Introduction
Cancer is a major public health problem worldwide.
Epidemiological and animal studies indicate that the consumption of
vegetables and fruits with natural chemopreventive agents, alone or
in a mixture, is associated with a reduced risk of cancer
development (1–3).
The human genome encodes at least 15 DNA polymerases
that perform cellular DNA synthesis (4,5).
Eukaryotic cells contain three replicative DNA polymerases (α, δ
and ε), mitochondrial DNA polymerase γ and at least eleven
repair-related DNA polymerases [β, ζ, η, θ, ι, κ, λ, μ, ν,
terminal deoxynucleotidyl transferase (TdT) and REV1] (4–6).
Targeted DNA polymerase inhibition exerts an antitumor effect,
since replicative DNA polymerases are essential for cancer cell
growth (5). As a result of
screening for DNA polymerase inhibitors, we observed that
monogalactosyl diacylglycerol (MGDG) and sulfoquinovosyl
diacylglycerol (SQDG) inhibit replicative DNA polymerase activity
(7–10). Higher plants contain, particularly
within the thylakoid membranes of the chloroplast, major
glycoglycerolipids, including MGDG, SQDG and digalactosyl
diacylglycerol (DGDG) (11). It is
known that glycoglycerolipids are contained in vegetables, fruits
and grains (12,13), and we observed that spinach was the
most abundant source of glycoglycerolipids of the vegetables tested
(14). Therefore, to examine the
glycoglycerolipid properties, we purified these three
glycoglycerolipids from spinach and observed that MGDG was obtained
in the highest amount (15). MGDG
purified from spinach also inhibited replicative DNA polymerases.
However, it had no inhibitory effect on the other DNA polymerases
tested (repair type-DNA polymerases β, η, ι, κ, λ and μ).
DGDG did not influence all mammalian DNA polymerases, whereas SQDG
inhibited the activities of all (both replicative and
repair-related) mammalian DNA polymerases (7).
MGDG has been shown to exert bioactive effects on
cells and animals, including cancer cell growth inhibition
(7) and anti-angiogenesis
(16,17) and anti-inflammatory activity
(18). However, the inhibition of
tumor growth in vivo following the oral administration of
MGDG has not been evaluated. MGDG has characteristic high viscosity
and low solubility; γ-cyclodextrin (CD) was used to solve this
problem. In the present study, we investigated the bioactivity,
particularly the antitumor activity, of orally administered spinach
MGDG mixed with CD (CD-MGDG complex). In addition, we discuss the
effects of the CD-MGDG complex on proliferation and angiogenesis in
colon tumors that were observed using a histopathological
technique. We evaluated the antitumor effects of orally
administered MGDG in mice to develop a food-derived anticancer
compound.
Materials and methods
Preparation of CD-MGDG complex
MGDG was purified from dried spinach (Spinacia
oleracea L.) as described previously (7). An MGDG purification grade of >98%
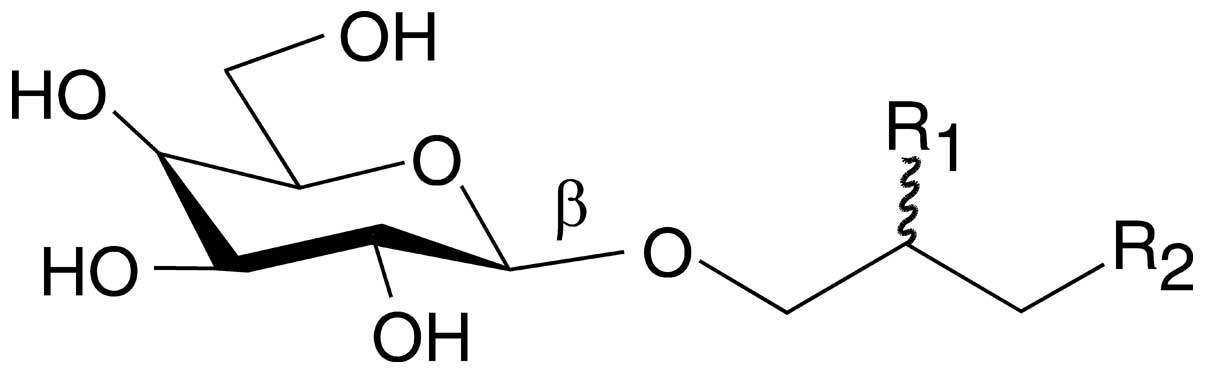
was used in this study. The chemical structure of MGDG is shown in
Fig. 1 and the composition of the
acyloxy groups of MGDG (R1 and R2 in Fig. 1) has been described previously
(7). MGDG (50 mg) was dissolved in
2.5 ml ethanol and a CD (Wako Pure Chemical Industries, Osaka,
Japan) solution (500 mg/2.5 ml in distilled water) was added. The
mixture of MGDG and CD was homogenized by mixing at 2500 rpm for 30
min at room temperature. After incubation overnight at room
temperature in the dark, the mixture of MGDG and CD was
freeze-dried in a vacuum at −50°C overnight.
In vitro anti-cell proliferation
activity
The mouse colon adenocarcinoma cell line, Colon26,
was provided by the Cell Resource Center for Biomedical Research
(Tohoku University, Sendai, Japan). The Colon26 cells were cultured
in RPMI-1640 (Wako Pure Chemical Industries) supplemented with 10%
fetal bovine serum (Equitech-Bio, Kerrville, TX, USA), penicillin
(100 U/ml, Nacalai Tesque, Kyoto, Japan) and streptomycin (100
μg/ml, Nacalai Tesque). The cells were cultured in an
atmosphere of 95% air and 5% CO2 at 37°C. Cell
proliferation was measured by the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT; Sigma-Aldrich, St. Louis, MO, USA) assay (19). The Colon26 cells were trypsinized
and plated in a 96-well plate at 5,000 cells per well (n=5) in
medium overnight. Following cell adhesion, MGDG (0–50 μM)
was added to the Colon26 cells. After 24 h, MTT solution was added
to all wells and the cells were incubated for 4 h at 37°C. The
medium containing MTT was removed and dimethyl sulfoxide (DMSO) was
added to each well to dissolve formazan crystals. The absorbance of
each well was measured using a microplate reader (Vmax; Molecular
Devices, Osaka, Japan) at test and reference wavelengths of 570 and
630 nm, respectively.
In vivo assessment of antitumor
activity
Five-week-old specific pathogen-free female Balb/c
mice were provided by Japan SLC (Shizuoka, Japan). The mice were
fed a standard diet (MF; Oriental Yeast Co., Ltd., Osaka, Japan)
and had free access to water. The present study was approved by the
Kobe-Gakuin University Animal Committee according to the guidelines
for the ‘Care and Use of Laboratory Animals’ of the University.
Following one week of breeding, s.c. (hypodermic
injection) tumors were induced by the inoculation of
1×106 Colon26 cells s.c. into the Balb/c mice. The
tumor-bearing mice were divided randomly into three groups and
treatment was initiated with the CD-MGDG complex or vehicle control
(CD alone) 5 days after tumor inoculation, when the tumors had
achieved a tumor volume [tumor volume = length ×
(width)2 × 0.5] of 25–50 mm3. The CD-MGDG
complex groups (44 or 220 mg/kg; n=5 or 6, respectively) were
treated orally (p.o.) with 4 or 20 mg/kg equivalent (eq.) of MGDG,
daily for 26 days. The control mice received CD alone p.o. (200
mg/kg; n=6) daily prior to examination.
When the treatment was completed, all mice were
examined at necropsy for gross organ abnormalities. The lungs,
heart, spleen, stomach, liver, pancreas, kidney, intestine and
brain were collected, fixed in 10% formalin, and embedded in
paraffin for histopathological evaluation with hematoxylin and
eosin (H-E) staining. The tumors were also embedded in paraffin for
H-E and proliferating cell nuclear antigen (PCNA) staining. The
remainder of the tumor was embedded in optimal cutting temperature
(OCT) compound (Sakura Finetek Japan, Tokyo, Japan) for
histopathological evaluation by CD31 staining and a terminal
deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL)
assay.
Assessment of tumor cell
proliferation
Tumor cell proliferation was measured using the
mitotic index and PCNA expression with H-E and immunohistochemical
staining. Sections (3 μm) were deparaffinized in xylene and
alcohol, and transferred to phosphate-buffered saline (PBS). The
deparaffinized sections were stained with H-E to calculate the
number of mitotic cells. The mitotic index [number of cells in
mitosis/high power field (HPF)] of five random non-necrotic fields
at ×400 magnification was determined. The remaining deparaffinized
sections were stained with PCNA monoclonal antibody (sc-53; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA) to evaluate cell
proliferation. Sections (3 μm) were incubated with PCNA
antibody (1:500 dilution) overnight at 4°C. The sections were then
rinsed three times with PBS for 5 min each and the slides were
incubated with a secondary goat anti-mouse antibody conjugated to
peroxidase (Nichirei Biosciences, Tokyo, Japan) for 10 min at room
temperature. The sections were washed three times with PBS for 5
min. Positive reactions were rendered visible by incubating the
slides with 3,3′-diaminobenzidine (DAB; Nichirei Biosciences) for 5
min at room temperature. The sections were rinsed with distilled
water, counterstained with hematoxylin for 5 sec and mounted. To
quantify PCNA expression, the number of positive cells was counted
in 3–5 fields at ×400 magnification.
Immunofluorescence staining for CD31
Fresh frozen tissues were cut into 10-μm
sections and mounted on positively charged slides. The sections
were fixed in cold methanol/acetone (1:1) for 10 min and then
washed three times with PBS for 5 min each time. The slides were
placed in a humidified chamber and incubated with 5% goat serum for
20 min at room temperature. The sections were then incubated
overnight with a 1:400 dilution of rat anti-mouse CD31 antibody (BD
Pharmingen, San Diego, CA, USA) at 4°C. The sections were then
rinsed three times with PBS for 5 min each and the slides were
incubated with a secondary goat anti-rat IgG antibody conjugated to
Alexa 594 (1:500 dilution; Invitrogen Japan K.K., Tokyo, Japan) for
10 min at room temperature. The slides were then washed three times
with PBS for 5 min each, mounting medium was placed on each slide
and the slides were covered with glass coverslips. To quantify CD31
expression, five fields at ×250 magnification were examined for
each tumor with a confocal laser scanning microscope (LSM 510 META,
Carl Zeiss MicroImaging, Tokyo, Japan). The fluorescence images
were analyzed using ZEN software (Carl Zeiss MicroImaging).
TUNEL assay
The TUNEL assay was performed using an Apoptosis
Detection kit (Takara Bio, Shiga, Japan). Fresh frozen sections
were fixed with 4% paraformaldehyde for 30 min at 4°C and then
washed with PBS for 30 min. The sections were incubated with 0.3%
H2O2 in methanol for 30 min to block
endogenous peroxidase and then washed three times with PBS for 5
min each. The sections were permeabilized with permeabilization
buffer on ice for 5 min. The slides were placed in a humidified
chamber and incubated with TdT enzyme including fluorescein
isothiocyanate (FITC)-conjugated dUTP for 60 min at 37°C. The
slides were washed three times with PBS for 5 min. The sections
were incubated with anti-FITC HRP conjugate for 30 min at 37°C and
the slides were washed three times with PBS for 5 min. Positive
reactions were rendered visible by incubating the slides with DAB
for 10 min at room temperature. The sections were rinsed with
distilled water, conterstained with hematoxylin for 5 sec and
mounted. To quantify TUNEL-positive expression, the number of
positive cells was counted in five fields at ×400
magnification.
Statistical analysis
All experiments show the mean ± SE between groups.
Comparisons were made using the Mann-Whitney U test or Steel’s test
using the KyPlot 5.0 software package (LyensLab, Tokyo, Japan).
P<0.05 was considered to indicate a statistically significant
result.
Results
Effect of spinach MGDG on in vitro cancer
cell proliferation
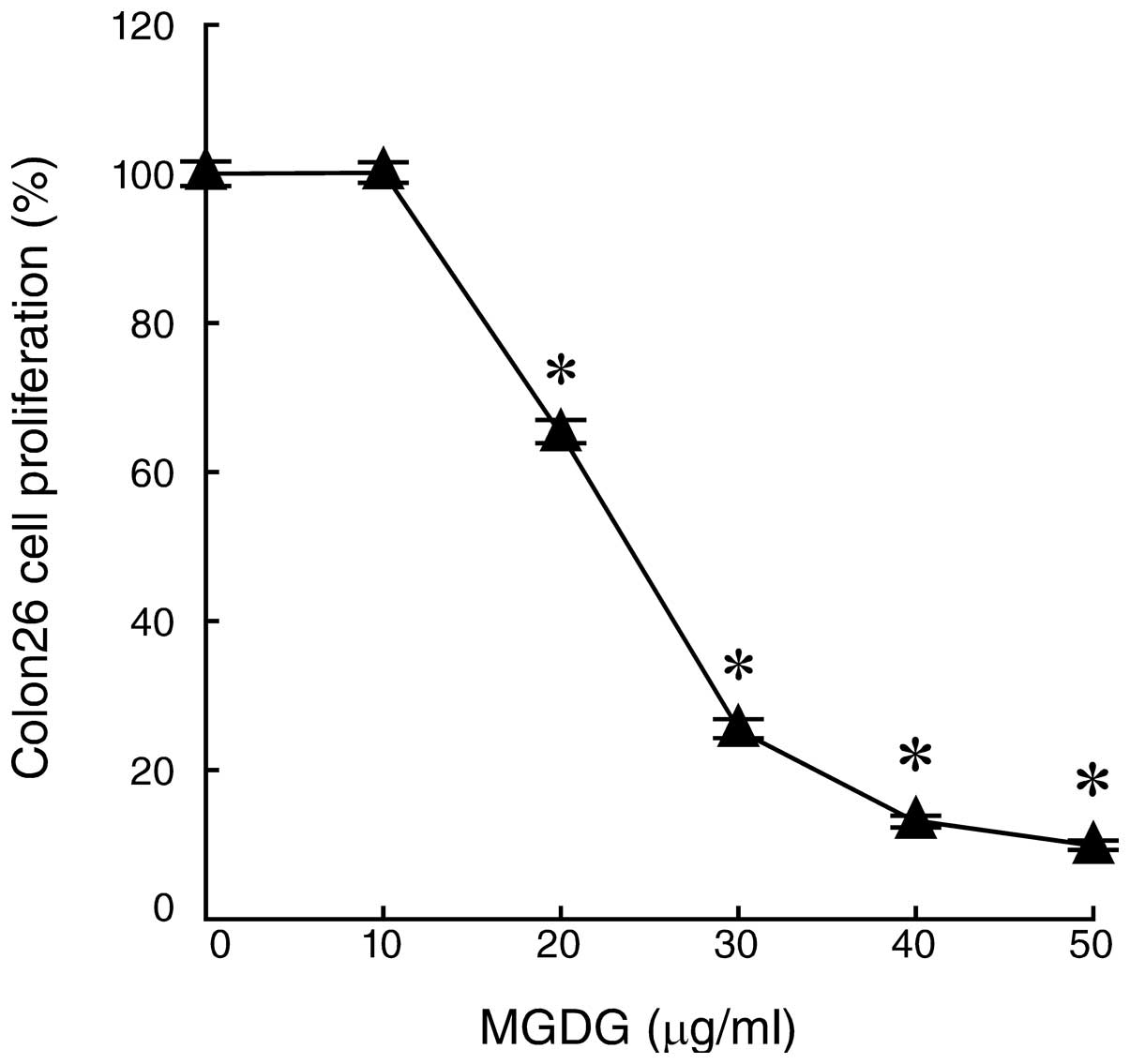
First, we investigated the effects of purified MGDG
from spinach (Fig. 1) on the cell
growth suppression of a mouse colon cancer cell line, Colon26,
in vitro. These cultured cells were tested using an MTT
assay following incubation with 0, 10, 20, 30, 40 or 50
μg/ml MGDG for 24 h. MGDG significantly inhibited the
proliferation of the Colon26 cells in a dose-dependent manner and
the LD50 value was 24 μg/ml (Fig. 2). Previously, we reported that MGDG
suppressed the human gastric cancer cell line NUGC-3 to almost the
same extent as the Colon26 cell line (7). These results indicate that MGDG is an
effective inhibitor of colon cancer cell growth.
Effect of CD-MGDG complex on in vivo
tumor graft growth in mice
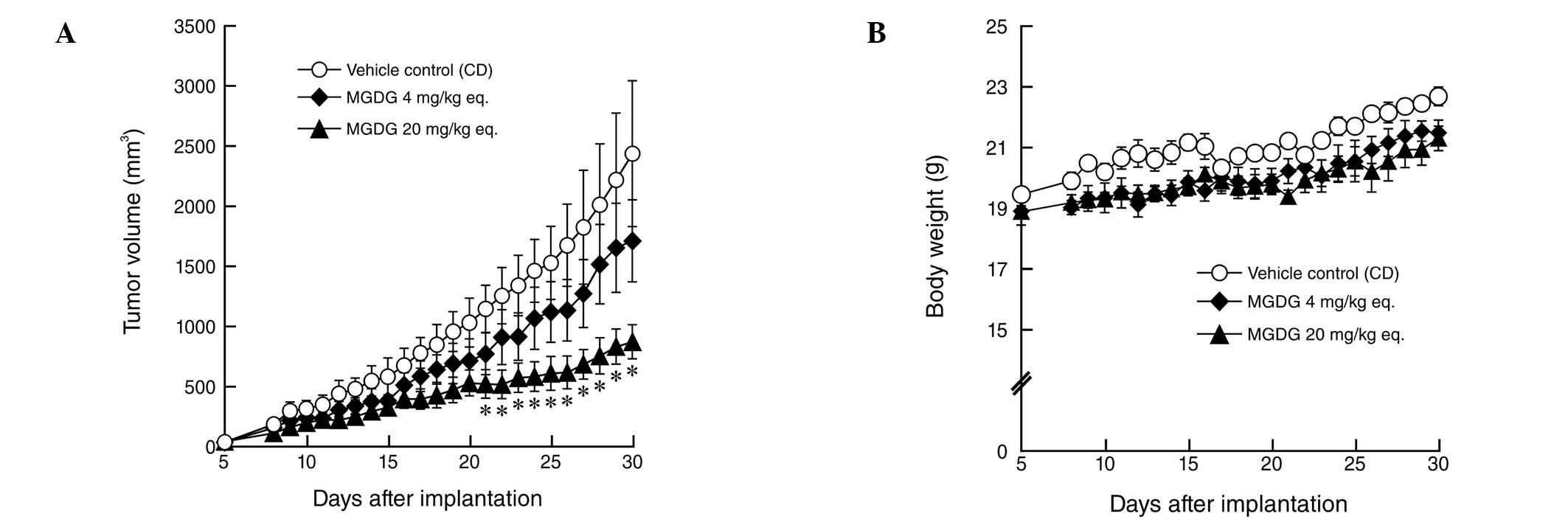
The Colon26 mouse colon cells (1×106)
were inoculated s.c. into mice. Five days later, the mice received
200 mg/kg CD alone (control), CD (40 mg/kg)-MGDG (4 mg/kg) complex
or CD (200 mg/kg)-MGDG (20 mg/kg) complex daily for 26 days. As
shown in Fig. 3A, tumors in the
control group continued to grow rapidly. By contrast, in the mice
treated with purified MGDG from spinach, tumor growth was
suppressed in a dose-dependent manner. Treatment of the mice with 4
and 20 mg/kg MGDG eq. inhibited Colon26 tumor growth by 29.8
(P>0.1) and 64.2% (P<0.05), respectively, relative to
vehicle-treated controls. In the present study, mice treated with
MGDG appeared healthy and showed no marked weight loss compared
with vehicle-treated controls (Fig.
3B). Histopathological analysis showed that the lungs, heart,
spleen, stomach, liver, pancreas, kidney, intestine and brain of
MGDG-treated mice were normal. These findings suggest that MGDG did
not have side effects, including mortality or evident toxicity,
loss of body weight and/or major organ damage, in mice.
Effect of CD-MGDG complex on cell
proliferation of mouse colon tumor tissue in vivo
To assess the in vivo effect of CD-MGDG
complex consumption on the proliferation of tumor tissue in mice,
the samples were analyzed by H-E staining and PCNA immunostaining.
Qualitative histopathologic analysis of H-E-stained sections
revealed a substantial decrease in the number of mitotic cells in
the tumor tissue of the MGDG-treated mice compared with that of the
vehicle-treated controls (Table 1;
P<0.05 for 20 mg/kg MGDG eq.). Qualitative histopathological
analysis of PCNA-stained sections revealed a decrease in the number
of PCNA-positive cells in the tumor tissue of the MGDG-treated mice
compared with that of the vehicle-treated controls (Table I; P<0.05 for each dosage).
Compared with tumors from mice receiving vehicle alone, the number
of mitotic cells and percentage of PCNA-positive cells were reduced
by 38.7 and 22.9%, respectively, in the tumors from mice that
received 20 mg/kg MGDG eq. These results suggest that orally
administered MGDG prevented the mitotic cell proliferation of
Colon26 solid tumors in mice in a dose-dependent manner.
 | Table ICounts of mitotic and PCNA-positive
cells in Colon26 tumor sections. |
Table I
Counts of mitotic and PCNA-positive
cells in Colon26 tumor sections.
| Variable | Mitosis (/HPF) | PCNA (%) |
|---|
| Vehicle control
(CD) | 19.1±1.6 | 77.0±1.4 |
| CD-MGDG complex
(MGDG 4 mg/kg eq.) | 14.7±1.5 | 67.3±2.2a |
| CD-MGDG complex
(MGDG 20 mg/kg eq.) | 11.7±1.3a | 59.4±2.5a |
Immunofluorescence staining for CD31 in
colon tumor tissue from mice
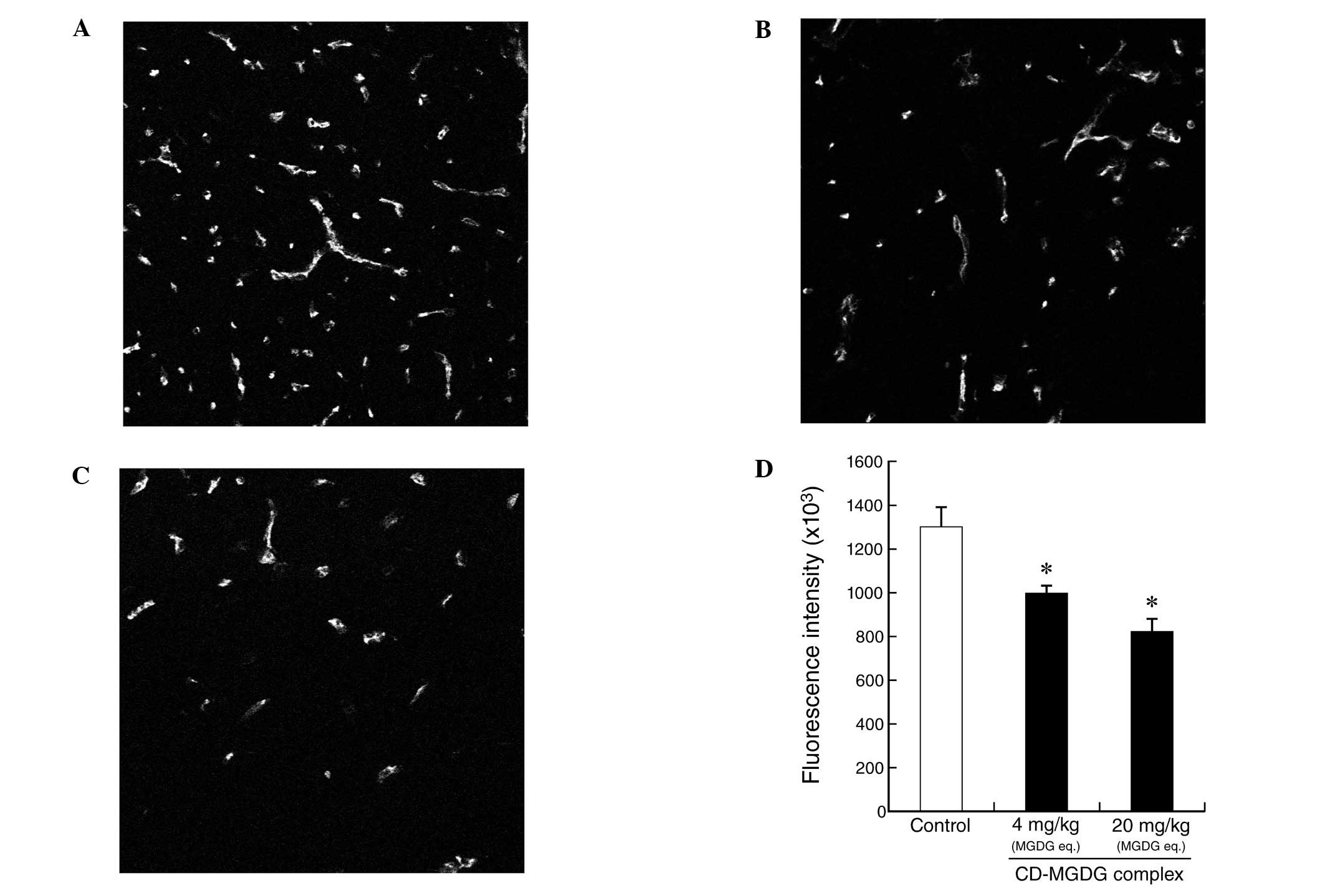
To evaluate the anti-angiogenesis effect in
vivo, MGDG was evaluated for dose-dependent modulation of
microvessels by immunostaining for CD31. As shown in Fig. 4A–C, a dose-dependent reduction in
the staining intensity of the CD31-positive endothelial cells by
MGDG was observed at doses of 4 and 20 mg/kg/day in Colon26 solid
tumor tissue, using a microscope equipped for immunofluorescence
analysis. Mice treated with 4 mg/kg and 20 mg/kg MGDG eq. had
fluorescence intensities of 999±35×103 (P<0.05) and
824±58×103 P<0.05) and inhibited CD31 expression by
23.2 and 36.7%, respectively, compared with the vehicle control
(1301±91×103; Fig. 4D).
The typical averages indicate that MGDG prevented not only
CD31-positive endothelial cell expression intensity but also
microvessel formation (Fig.
4A–C).
Effect of CD-MGDG complex on apoptosis
induction of colon tumor tissue in mice
The histopathologist randomly observed tumor tissue
sections with H-E staining. The number of apoptotic cells tended to
increase as the dose of MGDG increased (data not shown). We used a
TUNEL technique to detect apoptotic cells in Colon26 tumor tissue
sections. The number of TUNEL-positive apoptotic cells was
increased in the tumors from the mice treated with MGDG in a
dose-dependent manner compared with that in tumors from mice
treated with the vehicle (Table
II). These data indicate that MGDG administration effectively
induces apoptosis in tumor cells in vivo, as has been
demonstrated in vitro in a previous study (7).
| Table IITUNEL analysis of Colon26 tumor
sections. |
Table II
TUNEL analysis of Colon26 tumor
sections.
| Variable | TUNEL-positive
cells (/HPF) |
|---|
| Vehicle control
(CD) | 5.2±0.9 |
| CD-MGDG complex
(MGDG 4 mg/kg eq.) | 9.0±0.8 |
| CD-MGDG complex
(MGDG 20 mg/kg eq.) | 13.7±1.8a |
Discussion
In the current study, we evaluated the efficacy of
oral CD-MGDG complex administration for the treatment of implanted
solid tumors in mice. CD is able to render fat-soluble materials
water-soluble, therefore, the CD-MGDG complex may be useful as an
anticancer functional food and/or drug. CD itself is digested in
the body and is a safe agent (20). In the in vitro cultured
cancer cell growth assay, MGDG alone (without CD) significantly
suppressed the proliferation of colon cancer cells (Fig. 2). The vehicle control (CD alone)
had no effective antitumor activity (Fig. 3A), therefore, purified MGDG from
spinach may be effective. These results show that MGDG inhibits
tumor growth in a dose-dependent manner and immunohistochemical
analysis suggests that the antitumor efficacy of MGDG may be
associated with anti-angiogenetic, anti-proliferative and apoptotic
effects in tumor tissue without adverse health effects.
MGDG is a non-nutrient compound contained in
vegetables, grains and fruits. MGDG content differs among plants
(12) and is ingested daily in
food. The chemical structure of MGDG comprises two acyloxy groups
(R1 and R2 in Fig. 1) derived from fatty acid molecules.
In the present study, we used spinach MGDG which is rich in n-3
α-linolenic acid (26.3% of the total fatty acids in spinach MGDG)
(7). MGDG from wheat flour
includes non-n-3 fatty acids, such as linoleic acid, which is an
n-6 fatty acid, and saturated fatty acids (21). However, the fatty acid composition
influences the antitumor effect (22). Therefore, these findings suggest
that researchers should observe and note the lipid contents and
fatty acid composition in MGDG studies.
As few studies have investigated MGDG, it is unknown
whether MGDG is absorbed. Previously, we reported that mammalian
lipase is able to hydrolyze some MGDG into monogalactosyl
monoacylglycerol (MGMG) in vitro(15). A study of orally administered MGDG
in vivo revealed that it is digested to MGMG and
monogalactosyl glycerol (MGG) by a digestive enzyme and
enterobacteria, respectively, and MGG is not absorbed (21). However, the oral administration of
MGDG demonstrated strong and dose-dependent inhibition of tumor
growth in a mouse model in the current study. These results suggest
that part of the MGDG compound, such as MGMG and/or MGG, was
absorbed and re-synthesized or that undifferentiated MGDG entered
the blood stream. In addition, these results suggest that MGDG is
not completely degraded and possesses antitumor activity, since
components of MGDG, galactose and glycerol, do not have an
antitumor effect, and fatty acids have a weak antitumor effect
(23,24).
As shown in Tables
I and II and Fig. 4, we analyzed tumor tissue following
the administration of MGDG to mice. These results show that MGDG
prevented tumor growth by inhibiting angiogenesis and and reduced
the count of cells that stained positive for PCNA, which is a
proliferation marker (25,26), and was accompanied by an increase
of apoptosis. Certain studies have suggested that replicative DNA
polymerases (α, δ and ε) and tumor angiogenesis have potential as
cancer therapeutic targets (27–31).
In particular, replicative DNA polymerases are essential for cancer
cell proliferation. In addition, tumor growth depends on
angiogenesis, since tumors have to be located within 200 μm
of blood vessels to obtain nutrients and oxygen (32). We previously observed that MGDG
from spinach inhibited the activities of mammalian replicative DNA
polymerases, but had no inhibitory effect on other mammalian DNA
polymerases, including repair-related β, η, ι, κ, λ and μ,
and MGDG suppressed human umbilical vein endothelial cells (HUVEC)
tube formation, HUVEC proliferation and tumor angiogenesis in
vitro and ex vivo(16,17).
However, the data in Fig. 2 and
Table I suggest that the antitumor
effects of MGDG are not entirely dependent on angiogenesis. The
present in vitro study did not include endothelial cells and
it appears that MGDG has a direct effect on tumor cells.
In conclusion, the present study showed that the
natural product MGDG obtained from vegetables, fruits and grains is
safe with a potent oral antitumor effect, including
anti-proliferative, anti-angiogenesis and apoptosis-inducing
activity. Our results suggest that MGDG from food has
cancer-preventive and health-promotion effects.
Abbreviations:
|
MGDG
|
monogalactosyl diacylglycerol
|
|
CD
|
cyclodextrin
|
|
PCNA
|
proliferating cell nuclear antigen
|
|
TUNEL
|
terminal deoxynucleotidyl transferase
dUTP nick-end labeling
|
|
SQDG
|
sulfoquinovosyl diacylglycerol
|
|
DGDG
|
digalactosyl diacylglycerol
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium
bromide
|
|
H-E
|
hematoxylin and eosin
|
|
HUVEC
|
human umbilical vein endothelial
cell
|
Acknowledgements
We thank Dr Mami Yamaguchi, Mr Seiji
Ohtani and Mr Michitoshi Kimura of Sapporo Medical University
School of Medicine for the histopathological examination and
technical advice on the tumor samples.
Y.M. acknowledges Grant-in-Aids for Scientific
Research (C) (No. 24580205) from MEXT (Ministry of Education,
Culture, Sports, Science and Technology, Japan), Adaptable and
Seamless Technology transfer Program through target-driven R&D
(A-STEP) from JST (Japan Science and Technology Agency), Takeda
Science Foundation (Japan) and the Nakashima Foundation (Japan).
This study was supported in part by the MEXT-Supported Program for
the Strategic Research Foundation at Private Universities,
2012–2016.
References
|
1
|
Terry P, Giovannucci E, Michels KB,
Bergkvist L, Hansen H, Holmberg L and Wolk A: Fruit, vegetables,
dietary fiber, and risk of colorectal cancer. J Natl Cancer Inst.
93:525–533. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Surh YJ: Cancer chemoprevention with
dietary phytochemicals. Nat Rev Cancer. 3:768–780. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu RH: Potential synergy of
phytochemicals in cancer prevention: mechanism of action. J Nutr.
134:3479S–3485S. 2004.PubMed/NCBI
|
|
4
|
Hubscher U, Maga G and Spadari S:
Eukaryotic DNA polymerases. Annu Rev Biochem. 71:133–163. 2002.
View Article : Google Scholar
|
|
5
|
Bebenek K and Kunkel TA: DNA repair and
replication. Advances in Protein Chemistry. Yang W: 69. Elsevier;
San Diego, CA: pp. 137–165. 2004
|
|
6
|
Takata K, Shimizu T, Iwai S and Wood RD:
Human DNA polymerase N (POLN) is a low fidelity enzyme capable of
error-free bypass of 5S-thymine glycol. J Biol Chem.
281:23445–23455. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Murakami C, Kumagai T, Hada T, Kanekazu U,
Nakazawa S, Kamisuki S, Maeda N, Xu X, Yoshida H, Sugawara F,
Sakaguchi K and Mizushina Y: Effects of glycolipids from spinach on
mammalian DNA polymerases. Biochem Pharmacol. 65:259–267. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ohta K, Mizushina Y, Hirata N, Takemura M,
Sugawara F, Matsukage A, Yoshida S and Sakaguchi K:
Sulfoquinovosyldiacylglycerol, KM043, a new potent inhibitor of
eukaryotic DNA polymerases and HIV-reverse transcriptase type 1
from a marine red alga, Gigartina tenella. Chem Pharm Bull
(Tokyo). 46:684–686. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hanashima S, Mizushina Y, Ohta K, Yamazaki
T, Sugawara F and Sakaguchi K: Structure-activity relationship of a
novel group of mammalian DNA polymerase inhibitors, synthetic
sulfoquinovosylacylglycerols. Jpn J Cancer Res. 91:1073–1083. 2000.
View Article : Google Scholar
|
|
10
|
Maeda N, Hada T, Yoshida H and Mizushina
Y: Inhibitory effect on replicative DNA polymerases, human cancer
cell proliferation, and in vivo anti-tumor activity by glycolipids
from spinach. Curr Med Chem. 14:955–967. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Roughan PG and Batt RD: The glycerolipid
composition of leaves. Phytochemistry. 8:363–369. 1969. View Article : Google Scholar
|
|
12
|
Sugawara T and Miyazawa T: Separation and
determination of glycolipids from edible plant sources by
high-performance liquid chromatography and evaporative
light-scattering detection. Lipids. 34:1231–1237. 1999.PubMed/NCBI
|
|
13
|
Yunoki K, Sato M, Seki K, Ohkubo T, Tanaka
Y and Ohnishi M: Simultaneous quantification of plant
glyceroglycolipids including sulfoquinovosyldiacylglycerol by
HPLC-ELSD with binary gradient elution. Lipids. 44:77–83. 2009.
View Article : Google Scholar
|
|
14
|
Kuriyama I, Musumi K, Yonezawa Y, Takemura
M, Maeda N, Iijima H, Hada T, Yoshida H and Mizushina Y: Inhibitory
effects of glycolipids fraction from spinach on mammalian DNA
polymerase activity and human cancer cell proliferation. J Nutr
Biochem. 16:594–601. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Maeda N, Hada T, Murakami-Nakai C,
Kuriyama I, Ichikawa H, Fukumori Y, Hiratsuka J, Yoshida H,
Sakaguchi K and Mizushina Y: Effects of DNA polymerase inhibitory
and antitumor activities of lipase-hydrolyzed glycolipid fractions
from spinach. J Nutr Biochem. 16:121–128. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Matsubara K, Matsumoto H, Mizushina Y,
Mori M, Nakajima N, Fuchigami M, Yoshida H and Hada T: Inhibitory
effect of glycolipids from spinach on in vitro and ex vivo
angiogenesis. Oncol Rep. 14:157–160. 2005.PubMed/NCBI
|
|
17
|
Maeda N, Matsubara K, Yoshida H and
Mizushina Y: Anti-cancer effect of spinach glycoglycerolipids as
angiogenesis inhibitors based on the selective inhibition of DNA
polymerase activity. Mini Rev Med Chem. 11:32–38. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bruno A, Rossi C, Marcolongo G, Di Lena A,
Venzo A, Berrie CP and Corda D: Selective in vivo anti-inflammatory
action of the galactolipid monogalactosyldiacylglycerol. Eur J
Pharmacol. 524:159–168. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mosmann T: Rapid colorimetric assay for
cellular growth and survival: application to proliferation and
cytotoxicity assays. J Immunol Methods. 65:55–63. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fukuda K, Teramoto Y, Goto M, Sakamoto J,
Mitsuiki S and Hayashida S: Specific inhibition by cyclodextrins of
raw starch digestion by fungal glucoamylase. Biosci Biotechnol
Biochem. 56:556–559. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sugawara T and Miyazawa T: Digestion of
plant monogalactosyldiacylglycerol and digalactosyldiacylglycerol
in rat alimentary canal. J Nutr Biochem. 11:147–152. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Matsui Y, Hada T, Maeda N, Sato Y,
Yamaguchi Y, Takeuchi T, Takemura M, Sugawara F, Sakaguchi K,
Yoshida H and Mizushina Y: Structure and activity relationship of
monogalactosyl diacylglycerols, which selectively inhibited in
vitro mammalian replicative DNA polymerase activity and human
cancer cell growth. Cancer Lett. 283:101–107. 2009. View Article : Google Scholar
|
|
23
|
Grammatikos SI, Subbaiah PV, Victor TA and
Miller WM: n-3 and n-6 fatty acid processing and growth effects in
neoplastic and non-cancerous human mammary epithelial cell lines.
Br J Cancer. 70:219–227. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Coakley M, Banni S, Johnson MC, Mills S,
Devery R, Fitzgerald G, Paul Ross R and Stanton C: Inhibitory
effect of conjugated α-linolenic acid from bifidobacteria of
intestinal origin on SW480 cancer cells. Lipids. 44:249–256.
2009.
|
|
25
|
Kubben FJ, Peeters-Haesevoets A, Engels
LG, Baeten CG, Schutte B, Arends JW, Stockbrugger RW and Blijham
GH: Proliferating cell nuclear antigen (PCNA): a new marker to
study human colonic cell proliferation. Gut. 35:530–535. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bromley M, Rew D, Becciolini A, Balzi M,
Chadwick C, Hewitt D, Li YQ and Potten CS: A comparison of
proliferation markers (BrdUrd, Ki-67, PCNA) determined at each cell
position in the crypts of normal human colonic mucosa. Eur J
Histochem. 40:89–100. 1996.PubMed/NCBI
|
|
27
|
Maeda N, Kokai Y, Ohtani S, Sahara H,
Kuriyama I, Kamisuki S, Takahashi S, Sakaguchi K, Sugawara F,
Yoshida H, Sato N and Mizushina Y: Anti-tumor effects of
dehydroaltenusin, a specific inhibitor of mammalian DNA polymerase
α. Biochem Biophys Res Commun. 352:390–396. 2007.PubMed/NCBI
|
|
28
|
Kuriyama I, Mizuno T, Fukudome K,
Kuramochi K, Tsubaki K, Usui T, Imamoto N, Sakaguchi K, Sugawara F,
Yoshida H and Mizushina Y: Effect of dehydroaltenusin-C12
derivative, a selective DNA polymerase α inhibitor, on DNA
replication in cultured cells. Molecules. 13:2948–2961.
2008.PubMed/NCBI
|
|
29
|
Berdis AJ: DNA polymerases as therapeutic
targets. Biochemistry (Moscow). 47:8253–8260. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Folkman J: Tumor angiogenesis: therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Folkman J: Tumor angiogenesis. Adv Cancer
Res. 43:175–203. 1985. View Article : Google Scholar
|
|
32
|
Bossi P, Viale G, Lee AK, Alfano R, Coggi
G and Bosari S: Angiogenesis in colorectal tumors: microvessel
quantitation in adenomas and carcinomas with clinicopathological
correlations. Cancer Res. 55:5049–5053. 1995.PubMed/NCBI
|