Introduction
Phytic acid (PA) is a natural plant inositol
hexaphosphate comprising up to 10–30 g/kg dry matter in nuts,
cereals, edible legumes and oil seeds (1,2). The
nutritional importance of PA in limiting the availability of
essential dietary minerals has been emphasized (1,2). Most of
the reported actions of PA, including its effect on mineral
absorption, have been linked to the chemical properties of the
phosphate groups of its myo-inositol (MI) ring (1,2). A recent
study focused on the similarities between PA and MI in nutritional
or physiological functions (3). The
results suggested that PA and MI increase insulin sensitivity in
adipocytes by increasing lipid storage capacity, improving glucose
uptake and inhibiting lipolysis (3).
Additionally, experiments on their anticancer effects by Vucenik
and Shamsuddin (4) demonstrated that
PA and MI prevented various types of cancer. To date, it has been
demonstrated that equimolar concentrations of dietary PA and MI
prevent sucrose- or xenobiotic-induced fatty liver by reducing
hepatic lipogenic enzyme activity in rats (5–7). In
addition, a practical level of dietary PA (approximately 0.035%)
has been reported to prevent fatty liver caused by a high-sucrose
(HSC) diet in rats (8). However, in
rats fed a high-starch (HSR) diet, dietary PA and MI did not affect
hepatic triglyceride (TG) concentration or hepatic lipogenic enzyme
activity (5–7).
It has been reported that digestion of PA in
monogastric animals may be relatively inefficient and, like dietary
fibers or oligosaccharides, undigested PA or partially digested
inositol phosphates may reach the colon (9,10). Vucenik
and Shamsuddin (4) demonstrated that
PA intake is effective in preventing colon carcinogenesis in rats
and mice. Furthermore, dietary PA may modulate the composition of
cecal organic acids, mucins and microbiota in rats fed a high-fat
diet (11).
Studies have suggested that gut microbiota serve a
role in the etiology of nonalcoholic fatty liver disease (12,13). The
preventive effect of PA on fatty liver caused by an HSC diet may be
associated with the modulation of gut microbiota. In relation to
this, a recent study by our group demonstrated that the preventive
effect of dietary PA on fatty liver caused by an HSC diet was
mediated by downregulation of the gene expression of hepatic
lipogenic enzymes. In addition, PA intake was indicated to increase
the fecal ratio of Lactobacillus spp. and depress the ratio
of Clostridium coccoides (C. coccoides) in rats fed
the HSC diet (14). However, the
effect of PA intake on hepatic lipogenic gene expression and gut
microbiota in rats fed an HSR diet remains unknown.
The present study aimed to evaluate whether the
modulatory effects of PA intake on the gene expression of hepatic
lipogenic enzymes and modulation of gut microbiota in rats fed an
HSC diet depend on the dietary carbohydrate source and are
attributed to the MI ring of PA. Cecal organic acids (OAs), which
are products of bacterial fermentation, affect the levels of
hepatic lipogenesis and cholesterogenesis (15,16).
Therefore, the effect of the dietary treatment on the cecal OA
profile was also investigated.
Materials and methods
Animals and diets
A total of 42 male Sprague-Dawley rats (3-weeks-old,
54–67 g) were purchased from Japan SLC, Inc. (Hamamatsu, Japan).
The animals were maintained according to the ‘Guide for the Care
and Use of Laboratory Animals’ established by Fuji Women's
University (Ishikari, Japan), and the study was approved by the
university's ethics committee. The rats were individually housed in
a room with a controlled temperature (22–24°C), relative humidity
(55–65%) and under a 12-h light/dark cycle (light from 08:00 a.m.
to 08:00 p.m.). Following ad libitum access to a
non-purified commercial rodent powder diet (CE-2; CLEA Japan, Inc.,
Tokyo, Japan) for 3 days, the rats (78–90 g) were assigned to
weight-matched groups (n=6 rats per group) and given ad
libitum access to experimental diets and deionized water. Food
intake and body weight were measured daily at 08:00 a.m. In
experiment 1, the rats from four groups were fed an HSR or HSC diet
with or without 1.02% dodecasodium phytate (sodium PA;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 12 days. In
experiment 2, the rats from three groups were fed the HSC diet or
HSC diets with 1.02% sodium PA or 0.2% MI (Wako Pure Chemical
Industries, Ltd., Osaka, Japan) for 12 days. Table I lists the compositions of the
experimental diets (17). The molar
concentration of the added MI was equivalent to that of the added
PA. Feces were collected over the last 3 days of the experiments,
stored at −20°C, and subsequently freeze-dried and milled. At the
end of the feeding period, the rats were anesthetized with 3.0%
isoflurane (Wako Pure Chemical Industries, Ltd.) and sacrificed to
collect whole blood from the abdominal aorta. The serum was
obtained by centrifugation at 2,000 × g for 20 min at 4°C and then
stored at −80°C. The liver and cecum were removed and weighed. A
part of the liver was suspended in RNAlater stabilization reagent
(Ambion; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
stored at −80°C until RNA extraction. The remaining portions of
liver and cecum were immediately frozen using liquid nitrogen and
stored at −80°C until subsequent analyses.
 | Table I.Composition of basal diets. |
Table I.
Composition of basal diets.
| Component, %
(w/w) | High-starch | High-sucrose |
|---|
| α-corn starch | 64.95 | – |
| Sucrose | – | 64.95 |
| Casein | 20.00 | 20.00 |
| Corn oil |
5.00 |
5.00 |
| Cellulose |
5.00 |
5.00 |
| Vitamin
mixturea |
1.00 |
1.00 |
| Mineral
mixtureb |
3.50 |
3.50 |
| L-cystine |
0.30 |
0.30 |
| Choline
bitartrate |
0.25 |
0.25 |
Liver lipid and enzyme assays
Liver lipids were extracted using
chloroform-methanol (2:1) solution as previously described
(5–7).
The solvent was removed by evaporation, and the total lipid (TL)
was measured gravimetrically (5–7). The TG
and cholesterol (CH) concentrations were measured using enzymatic
kits (total cholesterol C-Test Wako, cat. no. 439–17501 and
triglyceride G-Test Wako, cat. no. 432-40201; Wako Pure Chemical
Industries, Ltd.). The frozen liver was homogenized in nine volumes
of 0.14 M KCl and centrifuged at 20,000 × g for 30 min at 4°C. The
supernatant was used as the enzyme source. The activities of liver
glucose-6-phosphate dehydrogenase (G6PD) and malic enzyme (ME) were
determined spectrophotometrically by monitoring the formation of
NADPH at 340 nm, as previously described (5–7). The
reaction mixtures were as follows: 0.017 M glycylglycine,
3.5×10−3 M MgSO4, 0.33×10−3 M
NADP, 0.67×10−3 M glucose 6-phosphate for G6PD activity;
and 0.017 M glycylglycine, 7×10−3 M MgSO4,
0.7×10−3 M MnSO4, 0.33×10−3 M
NADP, 0.33×10−3 M malate for ME activity. All reagents
were obtained from Wako Pure Chemical Industries, Ltd.
It has been recently demonstrated that dietary MI
increases the plasma level of adiponectin in C57BL/6 mice fed a
high-fat diet (18). Thus, the level
of serum adiponectin was measured by using an enzyme-linked
immunosorbent assay (cat. no. TI 25462309; Otsuka Pharmaceutical
Co., Ltd., Tokushima, Japan) in experiment 2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
mRNA expression levels were determined by RT-qPCR
(14,19). Total RNA was isolated using a
NucleoSpin RNA kit (Takara Bio, Inc., Otsu, Japan) according to the
manufacturer's instructions. Purified total RNA (400 ng) was
converted to cDNA using a PrimeScript RT Master Mix (Takara Bio,
Inc.). Real-time PCR was performed using SYBR Premix Ex Taq II
(Takara Bio, Inc.), and samples were amplified in a LightCycler 480
Instrument (Roche Diagnostics GmbH, Mannheim, Germany) under the
following cycle conditions: 95°C for 30 sec followed by 40 cycles
of 95°C for 5 sec and 60°C for 30 sec. The primer sets for
G6PD forward, GATGACATCCGCAAACAGAGTGA and reverse, GCTACA
TAGGAGTTACGGGCAAAGA; ME1 forward, CAGGCCTC CGTTAGCTTTGTTC
and reverse, GCACAAGGACTGTAA ACAGCAGTGA; and fatty acid synthetase
(FAS) forward, GCTGCTACAAACAGGACCATCAC and reverse, TCTT
GCTGGCCTCCACTGAC were purchased from Takara Bio, Inc. (14,19).
Melting curve analysis was performed following amplification to
distinguish the targeted PCR product from the non-targeted PCR
product using the LightCycler 480 Basic Software v1.5. Relative
expression levels were calculated for each sample following
normalization to those of the reference gene GAPDH (forward,
GGCACAGTCAAGGCTGAGAATG and reverse, ATGGTGGTGAAGACGCCAGTA), which
was used as the endogenous control gene (19).
Microbiota analysis using qPCR
Fecal and cecal microbiota were analyzed as
previously described (20). Briefly,
bacterial genomic DNA was isolated from the freeze-dried and milled
feces or the frozen cecal content using an UltraClean Fecal DNA
extraction kit (MO BIO Laboratories; Qiagen, Inc., Valencia, CA,
USA) according to the manufacturer's instructions. DNA was
quantified using a Qubit dsDNA BR Assay kit (Thermo Fisher
Scientific, Inc.). Bacterial groups were quantified by qPCR using
the LightCycler 480 Instrument (Roche Diagnostics GmbH) and the
group-specific primers, which have been provided previously
(20). Following initial denaturation
at 95°C for 30 sec, 40 PCR cycles were performed with denaturation
at 95°C for 5 sec, annealing at 55°C [total bacteria,
Lactobacillus spp., Bifidobacterium spp., C.
coccoides, C. Clostridium leptum (C. leptum) and
Bacteroides] for 30 sec, and extension at 72°C for 15 sec
(total bacteria, Lactobacillus spp., Bifidobacterium
spp. and Bacteroides) or 1 min (C. coccoides and
C. leptum). Melting curve analysis was performed following
amplification to distinguish the targeted PCR product from the
non-targeted PCR product. Data were analyzed using the second
derivative maximum method of the LightCycler 480 Basic Software
v1.5. The amplification efficiencies (e) of real-time PCR for each
primer set were estimated using a linear regression of the crossing
point for each fecal DNA dilution versus the log dilution using the
formula: e = × - 1/slope, where ‘x’ was fold dilution. Efficiencies
were between 1.94 and 1.99 (optimum value of 2.0). The relative
abundance of microbial populations was expressed as the proportion
of total bacterial 16S rRNA genes.
Cecal pH and OAs
The pH of cecal digesta was measured directly using
a compact pH meter (B-212; Horiba, Ltd., Kyoto, Japan). The empty
cecum was weighed and the weight of cecal digesta was calculated as
the difference between the weight of the cecum and the weight of
the empty cecum. Cecal OAs were measured by the internal standard
method using high-performance liquid chromatography (HPLC; L-2130;
Hitachi, Ltd., Tokyo, Japan) equipped with an Aminex HPX-87H ion
exclusion column (7.8 mm i.d. × 30 cm; Bio-Rad Laboratories, Inc.,
Hercules CA, USA) as previously described (20,21).
Briefly, 500 mg cecal digesta was homogenized in 5 ml of 50 mmol/l
H2SO4 containing 10 mmol/l 2,2-dimethyl
butyric acid as an internal standard and subsequently centrifuged
at 17,000 × g at 2°C for 20 min. The supernatant was ultrafiltered,
and the filtrate was examined by HPLC (column at 60°C). The mobile
phase (5 mM H2SO4) was delivered at a flow
rate of 0.7 ml/min. Cecal OAs were detected at 210 nm using a
variable wavelength detector (L-2400; Hitachi, Ltd.).
Statistical analysis
All values were expressed as means ± standard error
of the mean. Normally distributed data were analyzed using two-way
analysis of variance (ANOVA), whereas non-normally distributed data
were analyzed using the non-parametric Kruskal-Wallis one-way
ANOVA. The Tukey-Kramer post hoc test was performed when a
significant effect was detected by two-way ANOVA. The Steel-Dwass
post hoc test was conducted when a significant effect was detected
by the Kruskal-Wallis one-way ANOVA. Associations between liver TGs
and other parameters (liver lipogenic enzyme activity and gene
expression, serum adiponectin, fecal and cecal microbiota and cecal
OAs) were analyzed using the Spearman rank correlation analysis.
Data analysis was performed using Excel Statistics 2012 for Windows
(SSRI Co., Ltd., Tokyo, Japan). P<0.05 was considered to
indicate statistical significance.
Results
Dietary PA alleviates HSC diet-induced
fatty liver pathology
In experiment 1, body weight gain was not affected
by supplementation of the dietary carbohydrate source with PA
(Table II). Compared with the HSR
diet, the HSC diet marginally increased food intake (P<0.05),
whereas PA intake had no effect on food intake (Table II). The HSC diet induced significant
increases in the liver weight, hepatic TL and TG concentrations,
hepatic activities of G6PD and ME, and hepatic expression of
G6PD, ME and FAS (P<0.05; Table II). The supplementation of PA in the
diet effectively prevented the increases in hepatic TL and TG
concentrations and significantly decreased the hepatic expression
of G6PD and ME1 in rats fed the HSC diet (P<0.05).
Similarly, PA intake decreased the hepatic activities of G6PD and
ME, and the hepatic expression of FAS in rats fed the HSC
diet, though these changes were determined as non-significant. In
rats fed the HSR diet, PA intake did not alter these lipid
metabolic parameters except for marginally decreasing the hepatic
TG concentration (P<0.05).
| Table II.Body weight, food intake and lipid
metabolic parameters in rats fed an HSR or HSC diet with or without
PA (experiment 1). |
Table II.
Body weight, food intake and lipid
metabolic parameters in rats fed an HSR or HSC diet with or without
PA (experiment 1).
|
| HSR | HSR + PA | HSC | HSC + PA |
|---|
|
|
|
|
|
|
|---|
| Variables | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM |
|---|
| Final body weight,
g | 171 | 4 | 176 | 4 | 178 | 1 | 183 | 2 |
| Food intake, g/12
day | 174a | 4 | 177a,b | 4 | 188b,c | 2 | 193b | 3 |
| Liver |
|
|
|
|
|
|
|
|
| Weight,
g/100 g body wt. | 4.64a | 0.10 | 4.71a | 0.12 | 5.84b | 0.13 | 5.70b | 0.06 |
| Total
lipids, mg/g liver | 64.2a | 3.6 | 57.8a | 1.2 | 106.2b | 6.2 | 62.5a | 1.4 |
|
Triglyceride, mg/g liver | 17.5b | 1.9 | 12.0a | 0.3 | 51.6c | 5.6 | 23.2b | 1.6 |
|
Cholesterol, mg/g liver | 3.22 | 0.29 | 3.13 | 0.15 | 3.56 | 0.15 | 3.04 | 0.13 |
| G6PD
activity, mU/mg protein | 34.1a | 5.4 | 36.7a | 1.7 | 69.1b | 7.8 | 53.1a,b | 6.6 |
| ME
activity, mU/mg protein | 28.9a | 2.9 | 32.1a | 2.8 | 50.6b | 5.6 | 35.7a,b | 3.5 |
|
G6PD
expression | 1.00a | 0.12 | 0.89a | 0.10 | 1.71b | 0.09 | 1.12a | 0.09 |
| ME1
expression | 1.00a,b | 0.06 | 0.72a | 0.05 | 2.08c | 0.07 | 1.09b | 0.06 |
| FAS
expression | 1.00a | 0.11 | 0.88a | 0.16 | 2.79b | 0.78 | 2.21a,b | 0.06 |
PA intake, carbohydrate source, gut
microbiota and cecal OAs
Dietary PA significantly increased fecal weight in
rats fed the HSC diet (P<0.05; Table
III). The supplementation of PA in the HSC diet significantly
increased the fecal ratio of Lactobacillus spp. and reduced
the ratio of C. coccoides (P<0.05; Table III). In rats fed the HSR diet,
dietary PA did not have a significant effect on the fecal ratios of
Lactobacillus spp. and C. coccoides. The fecal ratio
of Bacteroides was higher in rats fed the HSC diet with PA
compared with that in rats fed the HSR diet with PA (P<0.05). PA
intake did not have a significant effect on the fecal ratios of
Bifidobacterium spp. regardless of the dietary carbohydrate
source (Table III).
| Table III.Fecal and cecal microbiota and cecal
organic acids in rats fed an HSR or HSC diet with or without PA
(experiment 1). |
Table III.
Fecal and cecal microbiota and cecal
organic acids in rats fed an HSR or HSC diet with or without PA
(experiment 1).
|
| HSR | HSR + PA | HSC | HSC + PA |
|---|
|
|
|
|
|
|
|---|
| Variables | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM |
|---|
| Feces |
|
|
|
|
|
|
|
|
| Weight,
g/3 days | 3.89a,b | 0.14 | 3.96a,b | 0.13 | 3.54a | 0.07 | 4.11b | 0.14 |
|
Microbiota,% of total
bacteria |
|
|
|
|
|
|
|
|
|
Lactobacillus
spp. | 4.76a,b | 1.81 | 7.20b | 2.11 | 0.15a | 0.03 | 9.29b | 3.01 |
|
Bifidobacterium
spp. | 0.077 | 0.044 | 0.153 | 0.090 | 0.013 | 0.009 | 0.006 | 0.001 |
|
Clostridium
coccoides | 7.32a | 1.39 | 3.56b | 0.61 | 7.17b | 1.26 | 0.02a | 0.01 |
|
Bacteroides | 15.9a,b | 2.1 | 14.7a | 2.9 | 17.1a,b | 2.0 | 27.4b | 4.3 |
| Cecum |
|
|
|
|
|
|
|
|
| Tissue
weight, g/100 g body wt. | 0.380b | 0.029 | 0.335a,b | 0.011 | 0.264a | 0.017 | 0.322a,b | 0.022 |
| Cecal digesta |
|
|
|
|
|
|
|
|
|
Weight, g | 2.47a | 0.12 | 3.22b | 0.19 | 1.93a | 0.12 | 2.25a | 0.11 |
|
pH | 6.90b | 0.18 | 6.22a | 0.05 | 7.62c | 0.15 | 6.92b | 0.09 |
|
Microbiota, % of total
bacteria |
|
|
|
|
|
|
|
|
|
Lactobacillus
spp. | 1.66a,b | 0.79 | 1.88b | 0.71 | 0.04a | 0.01 | 0.50b | 0.06 |
|
Bifidobacterium
spp. | 0.0077a,b | 0.0043 | 0.0277a,b | 0.0083 | 0.0024a | 0.0003 | 0.0060b | 0.0015 |
|
Clostridium
coccoides | 8.34b | 1.69 | 2.36a,b | 0.78 | 5.39b | 0.79 | 0.48a | 0.36 |
|
Bacteroides | 30.2 | 1.8 | 45.7 | 8.8 | 30.3 | 2.0 | 53.4 | 13.9 |
| Organic acids
(µmol/total digesta) |
|
|
|
|
|
|
|
|
|
Succinate | 2.1a | 0.8 | 129.3b | 12.3 | 0.9a | 1.3 | 59.2b | 18.3 |
|
Lactate | 2.5 | 1.2 | 7.7 | 2.7 | 1.3 | 0.4 | 2.0 | 0.6 |
|
Acetate | 93.4a,b | 12.4 | 129.9b | 17.7 | 69.2a | 3.5 | 67.4a,b | 6.6 |
|
Propionate | 34.6b | 3.5 | 39.4a,b | 8.5 | 23.6a,b | 1.5 | 18.0a | 1.7 |
|
n-Butyrate | 34.5 | 7.2 | 34.7 | 3.5 | 25.9 | 1.2 | 45.2 | 25.8 |
| Total
organic acids | 165.3a,b | 24.2 | 341.2c | 23.3 | 121.2a | 4.9 | 190.7b | 25.3 |
The weight of cecal tissue was lower in rats fed the
HSC diet compared with that in rats fed the HSR diet, and the
weight of cecal digesta was significantly enhanced by PA intake in
rats fed the HSR diet (P<0.05; Table
III). The cecal pH was higher in rats fed the HSC diet compared
with that in rats fed the HSR diet. PA intake significantly lowered
the cecal pH regardless of the dietary carbohydrate source (P<
0.05; Table III). In rats fed the
HSC diet, dietary PA significantly increased not only the fecal
ratio but also the cecal ratio of Lactobacillus spp. and
significantly decreased the cecal ratio of C. coccoides
(P<0.05; Table III). In
addition, in rats fed the HSC diet, dietary PA significantly
increased the cecal ratio of Bifidobacterium spp.
(P<0.05). In rats fed the HSR diet, dietary PA had no
significant effect on the cecal microbiota composition (Table III).
In rats fed the HSR diet or the HSC diet, the cecal
succinate level was markedly enhanced by PA intake (P<0.05;
Table III). The cecal acetate level
was significantly higher in rats fed the HSR diet with PA compared
with that in rats fed the HSC diet without PA. Furthermore, the
cecal propionate level was significantly higher in rats fed the HSR
diet without PA compared with that in rats fed the HSC diet with PA
(P<0.05; Table III).
Similar effects of PA and MI intake on
HSC-induced fatty liver pathology
In rats fed the HSC diet (experiment 2), dietary PA
and MI did not affect body weight gain or food intake (Table IV). Liver weight was significantly
reduced by MI intake (P<0.05), and moderately by PA intake
(Table IV), and PA or MI intake
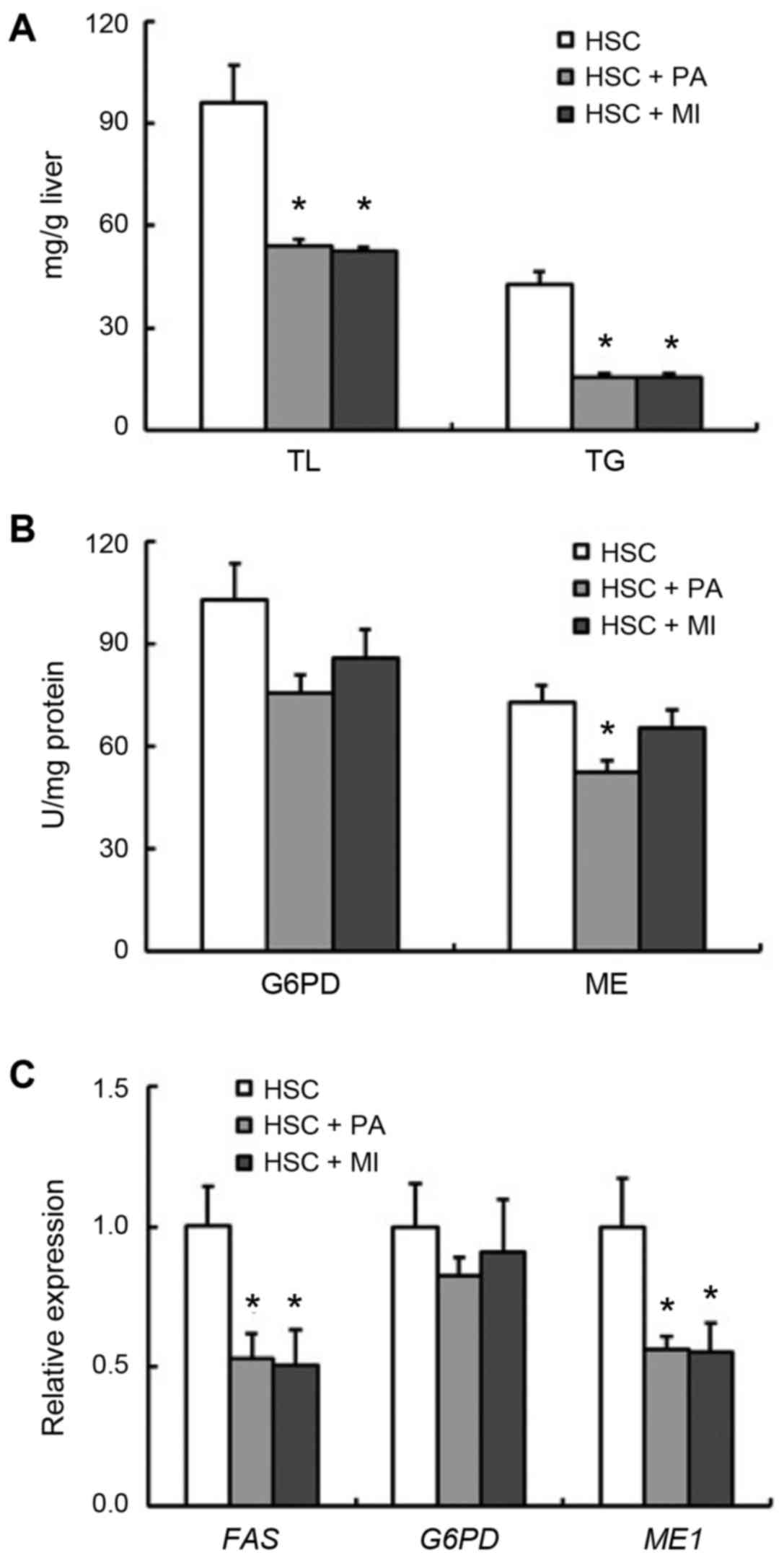
decreased the liver TL and TG concentrations (P<0.05; Fig. 1A). The activity of hepatic ME was
significantly decreased by dietary PA intake (P<0.05), and
moderately by MI intake (Fig. 1B).
Although the activity and expression of hepatic G6PD were not
significantly affected by dietary PA or MI, the expression of
hepatic ME1 and FAS in rats fed the HSC diet with PA
or MI was decreased (P<0.05; Fig. 1B
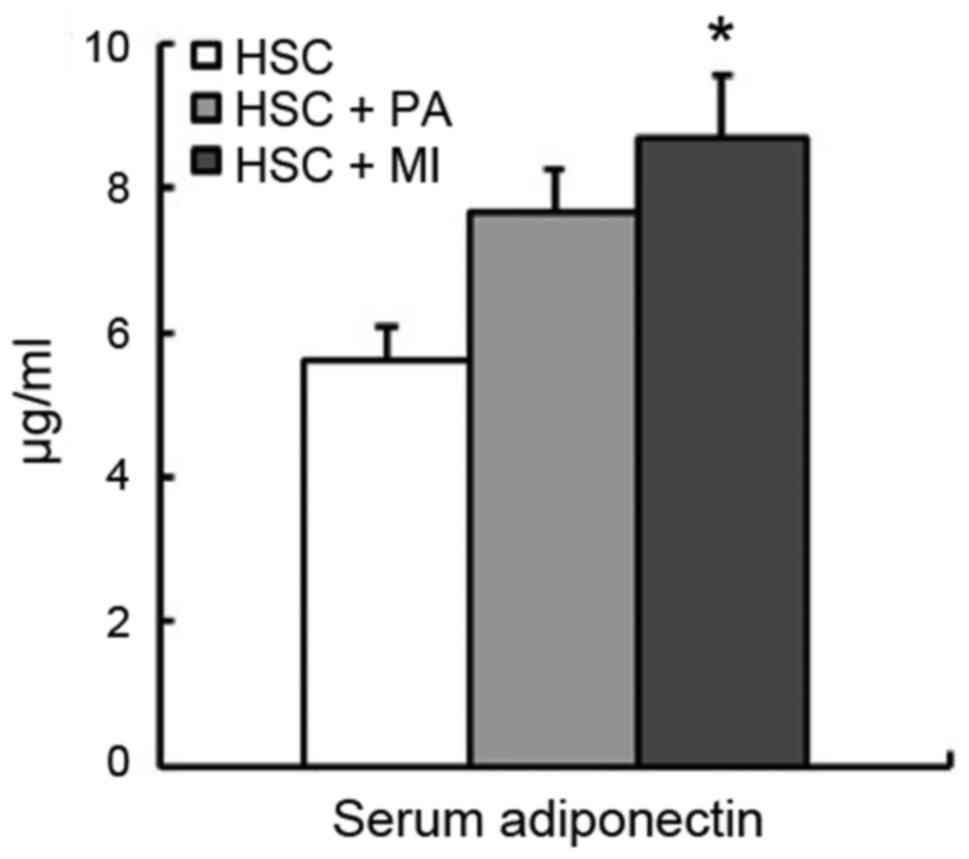
and C). The serum adiponectin level was significantly increased
by dietary MI (P<0.05), whereas it was moderately increased by
PA in rats fed the HSC diet (Fig.
2).
| Table IV.Fecal and cecal microbiota and cecal
organic acids in rats fed an HSC diet with or without MI or PA
(experiment 2). |
Table IV.
Fecal and cecal microbiota and cecal
organic acids in rats fed an HSC diet with or without MI or PA
(experiment 2).
|
| HSC | HSC + PA | HSC + MI |
|---|
|
|
|
|
|
|---|
| Variables | Mean | SEM | Mean | SEM | Mean | SEM |
|---|
| Final body weight,
g | 175 | 4 | 179 | 1 | 178 | 4 |
| Food intake, g/12
days | 183 | 4 | 183 | 2 | 185 | 5 |
| Liver |
|
|
|
|
|
|
| Weight,
g/100g body wt. | 5.83a | 0.08 | 5.52a,b | 0.08 | 5.31b | 0.12 |
| Feces |
|
|
|
|
|
|
| Weight,
g/3 days | 3.51 | 0.08 | 3.89 | 0.18 | 3.60 | 0.19 |
| Cecum |
|
|
|
|
|
|
| Tissue
weight, g/100 g body wt. | 0.245 | 0.005 | 0.291 | 0.011 | 0.270 | 0.017 |
| Cecal
digesta weight, g | 1.73a | 0.05 | 2.08b | 0.09 | 1.85a,b | 0.02 |
|
pH | 7.65c | 0.09 | 6.87a | 0.05 | 7.10b | 0.04 |
| Microbiota, % of
total bacteria |
|
|
|
|
|
|
|
Lactobacillus
spp. | 0.006a | 0.002 | 0.171b | 0.143 | 0.434b | 0.325 |
|
Bifidobacterium
spp. | 0.0031a | 0.0005 | 0.0197b | 0.0098 | 0.0178a,b | 0.0098 |
|
Clostridium
coccoides | 4.54b | 0.50 | 1.02a | 0.51 | 4.30b | 0.36 |
|
Bacteroides | 31.9 | 1.5 | 52.6 | 11.3 | 26.4 | 1.9 |
| Organic acids,
µmol/total digesta |
|
|
|
|
|
|
|
Succinate | 14.4a | 3.6 | 36.7b | 4.3 | 9.6a | 2.5 |
|
Lactate | 3.9 | 2.0 | 3.8 | 1.8 | 4.6 | 0.7 |
|
Acetate | 51.2 | 2.6 | 55.0 | 3.6 | 52.7 | 3.6 |
|
Propionate | 17.9 | 1.8 | 18.1 | 1.4 | 18.9 | 0.6 |
|
n-Butyrate | 18.8b | 1.2 | 21.2a,b | 1.0 | 23.4a | 1.8 |
| Total
organic acids | 106.2a | 7.2 | 134.8b | 4.8 | 109.2a,b | 6.8 |
Comparative effect of PA and MI on gut
microbiota and cecal OA
In experiment 2, the fecal and cecal weights were
not significantly affected by dietary PA or MI (Table IV). In accordance with experiment 1,
PA intake significantly increased the weight and lowered the pH of
the cecal digesta (P<0.05). Similarly, MI intake moderately
increased the digesta weight and significantly decreased the
digesta pH (Table IV). The
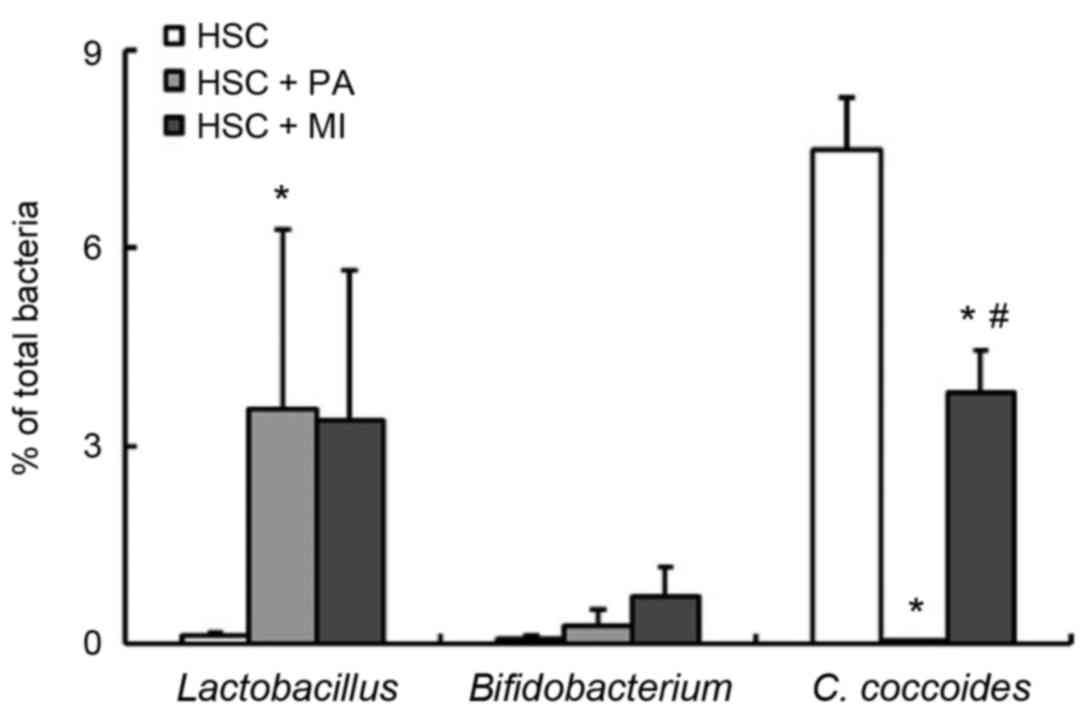
supplementation of PA in the HSC diet significantly increased the
fecal ratio of Lactobacillus spp. and decreased the fecal
ratio of C. coccoides compared with the HSC diet alone
(P<0.05; Fig. 3). Similar to PA
intake, MI intake decreased the ratio of C. coccoides
(P<0.05) and moderately increased the ratio of
Lactobacillus spp. (Fig. 3).
No significant differences were detected in the fecal ratio of
Bifidobacterium spp. among the three groups (Fig. 3). As presented in Table IV, dietary PA or MI significantly
increased the cecal ratio of Lactobacillus spp. The cecal
ratio of C. coccoides was decreased by dietary PA
(P<0.05), but was not affected by dietary MI (Table IV). Dietary PA significantly
increased the cecal ratio of Bifidobacterium spp.
(P<0.05; Table IV); dietary MI
also increased the ratio, but its effect was not significant
(Table IV). Dietary PA or MI did not
affect the cecal ratio of Bacteroides (Table IV). The cecal succinate level was
increased by the supplementation of PA in the HSC diet, as in
experiment 1; whereas it was unaffected by MI intake. MI intake
also marginally increased the cecal butyrate level (P<0.05). The
levels of all other cecal OAs were unaffected by PA or MI
intake.
Relation between liver TG and other
parameters
The data from all rats were used for correlation
analysis. As depicted in Table V, the
liver TG concentration was positively correlated with the activity
and expression of hepatic lipogenic enzymes in experiments 1 and 2,
and negatively correlated with the serum adiponectin level in
experiment 2 (P<0.05). Fecal and cecal Lactobacillus spp.
correlated negatively with the liver TG concentration in
experiments 1 and 2 (P<0.05). A similar association was observed
between the abundance of Bifidobacterium spp. and liver TG
concentration, although this correlation was not significant for
fecal abundance in experiment 2. The liver TG concentration
exhibited a significant positive correlation with the fecal and
cecal abundance of C. coccoides in experiment 2 (P<0.05);
while this correlation was not significant in experiment 1. The
cecal levels of succinate, lactate, acetate and propionate
exhibited a significant negative correlation with the liver TG
concentration in experiment 1 (P<0.05): while this correlation
was not significant in experiment 2 (Table V).
| Table V.Associations between liver TG
concentration and lipid metabolic parameters, gut microbiota and
cecal OAs. |
Table V.
Associations between liver TG
concentration and lipid metabolic parameters, gut microbiota and
cecal OAs.
|
| Correlation
coefficient, r Liver TG (mg/g tissue) |
|---|
|
|
|
|---|
| Parameters | Experiment 1 | Experiment 2 |
|---|
| Liver |
|
|
| G6PD
activity, mU/mg protein |
0.763b |
0.769b |
| ME
activity, mU/mg protein |
0.805b |
0.747b |
| G6PD
expression |
0.741b |
0.516a |
| ME
expression |
0.876b |
0.706b |
| FAS
expression |
0.718b |
0.693b |
| Serum |
|
|
|
Adiponectin, µmol/dl | ND |
−0.559a |
| Feces microbiota,
% |
|
|
|
Lactobacillus spp. |
−0.569b |
−0.562a |
|
Bifidobacterium
spp. |
−0.672b | −0.317 |
|
Clostridium
coccoides |
0.230 |
0.793b |
|
Bacteroides |
0.302 |
0.249 |
| Cecum microbiota,
% |
|
|
|
Lactobacillus spp. |
−0.702b |
−0.739b |
|
Bifidobacterium
spp. |
−0.597b |
−0.821b |
|
Clostridium
coccoides |
0.212 |
−0.508a |
|
Bacteroides | −0.275 |
0.285 |
| Cecum OAs,
µmol/total digesta |
|
|
|
Succinate |
−0.541b | −0.099 |
|
Lactate |
−0.577b | −0.100 |
|
Acetate |
−0.490a |
0.044 |
|
Propionate |
−0.506a | −0.100 |
|
n-Butyrate | −0.376 | −0.103 |
Discussion
The present study demonstrated that the effect of
dietary MI was similar to that of dietary PA on the expression of
hepatic lipogenic enzymes and gut microbiota in rats fed an HSC
diet. Sakamoto et al (22)
reported that [3H]PA is absorbed and distributed to
various organs MI and MI-monophosphate in rats. They speculated
that soluble PA when administered in drinking water is rapidly
absorbed through the stomach and upper small intestine, becomes
quickly dephosphorylated within the mucosal cells and is
distributed to various organs as inositol and MI-monophosphate
(22). In a previous study, dietary
PA significantly increased the hepatic level of free MI in rats
(7). It is postulated that the MI
ring of PA may be responsible for the effects of PA on hepatic
lipid metabolism and gut microbiota in rats fed an HSC diet. The
present results suggest that MI, a vitamin-like substance contained
in various foods including milk and citrus fruits (23,24), may
be a modulating factor of gut microbiota in rats fed an HSC
diet.
In accordance with previous results (5,7,14), the current study demonstrated that
dietary PA may prevent fatty liver pathology, which may at least in
part occur via a reduction in the activity and expression of
hepatic lipogenic enzymes. Furthermore, in rats fed the HSC diet,
dietary PA increased the ratio of Lactobacillus spp. and
decreased the ratio of C. coccoides in the cecum and feces.
The current data also confirm our previous findings (5,6) that PA
intake does not affect the hepatic TG concentration and the
activity of hepatic lipogenic enzymes in rats fed the HSR diet. The
present study indicated that dietary PA did not significantly
affect the expression of hepatic lipogenic enzymes and gut
microbiota in rats fed the HSR diet. Thus, taken together with
previous reports (5,7,14), our
findings may suggest that dietary PA normalizes hepatic TG
concentration as well as the activity and expression of hepatic
lipogenic enzymes with a concomitant modulation of gut microbiota
in rats fed a lipogenic diet.
The gut microbiota including Lactobacillus
spp. and Clostridium spp. are established to regulate
hepatic lipogenesis and fat storage (12,13). In
the present study, the hepatic TG concentration was positively
correlated with the activity and expression of hepatic lipogenic
enzymes and negatively correlated with the fecal and cecal ratios
of Lactobacillus spp. Xu et al (25) reported that the hepatic TL
concentration was negatively correlated with the concentration of
gut Lactobacillus spp. in a nonalcoholic fatty liver rat
model. Ritze et al (26)
demonstrated that Lactobacillus rhamnosus protected against
fatty liver disease induced by a high-fructose diet in mice.
Collectively, these findings indicate that hepatic TL and TG
concentrations are associated with gut microbiota, particularly
with the abundance of Lactobacillus spp. Thus, dietary PA or
MI may ameliorate fatty liver pathology by modifying the abundance
of Lactobacillus spp. in the gut of rats fed an HSC
diet.
It is well known that the plasma level of
adiponectin is decreased in patients with nonalcoholic fatty liver
disease (27,28). The delivery of recombinant adiponectin
has been reported to alleviate alcoholic and nonalcoholic fatty
liver diseases in mice (29). The
present study observed that dietary MI increased the serum level of
adiponectin. The same trend was observed in rats fed the HSC diet
with PA. The serum adiponectin level was also negatively correlated
with the liver TG concentration. Croze et al (18) recently demonstrated that dietary MI
increased the plasma level of adiponectin in C57BL/6 mice fed a
high-fat diet. The effect of dietary MI and PA on the serum level
of adiponectin may be associated with their anti-fatty liver effect
in rats fed an HSC diet. At present, a cDNA microarray analysis is
in progress by our group to determine the gene expression profile
in the liver of rats fed an HSC diet in response to dietary PA and
MI. Liver transcriptome analysis has already revealed that PA or MI
intake suppressed the expression of genes associated with lipid,
fatty acid and long-chain fatty-acyl-CoA biosynthesis processes and
the NADP-binding domain, which were upregulated by the HSC diet
(unpublished data).
Cecal OAs, which are products of bacterial
fermentation, affect hepatic lipogenesis and cholesterogenesis in
rats (15,16). In the present study, dietary PA
increased the cecal succinate level in rats regardless of the
dietary carbohydrate source. However, dietary MI did not affect the
cecal succinate level in rats fed the HSC diet, and the liver TG
concentration did not correlate with the cecal succinate level in
experiment 2. Thus, it may be speculated that the levels of cecal
OAs are not linked to the anti-fatty liver effect of PA and MI in
rats fed an HSC diet. It has previously been demonstrated that
dietary PA increased the cecal butyrate level in rats fed a
high-fat diet (11). Dietary PA may
affect the levels of cecal OAs differently according to the dietary
composition. Further studies are necessary to investigate the
association between the effect of dietary PA on the levels of cecal
OAs with varying dietary composition in rats.
In conclusion, the present study indicated that
dietary PA prevents fatty liver pathology by reducing the activity
and expression of hepatic lipogenic enzymes. In addition, it may
modulate the gut microbiota in rats fed an HSC diet, while having
no effect on hepatic lipid metabolism or gut microbiota in rats fed
an HSR diet. Therefore, it may be suggested that in rats fed a
high-lipogenic diet, dietary PA normalizes hepatic lipid metabolism
and modulates the gut microbiota; an effect not observed in rats
fed a normal diet. The findings of the present study also
implicated that the MI ring of PA may be responsible for the
beneficial effects of dietary PA on hepatic lipid metabolism and
gut microbiota in rats fed an HSC diet. Further studies on the
effect of PA or MI intake on lipid metabolism in adipose tissue and
mineral metabolism in rats with fatty liver pathology should be
performed in order to comprehensively evaluate the properties,
bioavailability and safety of PA/MI intake.
Acknowledgements
Not applicable.
Funding
The current study was supported by Grants-in-Aid for
Scientific Research from the Tojuro Iijima Foundation for Food
Science and Technology in Japan (grant no. 15, G-9, 2015).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YO and TK conceived and designed the experiments. YO
and AS performed the experiments and analyzed the data. YO and TK
wrote the paper. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Fuji Women's University (Ishikari, Japan).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kumar V, Sinha AK, Makkar HPS and Becker
K: Dietary roles of phytate and phytase in human nutrition: A
review. Food Chem. 120:945–959. 2010. View Article : Google Scholar
|
|
2
|
Silva EO and Bracarense AP: Phytic acid:
From antinutritional to multiple protection factor of organic
systems. J Food Sci. 81:R1357–R1362. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim JN, Han SN and Kim H-K: Phytic acid
and myo-inositol support adipocyte differentiation and improve
insulin sensitivity in 3T3-L1 cells. Nutr Res. 34:723–731. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vucenik I and Shamsuddin AM: Cancer
inhibition by inositol hexaphosphate (IP6) and inositol: From
laboratory to clinic. J Nutr. 133 Suppl 1:3778S–3784S. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Katayama T: Effects of dietary
myo-inositol or phytic acid on hepatic concentrations of lipids and
hepatic activities of lipogenic enzymes in rats fed on corn starch
or sucrose. Nutr Res. 17:721–728. 1997. View Article : Google Scholar
|
|
6
|
Okazaki Y and Katayama T: Effects of
dietary carbohydrate and myo-inositol on metabolic changes in rats
fed 1,1,1-trichloro-2,2-bis (p-chlorophenyl) ethane (DDT). J Nutr
Biochem. 14:81–89. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Okazaki Y and Katayama T: Dietary inositol
hexakisphosphate, but not myo-inositol, clearly improves
hypercholesterolemia in rats fed casein-type amino acid mixtures
and 1,1,1-trichloro-2,2-bis (p-chlorophenyl) ethane. Nutr Res.
28:714–721. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Onomi S, Okazaki Y and Katayama T: Effect
of dietary level of phytic acid on hepatic and serum lipid status
in rats fed a high-sucrose diet. Biosci Biotechnol Biochem.
68:1379–1381. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sandberg AS and Andersson H: Effect of
dietary phytase on the digestion of phytate in the stomach and
small intestine of humans. J Nutr. 118:469–473. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miyazawa E, Iwabuchi A and Yoshida T:
Phytate breakdown and apparent absorption of phosphorus, calcium
and magnesium in germfree and conventionalized rats. Nutr Res.
16:603–613. 1996. View Article : Google Scholar
|
|
11
|
Okazaki Y and Katayama T: Dietary phytic
acid modulates characteristics of the colonic luminal environment
and reduces serum levels of proinflammatory cytokines in rats fed a
high-fat diet. Nutr Res. 34:1085–1091. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Solga SF and Diehl AM: Non-alcoholic fatty
liver disease: Lumen-liver interactions and possible role for
probiotics. J Hepatol. 38:681–687. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Festi D, Schiumerini R, Eusebi LH, Marasco
G, Taddia M and Colecchia A: Gut microbiota and metabolic syndrome.
World J Gastroenterol. 20:16079–16094. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sekita A, Okazaki Y and Katayama T:
Dietary phytic acid prevents fatty liver by reducing expression of
hepatic lipogenic enzymes and modulates gut microflora in rats fed
a high-sucrose diet. Nutrition. 32:720–722. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hara H, Haga S, Aoyama Y and Kiriyama S:
Short-chain fatty acids suppress cholesterol synthesis in rat liver
and intestine. J Nutr. 129:942–948. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim M and Shin HK: The water-soluble
extract of chicory influences serum and liver lipid concentrations,
cecal short-chain fatty acid concentrations and fecal lipid
excretion in rats. J Nutr. 128:1731–1736. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reeves PG, Nielsen FH and Fahey GC Jr:
AIN-93 purified diets for laboratory rodents: Final report of the
American Institute of Nutrition ad hoc writing committee on the
reformulation of the AIN-76A rodent diet. J Nutr. 123:1939–1951.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Croze ML, Géloën A and Soulage CO:
Abnormalities in myo-inositol metabolism associated with type 2
diabetes in mice fed a high-fat diet: Benefits of a dietary
myo-inositol supplementation. Br J Nutr. 113:1862–1875. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Takahashi Y: Soy protein and fish oil
independently decrease serum lipid concentrations but interactively
reduce hepatic enzymatic activity and gene expression involved in
fatty acid synthesis in rats. J Nutr Sci Vitaminol (Tokyo).
57:56–64. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang Y, Sitanggang NV, Kato N, Inoue J,
Murakami T, Watanabe T, Iguchi T and Okazaki Y: Beneficial effects
of protease preparations derived from Aspergillus on the
colonic luminal environment in rats consuming a high-fat diet.
Biomed Rep. 3:715–720. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Okazaki Y, Tomotake H, Tsujimoto K, Sasaki
M and Kato N: Consumption of a resistant protein, sericin, elevates
fecal immunoglobulin A, mucins, and cecal organic acids in rats fed
a high-fat diet. J Nutr. 141:1975–1981. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sakamoto K, Vucenik I and Shamsuddin AM:
[3H]phytic acid (inositol hexaphosphate) is absorbed and
distributed to various tissues in rats. J Nutr. 123:713–720. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Indyk HE, Saldo SC, White PM, Dole MN,
Gill BD and Woollard DC: The free and total myo-inositol contents
of early lactation and seasonal bovine milk. Int Dairy J. 56:33–37.
2016. View Article : Google Scholar
|
|
24
|
Masuda T, Kawano A, Kitahara K, Nagashima
K, Aikawa Y and Arai S: Quantitative determination of sugars and
myo-inositol in citrus fruits grown in Japan using high-performance
anion-exchange chromatography. J Nutr Sci Vitaminol (Tokyo).
49:64–68. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu RY, Wan YP, Fang QY, Lu W and Cai W:
Supplementation with probiotics modifies gut flora and attenuates
liver fat accumulation in rat nonalcoholic fatty liver disease
model. J Clin Biochem Nutr. 50:72–77. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ritze Y, Bárdos G, Claus A, Ehrmann V,
Bergheim I, Schwiertz A and Bischoff SC: Lactobacillus
rhamnosus GG protects against non-alcoholic fatty liver disease
in mice. PLoS One. 9:e801692014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pagano C, Soardo G, Esposito W, Fallo F,
Basan L, Donnini D, Federspil G, Sechi LA and Vettor R: Plasma
adiponectin is decreased in nonalcoholic fatty liver disease. Eur J
Endocrinol. 152:113–118. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Targher G, Bertolini L, Rodella S, Zoppini
G, Scala L, Zenari L and Falezza G: Associations between plasma
adiponectin concentrations and liver histology in patients with
nonalcoholic fatty liver disease. Clin Endocrinol (Oxf).
64:679–683. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu A, Wang Y, Keshaw H, Xu LY, Lam KSL and
Cooper GJS: The fat-derived hormone adiponectin alleviates
alcoholic and nonalcoholic fatty liver diseases in mice. J Clin
Invest. 112:91–100. 2003. View Article : Google Scholar : PubMed/NCBI
|