Introduction
Enterococcus faecalis (E. faecalis), a
Gram-positive bacterium, is a predominant cause of
hospital-associated infections (1).
E. faecalis is also involved in root canal infections
(2) and is considered a major pathogen
associated with endodontic treatment failure (3). Several virulence attributes that promote
bacterial colonization, the invasion of host tissues and the
evasion of host defense mechanisms are indicated in persistent
infections caused by E. faecalis (4). The virulence factors include lipoteichoic
acid (LTA), peptidoglycan, aggregation substances, cytolysin and
lytic enzymes. Among them, LTA is the key virulence factor due to
its major role in pathogenicity. It has been demonstrated that
E. faecalis LTA can stimulate leukocytes to release certain
mediators associated with inflammatory response (5) and plays a critical role in biofilm
formation (6). E. faecalis LTA
is also known to inhibit the repair mechanism of periapical bone by
decreasing the proliferation of human osteoblast-like cells and
inducing apoptosis (7).
E. faecalis LTA belongs to classical (type 1)
LTA. Its basic structure is comprised of a glycolipid moiety and
1,3-polyglycerolphosphate substituted at position C-2 with
D-alanine and kojibiose residues (8).
The alanyl esters with D-configured glycerophosphate influence the
antibiotic action, pathogenesis, adhesion, biofilm formation and
virulence of Gram-positive bacteria (6,9). The dlt
operon consists of 4 genes: dltA, dltB, dltC
and dltD, which are responsible for D-alanine synthesis
(10). The inactivation of the
dlt genes resulted in the absence of D-alanyl residues of
LTA and further decreased the virulence features of E.
faecalis (6).
Bacterial antigens are recognized by the pattern
recognition receptors (PRRs) of immune cells and subsequently
induce the production of pro-inflammatory cytokines. Toll-like
receptors (TLR) are ubiquitous PRRs that play a pivotal role in the
innate immune response (11). Of all
the TLRs, TLR2 is known to specifically interact with bacterial LTA
(12). Baik et al (5) found that E. faecalis LTA
stimulates RAW 264.7 cells to express tumor necrosis factor-α
(TNF-α). However, molecular mechanisms pertaining to the sensing
and signaling of clinically derived E. faecalis LTA and
human macrophages have not yet been studied. The cells of the THP-1
human monocytic cell line can differentiate into macrophages under
stimulation by phorbol 12-myristate 13-acetate (PMA) (13). The differentiated THP-1 macrophages are
widely used as human macrophages in vitro for studying
inflammatory diseases (14,15). In order to address this research gap,
highly purified, intact LTA was extracted from an E.
faecalis clinical strain and its effect on TNF-α production was
studied in differentiated THP-1 macrophages. Nuclear factor-κB
(NF-κB) and p38 mitogen-activated protein kinase (MAPK) signaling
pathways are activated by endodontic infectious contents (16). The present study also investigated the
involvement of NF-κB and MAPK signaling pathways in the production
of TNF-α.
Materials and methods
Bacteria and cell culture
E. faecalis P25RC was isolated from a
patient's root canal at the Peking University Hospital of
Stomatology (Beijing, China) by Zhu et al (17). E. faecalis P25RC was grown
anaerobically (90% N2, 5% CO2 and 5%
H2) overnight at 37°C without agitation in the
brain-heart infusion broth (Oxoid Ltd., Basingstoke, Hampshire,
UK). Once the bacterial growth reached the mid-exponential phase
(A600=0.5), 10 liters of the bacteria were harvested by
centrifugation and washed twice with phosphate-buffered saline
(PBS; pH 7.4) (Sigma-Aldrich, St. Louis, MO, USA). The bacteria
were examined by Gram staining and microscopy to rule out
contaminations. The bacteria were stored at −70°C until LTA
extraction.
THP-1 cells (ATCC® TIB-202™) were purchased from
American Type Culture Collection (ATCC, Manassas, VA, USA). THP-1
cells were maintained in RPMI-1640 medium (Life Technologies, Grand
Island, NY, USA) supplemented with 10% heat-inactivated fetal
bovine serum (HyClone, Logan, UT, USA), 50 µM 2-mercaptoethanol
(Sigma-Aldrich), 100 IU/ml penicillin and 100 µg/ml streptomycin
(Invitrogen Life Technologies, Carlsbad, CA, USA). The cells were
incubated at 37°C with 5% CO2. The experiments were
performed in 6-well plates at a density of 8×105
cells/ml. Before the LTA stimulation assays, THP-1 monocytes were
pretreated with 200 ng/ml PMA (Sigma-Aldrich) for 18 h to
differentiate them into macrophages, which facilitates the adhesion
of the cells (18).
Isolation and purification of LTA
The processes of isolation and purification of LTA
were performed as described previously (19). The bacteria were defrosted and
suspended in 100 ml of 0.1 M cold sodium citrate buffer (pH 4.7)
(Sigma-Aldrich). Following disruption by Vibra-Cell ultrasonication
(Sonic and Materials, Inc., Danbury, CT, USA) at 40% amplitude in
pulse mode of 5-sec on and 5-sec off for 30 min on ice, the cell
lysate was stirred with 100 ml n-butanol (Sigma-Aldrich) at
4°C for 30 min. Subsequently, centrifugation was carried out at
7,000 × g for 45 min. The lower aqueous phase was collected and 100
ml sodium citrate buffer was added again for re-extraction. The two
aqueous phases were pooled and dialyzed overnight in deionized
water using a dialysis membrane with MWCO 1000 (Spectrum
Laboratories Inc., Rancho Dominguez, CA, USA). The dialyzed sample
was treated with vacuum filtration (Millipore, Billerica, MA, USA)
and prepared for purification. The dialyzed sample was subjected to
hydrophobic interaction chromatography (HIC) on an octyl-sepharose
column (2.5×10 cm) (Sigma-Aldrich). The dialyzed sample was
prepared and transferred to HIC in 15% n-propanol in 0.1 M
ammonium acetate. The bond fractions were eluted with a linear
concentration gradient from 15 to 60% n-propanol in 0.1 M
ammonium acetate (pH 4.7).
The structure of the LTA was assayed by
1H nuclear magnetic resonance (NMR) spectrometry as
described previously (19). The purity
of the LTA was determined by measuring the protein content through
Coomassie Blue G-250 staining (Bio-Rad, Hercules, CA, USA)
following sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) in gradient gels containing 5–12% wt/vol acrylamide
(Bio-Rad). Precision Plus Protein Unstained standards (Bio-Rad)
were used as reference markers. The molecular weight of LTA was
shown in the SDS-PAGE gel stained with Coomassie Blue dye.
Endotoxin contamination was excluded using a Limulus amoebocyte
lysate (LAL) assay kit (Pierce, Rockford, IL, USA) according to the
manufacturer's instructions. Thus, the endotoxin was quantified and
the contamination was evaluated (19,20). DNA or
RNA contamination was determined by measuring ultraviolet (UV)
absorption at 260 and 280 nm (NanoDrop Spectrophotometer 2000;
Thermo Fisher Scientific, Wilmington, DE, USA) (21).
Extraction of the E. faecalis genome
and detection of dlt genes
The genome of E. faecalis P25RC was extracted
using the Wizard Genome DNA Purification kit (Promega Corp.,
Madison, WI, USA). The fragments of genes dltA and
dltB were amplified using polymerase chain reaction (PCR).
The sequences of primers were: dltA forward, 5′-TTGATGAAT
GGGCACGTAAA-3′ and reverse, 5′-GCCAATGCGGGA TAAATAGA-3′; and
dltB forward, 5′-AGGTGAATGCCA CATGA-3′ and reverse,
5′-GACACCACCAAGATAATGACTGA-3′ (6). The
PCR products were visualized on a 1.5% agarose gel stained with
Redsafe Nucleic Acid Staining Solution (1:10,000) (iNtRON, Seoul,
Korea).
LTA treatment
LTA-containing fractions were pooled and
lyophilized. LTA was weighed using an analytical balance
(Sartorious, Goettingen, Germany), reconstituted using PBS (pH 7.4)
to a concentration of 1 mg/ml and stored at −70°C until use. E.
faecalis LTA was added into RPMI-1640 culture medium and mixed
thoroughly in plastic tubes. PMA pre-activated THP-1 cells were
treated with E. faecalis LTA at a gradient concentration
between 0.1 and 50 µg/ml at 37°C for 18 h. Cells without LTA
treatment were used as controls. All the experiments were performed
in triplicate.
ELISA assays
Following the stimulation of the cells with E.
faecalis LTA, supernatants were collected and assayed for
inflammatory and anti-inflammatory cytokines using sandwich ELISA
with their well-matched antibodies. Cytokines, such as TNF-α and
interleukin (IL)-10, were detected using ELISA kits (Invitrogen
Life Technologies, Frederick, MD, USA) according to the
manufacturer's instructions (22).
Quantitative PCR (qPCR)
The cells were washed twice with PBS and harvested
by scraping using cell scrapers (Costar, Corning, NY, USA)
following incubation with LTA. The mRNA of each sample was
extracted and purified using the RNeasy Plus Mini kit (Qiagen,
Hilden, Germany) and quantified using a Nanodrop 2000
spectrophotometer (Thermo Fisher Scientific). The cDNA was obtained
through SuperScript® VILO™ MasterMix (Invitrogen Life Technologies)
using 1.5 µg mRNA as a template for the final reaction volume of 20
µl. The qPCR was performed using a StepOne qPCR System (Applied
Biosystems Inc., Carlsbad, CA, USA) in standard mode and the PCR
products were detected with SYBR-Green (Life Technologies, Austin,
TX, USA). The signaling pathway messengers, including TLR2,
CD14 and MyD88, were amplified using qPCR. Primer
sequences were as follows: TLR2 forward,
5′-TTATCCAGCACACGAATACACAG-3′ and reverse,
5′-AGGCATCTGGTAGAGTCATCAA-3′; CD14 forward,
5′-GACCTAAAGATAACCGGCACC-3′ and reverse,
5′-GCAATGCTCAGTACCTTGAGG-3′; MyD88 forward,
5′-GGCTGCTCTCAACATGCGA-3′ and reverse, 5′-CTG TGTCCGCACGTTCAAGA-3′;
and GAPDH forward, 5′-ATGTGTCCGTCGTGGATCTGA-3′ and reverse, 5′-ATG
CCTGCTTCACCACCTTCT-3′. The qPCR reaction conditions were performed
at 95°C for 10 min, followed by 40 cycles of 95°C for 15 sec and
60°C for 1 min. The experiment was performed in triplicate. The
relative expression of the mRNA was normalized with GAPDH using the
2−ΔΔCT method.
Western blot analysis
The phosphorylation of NF-κB and MAPKs, including
ERK1/2, JNK1/2 and p38, was detected by western blot analysis, as
previously described (21). The
nuclear and cytoplasmic proteins were extracted using NE-PER™
Nuclear and Cytoplasmic Extraction reagents (Thermo Fisher
Scientific, Rockford, IL, USA). The protein was quantified with a
BCA™ protein assay kit (Thermo Fisher Scientific). Briefly, cell
lysates containing 20 µg of protein were boiled for 5 min in Lane
Marker Sample buffers (Thermo Fisher Scientific) and the volume
ratio of sample buffer in the protein sample was 1:4. The protein
was subsequently separated by 5–12% SDS-PAGE (Bio-Rad) and blotted
onto polyvinylidene fluoride membranes (GE Healthcare, Uppsala,
Sweden). After blocking for 1 h in 20 mmol/l Tris-HCl, 150 mmol/l
NaCl and 0.05% Tween-20 (TBS-T) containing 5% skimmed milk, the
membranes were washed three times in 20 ml TBS-T and probed
overnight at 4°C with rabbit anti-human antibodies for
phospho-NF-κB p65 (monoclonal; 3033), NF-κB p65 (monclonal; 4764),
phospho-p38 (polyclonal; 9211), p38 (polyclonal; 9212), phospho
ERK1/2 (monoclonal; 4377), ERK1/2 (monoclonal; 4695),
phospho-SAPK/JNK (monoclonal; 4671), SAPK/JNK (polyclonal; 9252)
(Cell Signaling, Beverly, MA, USA) in TBS-T with 0.5% skimmed milk.
Following three washes in 20 ml TBS-T, the membranes were incubated
with secondary horseradish peroxidase (HRP)-conjugated anti-rabbit
antibody (Cell Signaling Technology, Inc., Beverly, MA, USA) for 2
h and washed three times in 20 ml TBS-T. Bands were detected using
enhanced chemiluminescence reagents (Thermo Fisher Scientific,
Wilmington, DE, USA). The proteins were normalized with β-actin
(Santa Cruz Biotechnology Inc., Dallas, TX, USA). The band
densities were measured using Quantity One 4.6.9 software
(Bio-Rad).
Statistical analysis
The qPCR, ELISA and western blot analysis data were
analyzed by analysis of variance using IBM SPSS software version 20
(IBM Corp., Armonk, NY, USA). The Bonferroni method was used to
compare the difference between two groups. P<0.05 was considered
to indicate a statistically significant difference.
Results
Extraction, purification and
identification of LTA
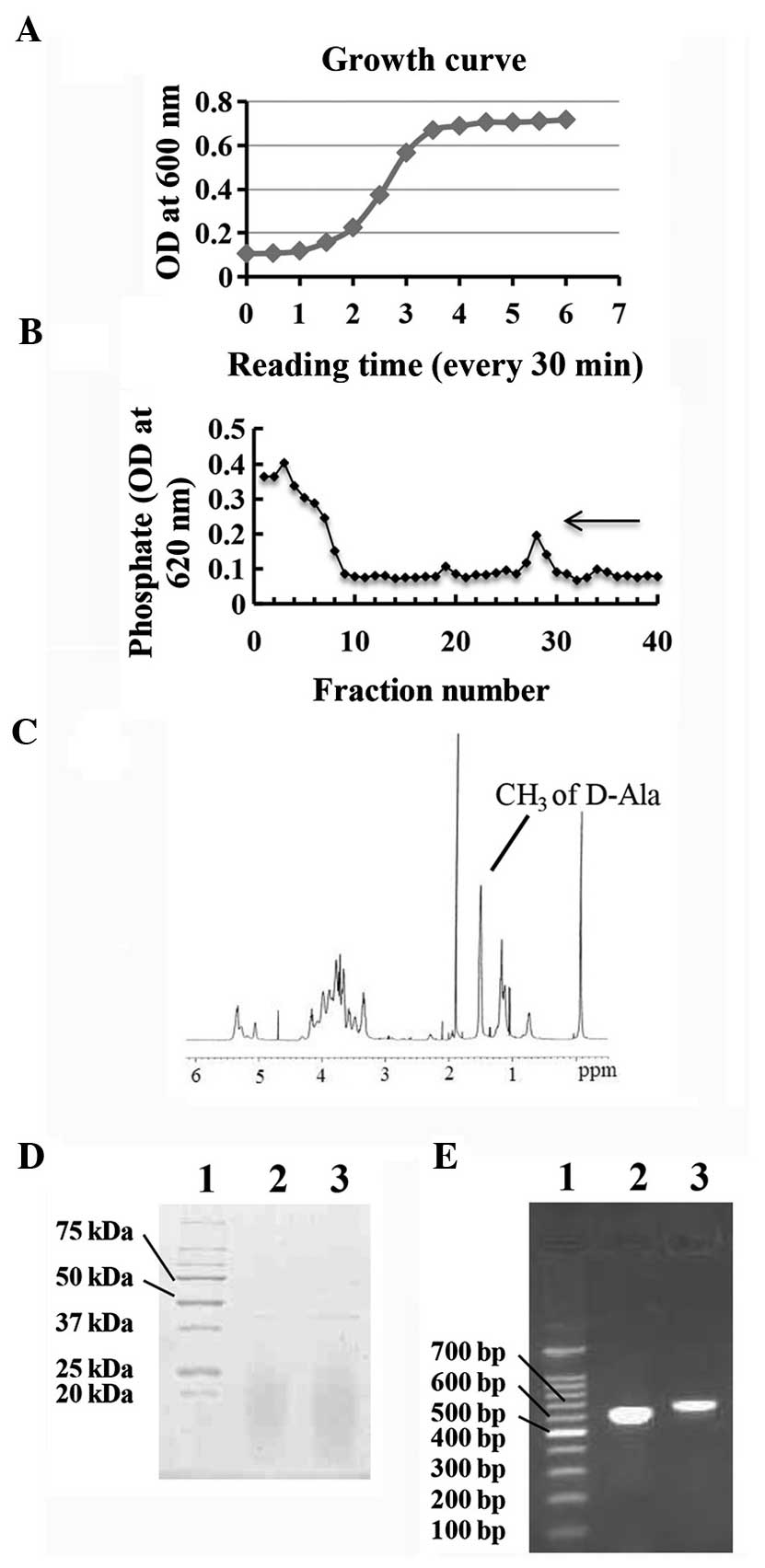
E. faecalis P25RC was grown and reached an
optical density of 0.5 at 600 nm for LTA extraction (Fig. 1A). LTA was extracted using the
n-butanol method and purified by HIC on octyl-sepharose. The
bond fractions were assayed for total phosphorus (Fig. 1B). The fractions corresponding to the
peak on the curve, which were considered the ones containing LTA,
were pooled. The structure of LTA was detected using 1H
NMR spectrometry (Fig. 1C). The purity
of LTA was determined by measuring the protein contamination
through Coomassie Blue staining following SDS-PAGE and the
molecular weight of LTA showed ~20 kDa (Fig. 1D). Endotoxin contamination was excluded
through an LAL assay and endotoxin content was <0.3 EU/mg in the
lyophilized LTA. DNA and RNA contaminations were determined by
measuring UV absorption at 260 and 280 nm. DNA and RNA accounted
for 5.4 and 3.4% (wt/wt), respectively. Conclusively, purified LTA
was prepared. The D-alanine residue was confirmed by amplifying the
fragments of the dltA and dltB operon genes using PCR
(Fig. 1E). The presence of PCR
products on the agarose gel showed the dltA (598 bp) and
dltB (649 bp) fragments. Preliminarily, LTA from E.
faecalis P25RC could be confirmed as D-alanyl LTA.
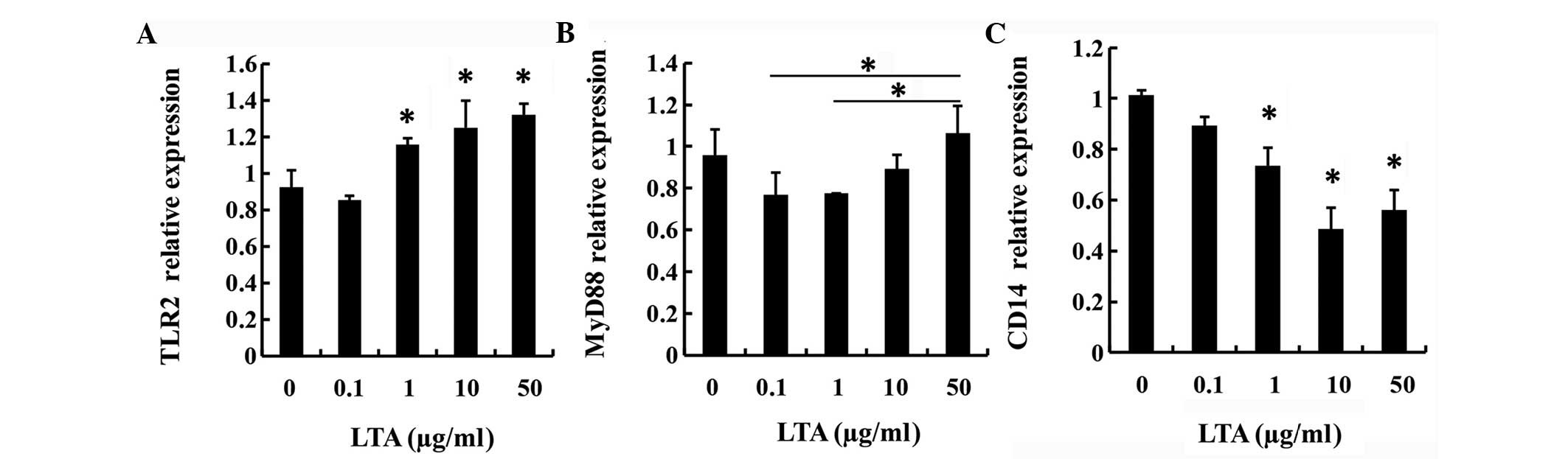
LTA promotes the gene expression of
TLR2 and MyD88
LTA from the E. faecalis P25RC clinical
strain was used to treat macrophages that were differentiated from
THP-1 cells. Cells without LTA treatment were used as controls. At
the concentration of 50 µg/ml LTA, the expression of TLR2
was enhanced ≤1.4-fold compared to the control and additionally,
the expression of MyD88 was enhanced ≤1.4-fold compared to
0.1 and 1 µg/ml LTA treatment. The expression of CD14 was
decreased by 0.5-fold compared to the control. Based on the results
of qPCR, D-alanyl LTA from E. faecalis P25RC promoted the
gene expression of TLR2 (P<0.05, Fig. 2A) and MyD88 (P<0.05, Fig. 2B), but LTA downregulated CD14
(P<0.05, Fig. 2C). In particular,
LTA from E. faecalis P25RC induced an inflammatory response
through TLR2-dependent and CD14-independent signaling
transduction.
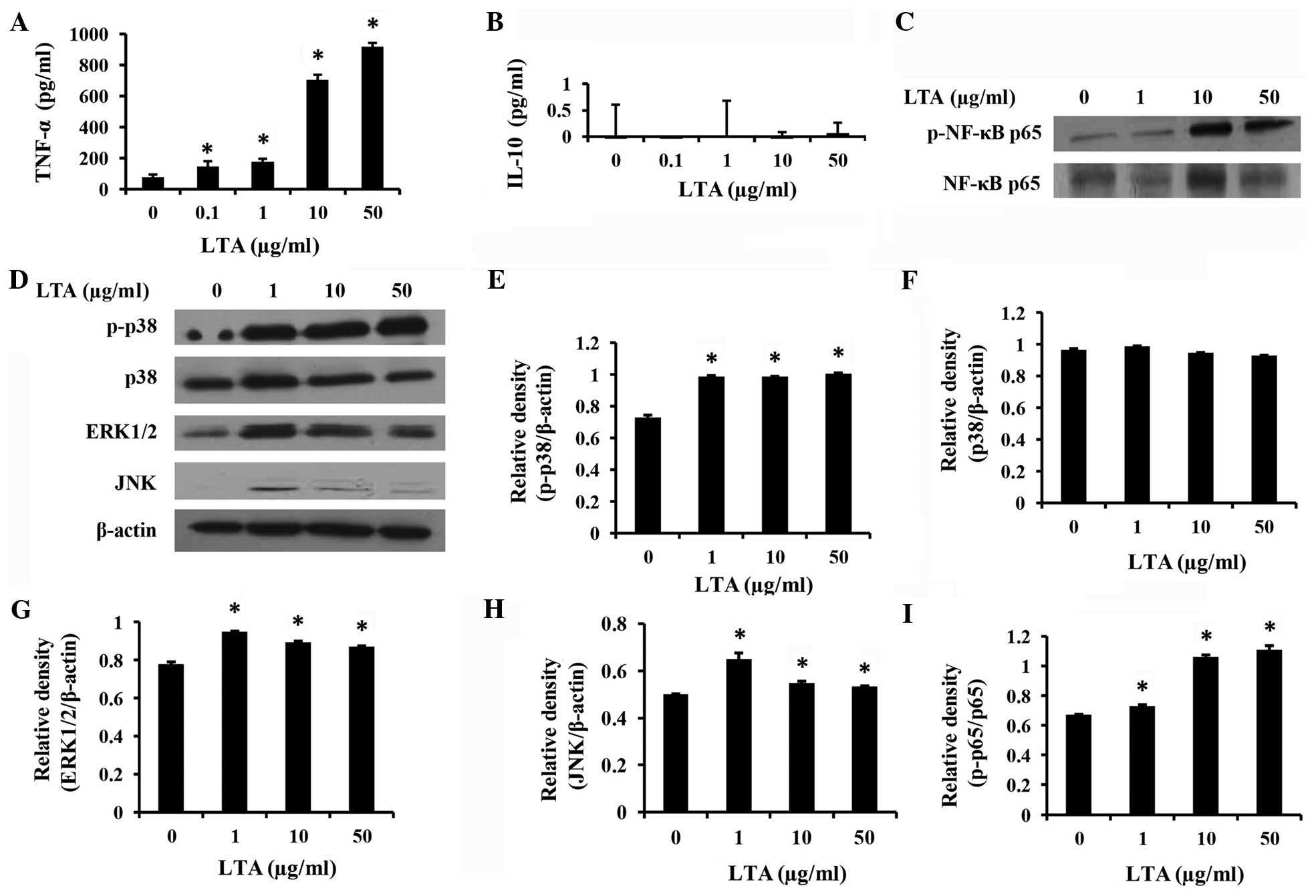
LTA promotes the expression of
TNF-α
The cytokines, such as TNF-α and IL-10, were
detected using ELISA sandwich kits. LTA stimulated the expression
of pro-inflammatory cytokine TNF-α (P<0.05, Fig. 3A), but not the anti-inflammatory
cytokine IL-10 (P>0.05, Fig.
3B).
LTA promotes p38 and NF-κB signaling
pathways
The three activated total MAPKs (p38, ERK1/2 and
JNK) (Fig. 3C) and NF-κB p65 (Fig. 3D) were assayed using western blot
analysis. In the present study, the results also suggest that
MAPKs, particularly p38, may play an important role in signal
transduction for TNF-α production. There was a dose-dependent
increase in p-p38 (P<0.05, Fig. 3E)
and p-NF-κB activity (P<0.05 compared to the control, Fig. 3I). By contrast, E. faecalis LTA
did not induce ERK1/2 or JNK activity. The total ERK1/2 (Fig. 3G) and JNK (Fig. 3H) were upregulated, but the activation
was not detected. Therefore, the p-p38 and p-NF-κB signaling
transduction pathways likely play a major role in E.
faecalis LTA-induced TNF-α expression in differentiated THP-1
macrophages.
Discussion
The present study examined the molecular mechanism
behind the interaction of a highly purified LTA derived from the
E. faecalis clinical strain with differentiated THP-1
macrophages. The n-butanol method and HIC on octyl-sepharose
were used to extract and purify LTA. The butanol method can be used
to extract structurally intact LTA and has been widely used to
extract LTA from numerous types of Gram-positive bacteria, such as
Staphylococcus aureus, Streptococcus pneumoniae,
E. faecalis and Streptococcus gordonii (6,19,23,24).
Furthermore, SDS-PAGE and Coomassie Blue G-250 staining were used
to detect protein contamination. Coomassie Blue G-250 staining
provides a detection limit of 1 ng protein and is sensitive enough
to exclude protein contamination. Of note, LTA could be exhibited
on the gel following staining by Coomassie Blue, while LTA was
previously examined using silver stain (25,26). LTA
showed a clear broad band of ~20 kDa. The LTA image was in
agreement with a previous study regarding LTA derived from other
Gram-positive bacteria (27).
Therefore, SDS-PAGE analysis with Coomassie Blue staining could
become a rapid and easy way to detect LTA. The NMR spectrum was
highly analogous to those reported in a previous study regarding
E. faecalis LTA (6). Taken
together, highly purified LTA was successfully isolated from an
E. faecalis clinical strain.
LTA is an amphiphile that is anchored to the cell
membrane via its glycolipid and extends its long chains of
glycerophosphate into the wall (20).
The dlt genes are involved in the production of D-alanine
residues and the inactivation of the dlt genes can lead to
the absence of D-alanine linked on the LTA glycerolphosphate
backbone. Furthermore, D-alanine is closely associated with the
formation of biofilm (9). Fabretti
et al (6) found that the
dltA deletion mutant of a strong biofilm producer (E.
faecalis 12030) was not as potent as the wild-type strain in
the production of biofilm. In the present study, the dltA
and dltB genes were detected in E. faecalis P25RC.
They are responsible for the production of D-alanine, suggesting
that E. faecalis P25RC LTA contains D-alanine. The E.
faecalis P25RC strain came from an infected root canal of a
patient suffering from persistent periapical periodontitis.
Therefore, understanding the immune-inflammatory activity of E.
faecalis P25RC may enhance our understanding of the apical
resorption process observed in endodontic infections.
As a surface molecule of Gram-positive bacteria, LTA
plays an important role in the stimulation of inflammatory
responses, in vivo and in vitro (21,28). In the
present study, gene expression of the signaling mediators was
detected using qPCR. LTA from the clinical strain used in the
present study induced an inflammatory response through
TLR2-MyD88-dependent, but CD14-independent signaling
transduction. There are 10 TLRs in human cells, each of which can
recognize different pathogen-associated molecular patterns
(26). LTA is a ligand against TLR2
and partially dependent on TLR1, TLR6 and CD14 (27). TLR2 may bind easier to E.
faecalis LTA than other TLRs in infected dental pulps, as an
overwhelming majority of TLR2-positive macrophages are found in
pulp inflammation (29).
The interaction of LTA with TLR2-dependent pathways
can activate intracellular messengers, such as MyD88 and MAPKs, and
subsequently transcription factors, including NF-κB, to stimulate
the expression of inflammatory cytokines (28,30). In the
present study, the release of cytokines was quantified using ELISA
and the protein expression of cells was assessed using western blot
analysis. The dose-dependent increment of TNF-α corresponded to the
activation of NF-κB at the LTA concentration of 10 µg/ml. NF-κB is
an important transcription factor that promotes the rapid release
of cytokines (31). The present
results show that NF-κB is responsible for the production of TNF-α,
in terms of the effect of the E. faecalis clinical strain
LTA on macrophages. The p38 was also activated, albeit not
simultaneously with NF-κB. Martinho et al (16) also reported that the phosphorylation of
NF-κB and p38 did not reach their peaks at the same time in the
study of signaling pathways associated with primary endodontic
infectious contents. In the signaling pathways, the upstream
adaptors could be responsible for this inconsistent phenomenon
regarding the activation of NF-κB and p38 (32).
In the present study, highly purified LTA was
successfully isolated from a clinical E. faecalis strain
derived from the infected root canal of a patient with persistent
apical periodontitis. E. faecalis LTA was also demonstrated
to promote the expression of pro-inflammatory cytokine TNF-α
through activating the p38 MAPK and NF-κB signaling pathways, but
did not influence the expression of anti-inflammatory cytokine
IL-10. Therefore, the present study provides a new insight into the
pathogenic mechanisms of E. faecalis in endodontic
infections and may possibly lead to the development of novel
therapeutic options targeting signaling pathways.
Acknowledgements
The Research Fund for the Control of Infectious
Diseases/Health and Health Services Research Fund (grant no.
12110772) supported the present study. The authors would like to
thank Mr. Raymond Tong, Ms. Edith Tong and Ms. Wendy Yu for their
technical assistance.
References
|
1
|
Jones ME, Draghi DC, Thornsberry C,
Karlowsky JA, Sahm DF and Wenzel RP: Emerging resistance among
bacterial pathogens in the intensive care unit - a European and
North American Surveillance study (2000–2002). Ann Clin Microbiol
Antimicrob. 3:142004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rôças IN, Siqueira JF Jr and Santos KR:
Association of Enterococcus faecalis with different forms of
periradicular diseases. J Endod. 30:315–320. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zoletti GO, Siqueira JF Jr and Santos KR:
Identification of Enterococcus faecalis in root-filled teeth
with or without periradicular lesions by culture-dependent
and-independent approaches. J Endod. 32:722–726. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kayaoglu G and Ørstavik D: Virulence
factors of Enterococcus faecalis: Relationship to endodontic
disease. Crit Rev Oral Biol Med. 15:308–320. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baik JE, Ryu YH, Han JY, Im J, Kum KY, Yun
CH, Lee K and Han SH: Lipoteichoic acid partially contributes to
the inflammatory responses to Enterococcus faecalis. J
Endod. 34:975–982. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fabretti F, Theilacker C, Baldassarri L,
Kaczynski Z, Kropec A, Holst O and Huebner J: Alanine esters of
enterococcal lipoteichoic acid play a role in biofilm formation and
resistance to antimicrobial peptides. Infect Immun. 74:4164–4171.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tian Y, Zhang X, Zhang K, Song Z, Wang R,
Huang S and Lin Z: Effect of Enterococcus faecalis
lipoteichoic acid on apoptosis in human osteoblast-like cells. J
Endod. 39:632–637. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Theilacker C, Kaczynski Z, Kropec A,
Fabretti F, Sange T, Holst O and Huebner J: Opsonic antibodies to
Enterococcus faecalis strain 12030 are directed against
lipoteichoic acid. Infect Immun. 74:5703–5712. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Neuhaus FC and Baddiley J: A continuum of
anionic charge: Structures and functions of D-alanyl-teichoic acids
in gram-positive bacteria. Microbiol Mol Biol Rev. 67:686–723.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rolph HJ, Lennon A, Riggio MP, Saunders
WP, MacKenzie D, Coldero L and Bagg J: Molecular identification of
microorganisms from endodontic infections. J Clin Microbiol.
39:3282–3289. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Iwasaki A and Medzhitov R: Toll-like
receptor control of the adaptive immune responses. Nat Immunol.
5:987–995. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Long EM, Millen B, Kubes P and Robbins SM:
Lipoteichoic acid induces unique inflammatory responses when
compared to other toll-like receptor 2 ligands. PLoS One.
4:e56012009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsuchiya S, Kobayashi Y, Goto Y, Okumura
H, Nakae S, Konno T and Tada K: Induction of maturation in cultured
human monocytic leukemia cells by a phorbol diester. Cancer Res.
42:1530–1536. 1982.PubMed/NCBI
|
|
14
|
Hjort MR, Brenyo AJ, Finkelstein JN,
Frampton MW, LoMonaco MB, Stewart JC, Johnston CJ and D'Angio CT:
Alveolar epithelial cell-macrophage interactions affect
oxygen-stimulated interleukin-8 release. Inflammation. 27:137–145.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ueki K, Tabeta K, Yoshie H and Yamazaki K:
Self-heat shock protein 60 induces tumour necrosis factor-alpha in
monocyte-derived macrophage: Possible role in chronic inflammatory
periodontal disease. Clin Exp Immunol. 127:72–77. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Martinho FC, Leite FR, Chiesa WM,
Nascimento GG, Feres M and Gomes BP: Signaling pathways activation
by primary endodontic infectious contents and production of
inflammatory mediators. J Endod. 40:484–489. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu X, Wang Q, Zhang C, Cheung GS and Shen
Y: Prevalence, phenotype, and genotype of Enterococcus
faecalis isolated from saliva and root canals in patients with
persistent apical periodontitis. J Endod. 36:1950–1955. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xie W, Li M, Xu N, Lv Q, Huang N, He J and
Zhang Y: MiR-181a regulates inflammation responses in monocytes and
macrophages. PLoS One. 8:e586392013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Draing C, Pfitzenmaier M, Zummo S, Mancuso
G, Geyer A, Hartung T and von Aulock S: Comparison of lipoteichoic
acid from different serotypes of Streptococcus pneumoniae. J
Biol Chem. 281:33849–33859. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Morath S, von Aulock S and Hartung T:
Structure/function relationships of lipoteichoic acids. J Endotoxin
Res. 11:348–356. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim HG, Lee SY, Kim NR, Ko MY, Lee JM, Yi
TH, Chung SK and Chung DK: Inhibitory effects of Lactobacillus
plantarum lipoteichoic acid (LTA) on Staphylococcus
aureus LTA-induced tumor necrosis factor-alpha production. J
Microbiol Biotechnol. 18:1191–1196. 2008.PubMed/NCBI
|
|
22
|
Gisch N, Kohler T, Ulmer AJ, Müthing J,
Pribyl T, Fischer K, Lindner B, Hammerschmidt S and Zähringer U:
Structural reevaluation of Streptococcus pneumoniae
Lipoteichoic acid and new insights into its immunostimulatory
potency. J Biol Chem. 288:15654–15667. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Morath S, Geyer A and Hartung T:
Structure-function relationship of cytokine induction by
lipoteichoic acid from Staphylococcus aureus. J Exp Med.
193:393–397. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
McCormick NE, Halperin SA and Lee SF:
Regulation of D-alanylation of lipoteichoic acid in
Streptococcus gordonii. Microbiology. 157:2248–2256. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Maurer JJ and Mattingly SJ: Molecular
analysis of lipoteichoic acid from Streptococcus agalactiae.
J Bacteriol. 173:487–494. 1991.PubMed/NCBI
|
|
26
|
Han SH, Kim JH, Martin M, Michalek SM and
Nahm MH: Pneumococcal lipoteichoic acid (LTA) is not as potent as
staphylococcal LTA in stimulating Toll-like receptor 2. Infect
Immun. 71:5541–5548. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cot M, Ray A, Gilleron M, Vercellone A,
LarrouyMaumus G, Armau E, Gauthier S, Tiraby G, Puzo G and Nigou J:
Lipoteichoic acid in Streptomyces hygroscopicus: Structural
model and immunomodulatory activities. PLoS One. 6:e263162011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Deininger S, Stadelmaier A, von Aulock S,
Morath S, Schmidt RR and Hartung T: Definition of structural
prerequisites for lipoteichoic acid-inducible cytokine induction by
synthetic derivatives. J Immunol. 170:4134–4138. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mutoh N, TaniIshii N, Tsukinoki K, Chieda
K and Watanabe K: Expression of toll-like receptor 2 and 4 in
dental pulp. J Endod. 33:1183–1186. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ryu YH, Baik JE, Yang JS, Kang SS, Im J,
Yun CH, Kim DW, Lee K, Chung DK, Ju HR, et al: Differential
immunostimulatory effects of Gram-positive bacteria due to their
lipoteichoic acids. Int Immunopharmacol. 9:127–133. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Albiger B, Dahlberg S, HenriquesNormark B
and Normark S: Role of the innate immune system in host defence
against bacterial infections: Focus on the Toll-like receptors. J
Intern Med. 261:511–528. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Garcia de Aquino S, Manzolli Leite FR,
Stach-Machado DR, Francisco da Silva JA, Spolidorio LC and Rossa C
Jr: Signaling pathways associated with the expression of
inflammatory mediators activated during the course of two models of
experimental periodontitis. Life Sci. 84:745–754. 2009. View Article : Google Scholar : PubMed/NCBI
|