Introduction
Carotid atherosclerotic plaque (CAP) is a
predominant established risk factor for stroke, which is associated
with high morbidity and mortality (1). Although the development of carotid
plaques is influenced by environmental factors, such as elevated
blood pressure and serum lipoprotein abnormalities (2), studies have been increasingly
focusing on the genetic causes associated with the formation of
atherosclerosis. ATP-binding cassette transporter 1 (ABCA1) has
been recognized as a major mediator in the initial step of the
reverse cholesterol transport, and was shown to facilitate the
efflux of cholesterol and phospholipids to lipid-poor
apolipoproteins (apo)A-I and apoE from peripheral cells, resulting
in the formation of nascent or pre-high-density lipoprotein (HDL)
(3,4). Such a cholesterol efflux process
prevents the accumulation of cholesterol or oxidative low-density
lipoprotein (LDL) in peripheral cells, including macrophages within
the arterial wall and, therefore, reduces the accumulation of
atherosclerotic plaque.
ABCA1 deficiency was indentified to be responsible
for Tangier disease and familial HDL deficiency. These conditions
are characterized by low levels of HDL, excessive deposition of
cholesteryl esters in tissue macrophages and an increased risk for
atherosclerotic disease (5). The
underlying mechanism is thought to result from the failure of ABCA1
to transport cholesterol and phospholipids out of cells to form
complexes with apolipoproteins to generate HDL (6). The intracellular sterol accumulation
and subsequent development of foam cells may, as a result, lead to
the atheromatous plaque. Due to its important role in
atherosclerosis, increasing attention has been focused on ABCA1, so
that therapeutic interventions are introduced to regulate its
expression and therefore reduce the development of
atherosclerosis.
The expression and significance of ABCA1 have been
well reported in animals with ABCA1 overexpression or inactivation
(7), as well as in individuals
with ABCA1 gene deficiency due to gene mutations (8). Although the systemic effects of loss
of function have been well investigated in patients with ABCA1
deficiency, few studies have addressed in detail the possible
effect of the transporter on plaque complication in localized
atherosclerosis in the absence of familial gene mutations.
Identification of the ABCA1 expression in localized lesions may
provide new insights into local genetic determinants of
atheromatous disease without mutations in the gene and therefore
the development of new therapeutic strategies.
In this study, we assessed the expression of the
ABCA1 gene and protein in human CAPs and compared it to human
arteries obtained from normal controls. Also, we classified the
plaques into three grades according to the American Heart
Association (AHA) and characterized the ABCA1 expression in the
CAPs of different grades. Furthermore, to gain further insight into
the regulatory mechanism underlying ABCA1 expression, we determined
the expression of one of its key regulators, retinoid X receptor α
(RXRα).
Materials and methods
Patients and specimens
We collected CAPs from 48 patients undergoing
carotid endarterectomy for symptomatic (transient ischemic attacks,
minor strokes) or asymptomatic stenosis >70% confirmed by MR
angiography or conventional arteriography. All patients underwent
detailed history assessment and a physical examination at baseline
that included routine blood biochemistry tests,
electrocardiography, chest radiography, ultrasound cardiography,
transcranial Doppler sonography, carotid CEMRI, computed
tomography, MRI and magnetic resonance angiography of the brain. In
addition, the presence of vascular risk factors as well as previous
antihypertensive, statin and antiplatelet treatments were recorded
(Table I). We also collected 10
inferior mesenteric arteries dissected from colectomy specimens of
subjects having elective operation as normal controls. The control
patients were assessed for the absence of symptomatic
atherosclerotic disease by history and examination.
 | Table I.Clinical characteristics of the
patients and controls. |
Table I.
Clinical characteristics of the
patients and controls.
| Patients | Controls |
|---|
| No. | 48 | 10 |
| Age (years) | 56.2±7.3 | 55.9±10.1 |
| Gender
(male/female) | 34/14 | 7/3 |
| Smokers | 26 | 4 |
| Diabetes | 12 | 0 |
| Hypertension | 30 | 3 |
| Coronary artery
disease | 14 | 0 |
| Family history of
IHD | 22 | 2 |
| Treatment | | |
| Aspirin | 20 | 1 |
| Clopidogrel | 2 | 0 |
| Warfarin | 4 | 0 |
| Statins | 14 | 0 |
The endarterectomy specimens included the
atheromatous plaque, the adjacent intima and the medial layers. The
collected 48 specimens were assigned to three grades according to
the modified AHA classification (9): 14 cases of grade I (type I and II),
which was defined as a diffuse intimal thickening or small
eccentric plaque without calcification; 22 cases of grade II (type
III and IV), which was described as plaque with a lipid or necrotic
core surrounded by fibrous tissue with possible calcification; 12
cases of grade III (type V and VI), presenting complex plaques with
a possible surface defect, hemorrhage or thrombus. The control
arteries were of full-thickness and consisted of the adventitial
layer. Plaques and control arteries were frozen immediately in
liquid nitrogen and stored at −80°C until further processing.
Sections from control arteries were stained with H&E and
assessed by a histopathologist for evidence of age-related
atheromatous change.
The study was approved by the Medical Ethics
Committee of the Fifth Affiliated Hospital of Zhengzhou University,
and informed consent was obtained from each patient.
Reverse transcriptase-polymerase chain
reaction (RT-PCR)
The ABCA1 and RXRα mRNA expression levels were
determined using semi-quantitative RT-PCR. Total RNA was isolated
from freshly isolated pulverized frozen tissue (∼25 mg) with TRIzol
reagent (Invitrogen) and solubilized in RNAase-free water. The
first-strand cDNA was synthesized from 2 μg of total RNA with
PrimeScript Reverse Transcriptase (Takara Bio). The sequences of
the specific primers used are shown in Table II. Specific primers for
glyceraldehyde-3-phosphate dehydrogenase (GAPDH; Clontech) were
added as a control. PCR was carried out by incubating each cDNA
sample with primers (0.5 μM each), Blend Taq polymerase
(1.25 units; Takara Bio) and a deoxynucleotide. For ABCA1, PCR was
run for 35 cycles with 30 sec/94°C denaturation, 40 sec/52°C
annealing and 45 sec/72°C elongation. For RXRα, PCR was run for 38
cycles with 30 sec/94°C denaturation, 45 sec/58°C annealing and 40
sec/72°C elongation. Reactions were performed in triplicate and in
a total volume of 25 μl. PCR products were electrophoresed on 1%
agarose gels. Bands were stained with ethidium bromide (Sigma) and
detected with a LAS-3000 mini CCD camera (Fuji Film). The Eagle Eye
II analyzer (USA) was used to determine the optical density (OD) of
target and reference mRNA. The mRNA levels were recorded as the
relative value of the OD of the target gene to that of GAPDH.
| Table II.Sequences of the primers for PCR. |
Table II.
Sequences of the primers for PCR.
| Forward primer
sequence | Reverse primer
sequence | Product fragment
(bp) |
|---|
| ABCA1 |
5′-GGGAGGCTCCCGGAGTT-3′ |
5′-GTATAAAAGAAGCCTCCGAGCATC-3′ | 302 |
| RXRα |
5′-GTATAAAAGAAGCCTCCGAGCATC-3′ |
5′-GCGGCGCCTCCAGCATCTCCATA-3′ | 800 |
| GAPDH |
5′-TGATGACATCAAGAAGGTGGTGAA-3′ |
5′-TCCTTGGAGGCCATGTAGGCCAT-3′ | 238 |
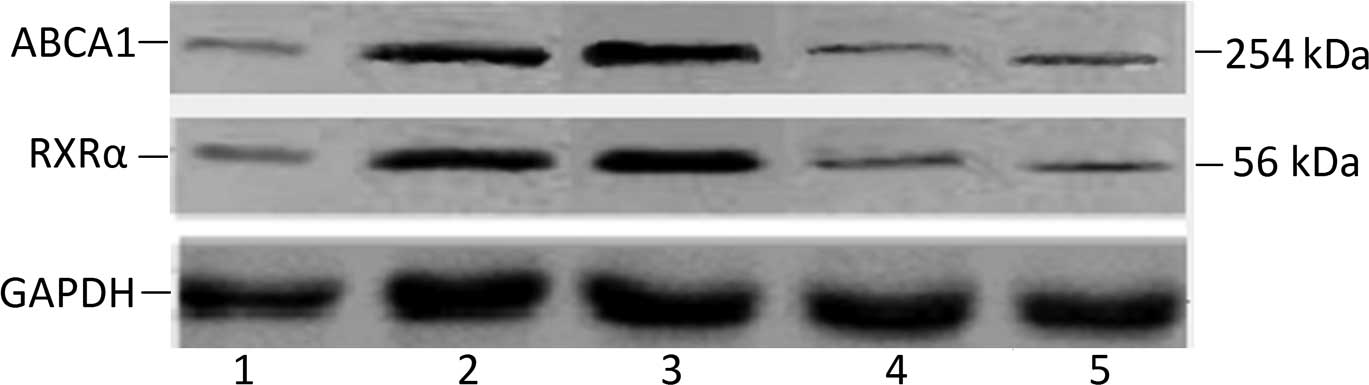
Western blotting
The content of ABCA1 and RARα protein was detected
by western blot analysis. The frozen tissues were pulverized and
sonicated in RIPA lysis buffer (Santa Cruz). After centrifugation
at 14,000 x g for 30 min at 4°C, 30 μg of the supernatants was
separated on 10% SDS-polyacrylamide gel and transferred onto
Immobilon-P transfer membrane (Millipore). After being blocked with
5% skim milk, the membrane was incubated with the primary ABCA1
antibody (dilution at 1:1,000; Abcam) or the primary RARα antibody
(dilution at 1:1,000; Millipore) at 4°C overnight. Membranes were
then treated with anti-rabbit secondary antibody conjugated with
horseradish peroxidase (dilution at 1:1,000; Santa Cruz). Complexes
on the membranes were then visualized using the Super Signal West
Dura Extended Duration Substrate (Pierce Biotechnology Inc.),
photographed and analyzed with the Eagle Eye II analyzer. The
relative value of the absorbance value of the target protein to the
reference protein was recorded.
Statistical analysis
Results are presented as the means ± SEM.
Statistical analysis was performed with SAS 9.0. Gender differences
between patient and control groups were assessed using the
χ2 test. Comparisons in the ABCA1 and RXRα expression
between patients and controls were performed using the Student’s
t-test or ANOVA. Relations between the ABCA1 and RXRα mRNA were
evaluated using Pearson correlations. Differences were considered
to denote statistical significance at P<0.05.
Results
Patient demographics
The demographic characteristics of the patients and
controls are presented in Table I.
There was no significant difference between patients and controls
in terms of gender, symptomatic carotid disease, individual or
combined risk factors, treatment or coexistence of other vascular
disease (Table I).
Expression of ABCA1 mRNA in carotid
plaques
Table III
summarizes the results for ABCA1 and RXRα mRNA levels in CAPs and
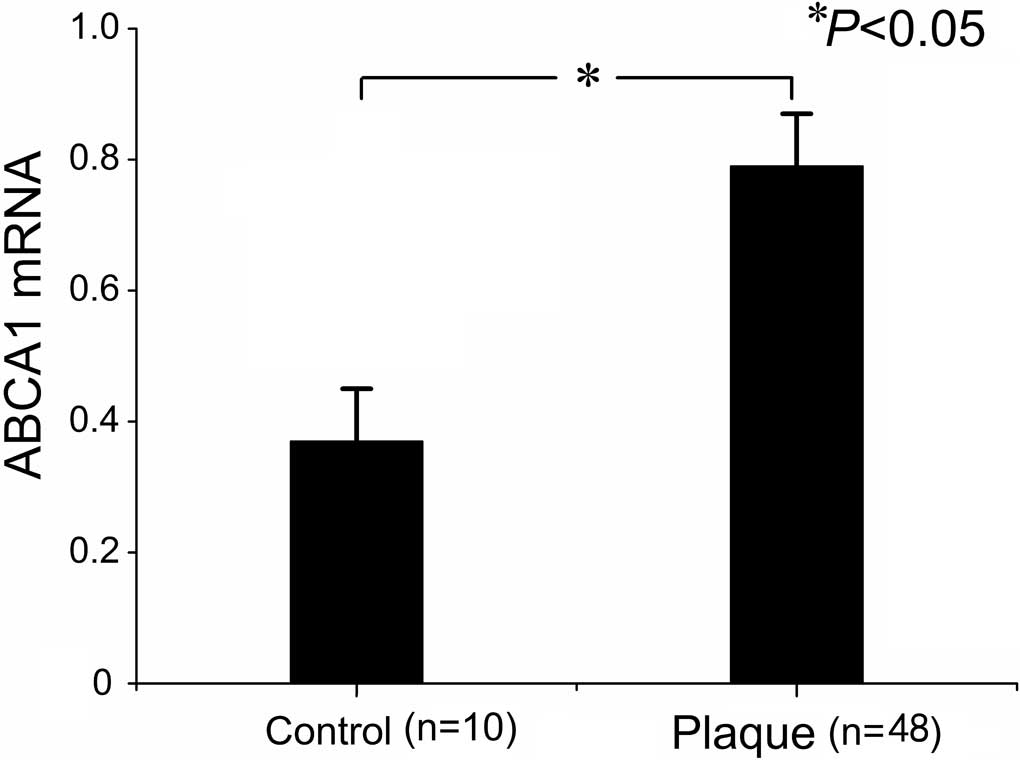
control arteries. As expected, ABCA1 mRNA was significantly
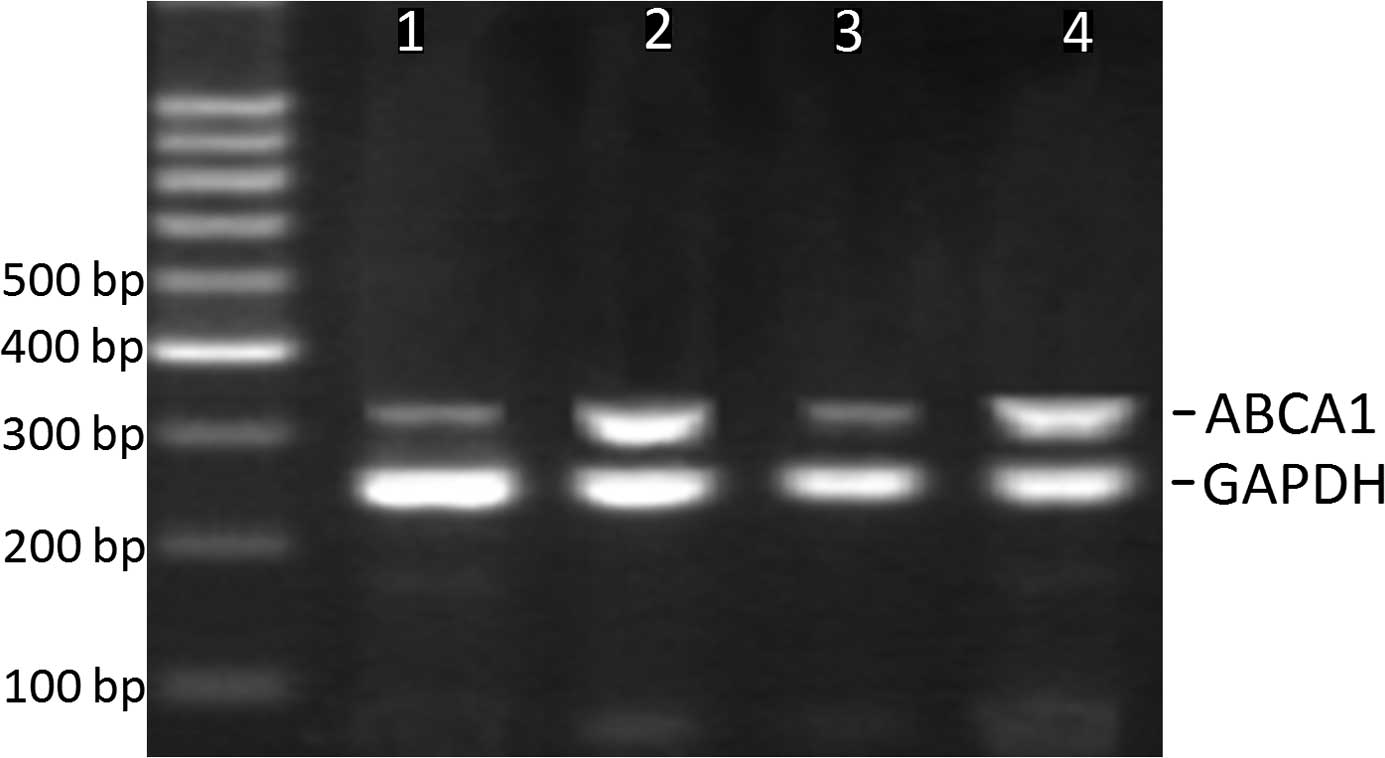
increased in the plaques compared to controls (Fig. 1). When we compared ABCA1 mRNA
expression across plaques of different grades, we found higher
ABCA1 mRNA levels in plaques of grade II and III compared to those
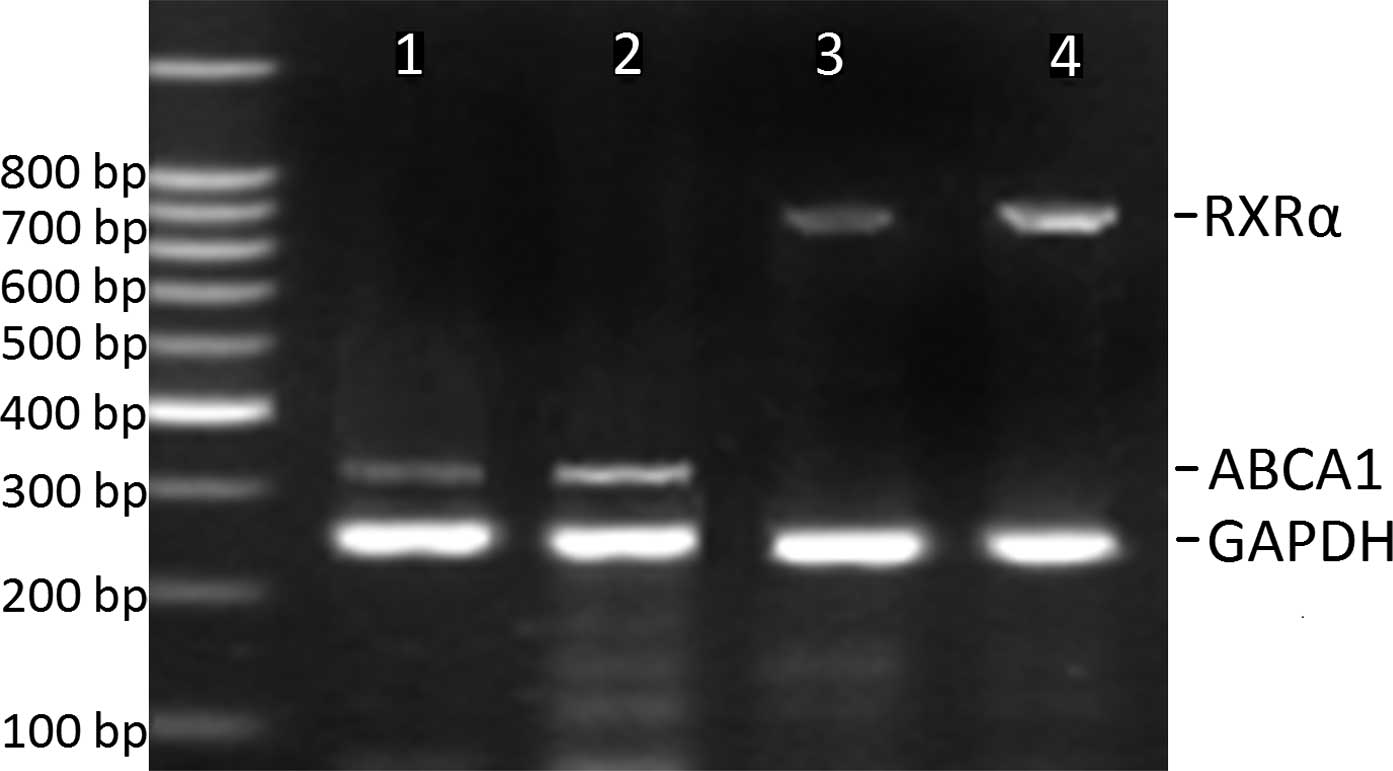
of grade I (Table III, Figs. 2 and 3). However, ABCA1 mRNA in plaques of
grade III was not significantly higher than that of grade II
(Table III). To explore the
potential underlying mechanism, the RXRα mRNA levels were also
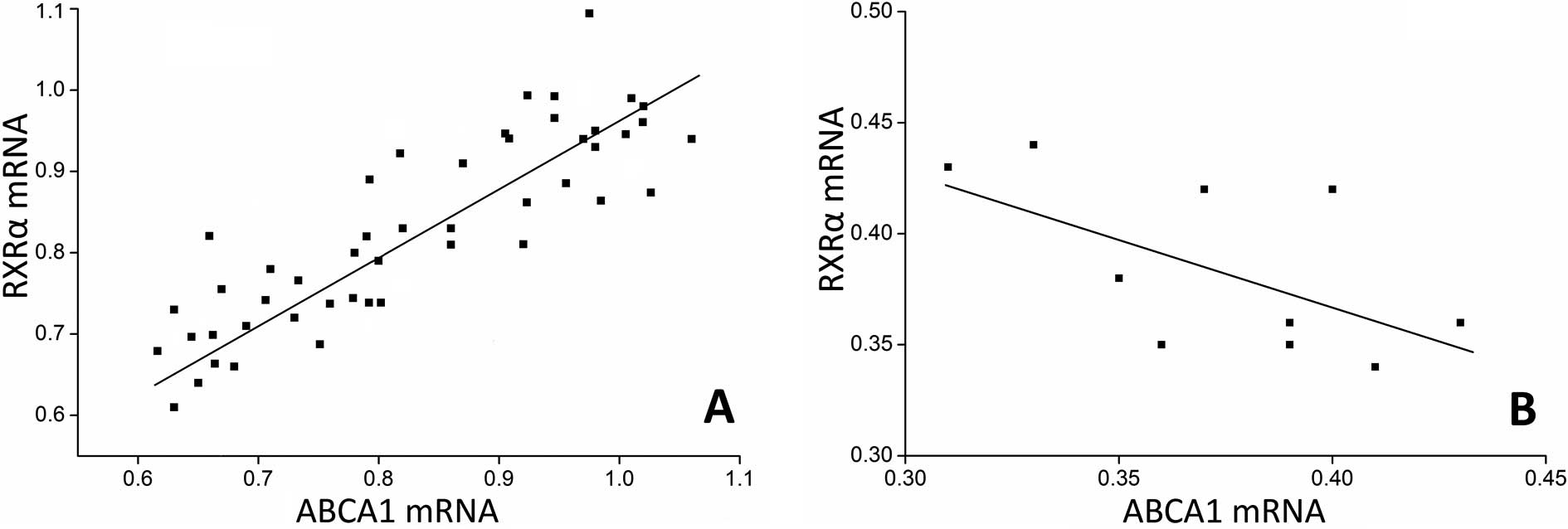
determined and correlated to the ABCA1 mRNA expression. As a
result, we found a parallel evolving pattern between RXRα and ABCA1
mRNA expression. Furthermore, a significant association between
ABCA1 and RXRα mRNA expression levels was observed in the plaques
(Fig. 4A), but no correlation was
found in the control arteries (Fig.
4B), suggesting the coordinative actions between ABCA1 and RXRα
mRNA in the plaques.
| Table III.Expression of ABCA1 and RXRα mRNA in
plaques and control arteries. |
Table III.
Expression of ABCA1 and RXRα mRNA in
plaques and control arteries.
| ABCA1 mRNA | RXRα mRNA |
|---|
| Plaques | | |
| Grade I (n=14) | 0.67±0.05a | 0.67±0.03a |
| Grade II
(n=22) |
0.97±0.05a,b |
0.95±0.05a,b |
| Grade III
(n=12) | 0.77±0.04a | 0.74±0.04a |
| Control arteries
(n=10) | 0.37±0.04 | 0.39±0.04 |
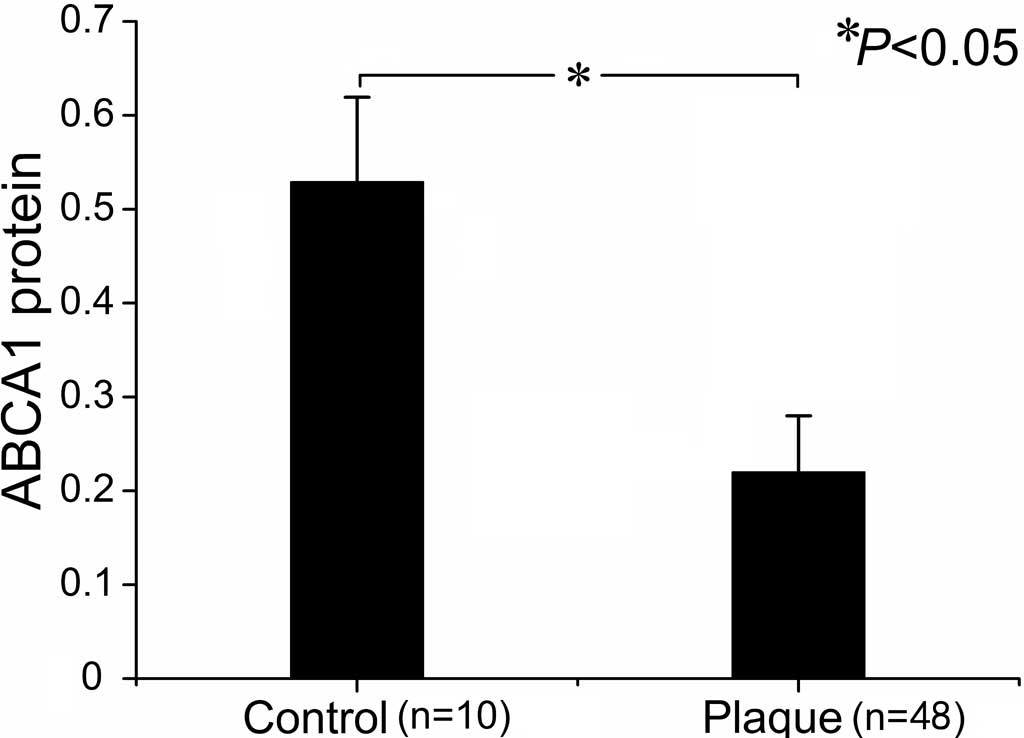
Expression of ABCA1 protein in carotid
plaques
Table IV summarizes
the results for ABCA1 and RXRα protein levels in CAPs and control
arteries. In contrast to mRNA levels, ABCA1 protein was
significantly reduced in the plaques as compared to control
arteries (Figs. 5 and 6, Table
IV). The plaques of grade II and III exhibited lower ABCA1
protein levels than those of grade I. However, no difference was
found in ABCA1 protein levels in plaques of grade II and III
(Table IV). A similar evolving
pattern was observed in the RXRα protein expression. In the
plaques, the RXRα protein levels were significantly reduced as
compared to control arteries. Although the RXRα protein levels were
significantly lower in plaques of grade II and III compared to
grade I, there was no difference between plaques of grade II and
III (Table IV).
| Table IV.Expression of ABCA1 and RXRα protein
in plaques and control arteries. |
Table IV.
Expression of ABCA1 and RXRα protein
in plaques and control arteries.
| ABCA1 protein | RXRα protein |
|---|
| Plaques | | |
| Grade I
(n=14) | 0.26±0.04a | 0.31±0.05a |
| Grade II
(n=22) |
0.16±0.05a,b |
0.18±0.03a,b |
| Grade III
(n=12) | 0.20±0.04a | 0.27±0.04a |
| Control arteries
(n=10) | 0.53±0.03 | 0.57±0.04 |
Discussion
In this study, we showed that in CAPs, both ABCA1
and RXRα mRNA levels were significantly increased compared to
control arteries, and a parallel correlation between the mRNA
levels of both genes was found. In these atherosclerotic tissues,
ABCA1 and RXRα mRNA levels in both grade II and III were notably
higher than those in grade I, but no significant difference was
found in mRNA levels between grade II and III. Despite the
up-regulation of mRNA expression, ABCA1 and RXRα protein levels
were notably decreased in atherosclerotic plaques. Furthermore,
although the plaques of grade I exhibited lower ABCA1 and RXRα
protein levels, there was no difference in the levels of both
proteins between plaques of grade II and III. These findings seem
to be important for improving our understanding of the
physiological roles of ABCA1 in localized atherosclerosis without
gene mutations, and thus raise the possibility of further new
therapeutic interventions targeted at ABC transporters in
atherosclerosis.
ABCA1 is a membrane transporter abundant in
macrophages that utilize ATP as a source of energy to transport
various molecules across membranes; its origin and pathological
significance in atherosclerotic tissues was identified in animal
studies of ABCA1 inactivation and human gene ABCA1 mutations
(10). In these studies, loss of
ABCA1 gene function was shown to play a key role in lipid
deposition and its sequelae (7,8). In
the present study, however, we found a significant increase in
ABCA1 gene expression in atherosclerotic tissues. In fact, mRNA
levels did not precisely reflect protein expression and,
particularly for ABCA1, there was a significant discordance between
ABCA1 mRNA and protein levels, implying that post-transcriptional
regulation plays a major role in ABCA1 protein expression (11). Since we determined the mRNA and
protein levels by analyzing the same specimen, we speculated that,
in the control arteries, the presence of ABCA1 protein reflected
the expression of the gene. In the CAP specimens, however, markedly
lower levels of ABCA1 protein were found compared to control
tissues. These findings suggested that, although the ABCA1
transcription was significantly increased in atherosclerotic
tissues, its protein expression was not increased, but reduced, in
agreement with results published by Albrecht et al (12).
We showed that the plaques of grade II and III
exhibited higher ABCA1 mRNA and lower protein levels than those of
grade I. In terms of plaque composition, plaques of grade II and
III consist of a lipid or necrotic core, which makes them unstable
(13). Thus, ABCA1 may serve as an
important marker for plaque instability. However, we found no
significant difference in both mRNA and protein levels between
plaques of grade II and III, suggesting that the unstable plaques
may share a similar pattern in ABCA1 distribution. Taking into
account that ABCA1 is mainly located in macrophages within the
atheromatous lesion (14), the
different ABCA1 expression patterns in plaques of different grades
may be associated with the composition and microenvironment of the
atherosclerotic lesions.
Atherosclerotic plaque is a dynamic structure
composed of lipids, cells and extracellular matrix (15). In unstable atherosclerotic lesions
(grade II and III), macrophages tend to accumulate large amounts of
sterols (16). Increased
intracellular sterol concentration in macrophages promotes ABCA1
mRNA expression not only in a time- and dose-dependent fashion
(17,18), but also by activating nuclear
receptors, such as LXR and RXRα (19–21),
which may explain the lower mRNA levels in plaques of grade I.
Although the accumulated sterol has also been shown to up-regulate
ABCA1 protein expression in a dose-dependent fashion (17), it accelerates the ABCA1 degradation
in macrophages in a direct and indirect way to a greater extent
(22). The sterol-rich environment
inside the plaque is amplified by the low ABCA1 levels (23), which in return degrade the ABCA1
protein. Furthermore, ABCA1 has been shown to contain a PEST
sequence that enhances protein degradation (24). The similar ABCA1 distribution
pattern in plaques of grade II and III may be attributed to the
similar composition of the three constituents (15).
The retinoid X receptor (RXR) is a pleiotropic
nuclear receptor transcription factor that regulates the expression
of its target genes. RXR expressed in macrophages plays an
important role in plaque physiology, as it activates and suppresses
the ABCA1 gene expression in a RXR-responsive element-dependent
manner (25). Previous studies
have shown that ABCA1 transcription is stimulated by retinoid X and
the LXR receptor (25), and that
the induction of ABCA1 expression reflected that of LXRα (12,26).
In this study, we found that in atherosclerotic plaques, the ABCA1
and RXRα mRNA levels were significantly elevated, and a parallel
increase in mRNA levels of both genes was observed. The observed
up-regulation of ABCA1 and RXRα mRNA may be associated with the
increased oxysterol concentration in the plaque. The ABCA1 protein
may be the key factor that modulates the cholesterol homeostasis in
macrophages. The increased degradation of ABCA1 protein in the
plaques could result in decreased lipid efflux, accumulation of
oxysterol and the stimulation of regulatory pathways through
RXRα.
In summary, our findings demonstrate that the ABCA1
protein was notably reduced in atherosclerotic plaques, whereas the
ABCA1 and RXRα mRNA levels were significantly increased. The
failure of ABCA1 mRNA to translate into ABCA1 protein suggests that
new interventions should be targeted at the transcriptional
up-regulation of ABCA1 and its protein stabilization. Our results
also show that the unstable atherosclerotic plaques exhibited
higher ABCA1 mRNA levels, proposing the ABCA1 mRNA as a potential
biomarker of a particular subgroup of unstable carotid plaques.
Acknowledgements
This study was supported by grants
from the Zhengzhou Committee of Science and Technology (No.
083SGY2612-9).
References
|
1.
|
Thorvaldsen P, Kuulasmaa K, Rajakangas AM,
et al: Stroke trends in the WHO MONICA project. Stroke. 28:500–506.
1997. View Article : Google Scholar
|
|
2.
|
Delcker A, Diener HC and Wilhelm H:
Influence of vascular risk factors for atherosclerotic carotid
artery plaque progression. Stroke. 26:2016–2022. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Remaley AT, Stonik JA, Demosky SJ, et al:
Apolipoprotein specificity for lipid efflux by the human ABCAI
transporter. Biochem Biophys Res Commun. 280:818–823. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Wang N, Silver DL, Thiele C, et al:
ATP-binding cassette transporter A1 (ABCA1) functions as a
cholesterol efflux regulatory protein. J Biol Chem.
276:23742–23747. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Burris TP, Eacho PI and Cao G: Genetic
disorders associated with ATP binding cassette cholesterol
transporters. Mol Genet Metab. 77:13–20. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Oram JF, Lawn RM, Garvin MR, et al: ABCA1
is the cAMP-inducible apolipoprotein receptor that mediates
cholesterol secretion from macrophages. J Biol Chem.
275:34508–34511. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Aiello RJ, Brees D, Bourassa PA, et al:
Increased atherosclerosis in hyperlipidemic mice with inactivation
of ABCA1 in macrophages. Arterioscler Thromb Vasc Biol. 22:630–637.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
van Dam MJ, de Groot E, Clee SM, et al:
Association between increased arterial-wall thickness and
impairment in ABCA1-driven cholesterol efflux: an observational
study. Lancet. 359:37–42. 2002.PubMed/NCBI
|
|
9.
|
Stary HC, Chandler AB, Dinsmore RE, et al:
A definition of advanced types of atherosclerotic lesions and a
histological classification of atherosclerosis. A report from the
Committee on Vascular Lesions of the Council on Arteriosclerosis,
American Heart Association. Arterioscler Thromb Vasc Biol.
15:1512–1531. 1995. View Article : Google Scholar
|
|
10.
|
Bodzioch M, Orso E, Klucken J, et al: The
gene encoding ATP-binding cassette transporter 1 is mutated in
Tangier disease. Nat Genet. 22:347–351. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Wellington CL, Walker EK, Suarez A, et al:
ABCA1 mRNA and protein distribution patterns predict multiple
different roles and levels of regulation. Lab Invest. 82:273–283.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Albrecht C, Soumian S, Amey JS, et al:
ABCA1 expression in carotid atherosclerotic plaques. Stroke.
35:2801–2806. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Virmani R, Kolodgie FD, Burke AP, et al:
Lessons from sudden coronary death: a comprehensive morphological
classification scheme for atherosclerotic lesions. Arterioscler
Thromb Vasc Biol. 20:1262–1275. 2000. View Article : Google Scholar
|
|
14.
|
Lawn RM, Wade DP, Couse TL, et al:
Localization of human ATP-binding cassette transporter 1 (ABC1) in
normal and atherosclerotic tissues. Arterioscler Thromb Vasc Biol.
21:378–385. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Soumian S, Albrecht C, Davies AH, et al:
ABCA1 and atherosclerosis. Vasc Med. 10:109–119. 2005. View Article : Google Scholar
|
|
16.
|
Tabas I: Free cholesterol-induced
cytotoxicity a possible contributing factor to macrophage foam cell
necrosis in advanced atherosclerotic lesions. Trends Cardiovasc
Med. 7:256–263. 1997.PubMed/NCBI
|
|
17.
|
Langmann T, Klucken J, Reil M, et al:
Molecular cloning of the human ATP-binding cassette transporter 1
(hABC1): evidence for sterol-dependent regulation in macrophages.
Biochem Biophys Res Commun. 257:29–33. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Liao H, Langmann T, Schmitz G, et al:
Native LDL upregulation of ATP-binding cassette transporter-1 in
human vascular endothelial cells. Arterioscler Thromb Vasc Biol.
22:127–132. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Costet P, Luo Y, Wang N, et al:
Sterol-dependent transactivation of the ABC1 promoter by the liver
X receptor/retinoid X receptor. J Biol Chem. 275:28240–28245.
2000.PubMed/NCBI
|
|
20.
|
Fu X, Menke JG, Chen Y, et al:
27-Hydroxycholesterol is an endogenous ligand for liver X receptor
in cholesterol-loaded cells. J Biol Chem. 276:38378–38387. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Wagner BL, Valledor AF, Shao G, et al:
Promoter-specific roles for liver X receptor/corepressor complexes
in the regulation of ABCA1 and SREBP1 gene expression. Mol Cell
Biol. 23:5780–5789. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Uehara Y, Engel T, Li Z, et al:
Polyunsaturated fatty acids and acetoacetate downregulate the
expression of the ATP-binding cassette transporter A1. Diabetes.
51:2922–2928. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Schwartz K, Lawn RM and Wade DP: ABC1 gene
expression and ApoA-I-mediated cholesterol efflux are regulated by
LXR. Biochem Biophys Res Commun. 274:794–802. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Wang N, Chen W, Linsel-Nitschke P, et al:
A PEST sequence in ABCA1 regulates degradation by calpain protease
and stabilization of ABCA1 by apoA-I. J Clin Invest. 111:99–107.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Cui H, Okuhira K, Ohoka N, et al:
Tributyltin chloride induces ABCA1 expression and apolipoprotein
A-I-mediated cellular cholesterol efflux by activating
LXRalpha/RXR. Biochem Pharmacol. 81:819–824. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Laffitte BA, Joseph SB, Walczak R, et al:
Autoregulation of the human liver X receptor alpha promoter. Mol
Cell Biol. 21:7558–7568. 2001. View Article : Google Scholar : PubMed/NCBI
|