Introduction
Mitochondria are the primary site of energy
production via the process of oxidative phosphorylation (OXPHOS).
This process involves the transfer of electrons from reduced
nicotine adenine dinucleotide (NADH) or flavin adenine dinucleotide
(FADH2) to oxygen through highly conserved mitochondrial
membrane-bound enzyme complexes (I-V) of the electron transport
chain (ETC) to create ATP (1).
Mitochondria are also an essential source of reactive oxygen
species (ROS) generation as by-products of normal mitochondrial
metabolism (2).
One of the mitochondria's unique features is that it
contains its own genome (mtDNA), separate and distinct from the
nuclear genome of the cell. Human mtDNA is a double-stranded and
circular molecule of 16,569 bp and contains two regions (3). The coding region encompasses 37 genes,
which encode 13 crucial protein subunits of the ETC, two ribosomal
(r)RNAs, and 22 transfer (t)RNAs. The control or regulatory
(D-loop) region consists of sites for replicating and transcribing
of the mtDNA. Except for complex II subunits, which are entirely
encoded by the nuclear DNA (nDNA), subunits of complex I, III, IV
and V are encoded by both nDNA and mtDNA. Specifically, mtDNA codes
for seven subunits (ND1, ND2, ND3, ND4, ND4L, ND5 and ND6) of
NADH-ubiquinone oxidoreductase of complex I, cytochrome b
(CYTB) subunit of ubiquinol-cytochrome c oxidoreductase of
complex III, three subunits (CO1, CO2 and CO3) of cytochrome
c oxidase of complex IV and two subunits (ATPase 6 and 8) of
ATP synthase of the complex.
The mtDNA is particularly susceptible to oxidative
damage and has a high mutational rate due to its proximity to the
site of ROS production, the lack of protective histones, and low
DNA repair capacity (4,5). Since mtDNA encodes essential components
of the ETC, these mutations can disrupt the mitochondria's ability
to generate energy for the cell (6).
Indeed, mtDNA mutations are linked with a wide range of human
diseases (6).
Although primary mutations in the mtDNA have been
observed in diseases of mitochondrial origin, secondary mutations
and new variants are also involved in aging (7,8) and may
underlie the predisposition of several common diseases, such as
neurodegenerative, metabolic and inflammatory conditions (8-10).
It is therefore useful to sequence the complete mitochondrial
genome to explore disease-related variants in the mtDNA (11,12).
Psoriasis (Ps) is a chronic immune-mediated
inflammatory skin disease characterized by hyperproliferative
keratinocytes and the infiltration of the dermis by various immune
cells (13,14). Ps affects ~3% of the population
worldwide (15), and its incidence
is also high in the Gulf countries, including Kuwait, where it
affects around ~3% of people (16-18).
Several studies have shown an association between Ps and metabolic
syndrome (19-21),
particularly type 2 diabetes (T2D), in which T2D was found to be
twice as prevalent in patients with Ps (22). T2D is a progressive metabolic disease
characterized by hyperglycaemia due to inadequate insulin secretion
from the β-cells and insulin resistance. T2D is a leading cause of
severe vascular complications, including cardiovascular disease
(23,24), which is frequently observed in Ps
patients (25,26).
Whilst the nature of the relationship between Ps and
T2D remains ambiguous, both of these diseases are multifactorial,
involving an interplay between genetic and environmental factors
(27). Amongst the genetic factors
that may explain the co-occurrence of Ps and T2D, variations in
mtDNA have been suggested. In this context, studies have shown a
potential role of mtDNA variants in the susceptibility or risk of
T2D in different populations, including in Asian (28), European (29) and other populations (30,31).
Similarly, the role of mtDNA variants in Ps has been observed in a
European population (32). However,
these studies have demonstrated ethnic diversity in the
distribution or the implications of mtDNA variants in Ps and
T2D.
To date, there are no studies that have investigated
variations in the mtDNA in patients with Ps alone or in patients
with Ps and T2D (Ps-T2D) in the Arab population, to the best of our
knowledge. Therefore, this study aimed to sequence and compare
whole mitochondrial genomes from Kuwaiti subjects with Ps, T2D,
Ps-T2D and healthy controls to identify mtDNA variants in Arab
individuals in Kuwait.
Materials and methods
Study subjects
In the present study, a total of 98 subjects were
enrolled, including 34 patients with Ps without T2D (male age range
34-76, median age 54; female age range 24-64, median age 37), 15
T2D patients with no history of skin diseases (male age range
35-60, median 54; female age range 35-57, median age 50), and 29
patients with Ps-T2D (male age range 43-73, median age 56; female
age range 38-65 and median 51), as well as 20 healthy controls
(male age range 24-57; median age 28; female age range 23-40,
median age 27). T2D patients were diagnosed according to the World
Health Organization criteria (33);
fasting glucose level >7.0 mmol/l and glycated haemoglobin
(HbA1c) levels of >6.5%. Patients diagnosed with plaque Ps with
and without T2D were recruited from the dermatology clinics of
Abdul Kareem Al-Saeed and Suaid Al-Subah Dermatology Centres in the
State of Kuwait. Healthy controls were free from inflammatory
dermatoses or autoimmune diseases and without a history of T2D.
Demographic and clinical parameters were obtained from the medical
reports of all participants. Written informed consent was obtained
from all participants under the protocols of the Joint Committee
for the Protection of Human Subjects in Research in Kuwait. The
study was approved by the Health Science Centre Ethics Committee at
Kuwait University and the Health and Medical Research Committee in
the Ministry of Health in Kuwait.
Blood sampling and genomic DNA
extraction
Whole blood samples (5 ml) were collected from
participants in EDTA tubes. Genomic DNA was extracted from whole
blood using a QIAamp DNA Blood Mini kit (Qiagen GmbH) according to
the manufacturer's protocol, and as previously described (9,34). The
purity of the DNA samples were assessed using a NanoDrop 1000
system (Thermo Fisher Scientific, Inc.) and the concentration was
measured using a Qubit 3.0 Fluorometer (Thermo Fisher Scientific,
Inc.).
Amplification of the mitochondrial
genome
The mitochondrial genome from the extracted DNA was
amplified by PCR using a Precision ID mtDNA Whole Genome Panel
(Applied Biosystems; Thermo Fisher Scientific, Inc.), which
consisted of a 2-pool multiplex assay that targets the entire human
mitochondrial genome. Amplification was performed according to the
manufacturer's protocol. Each pool contained 81 primer pairs, with
minimal primer overlap between pools. The mtDNA tiling approach was
also used to construct the Precision ID mtDNA Control Region Panel
which targets only the genome's control region, and was according
to the manufacturer's protocol.
Mitochondrial genome sequencing
The whole mitochondrial genome was sequenced using
the Ion Torrent S5™XL Next Generation Sequencing system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Library preparation
and purity were performed according to the manufacturer's protocol.
Raw signal data from the Ion Torrent S5 XL sequencing were
automatically transferred to the Torrent Server Hosting the Torrent
Suite Software, which converted the raw voltage semiconductor
sequencing data into DNA base calls. The pipeline included
processing, base calling, quality score assignment, adapter
trimming, read mapping to 19 reference human genomes, quality
control of mapping quality, coverage analysis with down sampling
and variant calling (thermofisher.com/kw/en/home/life-science/sequencing/next-generation-sequencing/ion-torrent-next-generation-sequencing-workflow/ion-torrent-next-generation-sequencing-data-analysis-workflow/ion-reporter-software.html).
Identification of variants was performed using the Ion Torrent
Variant Caller plug-in and Ion Reporter Software version 5.2.
Torrent Variant Caller version 5.2 was used for alignment and
variant detection according to the revised Cambridge Reference
Sequence of the human mitochondrial genome (35). The samples were multiplexed and
sequenced on an Ion 520 chip (3-6 Mb throughput). The average
throughput of the Ion 520 chip was 3.5 Mb. The datasets have been
registered in the Sequence Read Archive (SRA) repository with
reference PRJNA699142 (Table
SI).
Statistical analysis
SPSS version 15.0 (SPSS, Inc.) was used for
statistical analysis. Comparisons of demographic and clinical
parameters of multiple groups were performed using ANOVA followed
by a post hoc Tukey's LSD test. Pearson's χ2 was used to
assess differences in the mtDNA variants distribution between cases
and control. The results were evaluated with 95% confidence
intervals (CIs), and P<0.05 was considered to indicate a
statistically significant difference. mtDNA variants were
interpreted for disease association using the data from the MITO
synopsis (36), human mitochondrial
database (hmtdb.uniba.it) and CLINVAR database
(ncbi.nlm.nih.gov/clinvar/).
Results
Characteristics of the study
subjects
The study included 98 subjects, 34 patients with Ps,
15 patients with T2D, 29 patients with Ps-T2D and 20 healthy
controls. Table I shows the
characteristics of the study subjects. There was no significant
difference in the mean age between the study subjects.
Additionally, there was no significant difference in the sex
distribution amongst the study subjects. The mean value of fasting
glucose differed significantly between patients and controls and
was higher in the T2D patients and the Ps-T2D patients compared to
the Ps patients and controls (P<0.001).
 | Table ICharacteristics of study
subjects. |
Table I
Characteristics of study
subjects.
|
Characteristics | Ps | T2D | Ps-T2D | Controls |
|---|
| Sex, % | | | | |
|
Male | 56 | 47 | 59 | 50 |
|
Female | 44 | 53 | 41 | 50 |
| Age range (median),
year | | | | |
|
Male | 34-76(54) | 35-60(54) | 43-73(56) | 24-57(28) |
|
Female | 24-64(37) | 35-57(50) | 38-65(51) | 23-40(27) |
| Fasting glucose,
mmol/lc | 5.4±0.68 |
10.0±3.0a,b | 10.6±4a,b | 5.0±0.4 |
| Triglyceride,
mmol/lc |
1.7±1.2a |
2.1±1.1a |
2.5±1.6a | 0.8±0.2 |
| Total cholesterol,
mmol/lc |
5.0±0.8a | 4.5±0.9 | 4.8±1.6 | 4.3±1.0 |
A significant difference in the mean triglyceride
levels amongst the subject groups was observed (P≤0.001). The
triglyceride levels were normal in the Ps patients, but were
borderline high in the T2D patients and high in the Ps-T2D
patients. In contrast, a significant difference in the mean value
of total cholesterol was found between Ps patients compared with
controls (P<0.05), but not between any of the other groups
(P>0.05).
Novel mtDNA mutations in patients
Whole mitochondrial genome sequence analysis
revealed several novel mutations that were not previously reported,
were not associated with disease in the MitoMap, and are not listed
in the Single Nucleotide Polymorphism Database (36). These included synonymous and
non-synonymous mutations detected in patients with Ps, T2D and
Ps-T2D, which were not present in the controls. The identified
mutations and their characteristics are displayed in Tables
II-IV. The majority of the non-synonymous mutations were found
in the mtDNA coding region. Most of these were observed in subunit
genes of complex I, including ND2, ND4 and
ND5. The remaining mutations were found in the CYB
gene of complex III and ATP8 subunit gene of complex V.
Additionally, the synonymous mutations were found in the mtDNA
coding and control regions.
In the Ps group (Table
II), two missense mutations were found in the ND genes.
These included G5262A in the ND2 gene and A12397G in the
ND5 gene. The identified synonymous mutations in Ps patients
(Table II) were A3711G in the
ND1 gene, T5093C and C5303T in the ND2 gene, A10286G
in the ND3 gene, A10816G in the ND4 gene, T10667C in
the ND4L gene, A13101C in the ND5 gene and T6524C in
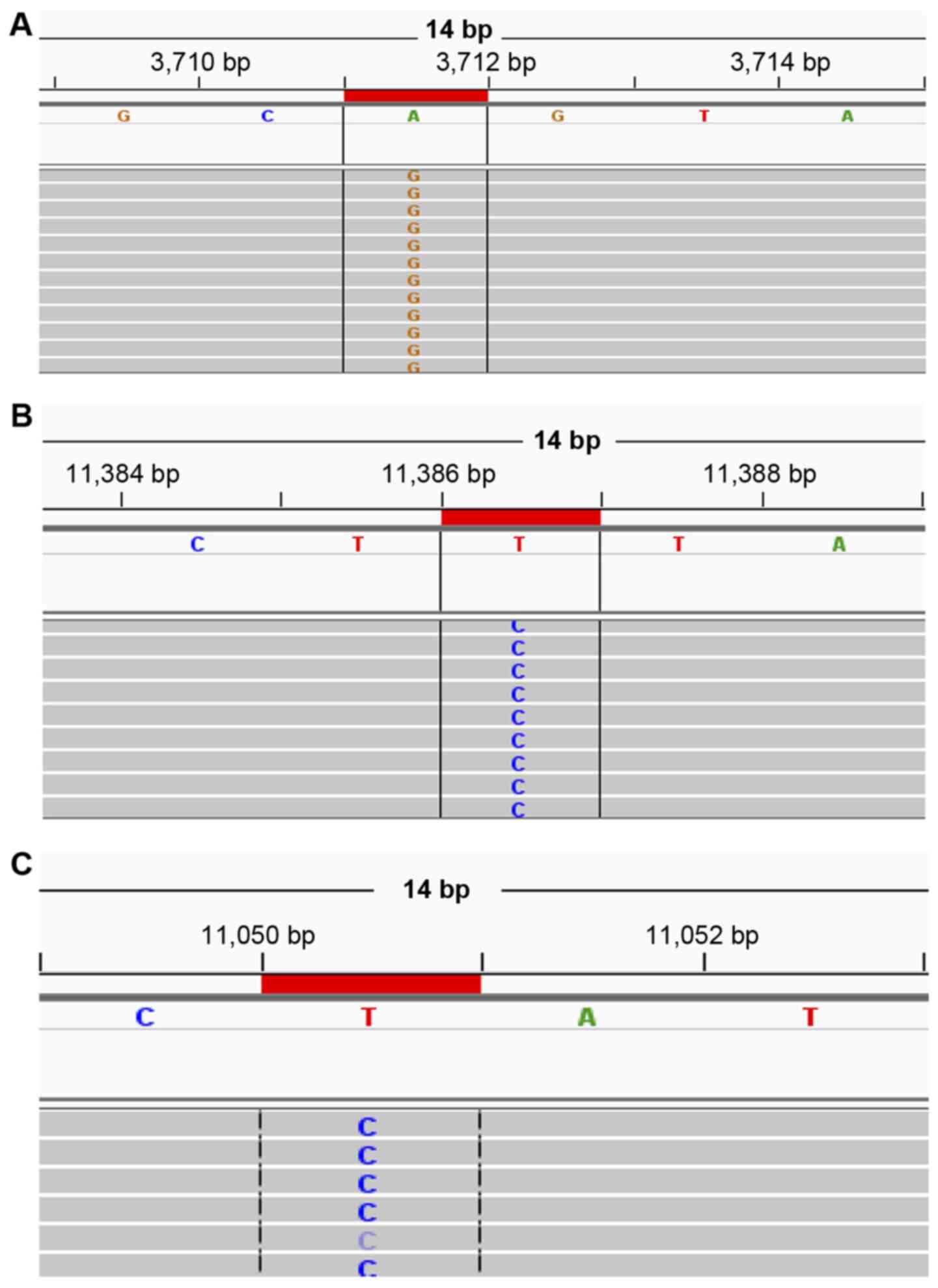
the CO1 gene. (Fig. 1A;
histograms show the A3711G synonymous mutation in the Ps
group).
| Table IINovel mitochondrial DNA mutations
present in the psoriasis patients. |
Table II
Novel mitochondrial DNA mutations
present in the psoriasis patients.
| Gene | Nucleotide
change | Amino acid
change | Type of
mutation | Nature of
mutation |
|---|
| ND1 | A3711G | No change | Synonymous | Homoplasmic |
| ND2 | T5093C | No change | Synonymous | Homoplasmic |
| ND2 | C5303T | No change | Synonymous | Homoplasmic |
| ND3 | A10286G | No change | Synonymous | Homoplasmic |
| ND4 | A10816G | No change | Synonymous | Homoplasmic |
| ND4L | T10667C | No change | Synonymous | Homoplasmic |
| ND5 | A13101C | No change | Synonymous | Homoplasmic |
| CO1 | T6524C | No change | Synonymous | Homoplasmic |
| ND2 | G5262A | Ala265Thr | Missense | Homoplasmic |
| ND5 | A12397G | Thr21Ala | Missense | Homoplasmic |
In the T2D (Table
III), five missense mutations were detected. The missense
mutations included, C12084T, G4959A and A11930G in the ND
genes, C14751T in the CYB gene, as well as T8951C in the
ATP6 gene. Moreover, the T1822C and T2226TA insertion
mutations were found in the 16S rRNA gene. The identified
synonymous mutations in the group of T2D patients (Table III) were T11386C, G11887A, T12136C,
C13077A, C13680T, T5196C, T14020C and A14500G in the ND
genes, C7648T and T7783C in the CO2 gene, and T15310C in the
CYB gene as well as the A16316G variant in the D-loop
control region (Fig. 1B.; histograms
show the T11386C synonymous mutation in the T2D group).
| Table IIINovel mitochondrial DNA mutations
present in the type 2 diabetes patients. |
Table III
Novel mitochondrial DNA mutations
present in the type 2 diabetes patients.
| Gene | Nucleotide
change | Amino acid
change | Type of
mutation | Nature of
mutation |
|---|
| ND4 | T11386C | No change | Synonymous | Homoplasmic |
| ND4 | G11887A | No change | Synonymous | Homoplasmic |
| ND4 | C12084T | Ser 442 Pro | Missense | Homoplasmic |
| ND4 | T12136C | No change | Synonymous | Homoplasmic |
| ND5 | C13077A | No change | Synonymous | Homoplasmic |
| ND5 | C13680T | No change | Synonymous | Homoplasmic |
| CYB | C14751T | Thr2IIe | Missense | Homoplasmic |
| CYB | T15310C | No change | Synonymous | Homoplasmic |
| CO2 | C7648T | No change | Synonymous | Homoplasmic |
| CO2 | T7783C | No change | Synonymous | Homoplasmic |
| ND2 | G4959A | Ala164Thr | Missense | Heteroplasmic |
| ND2 | T5196C | No change | Synonymous | Homoplasmic |
| ND4 | A11930G | IIe391Val | Missense | Homoplasmic |
| ND5 | T14020C | No change | Synonymous | Homoplasmic |
| ND6 | A14500G | No change | Synonymous | Homoplasmic |
| ATP6 | T8951C | Val142Ala | Missense | Homoplasmic |
| 16S
rRNA | T1822C | - | - | Homoplasmic |
| 16S
rRNA | T2226TA | - | - | Homoplasmic |
| D-loop | A16316G | - | - | Homoplasmic |
In the Ps-T2D (Table
IV), the C15735T missense mutation was found in the CYB
gene. The identified synonymous mutations in the Ps-T2D were
T5090C, T11050C, C10556T, C10628T, A13419T and A3720G in the
ND genes (Fig. 1C; histograms
show the T11050C synonymous mutation in PsT2D group).
| Table IVNovel mitochondrial DNA mutations
present in the psoriasis patients with type 2 diabetes. |
Table IV
Novel mitochondrial DNA mutations
present in the psoriasis patients with type 2 diabetes.
| Gene | Nucleotide
change | Amino acid
change | Type of
mutation | Nature of
mutation |
|---|
| ND2 | T5090C | No change | Synonymous | Homoplasmic |
| ND4 | T11050C | No change | Synonymous | Homoplasmic |
| ND4L | C10556T | No change | Synonymous | Homoplasmic |
| ND4L | C10628T | No change | Synonymous | Homoplasmic |
| ND5 | A13419T | No change | Synonymous | Homoplasmic |
| CYB | C15735T | Ala330Val | Missense | Homoplasmic |
| ND1 | A3720G | No change | Synonymous | Homoplasmic |
Several other variants that were previously reported
as either missense polymorphisms or synonymous mutations were also
found in all the patient groups and are shown in Table V.
| Table VKnown mitochondrial DNA sequence
variations present only in the patient groups. |
Table V
Known mitochondrial DNA sequence
variations present only in the patient groups.
| A, Ps group |
|---|
| Gene | Nucleotide
change | Amino acid
change | Type of
mutation | dbSNP (rs) |
|---|
| ND4 | A10819G | No change | Synonymous | rs28358283 |
| ND6 | T14212C | No change | Synonymous | rs28357672 |
| CO3 | A9377G | No change | Synonymous | rs28380140 |
| ATPase6 | A8860G | Thr112Ala | Missense | rs2001031 |
| ATPase8 | T8473C | No change | Synonymous | rs386829037 |
| B, T2D group |
| Gene | Nucleotide
change | Amino acid
change | Type of
mutation | dbSNP (rs) |
| ND1 | T3396C | No change | Synonymous | rs374875201 |
| ND1 | C4025T | Thr240Met | Missense | rs397515509 |
| ND1 | T4218C | No change | Synonymous | rs878853061 |
| ND1 | A4234G | Thr310Ala | Missense | rs2001030 |
| ND2 | C5187T | No change | Synonymous | rs879014605 |
| ND5 | G13145A | Ser270Asn | Missense | rs386829175 |
| ND5 | T13326C | No change | Synonymous | rs878889334 |
| ND5 | T14025C | No change | Synonymous | rs879073899 |
| ND6 | T14325C | Asn117Asp | Missense | rs397515505 |
| ND6 | T14577C | IIe33Val | Missense | rs386829219 |
| CYB | G14861A | Ala39Thr | Missense | rs2853505 |
| CO1 | A6891G | Ser330Gly | Missense | rs879091068 |
| CO1 | G7337A | No change | Synonymous | rs386829005 |
| CO2 | C7819A | No change | Synonymous | rs878853024 |
| CO2 | C7873T | No change | Synonymous | rs879161183 |
| CO3 | G9438A | Gly78Ser | Missense | rs267606611 |
| CO3 | T9530C | No change | Synonymous | rs879237361 |
| CO3 | T9950C | No change | Synonymous | rs3134801 |
| ATP6 | C8932T | Pro136Ser | Missense | rs878853013 |
| 12S
rRNA | G1503A | - | - | rs727503164 |
|
tRNAAla | C5601T | - | - | rs376884056 |
|
tRNAThr | CT15939Cdel | - | - | rs878981265 |
| D-loop | T42TC ins | - | - | rs377245343 |
| D-loop | CT151TC | - | - | rs386828863 |
| D-loop | T279C | - | - | rs879199276 |
| D-loop | A512G | - | - | rs1556422458 |
| D-loop | C16167T | - | - | rs371419667 |
| D-loop | T16209C | - | - | rs386829278 |
| C, Ps-T2D
group |
| Gene | Nucleotide
change | Amino acid
change | Type of
mutation | dbSNP (rs) |
| ND2 | A5390G | No change | Synonymous | rs41333444 |
| ND2 | T5426C | No change | Synonymous | rs878866102 |
| ND3 | G10143A | Gly29Ser | Missense | rs202131419 |
| ND4 | A10876G | No change | Synonymous | rs879036391 |
| ND5 | T13020C | No change | Synonymous | rs75577869 |
| ND5 | T13879C | Ser515Pro | Missense | rs879087566 |
| CYB | G15734A | Ala330Thr | Missense | rs386829259 |
| CO1 | C6045T | No change | Synonymous | rs879061193 |
| CO1 | T6515C | No change | Synonymous | rs878998677 |
| 12S
rRNA | G1598A | - | - | rs3135027 |
| 16S
rRNA | T2626C | - | - | rs879158835 |
|
tRNAThr | A15907G | - | - | rs41383248 |
| D-loop | T125C | - | - | rs144402189 |
| D-loop | C340T | - | - | rs117394573 |
| D-loop | A508G | - | - | rs113683159 |
| D-loop | C16214T | - | - | rs368055283 |
| D-loop | C16290T | - | - | rs386828866 |
| D-loop | C16295T | - | - | rs878874012 |
Known mtDNA sequence variations in
patients and controls
Analysis of whole mitochondrial genomes from Ps, T2D
and Ps-T2D patients and controls revealed the presence of numerous
known sequence variations in the coding and control regions of
mtDNA (Table VI). When the
frequency of these variants was compared between patients and
controls, significant results (P<0.05) with odd ratios (OR)>1
were found. Specifically, the G15301A variant in the CYB
gene was found at a higher frequency in the three groups of
patients, and appeared in 32% of the Ps patients (OR, 4.2; 95% CI,
2-9; P=0.0001), 20% of the T2D patients (OR, 2.2; 95% CI, 0.9-5;
P=0.04) and 21% of the Ps-T2D patients (OR, 2.4; 95% CI, 1-5.3;
P=0.04) compared with the controls (10%). Similarly, the C150T
variant in the D-loop was also found at an increased frequency in
the three groups of patients and appeared in 26% of the Ps patients
(OR, 3; 95% CI, 1.4-7; P=0.003), 20% of the T2D patients (OR, 2.2;
95% CI, 0.9-5; P=0.04) and 24% of the Ps-T2D patients (OR, 2.8; 95%
CI, 1.2-6.3l; P=0.008) compared with the controls (10%), whereas
the C12705T variant in the ND5 gene was found at increased
frequency in the Ps and Ps-T2D groups: 35% of Ps patients (OR, 3;
95% CI, 1.5-6; P=0.001) and 28% of the Ps-T2D patients (OR, 2.2;
95% CI, 1-4.4; P=0.03) compared with the controls (15%). The
variant A1438G in the 12S rRNA gene was observed in 100% of
Ps patients (OR, 11; 95% CI, 1.3-8.7; P=0.005) and 100% of the
Ps-T2D patients (OR, 11; 95% CI, 1.3-87; P=0.005) compared with the
controls (90%). Some of the identified variants appeared more
frequently in specific patient groups compared with the controls
(Table VI).
| Table VIKnown mitochondrial DNA sequence
variations in patients and controls. |
Table VI
Known mitochondrial DNA sequence
variations in patients and controls.
| | Ps | T2D | Ps-T2D | Controls |
|---|
| Gene | Nucleotide
change | % | OR, 95% CI | P-value | % | OR, 95% CI | P-value | % | OR, 95% CI | P-value | % |
|---|
| CYB | G15301A | 32 | 4.2, 2-9 | 0.0001c | 20 | 2.2, 0.9-5 | 0.04a | 21 | 2.4, 1-5.3 | 0.04a | 10 |
| D-loop | C150T | 26 | 3, 1.4-7 | 0.003b | 20 | 2.2, 0.9-5 | 0.04a | 24 | 2.8, 1.2-6.3 | 0.008b | 10 |
| ND5 | C12705T | 35 | 3, 1.5-6 | 0.001c | | | | 28 | 2.2, 1-4.4 | 0.03a | 15 |
| 12S
rRNA | A1438G | 100 | 11, 1.3-8.7 | 0.005b | | | | 100 | 11, 1.3-87 | 0.005b | 90 |
| ND3 | C10400T | 18 | 4, 1.4-11.7 | 0.003b | | | | | | | 5 |
| ND4 | T10873C | 35 | 3, 1.5-6 | 0.001c | | | | | | | 15 |
| CYB | T14783C | 18 | 4, 1.4-11.7 | 0.003b | | | | | | | 5 |
| CO3 | T9540C | 35 | 3, 1.5-6 | 0.001c | | | | | | | 15 |
| ATPase
6 | A8701G | 35 | 2, 1.1-4 | 0.01b | | | | | | | 20 |
| D-loop | C16223T | 38 | 2.4, 1.3-4.6 | 0.005b | | | | | | | 20 |
| D-loop | T16519C | 65 | 1.8, 1-3 | 0.03a | | | | | | | 50 |
| ND2 | A4769G | | | | 27 | 3.3, 1.5-7.3 | 0.002b | | | | 10 |
| ND4 | G11914A | | | | 26 | 3, 1.4-7 | 0.003b | | | | 10 |
| ND5 | C12633A | | | | 13 | 2.8, 0.9-8 | 0.04a | | | | 5 |
| ND5 | G13368A | | | | 20 | 2.2, 0.9-5 | 0.04a | | | | 10 |
| ND5 | G13590A | | | | 13 | 2.2, 0.9-5 | 0.04a | | | | 5 |
| ND6 | G14364A | | | | 13 | 2.2, 0.9-5 | 0.04a | | | | 5 |
| CYB | G15148A | | | | 13 | 2.8, 0.9-8 | 0.04a | | | | 5 |
| CYB | A15607G | | | | 20 | 2.2, 0.9-5 | 0.04a | | | | 10 |
|
tRNAThr | G15928A | | | | 20 | 2.2, 0.9-5 | 0.04a | | | | 10 |
|
tRNAArg | T10463C | | | | 20 | 2.2, 0.9-5 | 0.04a | | | | 10 |
| 16S
rRNA | G1719A | | | | 13 | 2.8, 0.9-8 | 0.04a | | | | 5 |
| 16S
rRNA | G1888A | | | | 20 | 2.8, 0.9-8 | 0.04a | | | | 10 |
| D-loop | G16274A | | | | 13 | 2.8, 0.9-8 | 0.04a | | | | 5 |
| D-loop | C16292T | | | | 13 | 2.8, 0.9-8 | 0.04a | | | | 5 |
| D-loop | C16294T | | | | 27 | 2, 1-4 | 0.03a | | | | 15 |
| D-loop | T195C | | | | 46 | 2.5, 1.4-4.6 | 0.002b | | | | 25 |
| D-loop | C16186T | | | | 13 | 2.8, 0.9-8 | 0.04a | | | | 5 |
|
tRNAArg | T10410C | | | | | | | 14 | 3, 1-9 | 0.02a | 5 |
| D-loop | G16390A | | | | | | | 14 | 3, 1-9 | 0.02a | 5 |
In the Ps group, higher frequencies of variants
(OR>1, P<0.05) were observed, namely C10400T and T10873C in
the ND genes, T14783C in the CYB gene, T9540C in the
CO3 gene, and A8701G in the ATPase 6 gene, as well as
C16223T and T16519C in the D-loop control region.
In the T2D group, increased frequencies of variants
(OR>1, P<0.05) were found in the coding region, including
A4769G, G11914A, C12633A, G13368A, G13590A and G14364A in the
ND genes, G15148A and A15607G in the CYB gene,
G15928A in the tRNAThr gene, T10463C in the
tRNAArg gene, and G1719A and G1888A in the 16S
rRNA gene. Variants in the D-loop control region, namely T195C,
C16186T, G16274A, C16292T and C16294T were also found.
In the Ps-T2D group, increased frequencies (OR>1,
P<0.05) were observed for the T10410C variant in the
tRNAArg gene and the G16390A variant in the
D-loop region.
When these variants' characteristics were analysed
(Table VII), the majority of the
identified variants were homoplasmic with no amino acid changes.
However, the A8701G variant in the ATPase 6 gene, which was
located at a higher frequency in 35% of the Ps patients (OR, 2; 95%
CI, 1.1-4; P=0.01) compared with 20% in the controls, was
identified as a missense mutation and exhibited a threonine to
alanine alteration (Thr59Ala).
| Table VIICharacteristics of mitochondrial DNA
sequence variations in patients and controls. |
Table VII
Characteristics of mitochondrial DNA
sequence variations in patients and controls.
| Gene | Nucleotide
change | Amino acid
change | Type of
mutation | Nature of
mutation | dbSNP (rs) |
|---|
| CYB | G15301A | No change | Synonymous | Homoplasmic | rs193302991 |
| D-loop | C150T | - | - | Homoplasmic | rs62581312 |
| ND5 | C12705T | No change | Synonymous | Homoplasmic | rs193302956 |
| 12S
rRNA | A1438G | - | - | Homoplasmic | rs2001030 |
| ND3 | C10400T | No change | Synonymous | Homoplasmic | rs28358278 |
| ND4 | T10873C | No change | Synonymous | Homoplasmic | rs2857284 |
| CYB | T14783C | No change | Synonymous | Homoplasmic | rs193302982 |
| CO3 | T9540C | No change | Synonymous | Homoplasmic | rs2248727 |
| ATPase
6 | A8701G | p. Thr59Ala | Missense | Homoplasmic | rs2000975 |
| D-loop | C16223T | - | - | Homoplasmic | rs2853513 |
| D-loop | T16519C | - | - | Homoplasmic | rs3937033 |
| ND2 | A4769G | No change | Synonymous | Homoplasmic | rs3021086 |
| ND4 | G11914A | No change | Synonymous | Homoplasmic | rs2853496 |
| ND5 | C12633A | No change | Synonymous | Homoplasmic | rs3926883 |
| ND5 | G13368A | No change | Synonymous | Homoplasmic | rs3899498 |
| ND5 | G13590A | No change | Synonymous | Homoplasmic | rs28359177 |
| ND6 | G14364A | No change | Synonymous | Homoplasmic | rs879086798 |
| CYB | G15148A | No change | Synonymous | Homoplasmic | rs527236206 |
| CYB | A15607G | No change | Synonymous | Homoplasmic | rs193302996 |
|
tRNAThr | G15928A | - | - | Homoplasmic | rs527236198 |
|
tRNAArg | T10463C | - | - | Homoplasmic | rs28358279 |
| 16S
rRNA | G1719A | - | - | Homoplasmic | rs3928305 |
| 16S
rRNA | G1888A | - | - | Homoplasmic | rs2897260 |
| D-loop | G16274A | - | - | Homoplasmic | rs144095641 |
| D-loop | C16292T | - | - | Homoplasmic | rs144417390 |
| D-loop | C16294T | - | - | Homoplasmic | rs140662392 |
| D-loop | T195C | - | - | Homoplasmic | rs66492218 |
| D-loop | C16186T | - | - | Homoplasmic | rs879166752 |
| D-loop | G16390A | - | - | Homoplasmic | rs41378955 |
|
tRNAArg | T10410C | - | - | Homoplasmic | rs200478835 |
Discussion
To the best of our knowledge, this is the first
study to perform a comprehensive analysis of mitochondrial DNA
(mtDNA) variants in Kuwaiti subjects with Ps, T2D and Ps-T2D, as
well as in healthy controls. The average coverage depth was
24625.2X and the mean read length was 144 bp. However, the average
total reads were 3,359,441, the frequency of reads was between
99.4-99.9%, and the coverage of reads was >100%. Whole
mitochondrial genome sequencing revealed 36 novel non-synonymous
and synonymous mutations and 51 sequence variations in the patient
groups that were not detected in the controls. Additionally,
several known sequence variations were seen in both patients and
controls.
In general, a synonymous mutation is the
substitution of a DNA base pair that does not result in a change in
the amino acid sequence; in contrast, a non-synonymous mutation is
the substitution of a DNA base pair that results in a single amino
acid change in a given polypeptide. Non-synonymous mutations
include a missense mutation (a point mutation in which a single
nucleotide change results in a codon that codes for a different
amino acid), a nonsense mutation (a point mutation in a sequence of
DNA that leads to the appearance of a stop codon, resulting in
premature termination of translation and the production of a
truncated protein), as well as insertion and deletion of one or
more DNA base pairs.
Amongst the novel mutations identified in the
patient groups, eight non-synonymous mutations resulted in amino
acid changes and were detected primarily in the subunit genes of
complexes I, III and V. These included missense mutations in the Ps
group, primarily found in subunit genes of complex I, including
G5262A in the ND2 gene and A12397G in the ND5 gene.
Moreover, missense mutations were detected in the T2D group. These
included the missense mutations C12084T and A11930G in the
ND4 gene and G4959A in the ND2 gene, as well as the
missense mutations C14751T in the CYB gene, and T8951C in
the ATP6 gene. Additionally, the C15735T missense mutation
in the CYB gene was found in the Ps-T2D group Moreover, 25
synonymous mutations were located in the coding and control regions
in patient groups. Known variants previously reported as either
missense or synonymous mutations were also identified. The majority
of these were located in the coding region, and only a few were
found in the control region.
Other known sequence variations were found in the
patients groups and controls. Some of these variations were
observed more frequently in all patient groups compared with the
controls. Specifically, the frequencies of the G15301A variant in
the CYB gene and the variant C150T in the D-loop region were
significantly higher in all patient groups compared with the
control. In contrast, the C12705T mutation in the ND5 gene
and the A1438G in the 12S rRNA gene were found at
significantly higher frequencies in the Ps and Ps-T2D groups
compared with the control group. Moreover, other variants were
found at higher frequencies in specific patient groups compared
with the control group. Whilst most of these variants were
synonymous, the A8701G variant in the ATPase6 gene that was
found at a higher frequency in the Ps patients compared with the
control group was identified as a missense mutation and resulted in
an amino-acid substitution from threonine to alanine
(Thr59Ala).
Mitochondria are the primary intracellular site of
energy production, and mutations in the mitochondrial genome can
affect mitochondrial function (6).
In humans, the mtDNA encodes 13 protein subunits of the ETC, two
rRNAs and 22 tRNAs, all of which are important for normal
mitochondrial function (3).
Mitochondria are also prone to damage from ROS, and several
mutations of the mtDNA-encoded genes can enhance ROS production
(37). Indeed, mitochondrial
impairment as a result of mtDNA mutations have been observed in
somatic tissues during normal aging (7,8), and
have also been linked to several diseases, where oxidative stress
serves a pivotal role in their development, such as in cancer and
neurodegenerative diseases (6,7,9). Moreover, mitochondrial dysfunction
serves a role in the pathogenesis of non-alcoholic fatty liver
disease (NAFLD), as it affects hepatic lipid homeostasis and
promotes ROS production and lipid peroxidation, and NAFLD has been
linked to both T2D and psoriasis (38).
In the present study, the identified missense, and
insertion mutations in the mtDNA genes were only observed in the
patient groups. Although the identified mutations were homoplasmic,
they showed changes in the amino acids of essential polypeptides
complexes of the mitochondrial ETC, as well as in rRNAs and tRNAs,
which are components of the mitochondrial gene expression system
and the non-coding region. A thousand copies of the mitochondrial
genome per cell gives rise to an essential feature of mitochondrial
genetics: Homoplasmy and heteroplasmy. Homoplasmy is the presence
of identical copies of mtDNA that may be normal or mutated.
Heteroplasmy is the presence of a mixture of normal and mutated
mtDNA. Whereas most deleterious mtDNA mutations are heteroplasmic
in nature, not all are pathogenic, as some heteroplasmic mutations
in the hypervariable D-loop region may be of little clinical
significance (39). Moreover, some
homoplasmic mutations have been reported to cause Leigh syndrome, a
severe neurological disorder (40),
or as secondary mutations that influence the disease severity of
Leber's hereditary optic neuropathy (41). Secondary homoplasmic mutations may
predispose an individual to specific symptoms of T2D, obesity and
Alzheimer's disease from different ethnic groups (31,42).
The present study identified novel mutations that
met at least 3 criteria classified as disease-causing mutations
(6); they were present in
structurally and functionally important regions of the mtDNA,
resulted in changes in the amino acids, and were not found in
healthy individuals. Therefore, these mutations may have
detrimental effects on the structure and function of the ETC
complexes. Notably, in the present study, most novel mutations were
found in the NADH dehydrogenase subunit genes of complex I, the
largest enzyme of the mitochondrial OXPHOS system, and the primary
source of ROS in mitochondria (43).
Altered complex I activity has been frequently observed in various
pathologies such as mitochondrial disorders, cancer,
neurodegenerative diseases and T2D (9,44,45).
The results of the present also showed several
synonymous mutations in patient groups. Although mutations that do
not result in amino changes are considered biologically silent,
they have been implicated in human diseases through their direct
effect on gene expression and function (46-48).
In addition to mtDNA pathogenic mutations, which are
rare in a population, mtDNA polymorphisms have been linked with the
susceptibility to or protection from various diseases. In this
context, previous population-based studies have found an
association between mtDNA variants with the susceptibility and risk
of T2D (28-31),
whereas a protective effect of mtDNA variants from Ps have also
been identified (32).
The current study identified numerous reported mtDNA
variations that are already present in the MITOMAP database, which
were found more frequently in ≥1 group of patients compared with
the controls. Although most of these were homoplasmic synonymous
variants with no amino acid changes, they were reported in several
disease conditions. The variants G15301A in the CYB gene of
complex III and the variant C150T in the hypervariable segment of
the D-loop region were found more frequently in all patient groups
(Ps, T2D and Ps-T2D) compared with the controls. These variants
have not been reported in any of the abovementioned diseases, but
were previously reported in other conditions. The G15301A variant
was described as a germline homoplasmic mtDNA mutation in 40% of
Malaysian females with breast cancer (49), whereas the C150T variant was
associated with the risk of cervical cancer and HPV infection
(50). The T10410C variant in the
tRNAArg gene and the G16390A variant in the
D-loop region were found at increased frequencies in Ps-T2D
patients compared with the controls. The T10410C variant was
previously reported in children with Leigh syndrome (51), and the G16390A variant was found to
be weakly associated with T2D in a Tunisian cohort (52). The variant A8701G in the ATPase
6 gene was found at a higher frequency in Ps patients compared
with controls. This homoplasmic variant was previously reported in
Japanese patients with T2D (53) and
patients with mitochondrial maternally inherited diabetes and
deafness (31). Numerous studies
have shown a clear association between Ps and T2D, and patients
with Ps are at increased risk of developing T2D (19-22).
The presence of the G16390A variant in Ps-T2D patients and the
A1438G variant in the Ps patients and their previous association
with T2D suggest a possible role of these variants to
predisposition of Ps and Ps-T2D.
In the present study, the variant A8701G, which
occurred at a higher frequency in the Ps patients compared with the
controls, was identified as a missense mutation and resulted in
amino-acid substitution from threonine to alanine (Thr59Ala) in the
ATPase6 subunit of complex V. This variant was previously
associated with maternally inherited hypertension and
cardiomyopathy in a Chinese pedigree of consanguineous marriage
(54). Although the Ps patients
triglyceride and total cholesterol levels were normal in the
present study, Ps patients are at higher risk of developing
cardiovascular diseases (25,26).
The present study has some limitations, including
the relatively low number of subjects affecting the statistical
power. Additionally, functional analysis should be performed to
determine the potential biological significance of these mutations
in the context of these diseases, which is lacking from the present
study.
In conclusion, the present study is the first study
to sequence and analyse the whole mitochondrial genome of Kuwaiti
patients with Ps, T2D and Ps-T2D, and compared these with healthy
controls. Novel mutations in patients that resulted in a change in
the coded amino acid, which may be co-responsible in the
determination of these diseases were identified. Additionally,
known variants were detected in higher frequencies in the patient
group compared with the controls, suggesting their role in
predisposing patients to these diseases. These results warrant
further functional analysis to determine the role of these variants
in T2D, Ps and Ps-T2D.
Supplementary Material
The URLs of the datasets registered in
the Sequence Read Archive (SRA) repository
Acknowledgements
We would like to thank Dr Ayda A.Ghader, Dr Shatha
Al. Roomi, and Dr Sakeena Salama, Consultant Dermatologists,
Ministry of Health who referred the majority of the psoriasis
patients included in this study. Additionally, we would like to
thank Ms Mona Alateeqi and Dr Shakir Bahzad (Molecular Genetics
Laboratory, Yacob Behbehani Center) for facilitating the use of the
NGS laboratory.
Funding
Funding: This study was supported by a scholarship program from
Kuwait University.
Availability of data and materials
The datasets generated in the present study have
been registered in the Sequence Read Archive repository (ref. no.
PRJNA699142). The reference BioSample accession nos. are
SAMN17766667-SAMN17766764, and the URLs of the datasets have been
uploaded as a supplementary file (Table
S1).
Author's contributions
MSA and SA conceived the study; MSA collected the
data and performed the experiments; MSA, GAK and MB contributed to
data analysis and interpretation, and wrote and edited the
manuscript. MB and GAK confirmed the authenticity of all the raw
data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
This study was performed in line with the principle
of the Declaration of Helsinki. Approval was granted by the Health
Science Center Ethics Committee at Kuwait University and Health and
Medical Research Committee in the Ministry of Health and registered
on No. 2016/496. Informed consent was obtained from all individual
participants included in the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Smeitink J, van den Heuvel L and DiMauro
S: The genetics and pathology of oxidative phosphorylation. Nat Rev
Genet. 2:342–352. 2001.PubMed/NCBI View
Article : Google Scholar
|
|
2
|
Andreyev AY, Kushnareva YE and Starkov AA:
Mitochondrial metabolism of reactive oxygen species. Biochemistry
(Mosc). 70:200–214. 2005.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Taanman JW: The mitochondrial genome:
Structure, transcription, translation and replication. Biochim
Biophys Acta. 1410:103–123. 1999.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Bohr VA: Repair of oxidative DNA damage in
nuclear and mitochondrial DNA, and some changes with aging in
mammalian cells. Free Radic Biol Med. 32:804–812. 2002.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Santos JH, Hunakova L, Chen Y, Bortner C
and Van Houten B: Cell sorting experiments link persistent
mitochondrial DNA damage with loss of mitochondrial membrane
potential and apoptotic cell death. J Biol Chem. 278:1728–1734.
2003.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Taylor RW and Turnbull DM: Mitochondrial
DNA mutations in human disease. Nat Rev Genet. 6:389–402.
2005.PubMed/NCBI View
Article : Google Scholar
|
|
7
|
Zapico SC and Ubelaker DH: mtDNA Mutations
and their role in aging, diseases and forensic sciences. Aging Dis.
4:364–380. 2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Li H, Slone J, Fei L and Huang T:
Mitochondrial DNA variants and common diseases: A mathematical
model for the diversity of age-related mtDNA mutations. Cells.
8(8)2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Alharbi MA, Al-Kafaji G, Khalaf NB,
Messaoudi SA, Taha S, Daif A and Bakhiet M: Four novel mutations in
the mitochondrial ND4 gene of complex I in patients with multiple
sclerosis. Biomed Rep. 11:257–268. 2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Escames G, López LC, García JA,
García-Corzo L, Ortiz F and Acuña-Castroviejo D: Mitochondrial DNA
and inflammatory diseases. Hum Genet. 131:161–173. 2012.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Da Pozzo P, Cardaioli E, Radi E and
Federico A: Sequence analysis of the complete mitochondrial genome
in patients with mitochondrial encephaloneuromyopathies lacking the
common pathogenic DNA mutations. Biochem Biophys Res Commun.
324:360–364. 2004.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Fauser S, Luberichs J, Besch D and
Leo-Kottler B: Sequence analysis of the complete mitochondrial
genome in patients with Leber's hereditary optic neuropathy lacking
the three most common pathogenic DNA mutations. Biochem Biophys Res
Commun. 295:342–347. 2002.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Peters BP, Weissman FG and Gill MA:
Pathophysiology and treatment of psoriasis. Am J Health Syst Pharm.
57:645–659; quiz 660-661. 2000.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Nestle FO, Kaplan DH and Barker J:
Psoriasis. N Engl J Med. 361:496–509. 2009.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Langley RG, Krueger GG and Griffiths CE:
Psoriasis: Epidemiology, clinical features, and quality of life.
Ann Rheum Dis. 64 (Suppl 2):ii18–ii23; discussion ii24-ii25.
2005.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Shelleh HH and Al-Hatiti HS: Pattern of
skin diseases in a hospital in southwestern Saudi Arabia. Saudi Med
J. 25:507–510. 2004.PubMed/NCBI
|
|
17
|
Alakloby OM: Pattern of skin diseases in
Eastern Saudi Arabia. Saudi Med J. 26:1607–1610. 2005.PubMed/NCBI
|
|
18
|
al-Fouzan AS and Nanda A: A survey of
childhood psoriasis in Kuwait. Pediatr Dermatol. 11:116–119.
1994.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Gisondi P, Tessari G, Conti A, Piaserico
S, Schianchi S, Peserico A, Giannetti A and Girolomoni G:
Prevalence of metabolic syndrome in patients with psoriasis: A
hospital-based case-control study. Br J Dermatol. 157:68–73.
2007.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Lønnberg AS, Skov L, Skytthe A, Kyvik KO,
Pedersen OB and Thomsen SF: Association of psoriasis with the risk
for type 2 diabetes mellitus and obesity. JAMA Dermatol.
152:761–767. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Gelfand JM: Psoriasis, Type 2 Diabetes
Mellitus, and Obesity: Weighing the Evidence. JAMA Dermatol.
152:753–754. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Cohen AD, Sherf M, Vidavsky L, Vardy DA,
Shapiro J and Meyerovitch J: Association between psoriasis and the
metabolic syndrome. A cross-sectional study. Dermatology.
216:152–155. 2008.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Cade WT: Diabetes-related microvascular
and macrovascular diseases in the physical therapy setting. Phys
Ther. 88:1322–1335. 2008.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Einarson TR, Acs A, Ludwig C and Panton
UH: Prevalence of cardiovascular disease in type 2 diabetes: A
systematic literature review of scientific evidence from across the
world in 2007-2017. Cardiovasc Diabetol. 17(83)2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Neimann AL, Shin DB, Wang X, Margolis DJ,
Troxel AB and Gelfand JM: Prevalence of cardiovascular risk factors
in patients with psoriasis. J Am Acad Dermatol. 55:829–835.
2006.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Gelfand JM, Neimann AL, Shin DB, Wang X,
Margolis DJ and Troxel AB: Risk of myocardial infarction in
patients with psoriasis. JAMA. 296:1735–1741. 2006.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Schwandt A, Bergis D, Dapp A, Ebner S,
Jehle PM, Köppen S, Risse A, Zimny S and Holl RW: Psoriasis and
diabetes: A Multicenter study in 222078 type 2 diabetes patients
reveals high levels of depression. J Diabetes Res.
2015(792968)2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Jiang W, Li R, Zhang Y, Wang P, Wu T, Lin
J, Yu J and Gu M: Mitochondrial DNA mutations associated with type
2diabetes mellitus in Chinese Uyghur population. Sci Rep.
7(16989)2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Poulton J, Luan J, Macaulay V, Hennings S,
Mitchell J and Wareham NJ: Type 2 diabetes is associated with a
common mitochondrial variant: Evidence from a population-based
case-control study. Hum Mol Genet. 11:1581–1583. 2002.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Charoute H, Kefi R, Bounaceur S, Benrahma
H, Reguig A, Kandil M, Rouba H, Bakhchane A, Abdelhak S and Barakat
A: Novel variants of mitochondrial DNA associated with type 2
diabetes mellitus in Moroccan population. Mitochondrial DNA A DNA
Mapp Seq Anal. 29:9–13. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Crispim D, Estivalet AAF, Roisenberg I,
Gross JL and Canani LH: Prevalence of 15 mitochondrial DNA
mutations among type 2 diabetic patients with or without clinical
characteristics of maternally inherited diabetes and deafness. Arq
Bras Endocrinol Metabol. 52:1228–1235. 2008.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Coto-Segura P, Santos-Juanes J, Gómez J,
Alvarez V, Díaz M, Alonso B, Corao AI and Coto E: Common European
mitochondrial haplogroups in the risk for psoriasis and psoriatic
arthritis. Genet Test Mol Biomarkers. 16:621–623. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
World Health O and International Diabetes:
Definition and diagnosis of diabetes mellitus and intermediate
hyperglycaemia. Report of a WHO/IDF Consultation. World Health
Organization, Geneva, 2006.
|
|
34
|
Al-Kafaji G, Aljadaan A, Kamal A and
Bakhiet M: Peripheral blood mitochondrial DNA copy number as a
novel potential biomarker for diabetic nephropathy in type 2
diabetes patients. Exp Ther Med. 16:1483–1492. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Bandelt HJ, Kloss-Brandstätter A, Richards
MB, Yao YG and Logan I: The case for the continuing use of the
revised Cambridge Reference Sequence (rCRS) and the standardization
of notation in human mitochondrial DNA studies. J Hum Genet.
59:66–77. 2014.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Brandon MC, Lott MT, Nguyen KC, Spolim S,
Navathe SB, Baldi P and Wallace DC: MITOMAP: A human mitochondrial
genome database - 2004 update. Nucleic Acids Res. 33:D611–D613.
2005.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Hahn A and Zuryn S: Mitochondrial Genome
(mtDNA) mutations that generate reactive oxygen species.
Antioxidants. 8(8)2019.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Tarantino G, Citro V and Capone D:
Nonalcoholic fatty liver disease: A challenge from mechanisms to
therapy. J Clin Med. 9(15)2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Wong LJ, Liang MH, Kwon H, Park J, Bai RK
and Tan DJ: Comprehensive scanning of the entire mitochondrial
genome for mutations. Clin Chem. 48:1901–1912. 2002.PubMed/NCBI
|
|
40
|
Negishi Y, Hattori A, Takeshita E, Sakai
C, Ando N, Ito T, Goto Y and Saitoh S: Homoplasmy of a
mitochondrial 3697G>A mutation causes Leigh syndrome. J Hum
Genet. 59:405–407. 2014.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Carelli V, Giordano C and d'Amati G:
Pathogenic expression of homoplasmic mtDNA mutations needs a
complex nuclear-mitochondrial interaction. Trends Genet.
19:257–262. 2003.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Itsara LS, Kennedy SR, Fox EJ, Yu S,
Hewitt JJ, Sanchez-Contreras M, Cardozo-Pelaez F and Pallanck LJ:
Oxidative stress is not a major contributor to somatic
mitochondrial DNA mutations. PLoS Genet.
10(e1003974)2014.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Hirst J: Towards the molecular mechanism
of respiratory complex I. Biochem J. 425:327–339. 2009.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Sharma LK, Lu J and Bai Y: Mitochondrial
respiratory complex I: Structure, function and implication in human
diseases. Curr Med Chem. 16:1266–1277. 2009.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Wu J, Luo X, Thangthaeng N, Sumien N, Chen
Z, Rutledge MA, Jing S, Forster MJ and Yan LJ: Pancreatic
mitochondrial complex I exhibits aberrant hyperactivity in
diabetes. Biochem Biophys Rep. 11:119–129. 2017.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Buske OJ, Manickaraj A, Mital S, Ray PN
and Brudno M: Identification of deleterious synonymous variants in
human genomes. Bioinformatics. 29:1843–1850. 2013.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Supek F, Miñana B, Valcárcel J, Gabaldón T
and Lehner B: Synonymous mutations frequently act as driver
mutations in human cancers. Cell. 156:1324–1335. 2014.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Gotea V, Gartner JJ, Qutob N, Elnitski L
and Samuels Y: The functional relevance of somatic synonymous
mutations in melanoma and other cancers. Pigment Cell Melanoma Res.
28:673–684. 2015.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Omasanggar R, Yu CY, Ang GY, Emran NA,
Kitan N, Baghawi A, Falparado Ahmad A, Abdullah MA, Teh LK and
Maniam S: Mitochondrial DNA mutations in Malaysian female breast
cancer patients. PLoS One. 15(e0233461)2020.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Zhai K, Chang L, Zhang Q, Liu B and Wu Y:
Mitochondrial C150T polymorphism increases the risk of cervical
cancer and HPV infection. Mitochondrion. 11:559–563.
2011.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Naess K, Freyer C, Bruhn H, Wibom R, Malm
G, Nennesmo I, von Döbeln U and Larsson NG: MtDNA mutations are a
common cause of severe disease phenotypes in children with Leigh
syndrome. Biochim Biophys Acta. 1787:484–490. 2009.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Hsouna S, Ben Halim N, Lasram K, Arfa I,
Jamoussi H, Bahri S, Ammar SB, Miladi N, Abid A, Abdelhak S, et al:
Association study of mitochondrial DNA polymorphisms with type 2
diabetes in Tunisian population. Mitochondrial DNA. 26:367–372.
2015.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Tawata M, Ohtaka M, Iwase E, Ikegishi Y,
Aida K and Onaya T: New mitochondrial DNA homoplasmic mutations
associated with Japanese patients with type 2 diabetes. Diabetes.
47:276–277. 1998.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Zhu Y, Gu X and Xu C: A Mitochondrial DNA
A8701G mutation associated with maternally inherited hypertension
and dilated cardiomyopathy in a Chinese pedigree of a
consanguineous marriage. Chin Med J (Engl). 129:259–266.
2016.PubMed/NCBI View Article : Google Scholar
|