1. Introduction
In 2003, proprotein convertase subtilisin/kexin type
9 (PCSK9) was identified by the bioinformatics method and DNA
microarray technology (1,2). PCSK9 is a member of the subtilisin
family of proprotein convertases and is involved in the degradation
of low-density lipoprotein receptor (LDLR) in the liver (3). Gain- and loss-of-function mutations of
PCSK9 may result in hyper- and hypocholesterolemia, respectively,
thereby affecting liver LDLR degradation and blood lipid levels.
Cohen et al (4) reported
that African-Americans harbouring nonsense mutations in PCSK9
exhibited a 28% decrease in LDL cholesterol levels and a 88% lower
risk of coronary heart disease (CHD), whereas Caucasians with less
severe mutations in PCSK9 exhibited a 15% decrease in LDL levels
and a 47% lower risk of CHD. Surprisingly, the LDL-lowering
mutations of PCSK9 were shown to exert a protective effect in CHD.
A previous phase II clinical trial of a monoclonal antibody to
PCSK9 demonstrated that patients with severe heterozygous familial
hypercholesterolemia on high-dose statins who were injected with
anti-PCSK9 monoclonal antibody exhibited maximum LDL-C levels
(5). The results, however, depend
on the phase III side effects of the anti-PCSK9 monoclonal antibody
(6). If proven efficient, the
combination of anti-PCSK9 anibodies with statins may become a
potent lipid-lowering and cardioprotective treatment that should be
used prior to the advancement of atherosclerosis, since the effects
of lowering LDL levels do not depend solely on the extent, but also
on the duration of the decrease (7). The extensive investigation regarding
PCSK9 revealed novel functions, in addition to those in lipid
metabolism, including cell apoptosis, inflammatory response,
neuronal development and tumor metastasis (8–11).
Neurodegeneration is a progressive loss of neurons
and synapses in the cerebral cortex and certain subcortical
regions. A number of neurodegenerative disorders (ND), such as
Parkinson’s disease, Alzheimer’s disease (AD) and Huntington’s
disease, occur as a result of neurodegenerative processes. AD is
the major form of ND and is characterized by senile plaques and
neurofibrillary tangles in the brain. Excessive generation and
accumulation of amyloid β peptide (Aβ) in the brain is an important
factor implicated in AD. Neuronal apoptosis is also a primary
mechanism underlying AD pathogenesis. Table I shows the numerous pro- and
anti-apoptotic factors that may mediate neuronal apoptosis.
Although several treatments are currently available to mitigate the
progression of the disease, there is no established medical method
to definitively prevent the progression of AD; the loss of neurons
is permanent, due to their lack of proliferative ability (12).
 | Table IPro- and anti-apoptotic factors in
neurons. |
Table I
Pro- and anti-apoptotic factors in
neurons.
2. Neuronal apoptosis rate is lower compared
to that in other cell types
Using New Zealand rabbit models on a high-fat diet
for 7 months, our research demonstrated that hyperlipidemia may
induce neuronal apoptosis. Higher rates of apoptosis in the brain
were observed in the high-fat diet group, compared to those in the
normal cholesterol group (0.83 vs. 0.30%, unpublished data). A
study by Cunningham et al (13) also reported that the apoptosis rate
in brain nerve cells induced by endotoxins in neurodegenerative
mouse models is lower, which is in accordance with the findings of
our study. However, compared to other systems, hyperlipidemia is
associated with significantly lower rates of neuronal apoptosis in
the brain. The apoptosis rate of smooth muscle cells in
atherosclerotic lesions was shown to be 11±7.8% in miniature pigs
on a high-fat diet for 37 weeks (14). Nematbakhsh et al (15) observed an endothelial cell apoptosis
rate of 8% in hyperlipidemic rabbits. However, the reason for the
significant difference in the apoptosis rate of nerve cells and
other cells exposed to hyperlipidemia has not been determined. It
was hypothesized that the presence of the blood-brain barrier (BBB)
was a possible explanation. However, the oxidized LDL (ox-LDL)
content was found to be increased in the brain and in the blood of
AD patients (16), indicating that
BBB does not play a major role under hyperlipidemic conditions.
Martins et al (17) proposed
two mechanisms that may underlie the increase in the lipid content
of the brain during hyperlipidemia. The first mechanism is BBB
damage. Ox-LDL may damage the BBB by inducing endothelial cell
apoptosis, which increases the permeability of the BBB membrane,
allowing lipids to cross the BBB into the brain. The second
mechanism involves necessary components involved in lipid
synthesis, such as unsaturated fatty acids, crossing the BBB,
thereby increasing endogenous lipid synthesis in the brain. The
brain lipidogram is significantly affected in hyperlipidemia.
However, the effect of the BBB on the apoptosis rate is minimal. We
further hypothesized that a pathway inhibiting neuronal apoptosis
may exist in the process of hyperlipidemia, along with an
additional pathway inducing brain nerve cell apoptosis; however,
further investigation is required to identify the common regulation
of the two diametrically opposed pathways.
3. PCSK9 promotes neuronal apoptosis
In this study, we mainly focused on the association
between cell apoptosis and PCSK9. The significant correlations
between PCSK9 and cell apoptosis are as follows: i) Wu et al
(8) reported that ox-LDL may
upregulate PCSK9 expression in human umbilical vein endothelial
cells (HUVECs), whereas PCSK9 siRNA inhibits HUVEC apoptosis
induced by ox-LDL through the Bcl/Bax-caspase 9-caspase 3 pathway;
and ii) previous studies suggested that PCSK9 may be associated
with neuronal apoptosis (18,19).
Cameron et al (20) reported
that berberine may decrease PCSK9 expression, whereas Ji and Shen
(21) indicated that berberine
exerts a potentially preventive effect on AD through a variety of
mechanisms. We therefore considered the potential involvement of
PCSK9 in the effect of berberine against AD. The link between PCSK9
and apoptosis was previously investigated. PCSK9 may regulate the
expression of inflammatory factors induced by ox-LDL through NF-κB
(9), whereas the NF-κB/B-cell
lymphoma 2 (Bcl-2) signaling pathway is crucial in mediating
apoptosis. The effect of PCSK9 in cerebellar granule neuron
apoptosis is reflected by the fact that the effects of wild-type
and mutant PCSK9 on apoptosis are partially reversed by BAF, a
caspase inhibitor. Moreover, apoptotic mediators, such as caspase 3
and death receptor 6, may be involved in neuronal apoptotic
signaling induced by PCSK9 (19). A
recent study demonstrated that PCSK9 promotes neuronal apoptosis
through the decreae of apolipoprotein E receptor 2 levels and
deregulation of anti-apoptotic signaling pathways (22). However, further investigations are
required to identify the precise PCSK9-related mechanism leading to
neuronal apoptosis.
4. PCSK9 inhibits neuronal apoptosis
Although PCSK9 is also referred to as neural
apoptosis-regulated convertase 1, certain studies failed to confirm
its pro-apoptotic effect. Ranheim et al (23) demonstrated that PCSK9 significantly
downregulated the poly(ADP-ribose) polymerase (PARP) family in
HepG2 cells through microarray analysis of D374Y-PCSK9, a
gain-of-function mutant. B16F1 melanoma cells were injected in
PCSK9−/− mice to induce liver metastasis. The findings revealed an
increased rate of apoptosis in the liver stroma and metastases.
Furthermore, the pro-apoptotic factor tumor necrosis factor-α
levels were increased and the anti-apoptotic factor Bcl-2 levels
were decreased (10), confirming
the anti-apoptotic function of PCSK9. However, the precise
mechanism through which PCSK9 inhibits neuronal apoptosis has not
been fully elucidated.
The following pathways are involved in amyloid
precursor protein (APP) metabolism: i) APP is cleaved by
α-secretase to generate soluble APP-α and C83; ii) APP is cleaved
by the β-site APP-cleaving enzyme 1 (BACE1) and γ-secretase to
generate insoluble Aβ. The former is generally considered to be the
major metabolic pathway. Under pathological conditions, the
generation of numerous Aβ peptides was suggested to cause AD
(24). The BACE1 level is increased
in the majority of sporadic types of AD. Consistent with the
increase in BACE1, higher concentrations of Aβ may induce neuronal
apoptosis and lead to AD.
Jonas et al (25) and and Ko and Puglielli (26) observed that the levels of BACE1 and
Aβ were increased in the brains of PCSK9(−/−) mice, the
overexpression of PCSK9 in CHO cells decreased the level of BACE1,
PCSK9 siRNA increased the level of BACE1 and secreted PCSK9 may
stimulate the degradation of BACE1. Those findings indicated that
PCSK9 possibly decreases Aβ generation to prevent neuronal
apoptosis through the degradation of BACE1 in nerve cells. However,
Liu et al (27) reported
that the levels of PCSK9 and BACE1 expression are not important;
thus, further investigations are required to determine whether they
affect BACE1 activity.
5. Inconsistencies and future
directions
Shibata et al (28) investigated two single-nucleotide
polymorphisms (SNPs) of PCSK9, namely rs11583680 and rs662145, and
suggested that SNPs are not associated with AD. In addition,
Reynolds et al (29)
investigated the association between lipid pathway genes and AD and
reported that PCSK9 is not associated with AD. However, although
the above-mentioned epidemiologic studies did not lead to a
definitive conclusion regarding the positive correlation between
PCSK9 and AD under site selection, case scale, or other reasons,
PCSK9 may still be involved in the pathogenesis of AD.
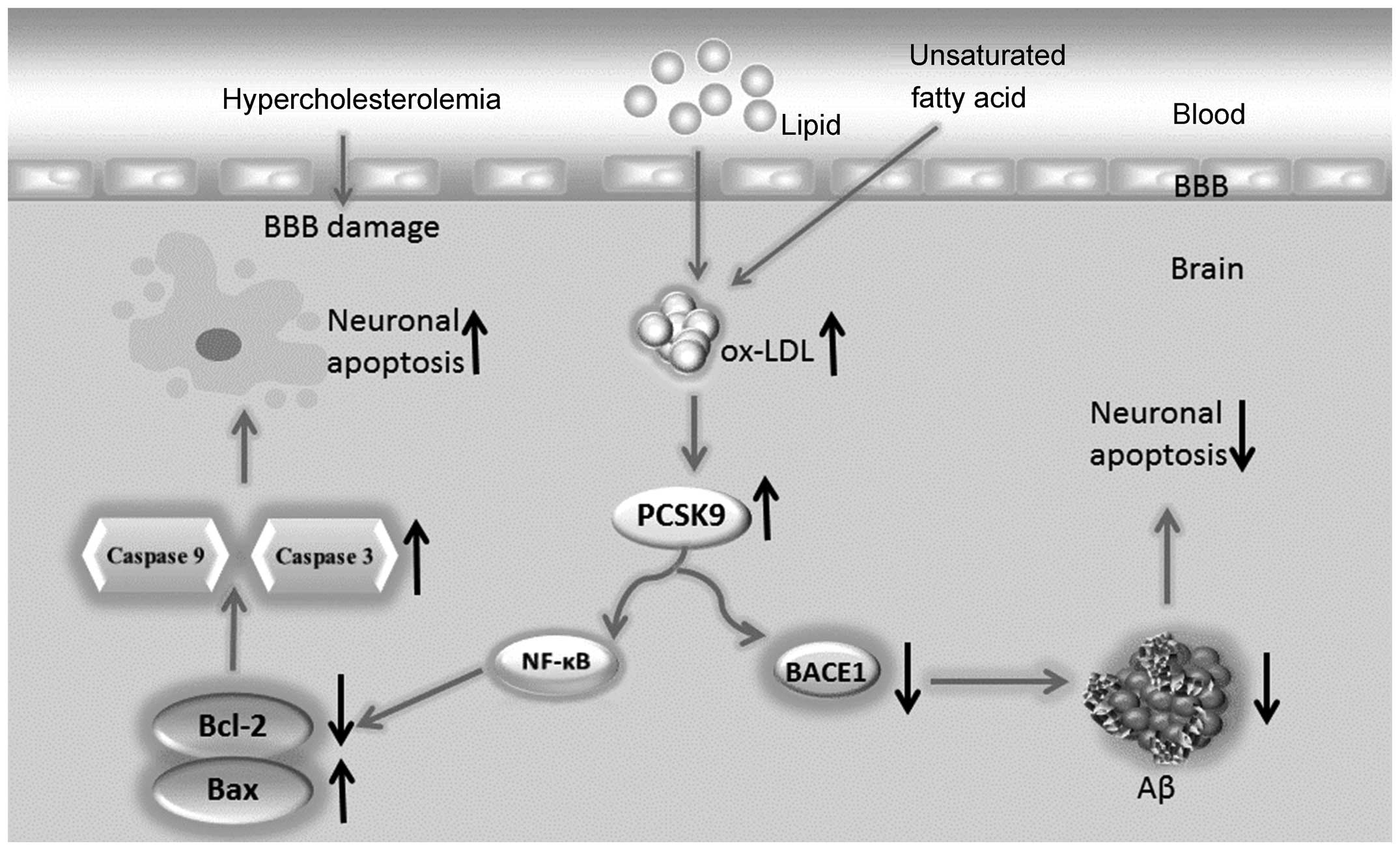
In this study, we proposed the following hypothesis:
hyperlipidemia increases the lipid content, particularly ox-LDL, of
the brain. The increased levels of ox-LDL may upregulate PCSK9
expression in nerve cells. Neuronal apoptosis is induced through
the NF-κB-Bcl-2/Bax-caspase 9-caspase 3 signaling pathway when
PCSK9 expression increases. Additionally, through the degradation
of BACE1, PCSK9 decreases Aβ generation via the inhibition of the
APP/A β metabolic pathway, which decreases neuronal apoptosis
induced by Aβ. Therefore, PCSK9 exerts a dual regulatory effect on
neuronal apoptosis (Fig. 1),
maintaining apoptosis at a low level or limiting its increase,
leading to the slow progression of ND. The elucidation of the
association among hyperlipidemia, PCSK9 expression and neuronal
apoptosis requires further investigations and animal testing.
Furthermore, along with the anti-PCSK9 antibody, which was proven
successful in lowering lipid levels (5), drugs targeting PCSK9 are eagerly
anticipated for evaluation in the treatment of AD.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (81200217), the Key Project of
the Educational Department of Hunan Province (10A105), the Project
of the Health Department of Hunan Province (B2011-042), the Aid
Program for Science and Technology Innovative Research Team in
Higher Educational Institutions of Hunan Province and the National
Natural Science Foundation of China (81370376).
References
|
1
|
Maxwell KN, Soccio RE, Duncan EM, Sehayek
E and Breslow JL: Novel putative SREBP and LXR target genes
identified by microarray analysis in liver of cholesterol-fed mice.
J Lipid Res. 44:2109–2119. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Seidah NG, Benjannet S, Wickham L, et al:
The secretory proprotein convertase neural apoptosis-regulated
convertase 1 (NARC-1): liver regeneration and neuronal
differentiation. Proc Natl Acad Sci USA. 100:928–933. 2003.
View Article : Google Scholar
|
|
3
|
Seidah NG: PCSK9 as a therapeutic target
of dyslipidemia. Expert Opin Ther Targets. 13:19–28. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cohen JC, Boerwinkle E, Mosley TH Jr and
Hobbs HH: Sequence variations in PCSK9, low LDL, and protection
against coronary heart disease. N Engl J Med. 354:1264–1272. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stein EA, Gipe D, Bergeron J, et al:
Effect of a monoclonal antibody to PCSK9, REGN727/SAR236553, to
reduce low-density lipoprotein cholesterol in patients with
heterozygous familial hypercholesterolaemia on stable statin dose
with or without ezetimibe therapy: a phase 2 randomised controlled
trial. Lancet. 380:29–36. 2012.
|
|
6
|
Hall SS: Genetics: a gene of rare effect.
Nature. 496:152–155. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brown MS and Goldstein JL: Biomedicine.
Lowering LDL - not only how low, but how long? Science.
311:1721–1723. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu CY, Tang ZH, Jiang L, Li XF, Jiang ZS
and Liu LS: PCSK9 siRNA inhibits HUVEC apoptosis induced by ox-LDL
via Bcl/Bax-caspase9-caspase3 pathway. Mol Cell Biochem.
359:347–358. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tang Z, Jiang L, Peng J, et al: PCSK9
siRNA suppresses the inflammatory response induced by oxLDL through
inhibition of NF-κB activation in THP-1-derived macrophages. Int J
Mol Med. 30:931–938. 2012.PubMed/NCBI
|
|
10
|
Sun X, Essalmani R, Day R, Khatib AM,
Seidah NG and Prat A: Proprotein convertase subtilisin/kexin type 9
deficiency reduces melanoma metastasis in liver. Neoplasia.
14:1122–1131. 2012.PubMed/NCBI
|
|
11
|
Poirier S, Prat A, Marcinkiewicz E, et al:
Implication of the proprotein convertase NARC-1/PCSK9 in the
development of the nervous system. J Neurochem. 98:838–850. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Citron M: Alzheimer’s disease: strategies
for disease modification. Nat Rev Drug Discov. 9:387–398. 2010.
|
|
13
|
Cunningham C, Wilcockson DC, Campion S,
Lunnon K and Perry VH: Central and systemic endotoxin challenges
exacerbate the local inflammatory response and increase neuronal
death during chronic neurodegeneration. J Neurosci. 25:9275–9284.
2005. View Article : Google Scholar
|
|
14
|
Verhamme P, Quarck R, Hao H, et al:
Dietary cholesterol withdrawal reduces vascular inflammation and
induces coronary plaque stabilization in miniature pigs. Cardiovasc
Res. 56:135–144. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nematbakhsh M, Haghjooyjavanmard S,
Mahmoodi F and Monajemi AR: The prevention of endothelial
dysfunction through endothelial cell apoptosis inhibition in a
hypercholesterolemic rabbit model: the effect of L-arginine
supplementation. Lipids Health Dis. 7:272008. View Article : Google Scholar
|
|
16
|
Kankaanpaa J, Turunen SP, Moilanen V,
Horkko S and Remes AM: Cerebrospinal fluid antibodies to oxidized
LDL are increased in Alzheimer’s disease. Neurobiol Dis.
33:467–472. 2009.PubMed/NCBI
|
|
17
|
Martins IJ, Berger T, Sharman MJ, Verdile
G, Fuller SJ and Martins RN: Cholesterol metabolism and transport
in the pathogenesis of Alzheimer’s disease. J Neurochem.
111:1275–1308. 2009.
|
|
18
|
Chiang LW, Grenier JM, Ettwiller L, et al:
An orchestrated gene expression component of neuronal programmed
cell death revealed by cDNA array analysis. Proc Natl Acad Sci USA.
98:2814–2819. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bingham B, Shen R, Kotnis S, et al:
Proapoptotic effects of NARC 1 (= PCSK9), the gene encoding a novel
serine proteinase. Cytometry A. 69:1123–1131. 2006.PubMed/NCBI
|
|
20
|
Cameron J, Ranheim T, Kulseth MA, Leren TP
and Berge KE: Berberine decreases PCSK9 expression in HepG2 cells.
Atherosclerosis. 201:266–273. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ji HF and Shen L: Berberine: a potential
multipotent natural product to combat Alzheimer’s disease.
Molecules. 16:6732–6740. 2011.PubMed/NCBI
|
|
22
|
Kysenius K, Muggalla P, Matlik K, Arumae U
and Huttunen HJ: PCSK9 regulates neuronal apoptosis by adjusting
ApoER2 levels and signaling. Cell Mol Life Sci. 69:1903–1916. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ranheim T, Mattingsdal M, Lindvall JM, et
al: Genome-wide expression analysis of cells expressing gain of
function mutant D374Y-PCSK9. J Cell Physiol. 217:459–467. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Di Paolo G and Kim TW: Linking lipids to
Alzheimer’s disease: cholesterol and beyond. Nat Rev Neurosci.
12:284–296. 2011.
|
|
25
|
Jonas MC, Costantini C and Puglielli L:
PCSK9 is required for the disposal of non-acetylated intermediates
of the nascent membrane protein BACE1. EMBO Rep. 9:916–922. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ko MH and Puglielli L: Two endoplasmic
reticulum (ER)/ER Golgi intermediate compartment-based lysine
acetyltransferases post-translationally regulate BACE1 levels. J
Biol Chem. 284:2482–2492. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu M, Wu G, Baysarowich J, et al: PCSK9
is not involved in the degradation of LDL receptors and BACE1 in
the adult mouse brain. J Lipid Res. 51:2611–2618. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shibata N, Ohnuma T, Higashi S, et al: No
genetic association between PCSK9 polymorphisms and Alzheimer’s
disease and plasma cholesterol level in Japanese patients.
Psychiatr Genet. 15:2392005.
|
|
29
|
Reynolds CA, Hong MG, Eriksson UK, et al:
Analysis of lipid pathway genes indicates association of sequence
variation near SREBF1/TOM1L2/ATPAF2 with dementia risk. Hum Mol
Genet. 19:2068–2078. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mattson MP and Camandola S: NF-κB in
neuronal plasticity and neurodegenerative disorders. J Clin Invest.
107:247–254. 2001.
|
|
31
|
Sola S, Aranha MM and Rodrigues CM:
Driving apoptosis-relevant proteins toward neural differentiation.
Mol Neurobiol. 46:316–331. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Culmsee C and Plesnila N: Targeting Bid to
prevent programmed cell death in neurons. Biochem Soc Trans.
34:1334–1340. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wong HK, Fricker M, Wyttenbach A, et al:
Mutually exclusive subsets of BH3-only proteins are activated by
the p53 and c-Jun N-terminal kinase/c-Jun signaling pathways during
cortical neuron apoptosis induced by arsenite. Mol Cell Biol.
25:8732–8747. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lindsten T, Zong WX and Thompson CB:
Defining the role of the Bcl-2 family of proteins in the nervous
system. Neuroscientist. 11:10–15. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Almeida A: Genetic determinants of
neuronal vulnerability to apoptosis. Cell Mol Life Sci. 70:71–88.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gotz R, Karch C, Digby MR, Troppmair J,
Rapp UR and Sendtner M: The neuronal apoptosis inhibitory protein
suppresses neuronal differentiation and apoptosis in PC12 cells.
Hum Mol Genet. 9:2479–2489. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Baratchi S, Kanwar RK and Kanwar JR:
Survivin: a target from brain cancer to neurodegenerative disease.
Crit Rev Biochem Mol Biol. 45:535–554. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dhandapani KM and Brann DW: Transforming
growth factor-β: a neuroprotective factor in cerebral ischemia.
Cell Biochem Biophys. 39:13–22. 2003.
|
|
39
|
Singh MH, Brooke SM, Zemlyak I and
Sapolsky RM: Evidence for caspase effects on release of cytochrome
c and AIF in a model of ischemia in cortical neurons.
Neurosci Lett. 469:179–183. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chiou SH, Chen SJ, Peng CH, et al:
Fluoxetine up-regulates expression of cellular FLICE-inhibitory
protein and inhibits LPS-induced apoptosis in hippocampus-derived
neural stem cell. Biochem Biophys Res Commun. 343:391–400. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Broughton BR, Reutens DC and Sobey CG:
Apoptotic mechanisms after cerebral ischemia. Stroke. 40:e331–e339.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Blaise S, Kneib M, Rousseau A, et al: In
vivo evidence that TRAF4 is required for central nervous system
myelin homeostasis. PLoS One. 7:e309172012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
LeBlanc AC: The role of apoptotic pathways
in Alzheimer’s disease neurodegeneration and cell death. Curr
Alzheimer Res. 2:389–402. 2005.
|
|
44
|
Joseph D’Ercole A and Ye P: Expanding the
mind: insulin-like growth factor I and brain development.
Endocrinology. 149:5958–5962. 2008.PubMed/NCBI
|
|
45
|
McCarthy MJ, Rubin LL and Philpott KL:
Involvement of caspases in sympathetic neuron apoptosis. J Cell
Sci. 110(Pt 18): 2165–2173. 1997.PubMed/NCBI
|
|
46
|
Ludwig-Galezowska AH, Flanagan L and Rehm
M: Apoptosis repressor with caspase recruitment domain, a
multifunctional modulator of cell death. J Cell Mol Med.
15:1044–1053. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gora-Kupilas K and Josko J: The
neuroprotective function of vascular endothelial growth factor
(VEGF). Folia Neuropathol. 43:31–39. 2005.PubMed/NCBI
|
|
48
|
Krantic S, Mechawar N, Reix S and Quirion
R: Apoptosis-inducing factor: a matter of neuron life and death.
Prog Neurobiol. 81:179–196. 2007. View Article : Google Scholar : PubMed/NCBI
|