Introduction
Phenotype modulation of arterial smooth muscle cells
(SMCs) is a key process during the development of atherosclerotic
plaque (1,2). Local factors around the vascular wall,
including growth factors, inflammatory mediators, mechanical
stress, injury and lipoprotein, induce a phenotypic switch from a
differentiated contractile state to a dedifferentiated
proliferative state, which is characterized by decreased expression
of contractile type SMC markers represented by α-smooth muscle
actin (α-SMA) (3). When vascular
SMCs are prepared for experiments, they are typically cultured in
the presence of growth factors and thus exhibit a proliferative
(dedifferentiated) phenotype (4).
To the best of our knowledge, there is limited
information on redifferentiation of SMCs, which is reversion to the
differentiated phenotype from the dedifferentiated phenotype
(5). In vitro
redifferentiation in human umbilical artery SMCs was induced by
serum deprivation (6). Similar
redifferentiation was observed in human uterine SMCs on the 6th day
after reaching 100% confluency (7).
There have been reports on molecules that are involved in
redifferentiation of vascular SMCs (8-10):
Hyperplasia suppressor gene has been shown to be involved in
redifferentiation of rat aortic SMCs (RASMCs) (8). C-type natriuremic peptide has been
reported to induce redifferentiation of RASMCs by stimulating the
cGMP cascade (9). Exendin-4 has
also been reported to promote redifferentiation of RASMCs via the
AMPK/sirtuin 1/FOXO3a signaling pathway (10). However, it is unknown whether and
how redifferentiation is induced in vitro in coronary artery
SMCs. Moreover, it remains to be determined whether protein markers
of differentiation also serve as markers of redifferentiation.
Redifferentiation has been shown in SMCs of neointimal tissue
following percutaneous transluminal coronary angioplasty (PTCA) in
autopsied patients (11).
Therefore, it is hypothesized to be clinically relevant to
determine protein markers of redifferentiation of HCASMCs.
The aim of the present study was therefore to
determine the culture conditions for inducing redifferentiation of
human coronary artery SMCs (HCASMCs). Proliferation and migration,
which are phenotypic characteristics of dedifferentiated SMCs, were
monitored, and expression levels of α-SMA, calponin, caldesmon and
SM22α/tagln, known specific differentiation markers of vascular
SMCs (12-14),
were evaluated as potential protein markers of redifferentiated
HCASMCs.
Materials and methods
Cell culture
HCASMCs were purchased from Kurabo Co., Ltd. (cat.
no. KS-4209) and cultured in medium (HuMedia-SB2; Kurabo Co. Ltd)
supplemented with 5% fetal bovine serum, human epidermal growth
factor (hEGF) at 0.5 ng/ml, human fibroblast growth factor B
(hFGF-B) at 2 µg/ml, insulin at 5 µg/ml, gentamicin at 50 µg/ml and
amphotericin B at 50 ng/ml (all obtained from Kurabo Co. Ltd) under
95% air-5% CO2 at 37˚C. For cell observation and protein
extraction or cell migration, cells were seeded at 90% confluency
(day 0) and reached 100% confluency after 48 h incubation (day 2).
Cells were seeded at 90% confluency (3.5x105 cells in a
35-mm dish for cell observation and western blotting and
7.5x105 cells in a 60-mm dish for the migration
experiment) (day 0). In experiments for testing the effects of
removal of growth factors on expression of phenotype marker
proteins, cells were seeded with a lower density of 10% confluency
(2.0x105) in a 100-mm dish in the presence or absence of
the aforementioned growth factors. Phase-contrast and fluorescence
images were captured by microscopy (TE300, Nikon Corporation) with
x10 magnification every day from day 0 to day 6 after seeding the
cells and representative images were obtained. Fluorescence images
were captured after staining with Calcein-AM (sc-203865, Santa Cruz
Biotechnology) for 1 h under 95% air-5% CO2 at 37˚C and
analyzed using a software (Lumina Version, MITANI Corporation,
Ver.3.3.6.1). Other chemicals were purchased from Sigma-Aldrich
(Merck KGaA) unless otherwise stated.
Western blotting
Western blotting was performed as described
previously (15). Percentages of
the SDS-PAGE used were 6% for caldesmon-1, 8% for calponin, α-SMA,
PCNA and GAPDH, and 15% for S100A4 and SM22α/tagln. Proteins were
extracted from HCASMCs at day 1 to day 6 after seeding the cells.
The proteins transferred onto the nitrocellulose membrane following
separation with SDS-PAGE were incubated with primary antibodies at
4˚C overnight. Antibodies to detect α-smooth muscle actin (α-SMA;
1:1,000, cat. no. 19245S), proliferating cell nuclear antigen
(PCNA; 1:2,000, cat. no. 2586S), caldesmon-1 (1:250, cat. no.
2980S), SM22α/tagln (1:1,000, cat. no. 40471S), S100A4 (1:250, cat.
no. 13018S), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH;
1:10,000, cat. no. #2118) were purchased from Cell Signaling
Technology, Inc. A calponin antibody (1:1,000, cat. no. 13938-1-AP)
was obtained from Proteintech Group, Inc. The proteins bound to the
primary antibodies were incubated with secondary antibodies at room
temperature for 1 h. The secondary antibodies used were anti-mouse
IgG, HRP-linked antibody (1:2,000, Cell Signaling, cat. no. #7076)
for PCNA and anti-rabbit IgG, HRP-linked antibody (1:2,000, Cell
Signaling, cat. no. #7074) for α-SMA, calponin, GAPDH, caldesmon-1,
S100A4 and SM22α/tagln. Immunoreactive protein bands were
visualized using an ECL plus detection system (PierceTM
ECL Plus Western Blotting Substrate 32132, Thermo Fisher
Scientific, Inc.) with LAS4000mini (Cytiva). The protein band
intensity was quantified by ImageQuant TL (ver.8.2, Cytiva) and the
relative changes of band intensities were calculated compared with
the control at the earliest time point (designated as 1.0). The
values were obtained from at least three independent
experiments.
Evaluation of migration
Migration activity of HCASMCs with 100% confluency
was evaluated in the presence of 5% fetal bovine serum by migration
assay, as previously described (16,17).
At 0 and 24 h after the culture from day 2 and day 5, the culture
was scratched with a sterile sharp knife and the medium
(HuMedia-SB2) was replaced with fresh medium. Following 23 h
incubation at 37˚C, cells were pre-stained using
calcein-acetoxymethyl ester (calcein-AM) at 1 µg/ml for 1 h at 37˚C
and images were taken. Migrating cells were counted manually under
a light microscopy (TE300, Nikon Corporation) with x10
magnification and a fluorescent microscopy as mentioned above.
Statistical analysis
All data are presented as the mean ± SEM of at least
three independent experiments. Differences in mean values between
groups for multiple comparisons were subjected to one-way ANOVA and
Dunnett or Tukey's post hoc test and comparisons between two groups
were performed using Student's unpaired t test. Statistical
analysis was performed using JMP Pro 16 (SAS Institute Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Changes in protein expression levels
of α-SMA, calponin and PCNA in HCASMCs after reaching 100%
confluency
Fig. 1A shows
representative phase-contrast images of HCASMCs on days 1, 2 and 6.
HCASMCs were seeded at 90% confluency on day 0 (the day when
culture was started). The confluency was similar on days 0 (data
not shown) and 1. This may be due to the fact that HCASMCs did not
start proliferating immediately after being seeded. The cells
reached 100% confluency on day 2, which was maintained without cell
death until day 6; no cells came off from the bottom face of the
dish, which indicated there were no dying cells. Thus, cell
viability was not changed post-confluence. Representative protein
expression of α-SMA and calponin, typical markers of
contractile-type SMCs, and PCNA, a marker protein of proliferating
cells (12-14),
in HCASMCs before and after reaching 100% confluency is shown in
Fig. 1B. The levels of α-SMA
increased in a time-dependent manner and were significantly higher
on days 5 and 6 compared with day 2 (Fig. 1B and C). The calponin expression also increased
in a time-dependent manner and was significantly higher on days 4-6
than on day 2 (Fig. 1B and D). By contrast, PCNA expression decreased
in a time-dependent manner and was significantly lower on day 6
than on day 2 (Fig. 1B and E).
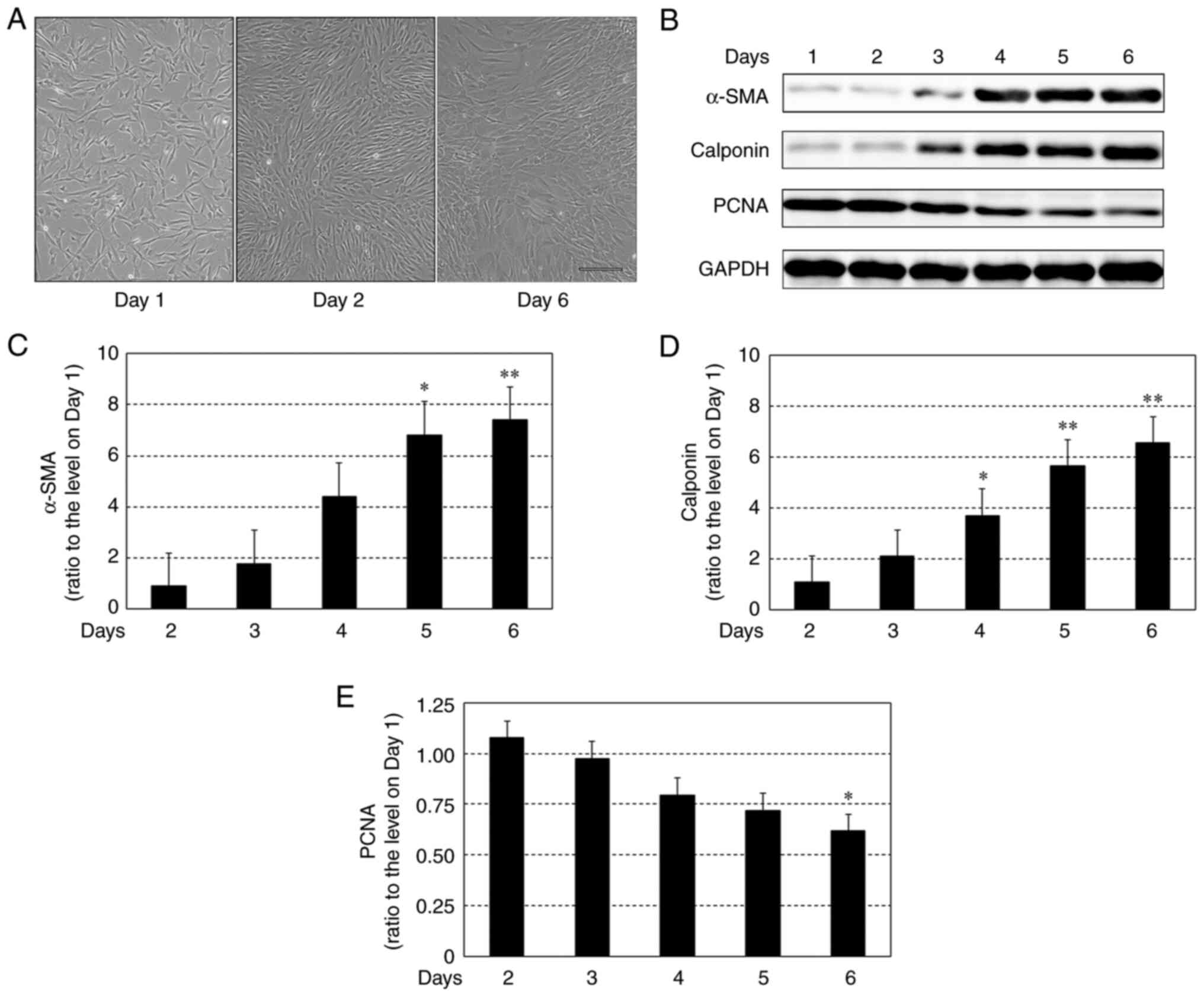 | Figure 1Time course of changes in protein
expression of α-SMA, calponin and PCNA in HCASMCs before and after
reaching 100% confluency. (A) Representative phase-contrast images
(x10 magnification) of HCASMCs on days 1, 2 and 6 after seeding at
90% confluency. Scale bar, 200 µm. (B) Representative western blots
for α-SMA, calponin, PCNA and GAPDH on each day. Densitometrical
analysis of expression of (C) α-SMA, (D) calponin and (E) PCNA.
Data are expressed as the ratio to GAPDH-corrected levels of each
protein on day 1 and are shown as the mean ± SEM of three
independent experiments. *P<0.05,
**P<0.01 vs. day 2. SMA, smooth muscle actin; PCNA,
proliferating cell nuclear antigen; HCASMC, human coronary artery
smooth muscle cell. |
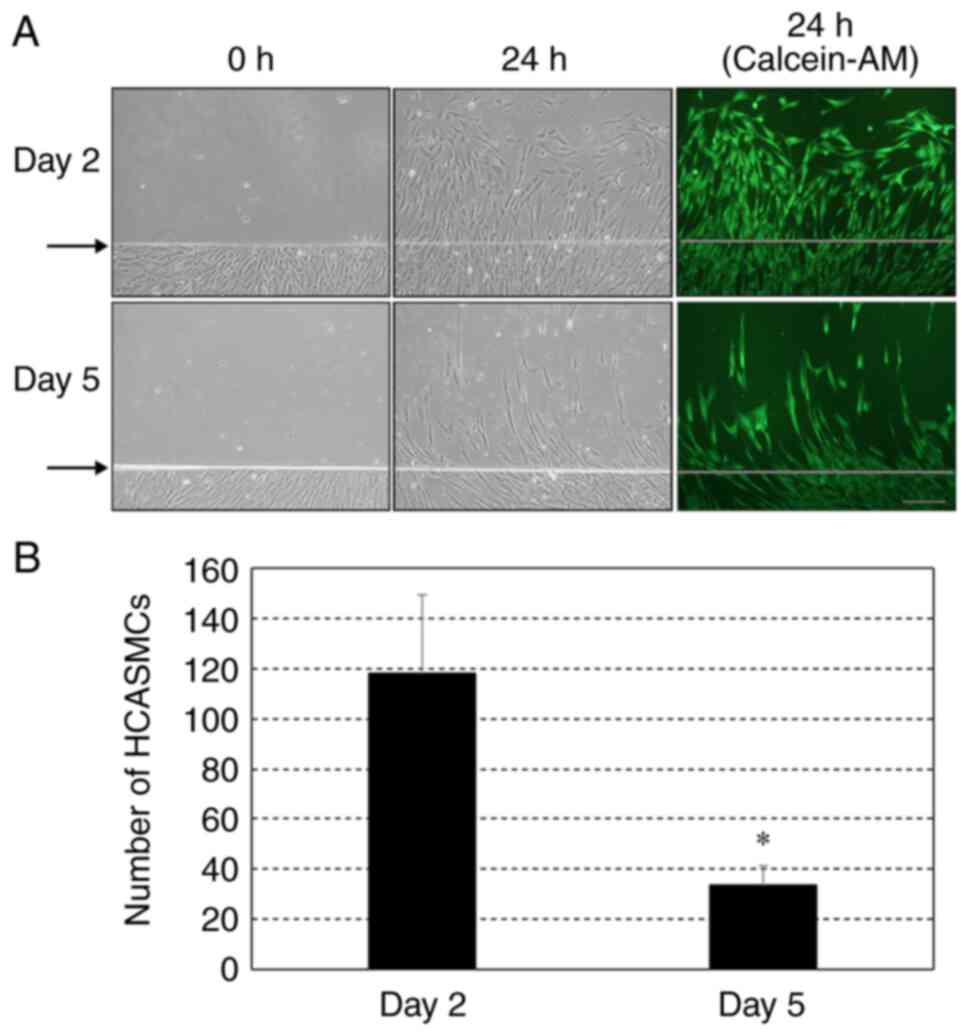
Migration activity of confluent and
post-confluent HCASMCs
Fig. 2A shows
representative phase-contrast and fluorescence images of migration
assay using HCASMCs. When culture from day 2 was scratched, cells
migrated from the baseline following 24 h incubation, while only a
small number of cells migrated at 24 h following injury when the
culture from day 5 was scratched. The number of migrating cells
from day 5 was significantly smaller than that from day 2 (Fig. 2B).
Effects of growth factors on protein
expression levels of α-SMA, calponin and PCNA in HCASMCs before
reaching 100% confluency
When culture of HCASMCs was started at 10%
confluency, the cells did not reach 100% confluency on day 4 in the
presence or absence of the growth factors (Fig. 3A). The number of cells was larger in
the presence of growth factors than in the absence but this was not
significant (Fig. 3B). Fig. 3C shows representative protein
expression of α-SMA, calponin and PCNA in cells cultured in the
presence or absence of growth factors. Expression levels of α-SMA
(Fig. 3D) and calponin (Fig. 3E) on day 4 were significantly higher
and those of PCNA (Fig. 3F) tended
to be lower (but not significantly) in the absence of the growth
factors.
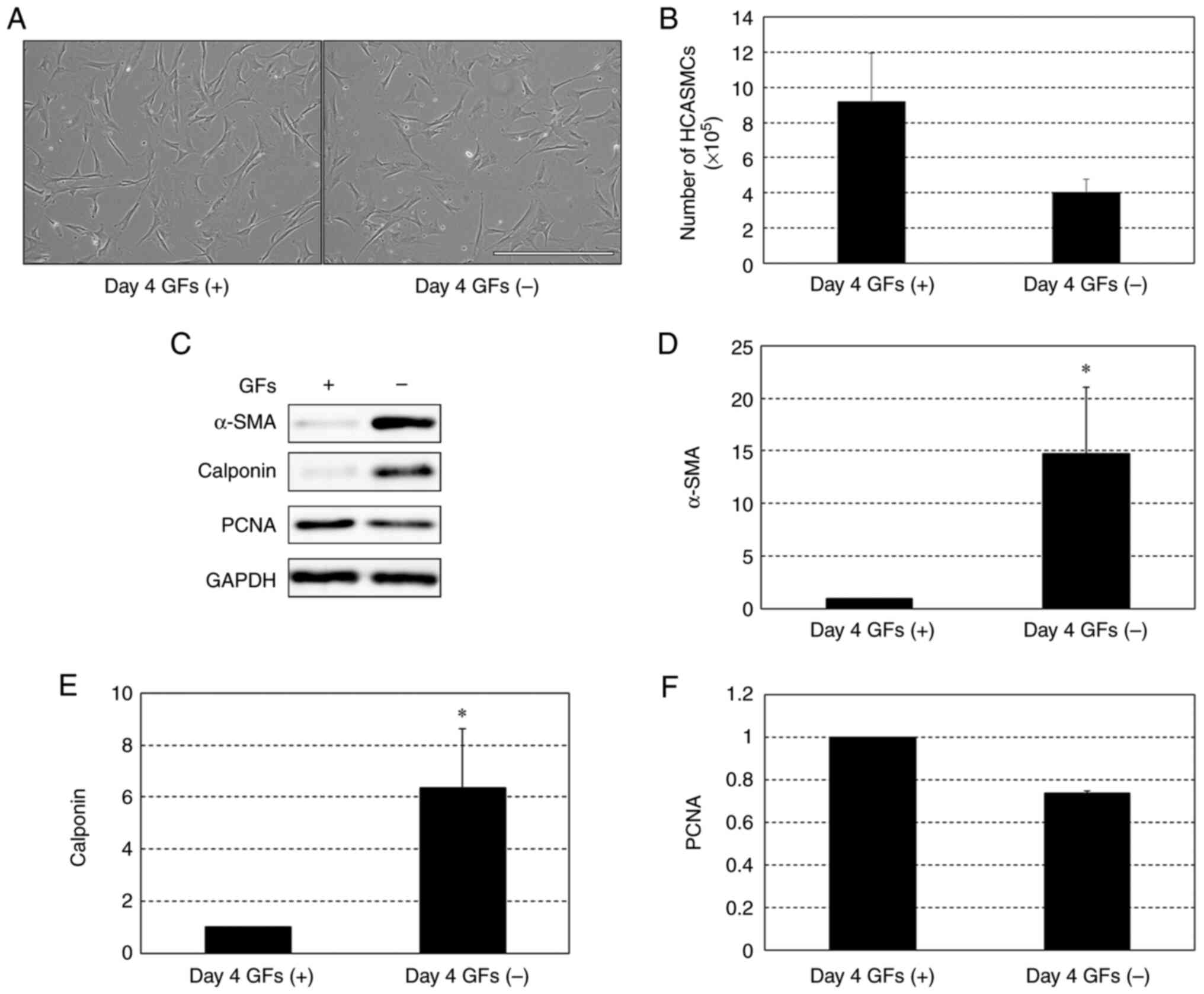 | Figure 3Protein expression levels of α-SMA,
calponin and PCNA in HCASMCs in the presence and absence of GFs,
including epidermal GF, fibroblast GF-B and insulin, on day 4 after
seeding at 10% confluency. (A) Representative phase-contrast images
(x10 magnification) on day 4 of HCASMCs cultured in the presence
and absence of GFs. Scale bar, 200 µm. (B) Numbers of HCASMCs on
day 4 in the presence and absence of the GFs. (C) Representative
western blots for α-SMA, calponin, PCNA and GAPDH expressed in
HCASMCs on day 4 in the presence and absence of GFs.
Densitometrical analysis of expression levels of (D) α-SMA, (E)
calponin and (F) PCNA. Data are expressed as the ratio to
GAPDH-corrected levels of each protein in the presence of GFs and
are shown as the mean ± SEM of three independent experiments.
*P<0.05 vs. GFs (+). SMA, smooth muscle actin; PCNA,
proliferating cell nuclear antigen; HCASMC, human coronary artery
smooth muscle cell; GF, growth factor. |
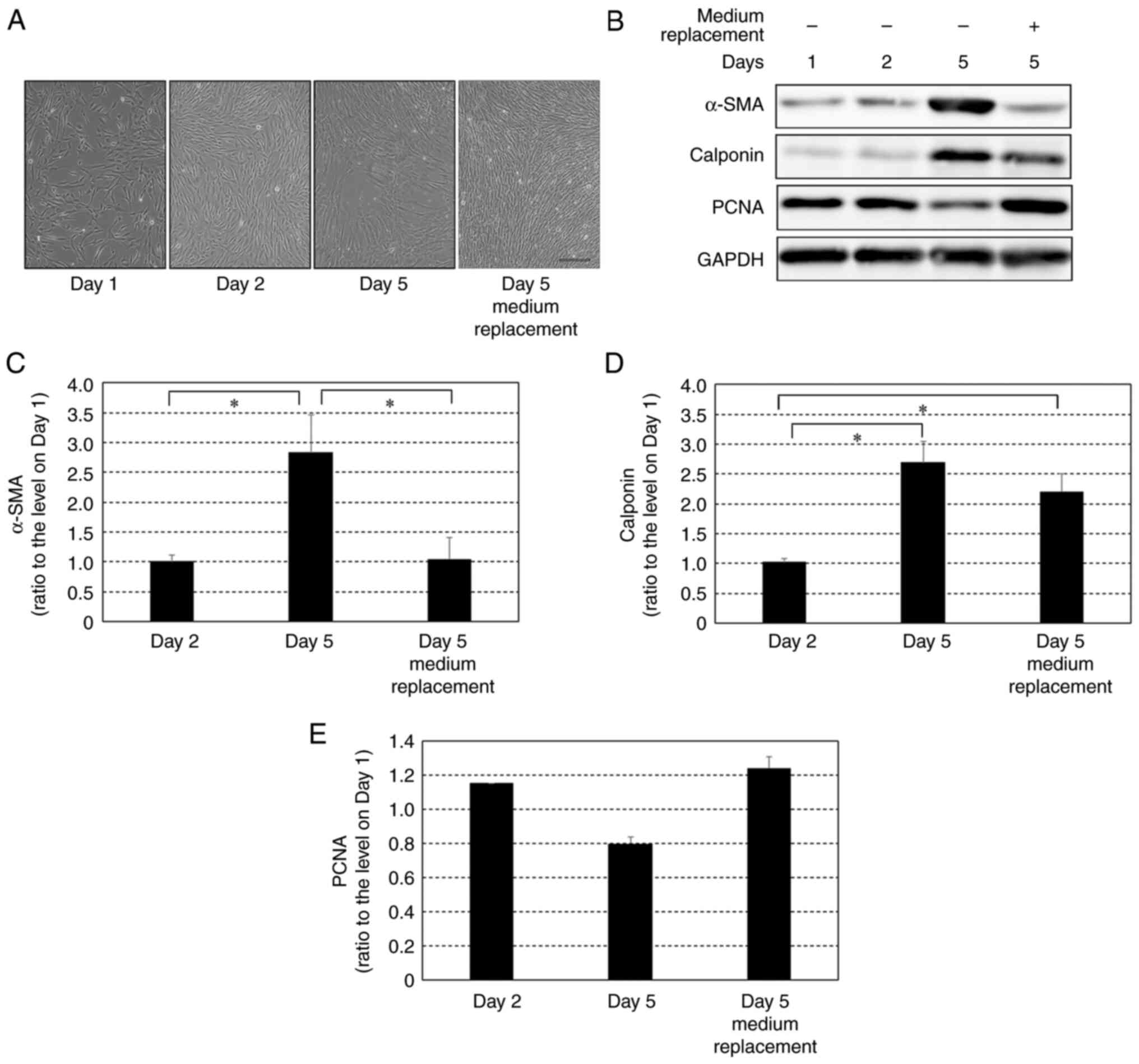
Effect of growth factors on protein
expression levels of α-SMA, calponin and PCNA in HCASMCs after
reaching 100% confluency
Cells seeded at 90% confluency were cultured and
maintained for five days with or without replacement of fresh
medium including growth factors every day (Fig. 4A). In the cells cultured in the
medium that was replaced every day with fresh medium, the protein
expression of α-SMA and PCNA on day 5 was similar to that on day 2
(Fig. 4B, C and E).
On the other hand, in the cells with daily replacement with fresh
medium, levels of calponin on day 5 were significantly higher than
those on day 2 (Fig. 4D). This
change in calponin expression levels was similar to the change
under a culture condition without daily medium replacement:
expression levels of calponin in the cells cultured without daily
medium replacement were significantly higher on day 5 than those on
day 2.
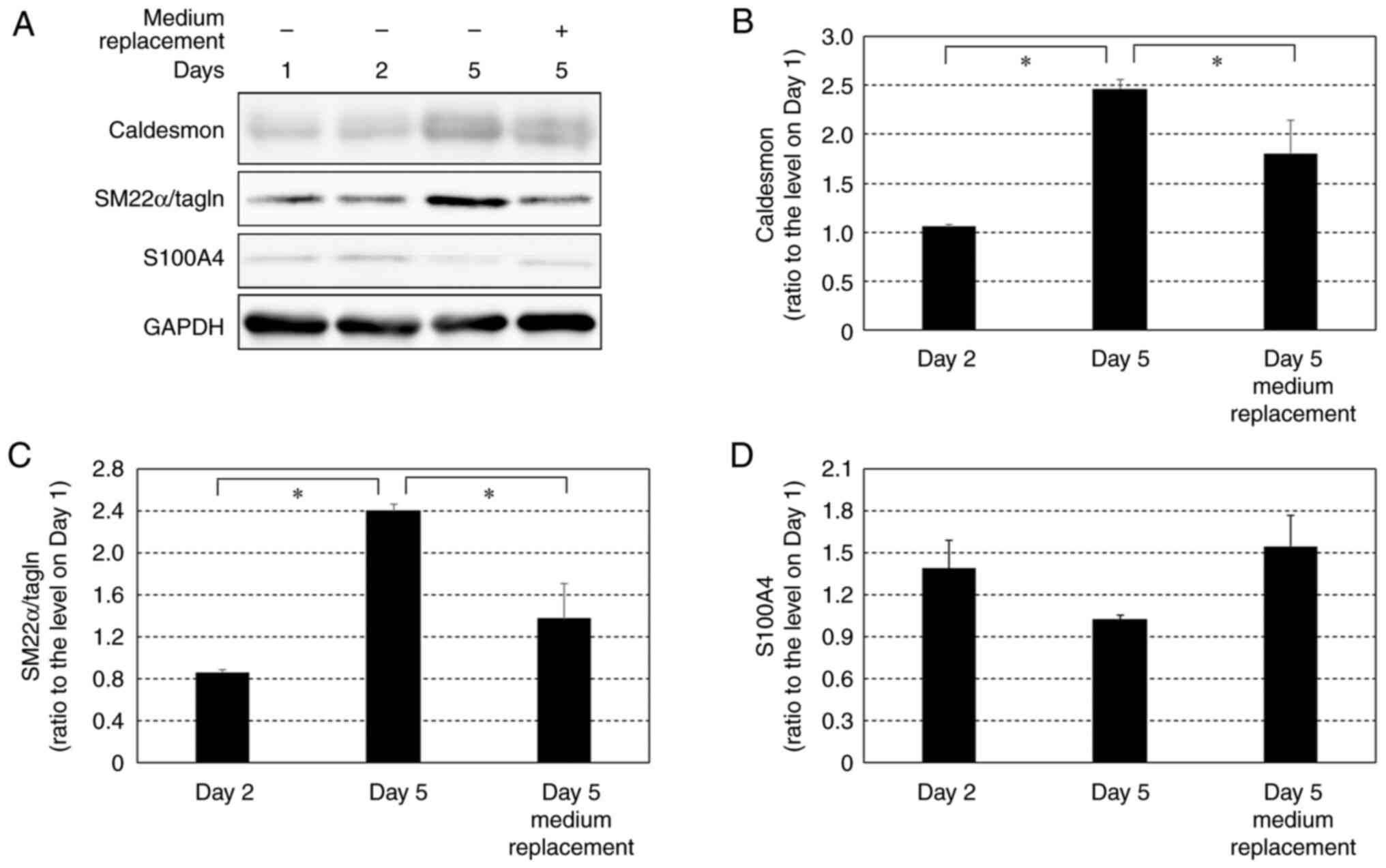
Effects of growth factors on protein
expression levels of caldesmon, SM22α/tagln and S100A4 in HCASMCs
after reaching 100% confluency
Protein expression levels of caldesmon, SM22 α/tagln
and S100A4 were compared on days 2 and 5 with and without daily
medium replacement (Fig. 5). The
expression levels of caldesmon and SM22α in cells without medium
replacement were significantly higher on day 5 than on day 2. The
levels of caldesmon and SM22α on day 5 in the cells with daily
medium replacement were significantly lower than in the cells
without medium replacement (Fig.
5A-C). The expression levels of S100A4 on day 5 in cells
without medium replacement tended to be lower than the expression
levels on day 2 but this was not significant. The expression levels
of S100A4 on day 5 in the cells with daily medium replacement
tended to be higher than in cells without medium replacement and
were comparable to those on day 2 (Fig.
5A and D); however, these
results were not significant.
Effects of growth factors on migration
of HCASMCs after reaching 100% confluency
Cell migration activity on day 5 without medium
replacement was significantly decreased compared with that on day
2. Cell migration on day 5 with daily medium replacement was
significantly higher than that without medium replacement and was
comparable to that on day 2 (Fig.
6A and B).
Discussion
In the present study, HCASMCs cultured post-100%
confluence without fresh medium replacement showed increased
expression levels of contractile phenotype markers and decreased
expression levels of proliferative phenotype markers. These results
suggested that redifferentiation was induced in post-confluent
HCASMCs. Similar changes in protein expression levels were observed
in HCASMCs cultured in medium without growth factors before
reaching 100% confluency. In the presence of a fresh medium
including growth factors, the aforementioned changes in expression
of phenotype marker proteins, except for calponin, were not
observed in post-confluent HCASMCs. Therefore, deprivation of
growth factors was required for induction of redifferentiation of
HCASMCs. This is consistent with the results of a previous study
showing that serum deprivation induces redifferentiation of
umbilical artery SMCs (6) since
serum contains growth factors. The present study investigated
changes in migration, which is a representative functional
characteristic of proliferative phenotype vascular SMCs, of
HCASMCs. Post-confluent HCASMCs showed lower migration activity
than that cells at 100% confluency; this decreased migration
activity was not found in the post-confluent cells when medium was
exchanged with a fresh medium every day. Accordingly, the results
of both protein expression and migration activity indicated that
deprivation of growth factors induces redifferentiation of HCASMCs.
Similar changes in expression of contractile phenotype markes such
as α-SMA and SM22α and migration activity were shown in RASMCs in
which redifferentiation was induced by serum deprivation (8). To the best of our knowledge, the
present study is the first to determine experimental conditions for
inducing redifferentiation of HCASMCs in vitro.
The present study demonstrated that deprivation of
growth factors, including EGF, FGF-B and insulin, induced
redifferentiation of HCASMCs. Redifferentiation of SMCs has been
demonstrated in post-PTCA neointimal tissues in a human specimen
(11). Blockade of signals
stimulated by growth factors may facilitate redifferentiation of
SMCs at loci of PTCA and in atherosclerotic plaque (11). This indicates potential therapeutic
application of gene suppression of growth factors and their
receptors in HCASMCs at atherosclerotic lesions.
A novel finding of this study was the difference in
marker proteins for differentiation and redifferentiation. Calponin
and α-SMA are protein markers of differentiated (contractile
phenotype) SMCs (12-14).
However, the present study obtained different results regarding
calponin and α-SMA in cells with and without daily replacement with
a fresh medium: Compared with expression level of calponin in
confluent HCASMCs, the calponin expression was increased in
post-confluent dedifferentiated HCASMCs in the presence of fresh
medium, although calponin is a phenotype marker of differentiated
arterial SMCs. On the other hand, α-SMA expression level was not
different in confluent and post-confluent dedifferentiated HCASMCs
in the presence of growth factors. In addition, the present study
investigated expression levels of caldesmon and SM22α, other
protein markers of differentiated-phenotype SMCs. Similar to the
results of α-SMA, levels of caldesmon and SM22α in post-confluent
cells with daily medium replacement were not significantly
different from those in confluent cells. These results suggested
that α-SMA, caldesmon and SM22α, but not calponin, may serve as
markers of redifferentiation of HCASMCs. Thus, medium replacement
maintained HCASMC dedifferentiation after 100% confluency, while
expression of calponin, a marker of differentiated arterial smooth
muscle cells, was increased in post-confluent cells in the presence
of growth factors. This means that calponin expression is more
prone to be upregulated in post-confluent HCASMCs and is less
affected by stimulation with growth factors compared with other
contractile-phenotype markers such as α-SMA, caldesmon and SM22α.
However, the reason for these findings remains unknown and future
studies are needed. Levels of calponin, α-SMA, caldesmon and SM22α
were all significantly higher in post-confluent cells without
medium replacement than in confluent cells. Therefore, one
potential explanation for the aforementioned difference in results
is a difference in signal transduction of growth factor-induced
inhibition of gene transcription of calponin and other
contractile-phenotype marker proteins. Myocardin, a transcriptional
coactivator of serum response factor, serves key roles in
differentiation of SMCs (18,19)
and expression levels of α-SMA, SM22 and calponin have been shown
to be similarly affected by modulating myocardin activity in human
aortic SMCs (20). Therefore, it is
a possibility that a myocardin-independent pathway is involved in
the expression of contractile-phenotype proteins in
redifferentiated HCASMCs. Expression levels of α-SMA in
post-confluent HCASMCs with daily medium replacement were
comparable to those in the confluent cells, while expression levels
of caldesmon and SM22α were slightly but not significantly higher
in post-confluent cells with daily medium replacement than in
confluent cells. Thus, α-SMA may be a more sensitive marker of
growth factor-dependent redifferentiation of HCASMCs than caldesmon
and SM22α. Although it would be ideal to confirm α-actin as a
marker for redifferentiation of HCASMCs functionally by using RNA
interference-α-SMA in vitro, the present study did not
perform such an experiment due to hypothesized damage to cells by
loss of α-SMA, a fundamental cytoskeletal element (21).
In conclusion, deprivation of growth factors,
including EGF, FGF-B and insulin, from culture medium induced
redifferentiation of HCASMCs. α-SMA, caldesmon and SM22α, but not
calponin, may serve as markers for redifferentiation of
HCASMCs.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
RS, RE and IW designed the study. RS performed the
experiments and data analysis. RE and IW contributed to data
interpretation. RS, RE and IW confirm the authenticity of all the
raw data. IW drafted the manuscript. RS and RE reviewed and edited
the manuscript. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gomez D and Owens GK: Smooth muscle cell
phenotypic switching in atherosclerosis. Cardiovasc Res.
95:156–164. 2012.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Bennett MR, Sinha S and Owens GK: Vascular
smooth muscle cells in atherosclerosis. Circ Res. 118:692–702.
2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Owens GK, Kumar MS and Wamhoff BR:
Molecular regulation of vascular smooth muscle cell differentiation
in development and disease. Physiol Rev. 84:767–801.
2004.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Thyberg J: Differentiated properties and
proliferation of arterial smooth muscle cells in culture. Int Rev
Cytol. 169:183–265. 1996.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Li S, Sims S, Jiao Y, Chow LH and
Pickering JG: Evidence from a novel human cell clone that adult
vascular smooth muscle cells can convert reversibly between
noncontractile and contractile phenotypes. Circ Res. 85:338–348.
1999.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Han M, Wen JK, Zheng B, Cheng Y and Zhang
C: Serum deprivation results in redifferentiation of human
umbilical vascular smooth muscle cells. Am J Physiol Cell Physiol.
291:C50–C58. 2006.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Vaes RDW, van den Berk L, Boonen B, van
Dijk DPJ, Olde Damink SWM and Rensen SS: A novel human cell culture
model to study visceral smooth muscle phenotypic modulation in
health and disease. Am J Physiol Cell Physiol. 315:C598–C607.
2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Jiang GJ, Han M, Zheng B and Wen JK:
Hyperplasia suppressor gene associates with smooth muscle
alpha-actin and is involved in the redifferentiation of vascular
smooth muscle cells. Heart Vessels. 21:315–320. 2006.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Doi K, Ikeda T, Itoh H, Ueyama K, Hosoda
K, Ogawa Y, Yamashita J, Chun TH, Inoue M, Masatsugu K, et al:
C-type natriuretic peptide induces redifferentiation of vascular
smooth muscle cells with accelerated reendothelialization.
Arterioscler Thromb Vasc Biol. 21:930–936. 2001.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Liu Z, Zhang M, Zhou T, Shen Q and Qin X:
Exendin-4 promotes the vascular smooth muscle cell
re-differentiation through AMPK/SIRT1/FOXO3a signaling pathways.
Atherosclerosis. 276:58–66. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Aikawa M, Sakomura Y, Ueda M, Kimura K,
Manabe I, Ishiwata S, Komiyama N, Yamaguchi H, Yazaki Y and Nagai
R: Redifferentiation of smooth muscle cells after coronary
angioplasty determined via myosin heavy chain expression.
Circulation. 96:82–90. 1997.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Samaha FF, Ip HS, Morrisey EE, Seltzer J,
Tang Z, Solway J and Parmacek MS: Developmental pattern of
expression and genomic organization of the calponin-h1 gene. A
contractile smooth muscle cell marker. J Biol Chem. 271:395–403.
1996.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Gao H, Steffen MC and Ramos KS:
Osteopontin regulates α-smooth muscle actin and calponin in
vascular smooth muscle cells. Cell Biol Int. 36:155–161.
2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lee M, San Martín A, Valdivia A,
Martin-Garrido A and Griendling KK: Redox-sensitive regulation of
myocardin-related transcription factor (MRTF-A) phosphorylation via
palladin in vascular smooth muscle cell differentiation marker gene
expression. PLoS One. 11(e0153199)2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Eguchi R and Wakabayashi I: HDGF enhances
VEGF-dependent angiogenesis and FGF-2 is a VEGF-independent
angiogenic factor in non-small cell lung cancer. Oncol Rep.
44:14–28. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Aljohi A, Matou-Nasri S, Liu D, Al-Khafaji
N, Slevin M and Ahmed N: Momordica charantia extracts protect
against inhibition of endothelial angiogenesis by advanced
glycation endproducts in vitro. Food Funct. 9:5728–5739.
2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Takaya K, Aramaki-Hattori N, Sakai S,
Okabe K, Asou T and Kishi K: Decorin inhibits dermal mesenchymal
cell migration and induces scar formation. Plast Reconstr Surg Glob
Open. 10(e4245)2022.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wang Z, Wang DZ, Pipes GC and Olson EN:
Myocardin is a master regulator of smooth muscle gene expression.
Proc Natl Acad Sci USA. 100:7129–7134. 2003.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zheng XL: Myocardin and smooth muscle
differentiation. Arch Biochem Biophys. 543:48–56. 2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhou YX, Shi Z, Singh P, Yin H, Yu YN, Li
L, Walsh MP, Gui Y and Zheng XL: Potential role of glycogen
synthase kinase-3β in regulation of myocardin activity in human
vascular smooth muscle cells. J Cell Physiol. 231:393–402.
2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Gerthoffer WT: Actin cytoskeletal dynamics
in smooth muscle contraction. Can J Physiol Pharmacol. 83:851–856.
2005.PubMed/NCBI View
Article : Google Scholar
|