Introduction
Skeleton is a highly organized tissue that provides
a structural framework to facilitate the locomotion and activity of
daily living. As such, it is metabolically active and is able to
sense and adapt to mechanical stimuli in order to maintain a
balance between bone formation and absorption (1). During orthodontic tooth movement,
mechanical stimuli also play a crucial role in bone remodeling as a
result of osteoblast and osteoclast.
Physiologically, tooth movement is a slow process
that occurs mainly in the buccal direction into cancellous bone. By
contrast, orthodontic tooth movement can occur rapidly or slowly,
depending on the physical characteristics of the applied force and
the size and biological response of the periodontal ligament (PDL).
This tooth movement has been defined as the result of a biologic
response to interference in the physiologic equilibrium of the
dentofacial complex by an externally applied force (2), which generates two different strains
in the PDLs, compression and tension. At the compression site, the
force that is generated by the root against the alveolar bone
induces bone resorption. At the tension site, PDL fibers are
stretched and bone tissue is formed (3,4).
Alveolar bone formation results from complex events that involve
the differentiation of osteoblast precursor cells from primitive
bone mesenchymal stem cells (BMSCs), maturation of osteoblasts, and
matrix formation, followed by its mineralization (4–6).
Nevertheless, several lines of evidence (7,8)
suggest that cells of the osteoblastic lineage are involved in
osteoclast formation. However, the role played by orthodontic
stretch regulating osteoclastogenesis induced by BMSCs during
initial osteoblastic differentiation in alveolar bone remains to be
determined.
Research in osteoclast differentiation has been
greatly advanced since the identification of the receptor activator
of nuclear factor-κB ligand (RANKL) as an osteoclast
differentiation factor. RANKL was identified as a key cytokine that
binds to its receptor on osteoclast progenitors, and then
stimulates the activity and recruitment (7,8).
Osteoprotegerin (OPG) has been identified as a competitive receptor
of RANKL that may prevent the activation of RANK resulting in
decreased osteoclast activity (9).
This association between osteoclasts and osteoblasts in bone
metabolism is greatly affected by the balance between RANKL and
OPG, which modulates the level of bone resorption on bone
surfaces.
In a previous study, we found that exposure to
intermittent traction stretch induced osteoblastic differentiation
of BMSCs (10,11). However, whether stretch-promoted
osteoblastic differentiation of BMSCs also affected
osteoclastogenesis remains to be elucidated. The present study was
designed to investigate the association of osteoclastogenesis with
osteoblastic differentiation of BMSCs. By evaluating the effects of
mechanical stretch on BMSCs insight may be gained into the
mechanisms of both bone regeneration and orthodontic movement in
vivo.
Materials and methods
Isolation and identification of
rBMSCs
The tibia and femur were isolated from 160±10 g,
male Sprague-Dawley rats (Shanghai SLAC Experimental Animal Center,
Shanghai, China). The bone marrow was flushed out with α-minimal
essential medium (α-MEM; Hyclone, Thermo, USA) supplemented with
100 U/ml penicillin, and 100 μg/ml streptomycin (Hyclone).
To remove the blood cells, the whole washouts were collected and
centrifuged at 1,800 × g for 10 min. The precipitate was mixed with
complete α-MEM supplemented with 10% fetal bovine serum (FBS,
Hyclone), 100 U/ml penicillin and 100 μg/ml streptomycin and
then plated into a culture flask maintained at 37°C in 5%
CO2. Non-adherent cells were removed by changing the
medium every 3 days for ∼10 days. When large colonies formed and
became confluent, the primary rat BMSCs were trypsinized with 10%
trypsin-EDTA (Hyclone) and passaged. Analysis of cell surface
molecules was performed on passages 2 using flow cytometry for
CD31, CD44 and CD90. BMSCs from passages 2–5 were used for
subsequent experiments.
Alkaline phosphatase (ALP) quantification
assay
To observe the osteoblastic differentiation
characteristic of BMSCs, cells were cultured either in the growth
or osteogenic medium, comprising Dulbecco’s modified Eagle’s medium
(DMEM, Hyclone) supplemented with 10% FBS, 10 mM
β-glycerophosphate, 100 nM dexamethasone, and 50 M ascorbic acid
(all from Sigma-Aldrich, St. Louis, MO, USA). BMSCs were rinsed
twice with ice-cold PBS, scraped from the dishes and suspended in
ddH2O. This was followed by three cycles of freezing and
thawing. ALP activity was determined at 405 nm using p-nitrophenyl
phosphate (pNPP) (Sigma-Aldrich) as the substrate. A 50 μl
of sample was mixed with 50 μl of pNPP (1 mg/ml) in 1 M
diethanolamine buffer containing 0.5 mM MgCl2 (pH 9.8)
and incubated at 37°C for 15 min on a bench shaker. The reaction
was stopped by the addition of 200 μl of 2 M NaOH per 200
μl of reaction mixture. Total protein content was determined
by the BCA method using a protein assay kit (Pierce, Rockford, IL,
USA). ALP activity was calculated as nmol p-nitrophenol per minute
per mg protein, and presented as fold changes over the un-induced
group at the respective time points. The experiments were conducted
in triplicate.
Mechanical tension load application
BMSCs were plated at a density of 1×105
cells/ml (unless stated otherwise) in 2 ml of medium on six-well
flexible silicone rubber BioFlex™ plates coated with collagen type
I (Flexcell International Corp., Hillsborough, NC, USA). The cells
were cultured for 48 h to allow them to attach and reach 80–90%
confluency, at which time the growth medium was replaced, and
mechanical strain was applied. A cyclic mechanical strain with a
0.5 Hz sinusoidal curve set at 10% elongation was applied twice
every day, for 4 h for each treatment, using a FX-4000T™ Flexcell
Tension Plus™ unit (Flexcell International Corp.). The cultures
were incubated in a humidified atmosphere at 37°C and 5%
CO2 while stretching. The BMSCs were collected after 1,
3, 5 and 7 days of stretch stimulation.
ALP staining
The presence of ALP in the cell layers was assessed
according to the manufacturer’s instructions (Beyotime, Suzhou,
China). The BMSCs from the control and stretch treatment groups
were rinsed with PBS three times and fixed with 4% paraformaldehyde
for 10 min. The fixed cells were soaked in 0.1% naphthol AS-MX
phosphate and 0.1% fast red violet LB salt in 56 mM
2-amino-2-methyl-1, 3-propanediol for 45 min at 37°C, washed with
ddH2O, and then visualized with a digital camera.
Real-time PCR
The total RNA of the cells was isolated using the
TRIzol reagent (Invitrogen, Carlsbad, CA, USA), according to the
manufacturer’s instructions. The RNA concentrations were determined
using a NanoDrop spectrophotometer (Thermo Scientific, Wilmington,
DE). Complimentary DNA (cDNA) was synthesized by means of a cDNA
Synthesis Reverse Transcription kit (Fermentas, Thermo Scientific,
Wilmington, DE, USA). Real-time PCR was performed using an ABI
7900HT system with SYBR Premix Ex Taq™ (Takara, Dalian,
China), according to the manufacturer’s instructions. The
conditions of the real-time PCR were as follows: denaturation at
95°C for 10 sec; 50 cycles at 95°C for 10 sec and 60°C for 30 sec;
and a final dissociation stage (95°C for 5 min) was added at the
end of the amplification procedure. β-actin was used as an internal
control. The data were analyzed using the comparative Ct
(2−ΔΔCt) (Pfaffl, 2001) method and expressed as a fold
change respective to the control. Each sample was analyzed in
triplicate. The primer sequences used in this study are listed in
Table I.
 | Table IList of primers used and their
respective forward and reverse sequences. |
Table I
List of primers used and their
respective forward and reverse sequences.
| Gene | Sequence |
|---|
| β-actin | F:
5′-GTAAAGACCTCTATGCCAACA-3′ |
| R:
5′-GGACTCATCGTACTCCTGCT-3′ |
| OPG | F:
5′-TTGAAATGGCAGTTGATTCCTTT-3′ |
| R:
5′-TATCCTCTTTCTCAGGGTGCTTG-3′ |
| RANK ligand | F:
5′-CATCGGGTTCCCATAAAG-3′ |
| R:
5′-GAAGCAAATGTTGGCGTA-3′ |
Statistical analysis
Data are presented as the mean ± SD. Significant
differences between non-load and stretch groups (shown in Figs. 2–4)
were determined using Student’s t-test. P<0.05 denotes
statistical significance.
Results
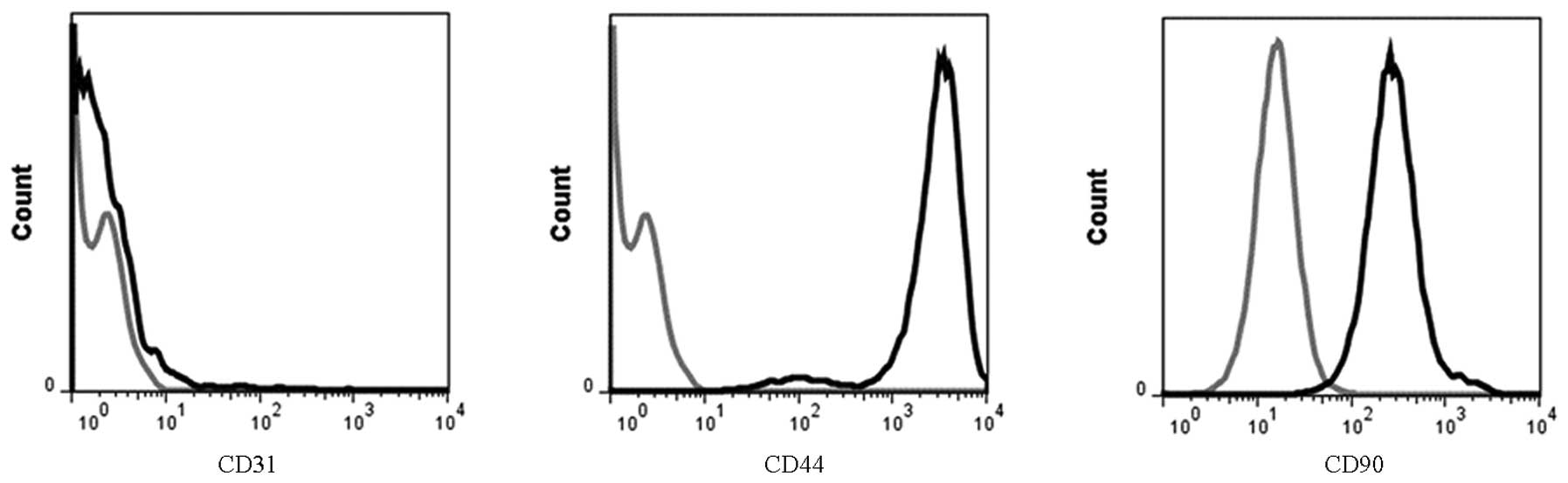
Identification of isolated BMSCs
To investigate the characterization of BMSCs, the
expression of antigens on the cell surface was detected by flow
cytometry. The positive rates of CD44 and CD90 were 72.0% and
70.8%, respectively, but the positive rate of CD31 was only 1.5%
(Fig. 1).
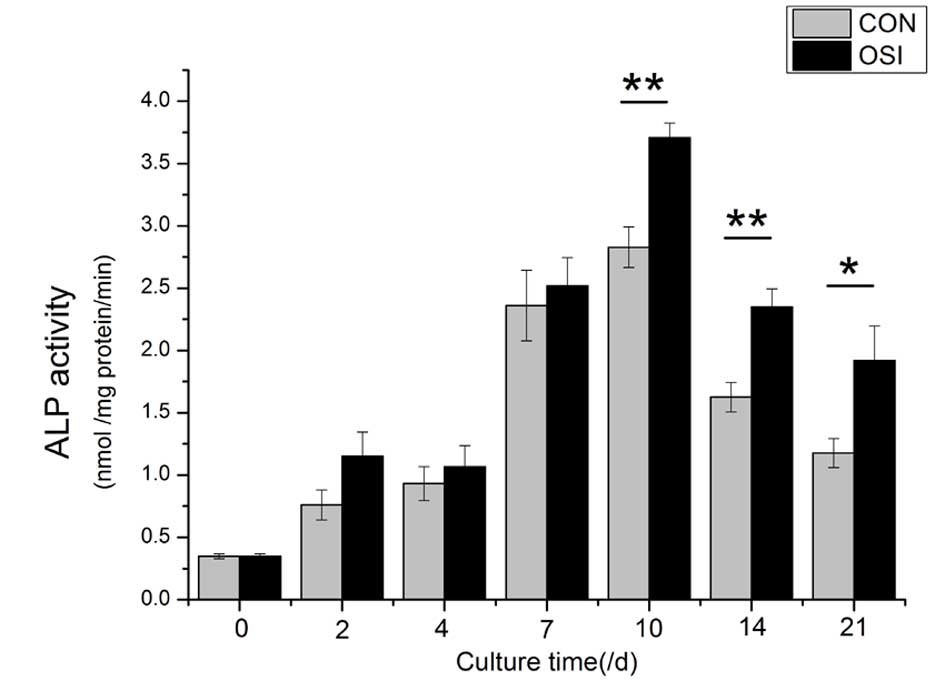
The quantitative analysis of ALP activity indicated
that the cells cultured in the osteogenic medium demonstrated
stronger ALP activity than those cultured in the growth medium, and
reached a peak at Day 10 (Fig.
2).
Intermittent traction stretch promotes
osteoblastic differentiation of BMSCs
In a previous study (10), we applied intermittent traction
stretch stimulation to BMSCs and examined whether or not the
progress of osteogenesis was switched on, as assessed by the
presence of the osteoblast differentiation markers ALP, Collagen I,
and OCN at the mRNA level. The results of those experiments showed
that the stretch induces a time-dependent increase in the
expression of osteoblastic genes and promoted the ALP activity
(Fig. 3) at 7 days.
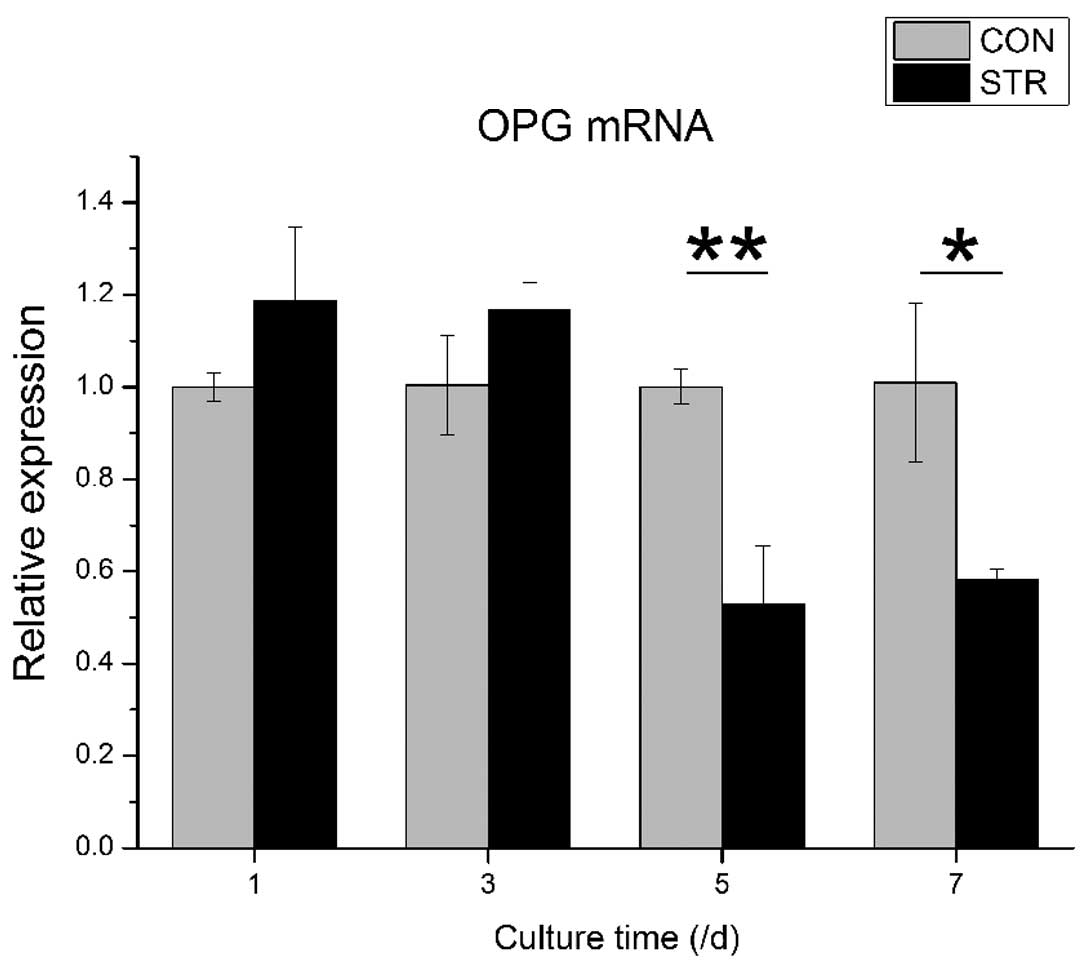
Expression of RANKL and OPG genes is
affected by intermittent traction stretch
When BMSCs were exposed to intermittent traction
stretch, the OPG mRNA level was increased slightly during the first
3 days. However, the OPG mRNA level decreased significantly at day
5 (P<0.01). At day 7, the OPG mRNA level of the stretching group
was increased again, but remained lower than the non-loading group
(P<0.05, Fig. 4).
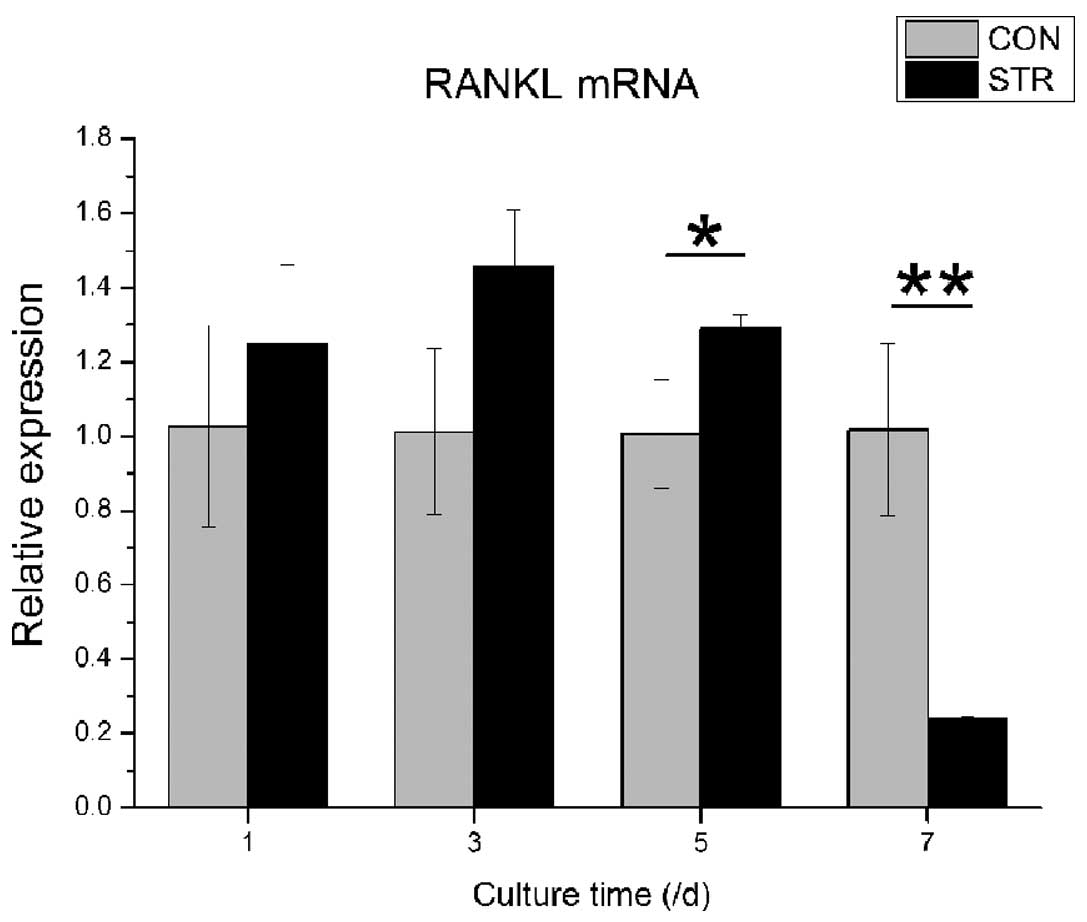
Dynamic stretch also showed different effects of
RANKL mRNA expression at various time points. In contrast to OPG,
RANKL gene expression was increased from the 1st day of loading to
the 5th day, followed by a decrease to approximately one fifth of
the non-loading group (P<0.05, Fig.
5).
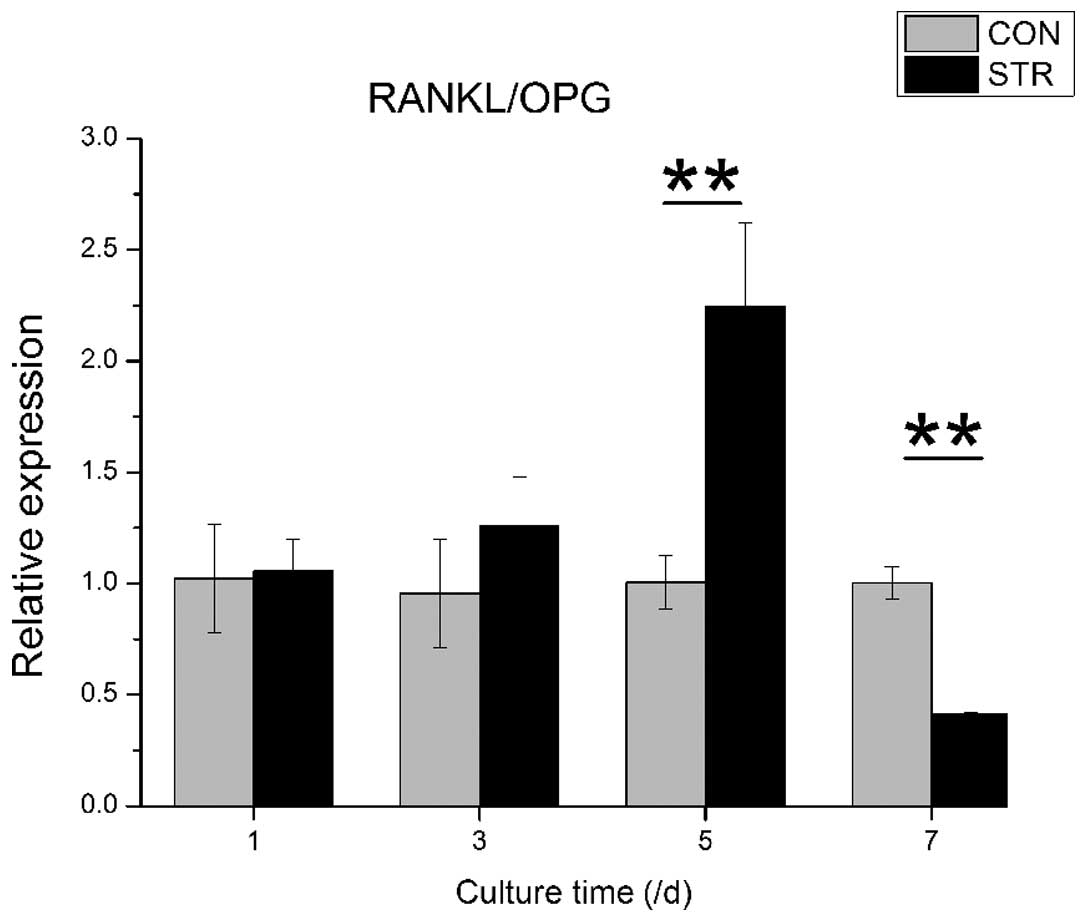
RANKL/OPG ratio of BMSCs is affected by
intermittent traction stretch
The RANKL/OPG ratio was increased gradually when
BMSCs were exposed to dynamic intermittent traction stretch, and
reached a climax (2.2-fold) at Day 5 (P<0.01). Of note is that
the ratio was reduced to more than half at Day 7 (P<0.01,
Fig. 6).
Discussion
Our previous studies have demonstrated that
intermittent traction stretch could promote osteoblastic
differentiation of BMSCs via the ERK-activated Cbfa1 signaling
pathway (10). In the present
study, we have shown that intermittent traction stretch affects the
cytokine expression and production regulating osteoclastogenesis
during the osteoblastogenesis process of BMSCs. Although several
studies (10–12) have demonstrated that mechanical
stress induces osteoblastic differentiation of osteoblastic cell
lines, this is the first study to investigate the effects of
intermittent traction stretch on osteoclastic-related cytokine
production in BMSCs. Our results showed that intermittent traction
stretch induced RANKL and OPG mRNA expression in a time-dependent
manner and altered the RANKL/OPG ratio at different time points,
initially upregulating the RANKL/OPG ratio and then downregulating
it. This suggests that in the process of osteoblastogenesis of
BMSCs induced by mechanical stretch, osteoclastic-related cytokine
is simultaneously activated leading to the initiation of osteoclast
activation. Nevertheless, osteoblastic differentiation of BMSCs was
continuously enhanced by appropriate mechanical stretch, the
osteoclastic-related cytokine was downregulated and then bone was
deposited in the tension side.
Different behaviors of RANKL and OPG expression in
various cells under different mechanical conditions have been
reported. In the human osteoblastic cell line MG-63, OPG production
increased subsequent to exposure of a cyclic strain; however, RANKL
expression was not altered (12). A
tensile strain upregulated OPG expression of mouse osteoblastic
MC3T3-E1 cells and downregulated RANKL expression. Moreover,
hydrostatic pressure changed the ratio of RANKL/OPG in favor of
RANKL both at the mRNA and protein levels in a magnitude- and
time-dependent manner (13). These
studies all demonstrated that mechanical stress suppressed
osteoclastogenesis and inhibited bone resorption. However, the
exact relative behaviors of RANKL and OPG differ slightly, which is
possibly due to the differences in mechanical conditions (strength,
time and quality) and types of cells in vitro. Of note is
that during tooth movement in vivo, mechanical stress
stimulates osteocytes to express OPN in order to initiate bone
remodeling (14–16), resulting in bone resorption and
apposition on opposite sides of the tooth.
Similarly, it has been previously suggested that in
response to mechanical forces, osteocytes regulate the recruitment
of osteoclasts to sites of bone resorption by inducing the
expression of RANKL by osteoblastic cells in the local
micro-environment (17). This
finding mainly consists of localization of RANKL-expression cells
in bone sections (18,19) and detection of RANKL expression in
cells isolated from rodent calvarias, which are rich in osteoblast
progenitors (7). However, the
results revealed in those studies are inconsistent with each other.
For instance, some studies detected RANKL in osteocytes while
others did not detect RANKL. Moreover, calvarial preparations are
heterogeneous and contain other cell types in addition to
osteoblast precursors (20,21), some of which may be sources of
RANKL. Furthermore, the conditional ablation of
osteocalcin-expressing cells in mice did not alter osteoclast
number and function (22),
indicating that mature osteoblasts and their immediate precursors
may not be an essential source of RANKL in bone. Thus, while
osteoblast precursors may be an important source of RANKL, this has
not yet been proven experimentally. In addition, increased bone
formation does not result in increased bone resorption in available
models (23,24). These latter studies dissociate the
well-elucidated role of cells of the osteoblastic lineage in
osteoclastogenesis from the potential influence of bone formation
on bone resorption. Taken together, this evidence suggests that
cells that support osteoclast formation may share a common
progenitor with cells that differentiate into matrix-synthesizing
osteoblasts. If this were found to be the case, it would provide
strong support for the hypothesis that the balance in bone turnover
is maintained, in part, by associating osteoclast and osteoblast
differentiation.
The bone remodeling cycle could be altered by a
number of stimulations, including mechanical stress. Our results
indicate that mechanical stretch induced osteoclastogenesis to a
certain extent. In the bone remodeling process, osteoclastogenesis
and bone resorption occur initially and subsequently bone formation
proceeds. In this study, we investigated the effect of mechanical
stretch on RANKL and OPG production in BMSCs over a period of time.
Our results seem to represent the initial and turning stage of the
bone remodeling cycle. Moreover, in the experiment with a model
loaded with long-term mechanical stress (25,26),
osteogenic marker upregulation was observed in osteoblasts loaded
with mechanical stress compared with that for non-loaded
osteoblasts.
A new mode of osteoclast-osteoblast communication
has been identified in which Sema4D secreted by osteoclasts acts as
a ligand and binds its receptor, Plexin-B1, on osteoblasts, thus
suppressing their differentiation (27). The osteoclasts were shown to highly
express Sema4D, and that expression is further increased during
RANKL-induced osteoclastogenesis. When the number of osteoblastic
cells increases, more RANKL is available to stimulate
osteoclastogenesis, which in turn produces more Sema4D to reduce
osteoblast differentiation, thereby serving as a negative feedback
loop to balance the supply of osteoclasts and osteoblasts. As
Sema4D plays an essential role in bone remodeling, it is necessary
to investigate Sema4D for analysis of the molecular mechanism in
bone remodeling induced by mechanical stress.
In conclusion, we have demonstrated that
intermittent traction stretch regulated the production of RANKL in
a time-dependent manner and initially changed the RANKL/OPG ratio
in favor of RANKL in BMSCs down-regulating the RANKL/OPG ratio
later. These results suggest that BMSCs play an important role in
cytokine production in response to mechanical stress and that
orthodontic force may support the maintenance of mandible bone
homeostasis by activating bone remodeling through
osteoclastogenesis in vivo.
Acknowledgements
This study was supported by the
National Nature Science Foundation of China (grant nos. 30901698
and 10972142), Collaborative Foundation of Medical and Engineering
Science of Shanghai Jiaotong University (YG2012MS40), Key Basic
Research Foundation of the Shanghai Committee of Science and
Technology, China (12JC1405700), Program for Innovative Research
Team of Shanghai Municipal Education Commission.
References
|
1
|
Heinonen A, Sievanen H, Kyrolainen H, et
al: Mineral mass, size, and estimated mechanical strength of triple
jumpers’ lower limb. Bone. 29:279–285. 2001.PubMed/NCBI
|
|
2
|
Dunphy L: Contemporary orthodontics, 5th
edition. Br Dent J. 213:2582012. View Article : Google Scholar
|
|
3
|
Krishnan V and Davidovitch Z: Cellular,
molecular, and tissue-level reactions to orthodontic force. Am J
Orthod Dentofacial Orthop. 129:e1–e32. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Masella RS and Meister M: Current concepts
in the biology of orthodontic tooth movement. Am J Orthod
Dentofacial Orthop. 129:458–468. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Davidovitch Z: Tooth movement. Crit Rev
Oral Biol Med. 2:411–450. 1991.PubMed/NCBI
|
|
6
|
Koka S and Reinhardt RA: Periodontal
pathogen-related stimulation indicates unique phenotype of primary
cultured human fibroblasts from gingiva and periodontal ligament:
implications for oral health disease. J Prosthet Dent. 77:191–196.
1997. View Article : Google Scholar
|
|
7
|
Lacey DL, Timms E, Tan HL, et al:
Osteoprotegerin ligand is a cytokine that regulates osteoclast
differentiation and activation. Cell. 93:165–176. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Teitelbaum SL: Osteoclasts: what do they
do and how do they do it? Am J Pathol. 170:427–435. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu Y, Zhang X, Zhang P, et al:
Intermittent traction stretch promotes the osteoblastic
differentiation of bone mesenchymal stem cells by the
ERK1/2-activated Cbfa1 pathway. Connect Tissue Res. 53:451–459.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang P, Wu Y, Jiang Z, et al: Osteogenic
response of mesenchymal stem cells to continuous mechanical strain
is dependent on ERK1/2-Runx2 signaling. Int J Mol Med.
29:1083–1089. 2012.PubMed/NCBI
|
|
12
|
Saunders MM, Taylor AF, Du C, et al:
Mechanical stimulation effects on functional end effectors in
osteoblastic MG-63 cells. J Biomech. 39:1419–1427. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yamamoto K, Yamamoto T, Ichioka H, et al:
Effects of mechanical stress on cytokine production in
mandible-derived osteoblasts. Oral Dis. 17:712–719. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fujihara S, Yokozeki M, Oba Y, et al:
Function and regulation of osteopontin in response to mechanical
stress. J Bone Miner Res. 21:956–964. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kuroda S, Balam TA, Sakai Y, et al:
Expression of osteopontin mRNA in odontoclasts revealed by in situ
hybridization during experimental tooth movement in mice. J Bone
Miner Metab. 23:110–113. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Terai K, Takano-Yamamoto T, Ohba Y, et al:
Role of osteopontin in bone remodeling caused by mechanical stress.
J Bone Miner Res. 14:839–849. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kartsogiannis V, Zhou H, Horwood NJ, et
al: Localization of RANKL (receptor activator of NF kappa B ligand)
mRNA and protein in skeletal and extraskeletal tissues. Bone.
25:525–534. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ikeda T, Utsuyama M and Hirokawa K:
Expression profiles of receptor activator of nuclear factor kappaB
ligand, receptor activator of nuclear factor kappaB, and
osteoprotegerin messenger RNA in aged and ovariectomized rat bones.
J Bone Miner Res. 16:1416–1425. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Silvestrini G, Ballanti P, Patacchioli F,
et al: Detection of osteoprotegerin (OPG) and its ligand (RANKL)
mRNA and protein in femur and tibia of the rat. J Mol Histol.
36:59–67. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guenther HL, Hofstetter W, Stutzer A, et
al: Evidence for heterogeneity of the osteoblastic phenotype
determined with clonal rat bone cells established from transforming
growth factor-beta-induced cell colonies grown anchorage
independently in semisolid medium. Endocrinology. 125:2092–2102.
1989. View Article : Google Scholar
|
|
21
|
Kalajzic I, Staal A, Yang WP, et al:
Expression profile of osteoblast lineage at defined stages of
differentiation. J Biol Chem. 280:24618–24626. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Corral DA, Amling M, Priemel M, et al:
Dissociation between bone resorption and bone formation in
osteopenic transgenic mice. Proc Natl Acad Sci USA. 95:13835–13840.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ducy P, Amling M, Takeda S, et al: Leptin
inhibits bone formation through a hypothalamic relay: a central
control of bone mass. Cell. 100:197–207. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jochum W, David JP, Elliott C, et al:
Increased bone formation and osteosclerosis in mice overexpressing
the transcription factor Fra-1. Nat Med. 6:980–984. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu J, Zhao Z, Zou L, et al:
Pressure-loaded MSCs during early osteodifferentiation promote
osteoclastogenesis by increase of RANKL/OPG ratio. Ann Biomed Eng.
37:794–802. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu J, Zhang X, Wang C, et al: Different
magnitudes of tensile strain induce human osteoblasts
differentiation associated with the activation of ERK1/2
phosphorylation. Int J Mol Sci. 9:2322–2332. 2008. View Article : Google Scholar
|
|
27
|
Negishi-Koga T, Shinohara M, Komatsu N, et
al: Suppression of bone formation by osteoclastic expression of
semaphorin 4D. Nat Med. 17:1473–1480. 2011. View Article : Google Scholar : PubMed/NCBI
|