Introduction
Pediatric patients may be at risk for later learning
and behavioral impairment when exposed to general anesthesia and
surgery (1–4). This is consistent with studies in
preclinical experiments by Yon et al (5). In the study, rats were exposed at
postnatal day 7 to compound anesthetic agents (midazolam,
isoflurane and nitrous oxide) and identified neuronal apoptosis in
the brain and persistent defects of memory and learning when the
rats were juvenile. In addition, these rats also showed the
impairment of spatial reference and working memory as they grew up.
Similar impairment of learning and memory induced by neonatal
anesthesia was also reported for adult patients and other animals
(1–4).
However, the exact mechanism underlying the neonatal anesthesia
remains unclear.
Rho kinase 2 (ROCK2), one of the members of the
family of serine/threonine protein kinase, is a downstream effector
of Rho protein A (RhoA) (6). RhoA
regulates the reorganization of cytoskeleton protein actin by
controlling ROCK2, affecting cell migration, apoptosis, gene
transcription, nerve regeneration and other biological processes
(7,8).
RhoA/Rho-kinase is an important pathway underlying neuronal injury
in rats of experimental spinal cord injury (9). In addition, the Rho/Rho-kinase pathway is
associated with the modulation of N-methyl-D-aspartate
(NMDA) receptor function and the NMDA receptor is associated with
learning and memory processing (10).
In a previous study, Lemkuil et al (11) demonstrated that isoflurane induced
neurotoxicity of mouse neurons by activating
p75NTR-RhoA, and inhibiting activation of RhoA
attenuated isoflurane-induced impairment. Pearn et al
(12) also reported that
propofol-induced apoptosis is involved in p75NTR and
RhoA kinase activation in developing neurons in vivo and
in vitro. These suggested that RhoA played important roles
in the neurotoxicity of anesthetics. However, it remains unclear
whether ROCK2, the key downstream molecule of the RhoA signals,
mediates the neurotoxicity of anesthetics cognitive impairment of
adult rats with neonatal exposure of sevoflurane.
The present study investigated i) the effects of
sevoflurane exposure of rats at postnatal days 7–9 on their
learning and memory at adulthood; and ii) the association of the
cognitive dysfunction induced by sevoflurane to expression changes
of RhoA and ROCK2. Sevoflurane exposure at postnatal days 7–9
impaired the learning and memory of rats after they grew up.
Corresponding to the behavioral changes of rats, hippocampal RhoA
and ROCK2 increased; and inhibiting ROCK2 expression reversed the
cognitive dysfunction induced by sevoflurane exposure.
Materials and methods
Animals
Fifty-six Sprague-Dawley rats (7 days old) were
obtained from the Experimental Animal Center of The Third Xiangya
Hospital of Central South University (Changsha, China). The
institutional guidelines regarding animal safety were strictly
followed and authorized by the Institutional Animal Care. The room
temperature was maintained at 22–24˚C under suitable humidity. The
rats were divided into four groups (S1, S3, F and C) randomly, with
14 in each group. The rats in group S1 inhaled 2% sevoflurane and
80% oxygen for 2 h. Rats in the S3 group were exposed to
sevoflurane three times from postnatal day 7 (2 h/time/day). The F
group rats were pretreated with ROCK2 inhibitor fasudil
hydrochloride (10 mg/kg, HY-10341; MedChem Express LLC, Princeton,
NJ, USA) and 2 h later were exposed to sevoflurane three times from
postnatal day 7 (2 h/time/day). The rats in group C inhaled 80%
oxygen. All the rats were placed in a self-made organic glass box
(30×40×30 cm) with soda lime in the bottom. The multi-function
monitor (Datex-Ohmeda, Helsinki, Finland) was used to monitor the
concentration of sevoflurane, O2 and CO2.
When experimental models were completed for 3 h, rats were
sacrificed by decapitation and the hippocampus was removed to
assess the expression of RhoA, ROCK2 and cleaved caspase-3
(Cl-Csp3) by western blotting. The neurobehavioral tests of the
Morris water maze (MWM) at P45-50 were used to examine the memory
function. Latency to locate the target platform, swimming distance
and speed in target zone were determined and compared in all the
groups.
Western blotting
The protein concentration of samples was determined
using a bicinchoninic acid protein assay kit (Wellbio, Changsha,
China) according to the manufacturer's instructions. Equal amounts
of protein samples (/lane) were loaded and separated by sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and transferred
to polyvinylidene fluoride membranes. Membranes were blocked with
10% skimmed milk in phosphate-buffered saline with Tween-20 buffer
for 2 h and subsequently incubated with primary antibodies [rabbit
anti-RhoA polyclonal antibody (1:600; cat. no. 10749-1-AP;
Proteintech, Wuhan, Hubei, China), rabbit anti-ROCK2 polyclonal
antibody, 1:200; cat. no. sc5561; rabbit anti-cleaved caspase3
polyclonal antibody (1:400; cat. no. sc22139; Santa Cruz
Biotechnology, Santa Cruz, CA, USA) and anti-β-actin (1:2,000; cat.
no. KGAA001-4; KeyGen BrioTech, Nanjing, China) overnight at 4˚C.
After three washes, membranes were incubated with the secondary
antibody [goat anti-rabbit IgG (H+L) (1:2,000; cat. no. KGAA35;
KeyGen BioTech)] at room temperature for 2 h. Finally,
visualization of the proteins was accomplished by ECL detection
reagents (Advansta Corp., Menlo Park, CA, USA). The images were
developed on luminescent image analyzer (ImageQuant 350 Capture; GE
Healthcare Life Sciences, Shanghai, China) and quantified by
densitometry (Beckman Coulter, Inc., Pasadena, CA, USA). Relative
expression levels of protein were normalized by the ratio of the
target protein (ROCK2, RhoA and Cl-Csp3) to β-actin.
MWM test
The MWM test was used to evaluate the learning and
memory of rats. A computerized video track system (Logitech,
Suzhou, China) was used to record the movement of the rats in the
water maze by following a previous method (13). Briefly, a transparent circular platform
was placed below the water surface of the southeast quadrant in a
circular black pool. During the training, rats were first placed on
the platform for 30 sec, and subsequently were put into the water
facing the tank wall. The maximum trial time was 60 sec, following
a relaxation of 20 sec on the platform. When a rat could not locate
the platform within 60 sec, it was guided to the platform and
remained there for 30 sec. All the rats were trained for 5 days
with three trials/day. Following training, the memory of the rats
was evaluated by the percentage of searching time and distance in
the targeted area.
Statistical analysis
Water maze data are presented as mean ± standard
error of the mean and were analyzed using analysis of variance for
repeated measures followed by the least significant difference
test. Western blotting data are presented as mean ± standard
deviation and were analyzed using a Student's t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Sevoflurane exposure at postnatal day
7–9 impairs the learning and memory of rats after they grew up
In the training, the latency and distance of rats in
the S3 group at postnatal days 45–50 (P45-50) were significantly
longer than that of the C group (P<0.05, respectively). There
was no significant difference of the latency and distance between
the C and S1 groups (P>0.05, respectively) (Fig. 1A and B). In all the groups, the
swimming speed did not show a significant difference on all the
days (P>0.05) (Fig. 1C). In the
probe trial test, the percentage of searching time and distance in
target quadrant in the S3 group was evidently less than that of the
C (P<0.05) and S1 groups (P<0.05). The percentage of
searching time and distance in target quadrant in the S1 group were
not different from that of the C group (P>0.05) (Fig. 1D-F). These suggested that sevoflurane
exposure at postnatal days 7–9 dose-dependently impaired the
learning and memory of rats after they grew up.
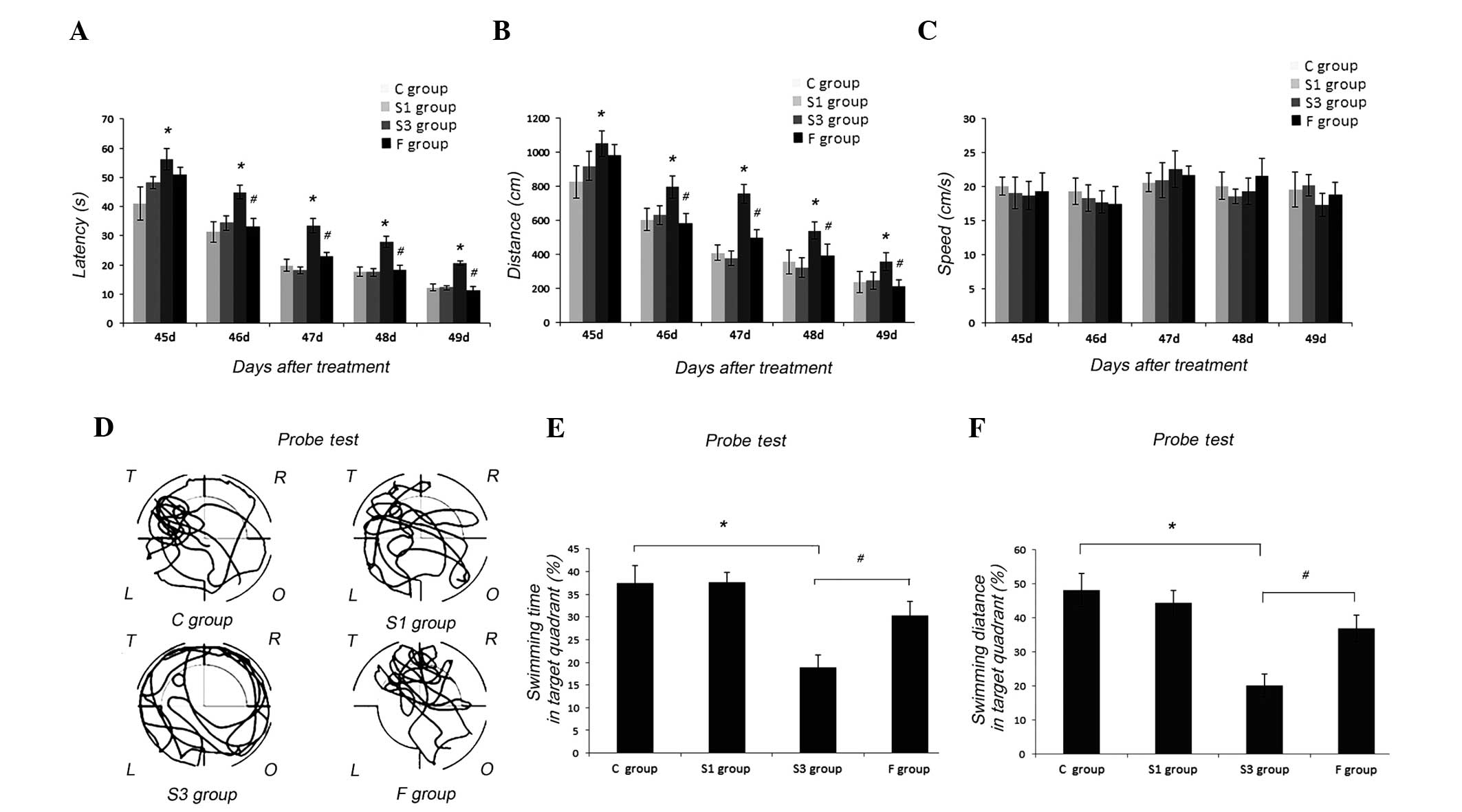 | Figure 1.Sevoflurane exposure at postnatal days
7–9 impaired the learning and memory of P45-50 rats in the Morris
water maze (MWM) test. (A) In the place navigation test, the
latency of the S3 group was longer than that of the C and F groups
(*P<0.05, C vs. S3 group at P45-49; #P<0.05, S3
vs. F group at P46-49). There was no significant difference of
latency between the C and S1 groups (P>0.05, respectively). (B)
The distance of the S3 group was longer than that of the C and F
groups (*P<0.05, C vs. S3 group at days 45–49;
#P<0.05, S3 vs. F group at P46-49). There was no
significant difference of the distance between the C and S1 groups
(P>0.05, respectively). (C) The swimming speed had no difference
in all the groups (P>0.05). (D) Representative routes of the C,
S1, S3 and F groups in the probe test on P50. T: target quadrant;
R, O, L quadrants: right, opposite or left of the target quadrant.
(E) In the probe trial test, the percentage of searching time in
the target quadrant of the S3 group was clearly less than that of
the C and F groups (*P<0.05, C vs. S3 group;
#P<0.05, S3 vs. F group). The percentage of searching
time in the target quadrant of the S1 group was not different from
that of the C group (P>0.05). (F) In the probe trial test, the
percentage of searching distance in the target quadrant of the S3
group was clearly less than that of the C and F groups (*P<0.05,
C vs. S3 group; #P<0.05, S3 vs. F group). The
percentage of searching distance in the target quadrant of the S1
group was nott different from that of the C group (P>0.05). Data
are expressed as mean ± standard error of the mean. |
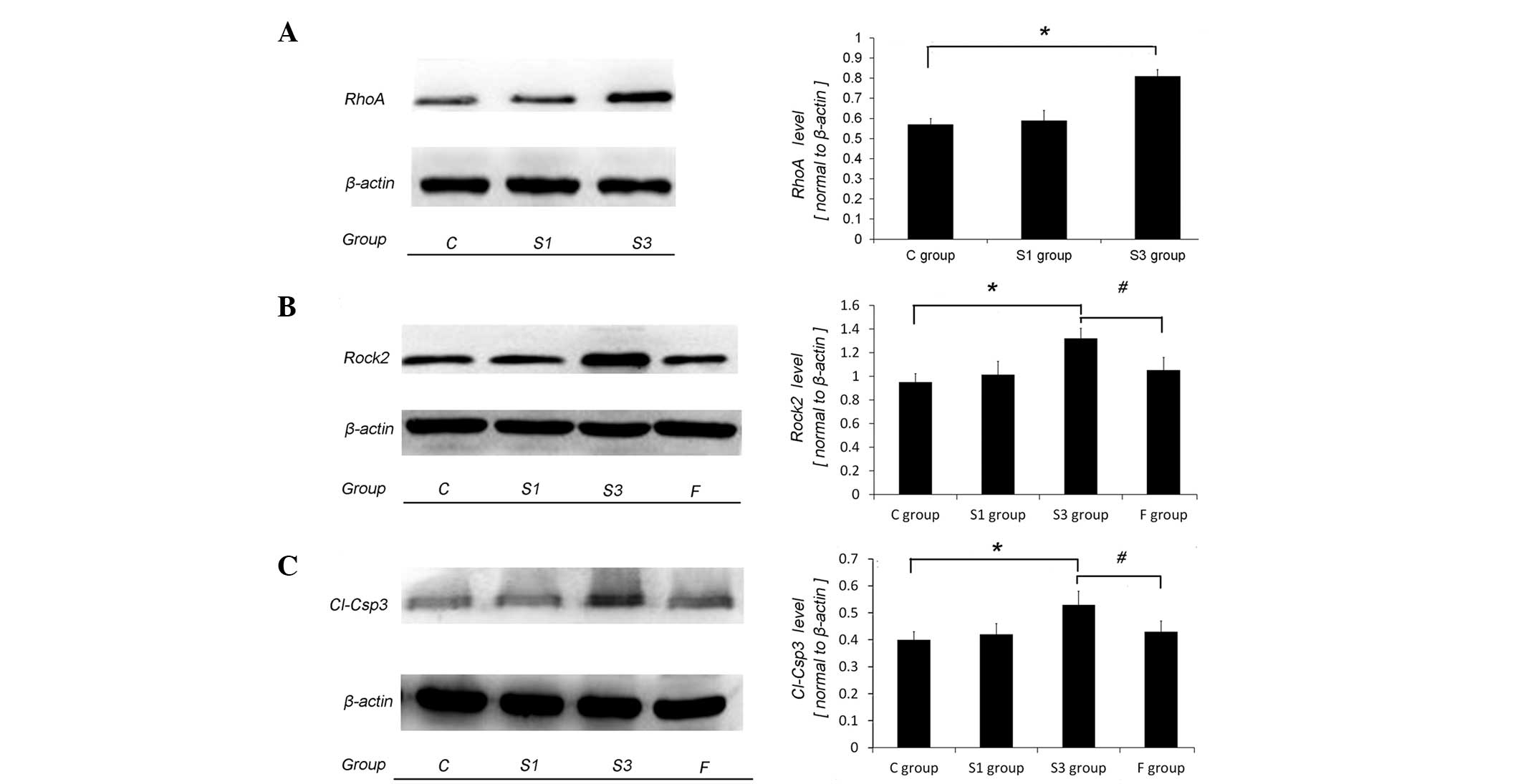
RhoA and ROCK2 upregulation of rat
hippocampus is involved in memory dysfunction induced by postnatal
exposure of sevoflurane
Corresponding to the behavioral changes of the S3
group, the levels of RhoA, ROCK2 and Cl-Csp3 in the hippocampus of
the S3 group significantly increased, compared to that of the C and
S1 groups (P<0.05) (Fig. 2A-C). The
ROCK2 inhibitor, fasudil hydrochloride, in the F group clearly
decreased the expression of ROCK2 and Cl-Csp3, and shortened the
latency in the training (P<0.05, P46-49 respectively) and probe
test (P<0.05), compared to that of the S3 group. In addition,
there was no significant difference in the expression of ROCK2 and
Cl-Csp3, and the latency in the place navigation test and probe
test between the C and F groups (P>0.05). These suggested that
the increasing expression of RhoA and ROCK2 in the hippocampus was
an important mechanism underlying the cognitive dysfunctions
induced by sevoflurane exposure.
Discussion
The aim of the present study was to investigate
whether ROCK2 played roles in the cognitive dysfunction of adult
rats exposed to sevoflurane during the postnatal early stage.
Sevoflurane exposure during the postnatal early stage
dose-dependently impaired the cognitive function of rats after they
grew up. Corresponding to the cognitive dysfunction of rats, the
level of RhoA, ROCK2 and Cl-Csp3 in the hippocampus in the
sevoflurane-treated group significantly increased, compared to the
control group. Additionally, ROCK2 inhibitor fasudil hydrochloride
decreased the level of ROCK2 and Cl-Csp3 in the hippocampus and
partly reversed the cognitive dysfunction in the
sevoflurane-treated rats. These showed that ROCK2 was closely
involved in the neurotoxicity of sevoflurane.
Neurotoxicity of anesthetics was well reported in
aged animals (13–15), natal animals (15) and cultured neurons (16). Yan et al (17) showed that the isoflurane-induced
decrease of nNOS is closely correlated with the cognitive
impairment in aged rats. In another study, Zheng et al
(18) found that sevoflurane use
during pregnancy may produce adverse effects on fetal and postnatal
rats. In the present study, 2% sevoflurane for 2 h at postnatal day
7 did not impair the cognitive function of rats when they grew up.
By contrast, 2% sevoflurane for 2 h at postnatal days 7–9 (2 h
sevoflurane/day) significantly impaired the cognitive function of
rats when they grew up. These results showed the dose-dependent
neurotoxicity of sevoflurane to the neurons. Consistent with our
data, Feng et al (19) found
that inhaling 2.3% sevoflurane for 6 h induced nerve cell death in
newborn rats. Shen et al (20)
found that rats in the growing stage (6 days of age) that inhaled
3% sevoflurane for 2 h at a time did not show cognitive impairment
and nerve inflammation, while those that inhaled 3% sevoflurane for
2 h for three days showed cognitive impairment and nerve
inflammation. Of note in the present study, corresponding to
dose-dependent neurotoxicity of sevoflurane to rats, 2% sevoflurane
of 2 h at postnatal day 7 did not induce clear expression changes
of hippocampal RhoA, ROCK2 and Cl-Csp3, but 2% sevoflurane of 2 h
at postnatal days 7–9 (2 h sevoflurane/day) significantly increased
the expression of hippocampal RhoA, ROCK2 and Cl-Csp3.
Additionally, when the expression of ROCK2 was inhibited by fasudil
hydrochloride, the expression of rat hippocampal ROCK2 and Cl-Csp3
decreased and rat cognitive dysfunction induced by sevoflurane
exposure also partly reversed. These showed the key roles of RhoA
and ROCK2 in sevoflurane neurotoxicity to natal rats. A previous
study showed that RhoA and ROCK2 regulated synaptic plasticity
(6,21).
Therefore, in the present study, we speculate that sevoflurane may
upregulate RhoA and ROCK2, damage synaptic plasticity, and
subsequently, induce impairment of learning and memory performance
of rats after they grew up. However, its specific mechanism remains
to be proved.
Acknowledgements
The present study was supported by the Hunan
Provincial Natural Science Foundation of China (grant no. 12JJ3110)
and the 125 Program of The Third Xiangya Hospital, Central South
University (Changsha, China).
References
|
1
|
McCann ME, Bellinger DC, Davidson AJ and
Soriano SG: Clinical research approaches to studying pediatric
anesthetic neurotoxicity. Neurotoxicology. 30:766–771. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wilder RT, Flick RP, Sprung J, Katusic SK,
Barbaresi WJ, Mickelson C, Gleich SJ, Schroeder DR, Weaver AL and
Warner DO: Early exposure to anesthesia and learning disabilities
in a population-based birth cohort. Anesthesiology. 110:796–804.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sun L: Early childhood general anaesthesia
exposure and neurocognitive development. Br J Anaesth. 105 (Suppl
1):i61–i68. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kalkman CJ, Peelen L, Moons KG, Veenhuizen
M, Bruens M, Sinnema G and de Jong TP: Behavior and development in
children and age at the time of first anesthetic exposure.
Anesthesiology. 110:805–812. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yon JH, Daniel-Johnson J, Carter LB and
Jevtovic-Todorovic V: Anesthesia induces neuronal cell death in the
developing rat brain via the intrinsic and extrinsic apoptotic
pathways. Neuroscience. 135:815–827. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wettschureck N and Offermanns S:
Rho/Rho-kinase mediated signaling in physiology and
pathophysiology. J Mol Med Berl. 80:629–638. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Negishi M and Katoh H: Rho family GTPases
as key regulators for neuronal network formation. J Biochem.
132:157–166. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Leong SY, Faux CH, Turbic A, Dixon KJ and
Turnley AM: The Rho kinase pathway regulates mouse adult neural
precursor cell migration. Stem Cells. 29:332–343. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sung JK, Miao L, Calvert JW, Huang L,
Louis Harkey H and Zhang JH: A possible role of RhoA/Rho-kinase in
experimental spinal cord injury in rat. Brain Res. 959:29–38. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nakazawa T, Watabe AM, Tezuka T, Yoshida
Y, Yokoyama K, Umemori H, Inoue A, Okabe S, Manabe T and Yamamoto
T: p250GAP, a novel brain-enriched GTPase-activating protein for
Rho family GTPases, is involved in the N-methyl-d-aspartate
receptor signaling. Mol Biol Cell. 14:2921–2934. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lemkuil BP, Head BP, Pearn ML, Patel HH,
Drummond JC and Patel PM: Isoflurane neurotoxicity is mediated by
p75NTR-RhoA activation and actin depolymerization. Anesthesiology.
114:49–57. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pearn ML, Hu Y, Niesman IR, Patel HH,
Drummond JC, Roth DM, Akassoglou K, Patel PM and Head BP: Propofol
neurotoxicity is mediated by p75 neurotrophin receptor activation.
Anesthesiology. 116:352–361. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He HJ, Wang Y, Le Y, Duan KM, Yan XB, Liao
Q, Liao Y, Tong JB, Terrando N and Ouyang W: Surgery upregulates
high mobility group box-1 and disrupts the blood-brain barrier
causing cognitive dysfunction in aged rats. CNS Neurosci Ther.
18:994–1002. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stratmann G, Sall JW, May LD, et al:
Isoflurane differentially affects neurogenesis and long-term
neurocognitive function in 60-day-old and 7-day-old rats.
Anesthesiology. 110:834–848. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shen X, Liu Y, Xu S, Zhao Q, Guo X, Shen R
and Wang F: Early life exposure to sevoflurane impairs adulthood
spatial memory in the rat. Neurotoxicology. 39:45–56. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang WY, Luo Y, Jia LJ, et al: Inhibition
of aberrant cyclin-dependent kinase 5 activity attenuates
isoflurane neurotoxicity in the developing brain.
Neuropharmacology. 77:90–99. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yan XB, Ouyang W, Li G and Duan KM:
Involvement of neuronal nitric oxide synthase in cognitive
impairment in isoflurane-treated rats. Neurosci Lett. 506:240–244.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zheng H, Dong Y, Xu Z, Crosby G, Culley
DJ, Zhang Y and Xie Z: Sevoflurane anesthesia in pregnant mice
induces neurotoxicity in fetal and offspring mice. Anesthesiology.
118:516–526. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Feng X, Liu JJ, Zhou X, Song FH, Yang XY,
Chen XS, Huang WQ, Zhou LH and Ye JH: Single sevoflurane exposure
decreases neuronal nitric oxide synthase levels in the hippocampus
of developing rats. Br J Anaesth. 109:225–233. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shen X, Dong Y, Xu Z, Wang H, Miao C,
Soriano SG, Sun D, Baxter MG, Zhang Y and Xie Z: Selective
anesthesia-induced neuroinflammation in developing mouse brain and
cognitive impairment. Anesthesiology. 118:502–515. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nakano-Kobayashi A, Kasri NN, Newey SE and
Van Aelst L: The Rho-linked mental retardation protein OPHN1
controls synaptic vesicle endocytosis via endophilin A1. Curr Biol.
19:1133–1139. 2009. View Article : Google Scholar : PubMed/NCBI
|