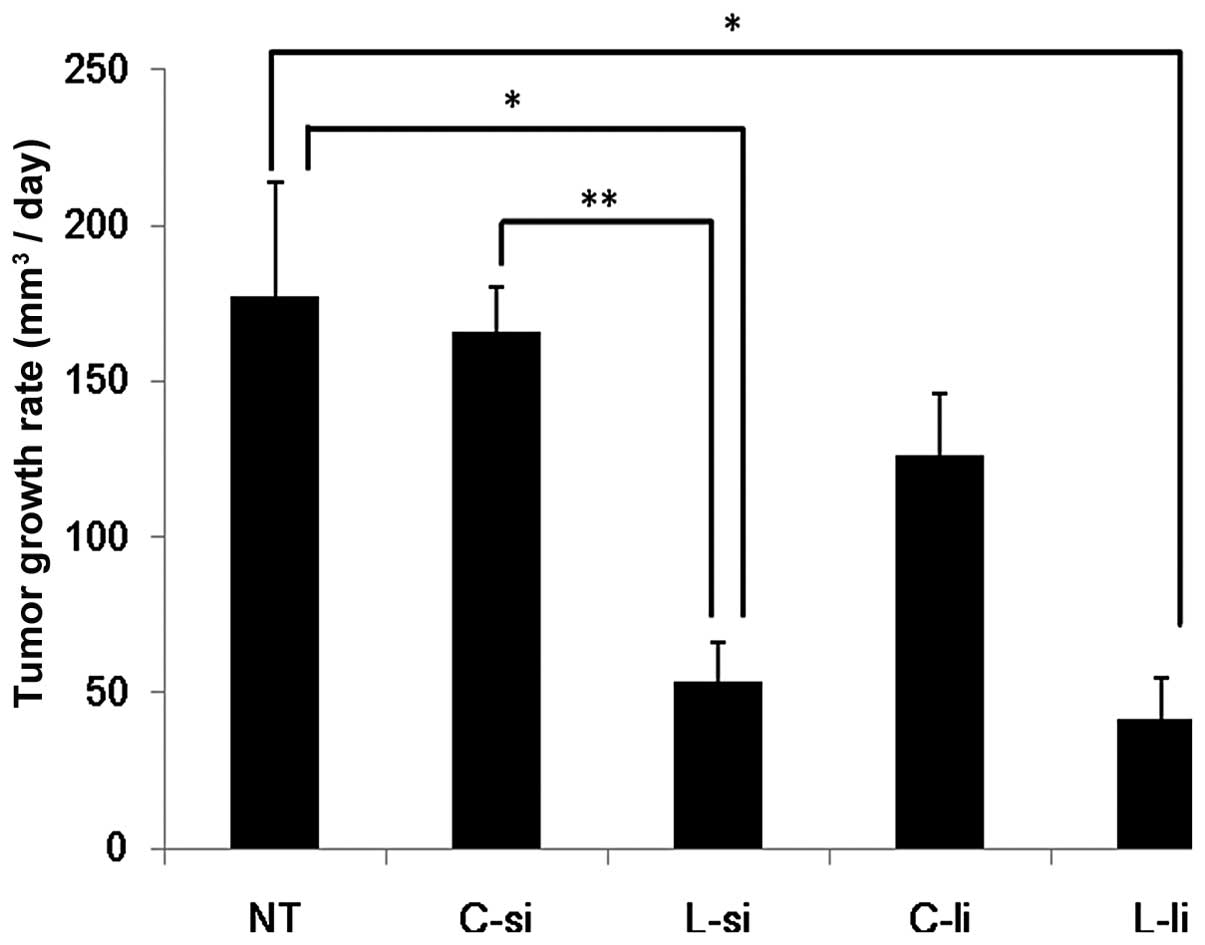
The tumor growth rates are shown in Fig. 1. In the L-si group (53.5±12.5
mm3/day), the growth rates of the tumor tissues were
significantly decreased compared to those observed in the NT
(177.5±36.4 mm3/day) and C-si (166.1±13.9
mm3/day) groups (P<0.05 vs. NT group and P<0.01
vs. C-si group). In the L-li group (41.8±12.7 mm3/day),
the growth rates of the tumor tissues were significantly decreased
compared to those observed in the NT group (P<0.05). In the L-li
group, the growth rates of the tumor tissues were slightly
decreased compared to those observed in the C-li group (126.0±19.0
mm3/day). There were no significant differences between
the L-si and L-li groups.
Effects of lupeol via systemic and local
administration on Ki-67 staining in the melanoma-bearing mouse
model
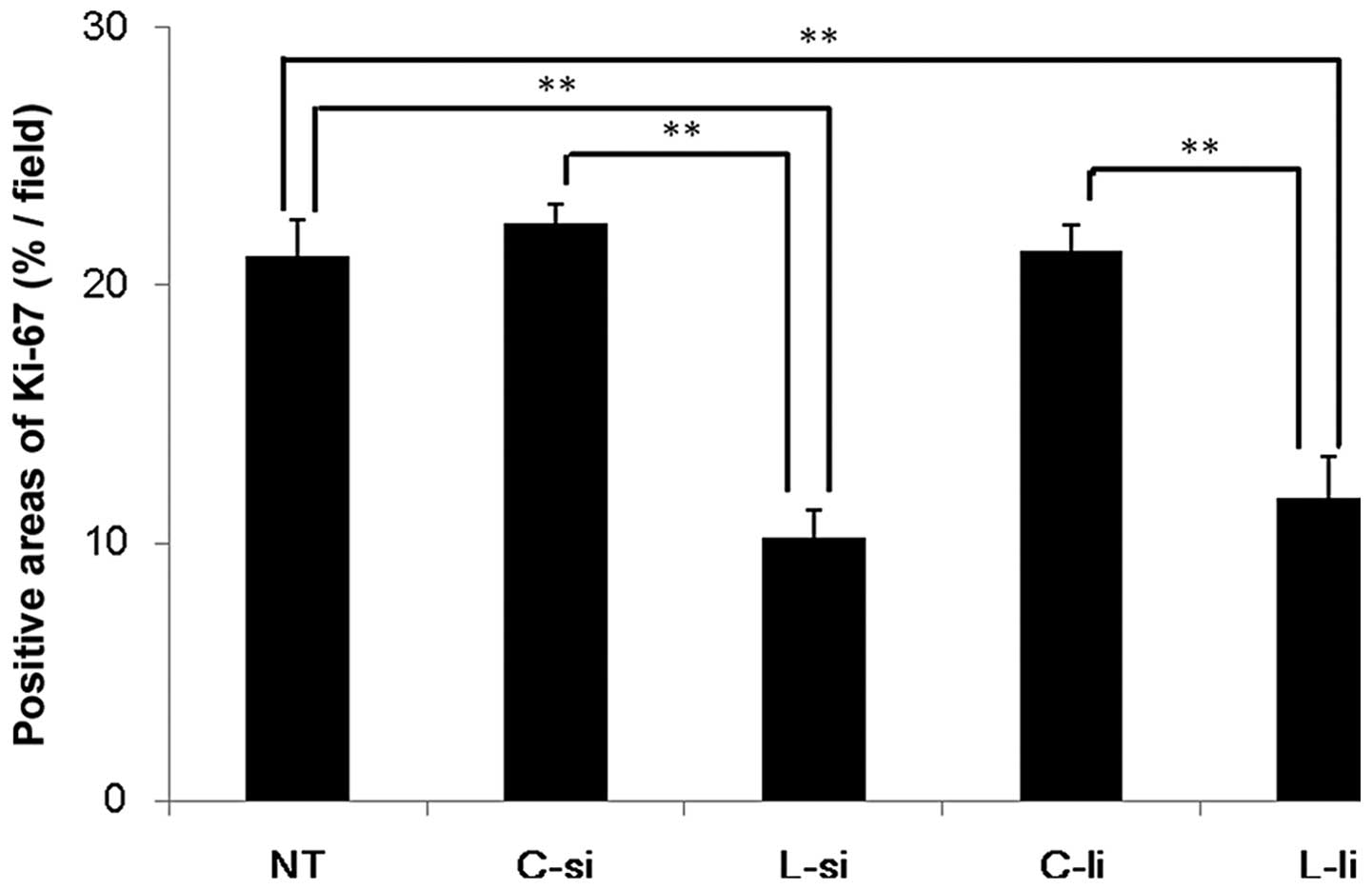
The results of the image analysis are shown in
Fig. 2. In the L-si group
(10.2±1.0%/field), the percentages of Ki-67-positive areas were
significantly decreased compared to those observed in the NT
(21.1±1.4%/field) and C-si (22.4±0.8%/field) groups (P<0.01). In
the L-li group (11.8±1.6%/field), the percentages of Ki-67-positive
areas were significantly decreased compared to those observed in
the NT and C-li (21.3±1.0%/field) groups (P<0.01). There were no
significant differences between the L-si and L-li groups.
Effects of lupeol via systemic and local
administration on PCNA staining in the melanoma-bearing mouse
model
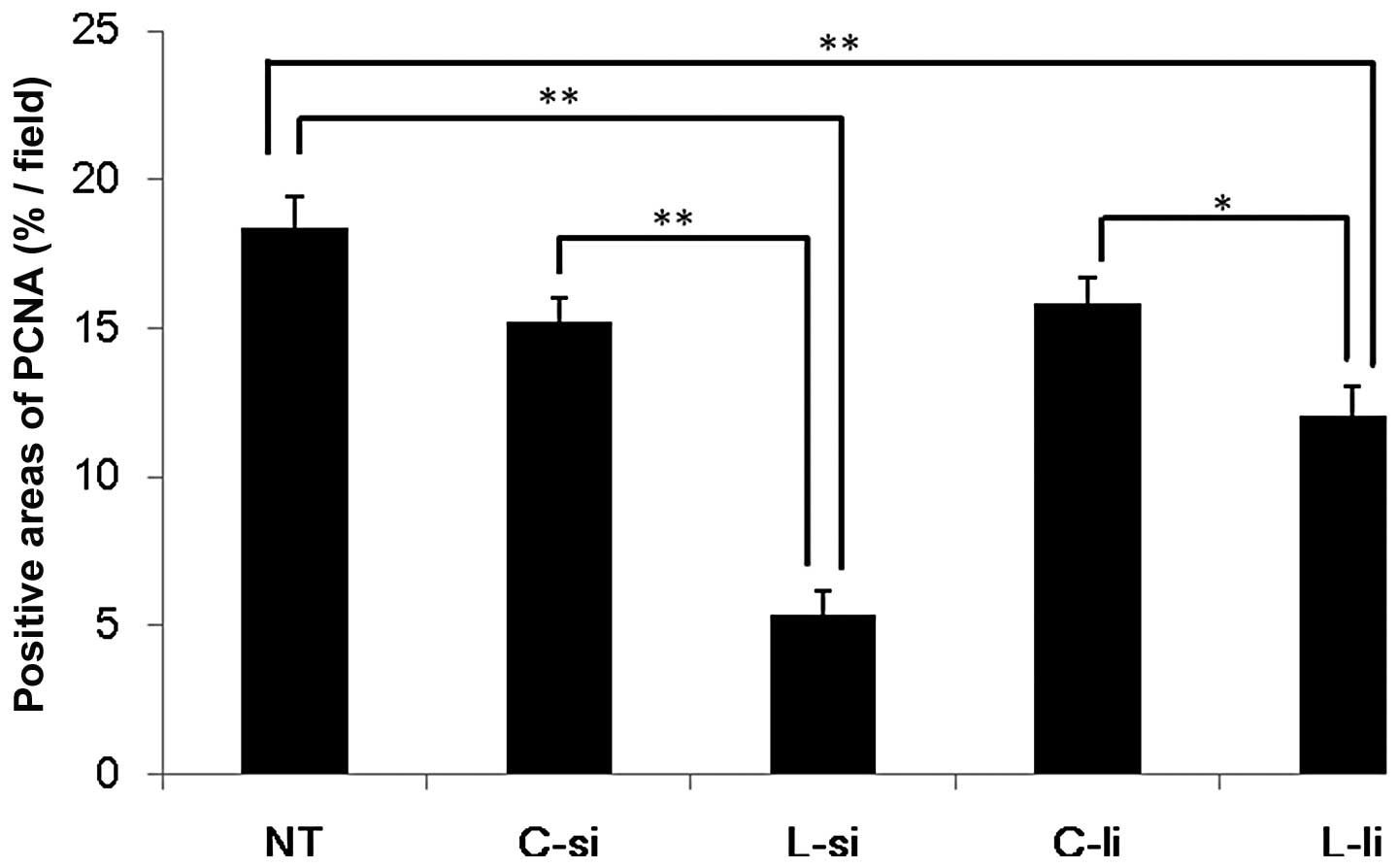
The results of the image analysis are shown in
Fig. 3. In the L-si group
(5.4±0.8%/field), the percentages of PCNA-positive areas were
significantly decreased compared to those observed in the NT
(18.4±1.1%/field) and C-si (15.2±0.8%/field) groups (P<0.01). In
the L-li group (12.1±0.9%/field), the percentages of PCNA-positive
areas were significantly decreased compared to those observed in
the NT and C-li (15.9±0.8%/field) groups (P<0.01 vs. NT group
and P<0.05 vs. C-li group). There were no significant
differences between the L-si and L-li groups.
Effects of lupeol on the expression of
markers of melanoma cell differentiation and proliferation
Administration of 5 μM of lupeol did not affect the
gene expression of β-glucuronidase (GUS, a housekeeping gene) in
the B16 2F2 cells. The markers of pigment cell differentiation
tyrosinase and Rab27a were elevated by 5 μM of lupeol at the mRNA
level; however, the agent markedly attenuated the expression of the
PCNA and Ki-67 genes (Table I).
Discussion
Several previous studies indicated that lupeol
exhibits anti-tumor activities against melanoma cells in
vitro (9,11–16). A
previous study by Saleem et al (11) demonstrated that intraperitoneal
injection of lupeol inhibits tumor growth in a human
melanoma-bearing mouse model. To incorporate lupeol into the
clinical treatment of melanoma patients, its effects must be
evaluated using an easy administration route. In the present study
it was demonstrated that systemic and local administration of
lupeol inhibits tumor growth in a melanoma-bearing mouse model.
In this study, systemic and local administration of
lupeol decreased the percentage of Ki-67- and PCNA-positive areas
in the tumor tissues. Ki-67 is a cell proliferation marker that is
detected during all the active phases of the cell cycle and is
absent in resting cells (17). The
levels of Ki-67 increase during the synthesis phase (S phase) until
mitosis, when its expression reaches a maximum. After cell
division, cells in the G1 phase exhibit a decrease in the Ki-67
expression until they re-enter the S phase, when the levels of
Ki-67 increase again (18). PCNA is
also a cell proliferation marker whose levels increase
progressively from the G1 to the S phase (19). In numerous melanoma patients the
expression of Ki-67 and PCNA in the tumor tissue is correlated with
the presence of malignancy and prognosis (20). The intraperitoneal injection of
lupeol has been reported to reduce the expression of Ki-67 and PCNA
in tumor tissue (11). Our data
indicated similar results, suggesting that systemic and local
administration of lupeol is associated with efficient cell cycle
arrest of melanoma cells.
The effects of lupeol on the expression of markers
of melanoma cell differentiation and proliferation were
investigated (Table I). The markers
of pigment cell differentiation, tyrosinase and Rab27a (15) were elevated by lupeol at the mRNA
level; however, the agent markedly attenuated the expression of the
PCNA and Ki-67 genes. These results suggest that lupeol suppresses
PCNA and Ki-67 by inducing B16 2F2 cell differentiation and they
are in agreement with the results shown in Figs. 2 and 3.
Several treatment methods have been used for
melanoma patients, including surgical resection, chemotherapy and
radiation therapy (21). Recently,
high-dose interferon-α therapy, high-dose interleukin-2 therapy,
antibody blockade of cytotoxic T-lymphocyte-associated antigen 4,
inhibitors of the MAPK pathway and adoptive cell therapy have also
been used in the treatment of melanoma (20–22).
Our results indicate that the systemic and local administration of
lupeol may be an effective novel therapeutic option for melanoma
patients.
Acknowledgements
This study was supported by a grant
from the Ministry of Education, Sports, Culture, Science and
Technology (no. 20580352).
References
|
1
|
Oliveria S, Dusza S and Berwick M: Issues
in the epidemiology of melanoma. Expert Rev Anticancer Ther.
1:453–459. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Korn EL, Liu PY, Lee SJ, Chapman JA,
Niedzwiecki D, Suman VJ, Moon J, Sondak VK, Atkins MB, Eisenhauer
EA, et al: Meta-analysis of phase II cooperative group trials in
metastatic stage IV melanoma to determine progression-free and
overall survival benchmarks for future phase II trials. J Clin
Oncol. 26:527–534. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bergman PJ: Canine oral melanoma. Clin
Tech Small Anim Pract. 22:55–60. 2007. View Article : Google Scholar
|
|
4
|
Saleem M, Kaur S, Kweon MH, Adhami VM,
Afaq F and Mukhtar H: Lupeol, a fruit and vegetable based
triterpene, induces apoptotic death of human pancreatic
adenocarcinoma cells via inhibition of Ras signaling pathway.
Carcinogenesis. 26:1956–1964. 2005. View Article : Google Scholar
|
|
5
|
Geetha T and Varalakshmi P:
Anti-inflammatory activity of lupeol and lupeol linoleate in rats.
J Ethnopharmacol. 76:77–80. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Prasad S, Kalra N and Shukla Y:
Hepatoprotective effects of lupeol and mango pulp extract of
carcinogen induced alteration in Swiss albino mice. Mol Nutr Food
Res. 51:352–359. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chaturvedi PK, Bhui K and Shukla Y:
Lupeol: connotations for chemoprevention. Cancer Lett. 263:1–13.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Saleem M: Lupeol, a novel
anti-inflammatory and anti-cancer dietary triterpene. Cancer Lett.
285:109–115. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hata K, Ishikawa K, Hori K and Konishi T:
Differentiation-inducing activity of lupeol, a lupane-type
triterpene from Chinese dandelion root (Hokouei-kon), on a mouse
melanoma cell line. Biol Pharm Bull. 23:962–967. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hata K, Mukaiyama T, Tsujimura N, Sato Y,
Kosaka Y, Sakamoto K and Hori K: Differentiation-inducing activity
of lupane triterpenes on a mouse melanoma cell line.
Cytotechnology. 52:151–158. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saleem M, Maddodi N, Abu Zaid M, Khan N,
bin Hafeez B, Asim M, Suh Y, Yun JM, Setaluri V and Mukhtar H:
Lupeol inhibits growth of highly aggressive human metastatic
melanoma cells in vitro and in vivo by inducing apoptosis. Clin
Cancer Res. 14:2119–2127. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hata K, Hori K and Takahashi S: Role of
p38 MAPK in lupeol-induced B16 2F2 mouse melanoma cell
differentiation. J Biochem. 134:441–445. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
You YJ, Nam NH, Kim Y, Bae KH and Ahn BZ:
Antiangiogenic activity of lupeol from Bombax ceiba.
Phytother Res. 17:341–344. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hata K, Hori K, Murata J and Takahashi S:
Remodeling of actin cytoskeleton in lupeol-induced B16 2F2 cell
differentiation. J Biochem. 138:467–472. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ogiwara K and Hata K: Melanoma cell
differentiation induced by lupeol separates into two stages:
morphological and functional changes. J Nat Med. 63:323–326. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hata K, Hori K, Mukaiyama T, Sakamoto K
and Takahashi S: Activators of melanin biosynthesis from Lactuca
indica. Nat Med. 57:238–241. 2003.
|
|
17
|
Brown DC and Gatter KC: Ki67 protein: the
immaculate deception? Histopathology. 40:2–11. 2002. View Article : Google Scholar
|
|
18
|
Lopez F, Belloc F, Lacombe F, Dumain P,
Reiffers J, Bernard P and Boisseau MR: Modalities of synthesis of
Ki67 antigen during the stimulation of lymphocytes. Cytometry.
12:42–49. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Moldovan GL, Pfander B and Jentsch S:
PCNA, the maestro of the replication fork. Cell. 129:665–679. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gould Rothberg BE, Bracken MB and Rimm DL:
Tissue biomarkers for prognosis in cutaneous melanoma: a systematic
review and meta-analysis. J Natl Cancer Inst. 101:452–474.
2009.PubMed/NCBI
|
|
21
|
Lee B, Mukhi N and Liu D: Current
management and novel agents for malignant melanoma. J Hematol
Oncol. 5:32012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Garbe C, Eigentler TK, Keilholz U,
Hauschild A and Kirkwood JM: Systematic review of medical treatment
in melanoma: current status and future prospects. Oncologist.
16:5–24. 2011. View Article : Google Scholar : PubMed/NCBI
|