Introduction
Serum albumin is synthesized mainly in the liver
(1) and secreted into blood where
it constitutes almost 50% of the proteins in the serum (2–5).
Albumin also produces the majority of the protein contents of
extracellular fluids such as interstitial, cerebrospinal, and
lymphatic (2,6,7).
Albumins are simple proteins without prosthetic groups,
glycosylation or lipid post-translational modification. Serum
albumin is a highly soluble, single-chain polypeptide comprising
almost 585 amino acids (8–10). The development of the serum albumin
gene has occurred due to gene triplication of monomeric serum
albumin (11,12). Results of multiple sequence
analysis of serum albumin revealed the well-characterized
replication of three long homologous domains of 180 amino acids
(13). Results of X-ray
crystallographic analyses of various albumins revealed that their
structures consist of only α helices (∼67%), connected by long
flexible loops (14,15). The 3D structure analysis of serum
albumin also demonstrated that this heart-shaped globular protein
comprises three homologous domains (I, II and III), each of which
consist of sub-domains A and B (16). Albumin is a disulfide-rich protein
(17 disulfide bonds) that provides rigidity in the molecule while
the long connecting loops provide flexibility and binding
properties (17–19).
Serum albumin performs several vital functions
(2,20,21)
including transport of hormones, endogenous and exogenous
molecules, fatty acids, bilirubin, numerous substances that are
toxic in free form, as well as involvement in inflammatory
responses, and prevention of the photo-degradation of folic acid
(2,22–25).
Serum albumins are multifunctional and economically viable due to
their extensive use in the pharmaceutical and biotechnological
industries (26,27). They are employed as stabilizers in
therapeutic proteins, vaccines and enzymes (28–30).
Serum albumins are amphiphilic in nature and could be utilized to
prevent adsorption of other active proteins on the surface of the
container, or could be included in the formulation to suppress
protein aggregation (31,32).
The camel (Camelus dromedarius) is an
extremely adaptable mammal capable of surviving in extremely hot
climates without water intake for several weeks. Therefore, camel
serum albumin (CSA) may possess unique ligand binding and
stability/folding properties. In the present study, we have adopted
a chromatographic method that may be used to purify CSA to
homogeneity and characterized some of its properties using
capillary electrophoresis, high-performance liquid chromatography
(HPLC) and mass spectrometry. The purpose of this study was to
explore the possibility of substituting HSA/BSA with CSA in all
cases. However, these proteins are currently not available or
cannot be utilized due to variety of considerations.
Materials and methods
Materials
Fresh camel blood was purchased from the
slaughterhouse. Blue-Sepharose, Q-Sepharose and Sephacryl S-100
column, as well as low molecular weight (LMW) markers were obtained
from GE Healthcare (Pittsburgh, PA, USA). ÄKTA purifier and
SDS-PAGE assembly were from GE Healthcare. HPLC (Agilent 1260
Infinity LC system) and capillary electrophoresis (CE) (7100
capillary electrophoresis) were from Agilent Technologies (Santa
Clara, CA, USA). Mass spectrometry-based proteomics analysis was
performed at the Proteomics Resource Facility, University of
Michigan, MI, USA, using multidimensional proteomic identification
technology. All the chemicals used were of analytical grade.
Plasma preparation
Camel blood was collected in a beaker containing
anticoagulant (10 mg/ml EDTA). Anticoagulant was mixed gently using
glass rod. Blood samples were transported on ice. Clear supernatant
was separated following centrifugation at 1,000 × g for 30 min at
4°C. Plasma was aliquoted and stored at −80°C.
Purification of CSA on Blue-Sepharose
column
Frozen plasma was thawed on ice and 5 ml of plasma
aliquot was centrifuged at 24,000 × g for 10 min at 4°C to remove
debris. The supernatant was passed through a 0.45 micron syringe
filter. To reduce ionic strength, plasma was diluted 10-fold in 20
mM Tris-HCl, pH 8.0. The Blue-Sepharose column was equilibrated
with 20 mM Tris-HCl, pH 8.0. Diluted plasma was passed through
pre-equilibrated Blue-Sepharose column. Flow-through was collected
for further analysis. The column was washed extensively (5 CV) with
20 mM Tris-HCl, pH 8.0. Bound protein was eluted with a linear
gradient of NaCl; the elution buffer was 20 mM Tris-HCl, 2 M NaCl,
pH 8.0. Fractions were loaded on 12% SDS-PAGE to analyze the
purity. Fractions containing a band corresponding to albumin were
pooled and concentrated by Amicon 8050 stirred cells. Concentrated
pooled protein was dialyzed (1:100 v/v) twice against 20 mM
Tris-HCl, pH 8.0.
Purification of CSA on Q-Sepharose
Dialyzed protein sample was centrifuged at 24,000 ×
g for 30 min at 4°C and filtered through 0.45 micron filter.
Protein sample was passed through a pre-equilibrated (20 mM
Tris-HCl, pH 8.0) Q-Sepharose column using a syringe. The column
was washed with 10 CV of 20 mM Tris-HCl, pH 8.0. Bound proteins
were eluted on fast protein liquid chromatography (FPLC) with a
buffer of 20 mM Tris-HCl, 2 M NaCl, pH 8.0 using a 0–50% gradient
of 2 M NaCl for 50 min at a 1 ml/min flow rate. Purity of the
eluted fractions was analyzed by 12% SDS-PAGE and pure fractions
were pooled for gel filtration chromatography.
Purification of CSA by gel
filtration
Sephacryl S-100 column was equilibrated with 20 mM
Tris-HCl, 300 mM NaCl, pH 8.0. Protein sample was loaded using 10
ml superloop. Fractions were collected at a 1 ml/ml flow-rate.
Purity of the fractions was analyzed by 12% SDS-PAGE. Pure
fractions were pooled and dialyzed three times against water (1:100
v/v).
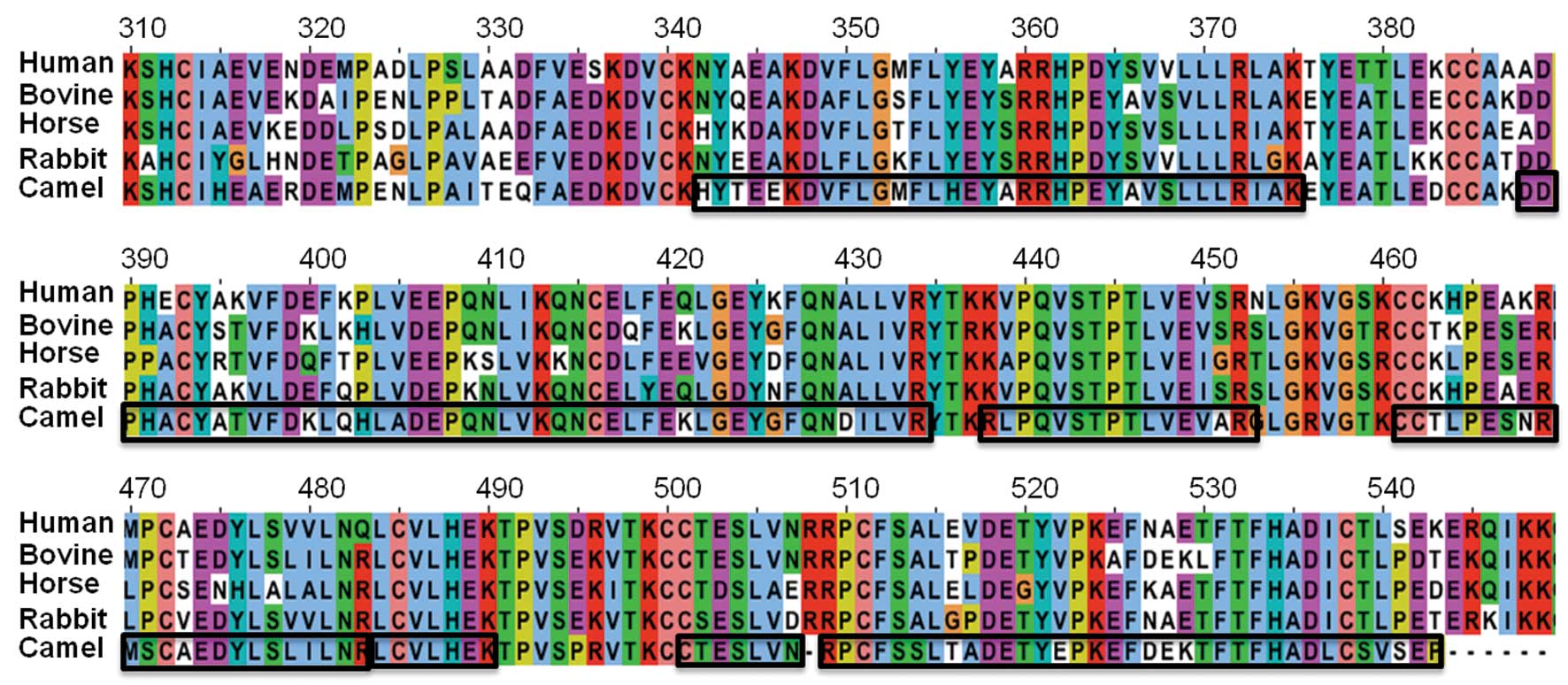
Multiple sequence alignment
Partial mRNA sequence of CSA (NCBI accession no.
HM640019.1) was translated into protein sequences using Ex PASy web
tools (http://web.expasy.org/translate/). Multiple sequence
alignment was performed using the Jalview program (http://www.jalview.org/) between human (P02768),
bovine (P02769), horse (P35747), and rabbit (P49065), as well as
the partial sequence of CSA.
Mass spectrometry and protein
analysis
Gel proteolysis was performed as described in a
previous study (33). The protein
band stained with Coomassie Brilliant Blue was excised from 12%
gradient SDS-PAGE gel. The excised gel piece was destained for 4 h
in 10 ml of 30% methanol. After destaining, the excised gel piece
was incubated for 30 min in 200 μl of 1:1 mixture of 100 mM
ammonium bicarbonate buffer with acetonitrile (buffer A). The gel
was then transferred into a reducing buffer comprising 10 mM
dithiothreitol (DTT) in 100 mM ammonium bicarbonate buffer, and was
incubated for 30 min. After washing in 150 μl of buffer A,
150 μl of alkylation buffer was added (50 mM iodoacetamide
in 100 mM ammonium bicarbonate). After two washes in buffer A for 5
min each, the gel plugs were diced/crushed into smaller cubes
followed by drying in Speed Vac for 10 min. Then, 30 μl (750
ng) of trypsin solution was added to the dried gel pieces and the
gel was swollen for 10 min at room temperature. Subsequently, 50
μl of 100 mM ammonium bicarbonate buffer was added until the
gel pieces were submerged. This was followed by incubation at 37°C
for 12 h and another 10 μl (250 ng) of sequencing grade
trypsin was added (Promega, Madison, WI, USA) and incubated for an
additional 2 h. The digest was removed and 150 μl of 60%
acetonitrile and 0.1% trifluroacetic acid (TFA) (buffer B) were
added followed by incubation for 30 min at 30°C. The solution was
removed and pooled with solution from the previous step. Extraction
was repeated and solutions were concentrated to a final volume of
15–20 μl.
Digest (2 μl) was separated on a reverse
phase column (Aquasil C18, 15 μm tip × 75 μm id × 5
cm Picofrit column, New Objectives, Woburn, MA, USA) using an
acetonitrile/1% acetic acid gradient system (5–75% acetonitrile
over 35 min followed by 95% acetonitrile wash for 5 min) at a flow
rate of 250 nl/min. Trypsin-cleaved peptides were directly
introduced into an ion-trap mass spectrometer equipped with a
nanospray source. The mass spectrometer was set for analyzing the
positive ions and obtaining a full MS scan and a collision-induced
dissociation spectrum on the most abundant ion from the full MS
scan (relative collision energy ∼30%). Dynamic exclusion was set to
collect 3 CID spectra on the most abundant ion and then exclude the
ion for 2 min. Data were searched against human and bovine
databases appended with human (P02768), bovine (P02769), horse
(P35747) and rabbit (P49065) albumin.
SDS-PAGE analysis
Purity of albumin after the different steps of
purification was analyzed by 12% SDS-PAGE. Protein samples (50
μl) were mixed with 10 μl 5X SDS-loading dye and
boiled for 2 min. Boiled samples were centrifuged at 10,000 × g for
10 sec. From each sample, 10 μl were loaded on SDS-PAGE.
Purity of CSA analyzed by HPLC
The homogeneity of the purified CSA was analyzed by
RP-HPLC. The mobile phase A consisted of 10% acetonitrile/90% water
containing 0.01% TFA and mobile phase B was 90% acetonitrile/10%
water containing 0.01% TFA. The column was 5 μm, 4.6×150 mm
waters symmetry C18 column and equilibrated with mobile phase A.
Purified CSA was bound on the column and eluted with a linear
gradient of mobile phase B.
Purity of CSA by capillary zone
electrophoresis
Polyvinyl alcohol (PVA)-coated CE capillary (75
μm, 56 cm in length) was fitted into high sensitivity
detection cell (Agilent Technologies). The capillary was
extensively washed with 20 mM triethanolamine buffer, pH 3.6.
Purified CSA (1 mg/ml) was diluted 10-fold in 20 mM triethanolamine
buffer, pH 3.6. Hydrodynamic pressure (100 mb) was applied for 10
sec to inject samples in the capillary and electrophoresis was
performed at 10 kV (positive polarity), 100 μA current and 6
W in 20 mM triethanolamine buffer, pH 3.6 for 30 min.
Results and Discussion
Purification of CSA
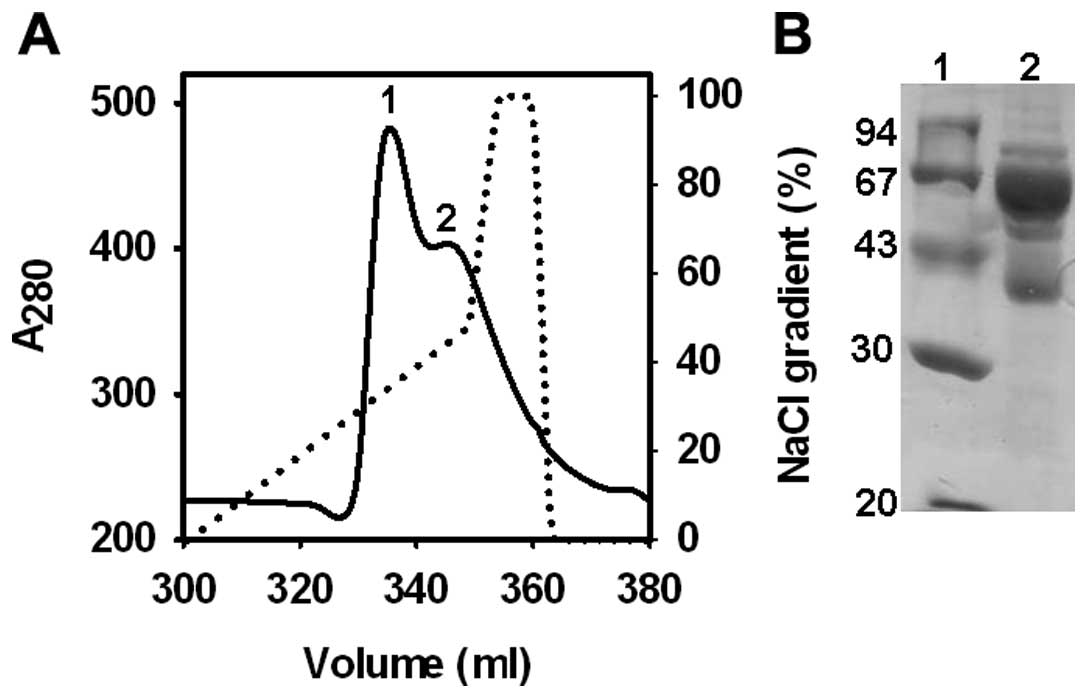
Centrifugation of EDTA-treated camel blood at 1,000
× g resulted in slightly reddish clear plasma. When diluted plasma
is passed through an equilibrated Blue-Sepharose column at a pH of
8.0, albumin binds with Cibacron Blue 3G on Blue-Sepharose 6 matrix
(34). Bound protein was eluted
with linear NaCl gradient (Fig.
1). It was observed that even when plasma containing 50–60 mg
albumin was applied on 20 ml Blue-Sepharose column (∼360 mg HSA
binding capacity), only the partial binding of CSA was observed.
Cibacron Blue-Sepharose efficiently is known to eliminate almost
all the albumin from human serum but not from bovine, sheep and
rabbit serum (35). At pH 8.0, HSA
binding on Blue-Sepharose was ∼90% while under identical conditions
only ∼50% BSA binds on Blue-Sepharose (35). The Kdiss of
defatted human, bovine, rabbit and sheep serum albumin for Cibacron
Blue were 19, 196, 150 and 150 μM, respectively (36). The binding of different mammalian
serum albumin on Blue-Sepharose was decreased as the degree of
pre-saturation of albumins with bilirubin, fatty acids and number
of carbon atoms in the chains of fatty acids increased (35,36).
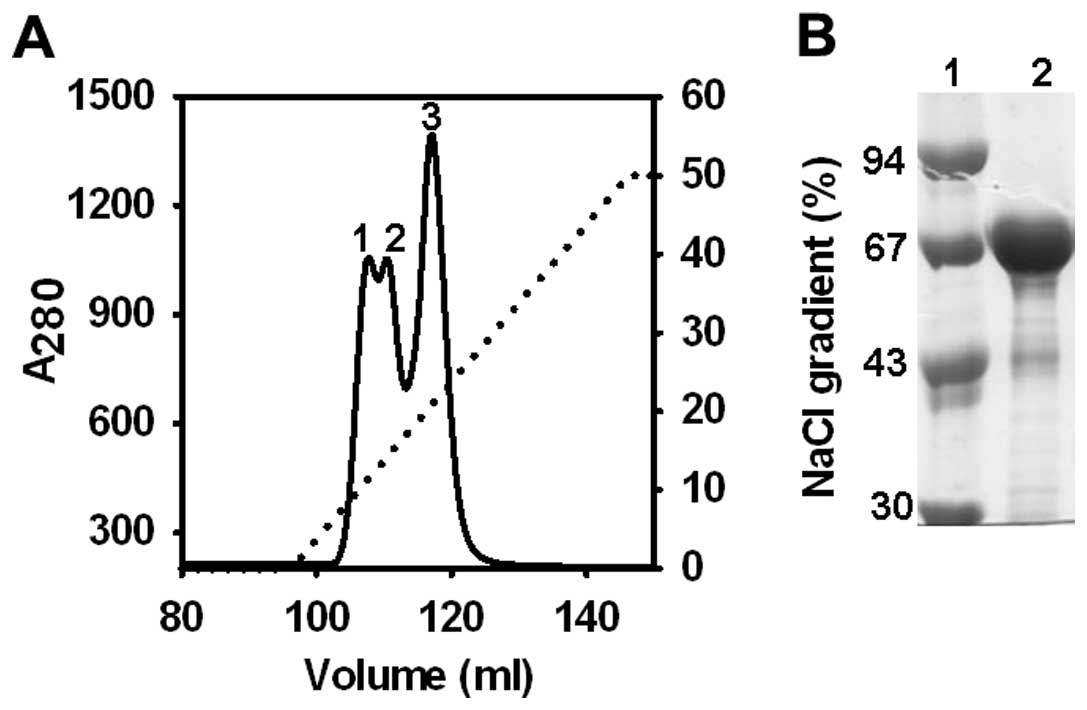
Following enrichment of CSA by Blue-Sepharose
affinity chromatography, partially purified CSA was further
purified by 5 ml HiTrap Q column (250 mg HSA binding capacity). At
pH 8.0, CSA was efficiently and tightly bound on HiTrap Q column.
No CSA was detected in the flow through and wash. Bound albumin was
eluted by the linear NaCl gradient and purity was analyzed by 12%
SDS-PAGE (Fig. 2). In the
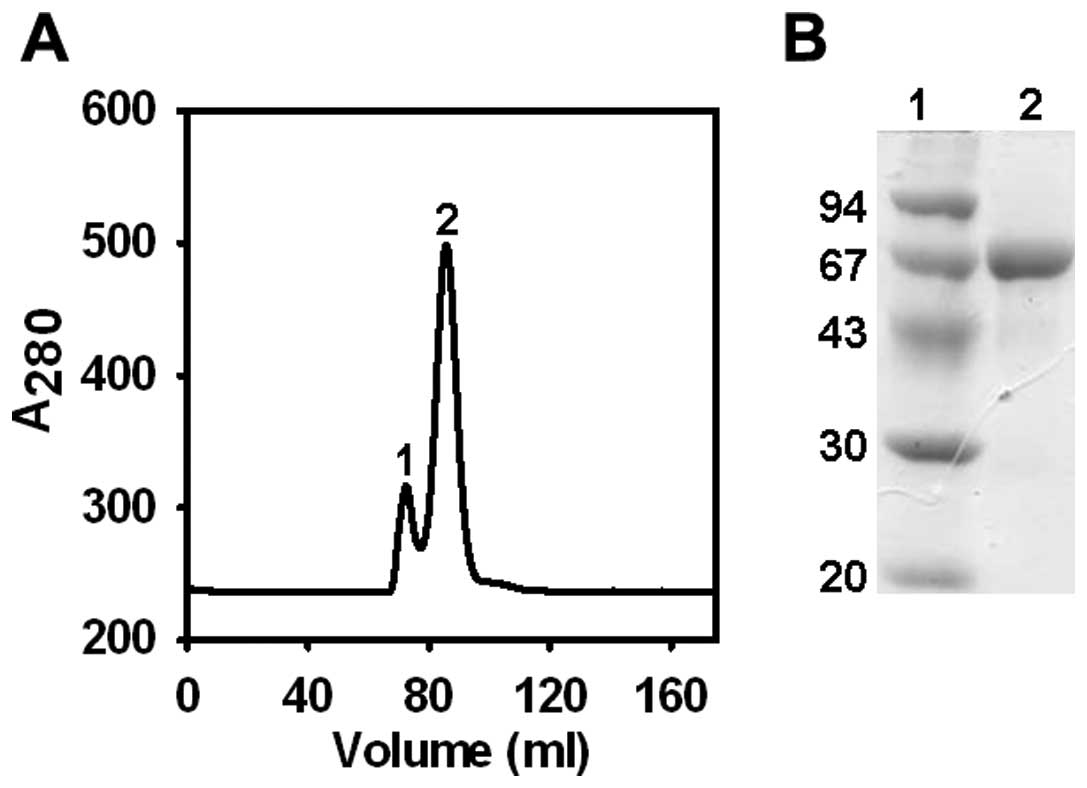
polishing step of purification, gel permeation chromatography was
performed using Sephacryl S-100 (Fig.
3). Using this protocol, highly pure CSA was obtained
reproducibly at ∼12 mg level from 1 ml of camel plasma in three
independent experiments.
Identification of purified protein by
mass spectrometry
SDS-PAGE of purified CSA was carried out and protein
band corresponding to the molecular weight of albumin (68 kDa) was
excised with a clean razor. Sliced gel was placed in a sterilized
Eppendorf tube and subjected to mass spectrometry. As shown in
Fig. 4, multiple fragments of CSA
were identified. Only the partial sequence of CSA (GenBank:
HM640019.1) was deposited in the gene bank (http://getentry.ddbj.nig.ac.jp/getentry/ddbj/HM640019?filetype=html).
When this partial sequence of CSA was aligned with the
corresponding sequence of human, bovine, rabbit and horse, 75, 79,
71 and 69% similarities were identified, respectively. Mass
spectrometric data revealed unique sequences (of 7–26 amino acids
in length) that correspond to the known partial sequence of CSA
(Fig. 4). These data also
indicated that identified peptides overlapped with the unique
sequence of CSA and confirmed that the purified protein was
CSA.
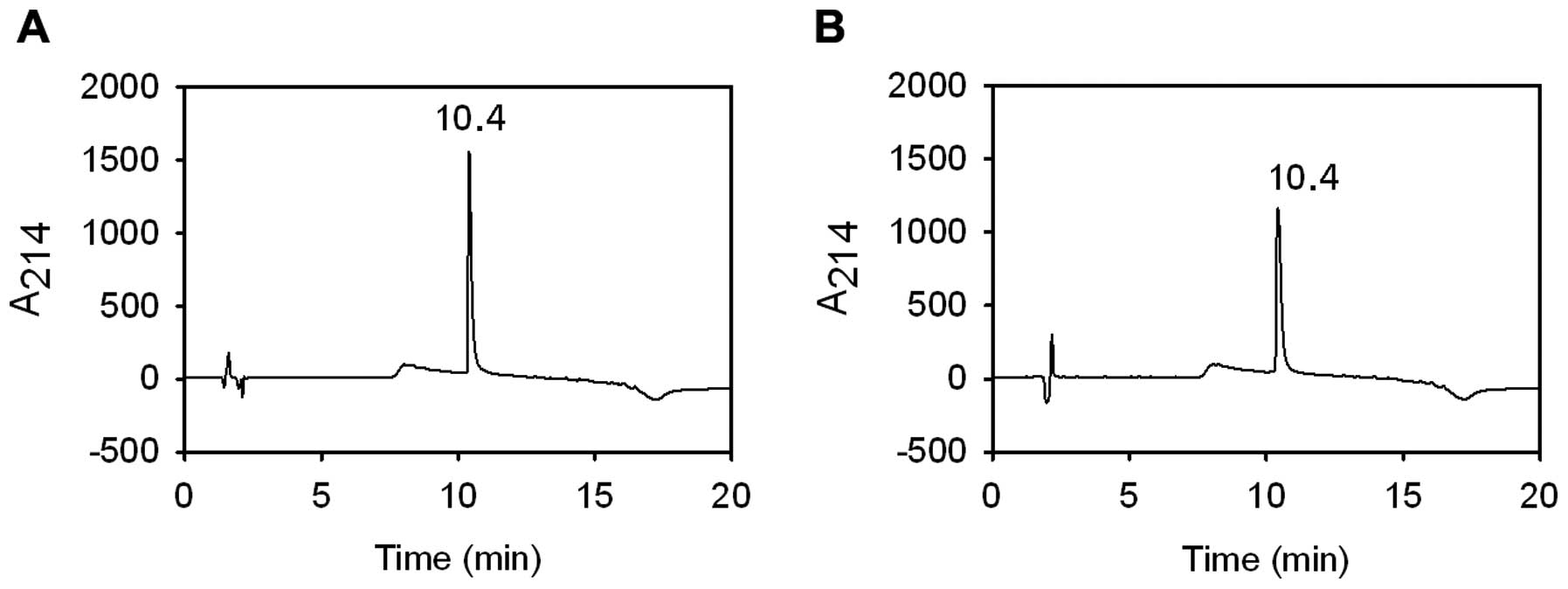
Analysis of CSA by RP-HPLC
The purity and homogeneity of the purified proteins
was analyzed by RP-HPLC, which separate the proteins based upon
surface hydrophobicity. To separate proteins, acetonitrile was
frequently used as an organic modifier and TFA as the ion-pairing
agent (37). When a linear
gradient of mobile phase B at room temperature was applied, CSA was
eluted in a single and sharp peak (Fig. 5). The peak retention time for
purified CSA was 10.4 min, which was the same as for commercial
BSA.
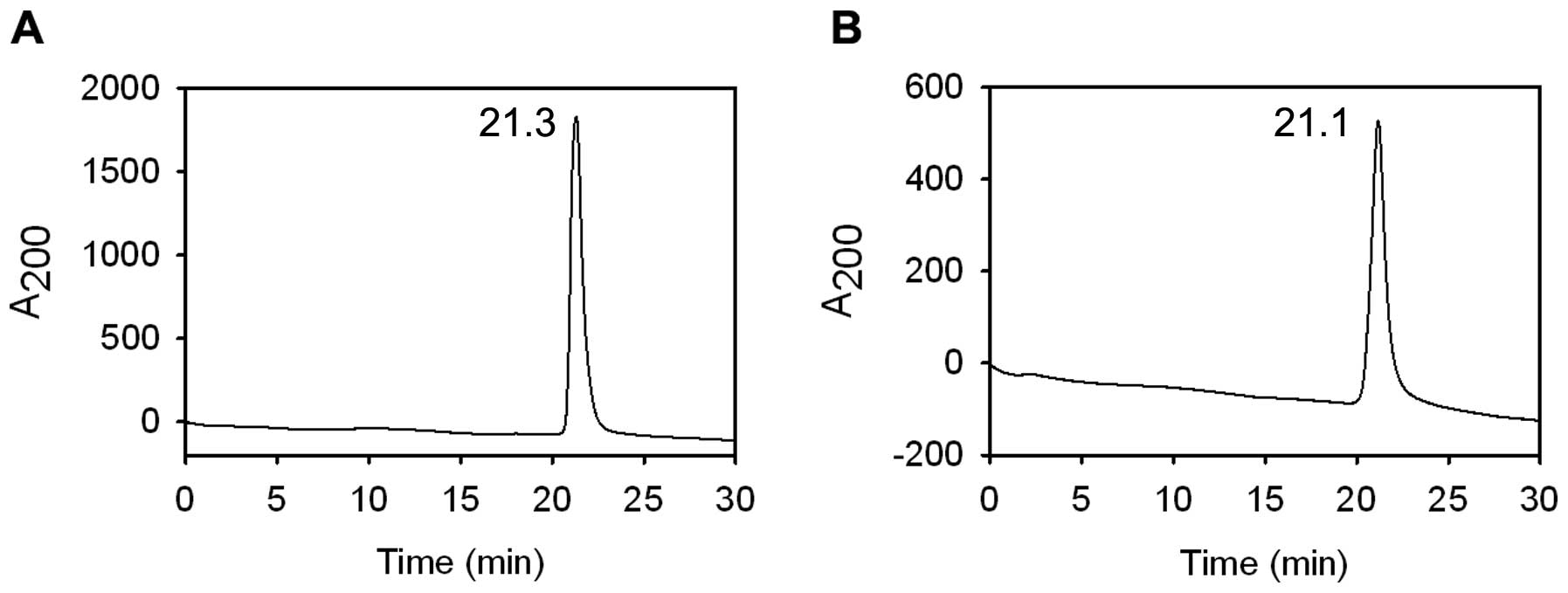
Analysis of CSA by capillary zone
electrophoresis
Capillary zone electrophoresis (CZE) separates
proteins based on their unique charge/mass ratio. CZE is a
complementary technique to RP-HPLC used for determination of the
homogeneity of proteins (38). The
capillary zone electropherogram of purified CSA showed a single and
symmetrical peak (Fig. 6A), which
confirmed the homogenous preparation of CSA. The retention time of
CSA and commercial BSA (Fig. 6B)
were very close (21.3 vs. 21.1 min).
Conclusion
Findings of SDS-PAGE, RP-HPLC and CZE indicate that
the current protocol for the isolation of CSA is efficient, with a
high yield and of pure grade. The mass spectrometric data confirmed
that purified protein is CSA. The described protocol for
purification of CSA is simple, efficient, and reproducible.
Furthermore, it yields a homogeneous product of CSA. In addition,
in this procedure mild conditions were applied to minimize the
denaturation of the CSA. In a number of biotechnological and
routine biochemical applications, HSA is replaced with BSA
(39). However, an investigation
into the structural, biochemical and immunological aspects of CSA,
potentially render it a viable alternative to HSA for certain
applications, such as cell culture, or indicate that it may possess
unique properties that may be beneficial in engineering albumin to
improve various biological and physical properties including ligand
binding specificity, affinity, and stability.
Abbreviations:
|
LMW
|
low molecular weight;
|
|
PVA
|
polyvinyl alcohol;
|
|
CE
|
capillary electrophoresis;
|
|
FPLC
|
fast protein liquid
chromatography;
|
|
CV
|
column volume;
|
|
TFA
|
trifluroacetic acid
|
Acknowledgements
This study was supported by King Saud
University, Deanship of Scientific Research, College of Science
Research Center.
References
|
1.
|
Lundsgaard-Hansen P: Physiology and
pathophysiology of colloid osmotic pressure and albumin metabolism.
Curr Stud Hematol Blood Transfus. 53:1–17. 1986.PubMed/NCBI
|
|
2.
|
Prajapati KD, Sharma SS and Roy N: Current
perspectives on potential role of albumin in neuroprotection. Rev
Neurosci. 22:355–363. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Georgiou HM, Rice GE and Baker MS:
Proteomic analysis of human plasma: failure of centrifugal
ultrafiltration to remove albumin and other high molecular weight
proteins. Proteomics. 1:1503–1506. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Jiang L, He L and Fountoulakis M:
Comparison of protein precipitation methods for sample preparation
prior to proteomic analysis. J Chromatogr A. 1023:317–320. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Olver CS, Webb TL, Long LJ, Scherman H and
Prenni JE: Comparison of methods for depletion of albumin and IgG
from equine serum. Vet Clin Pathol. 39:337–345. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Kim SH, Kim UK, Lee WS, et al:
Albumin-like protein is the major protein constituent of luminal
fluid in the human endolymphatic sac. PLoS One. 6:e216562011.
View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Wiig H, Reed RK and Tenstad O:
Interstitial fluid pressure, composition of interstitium, and
interstitial exclusion of albumin in hypothyroid rats. Am J Physiol
Heart Circ Physiol. 278:H1627–H1639. 2000.PubMed/NCBI
|
|
8.
|
Shaklai N, Garlick RL and Bunn HF:
Nonenzymatic glycosylation of human serum albumin alters its
conformation and function. J Biol Chem. 259:3812–3817.
1984.PubMed/NCBI
|
|
9.
|
Spector AA: Fatty acid binding to plasma
albumin. J Lipid Res. 16:165–179. 1975.PubMed/NCBI
|
|
10.
|
Ashbrook JD, Spector AA, Santos EC and
Fletcher JE: Long chain fatty acid binding to human plasma albumin.
J Biol Chem. 250:2333–2338. 1975.PubMed/NCBI
|
|
11.
|
Sargent TD, Jagodzinski LL, Yang M and
Bonner J: Fine structure and evolution of the rat serum albumin
gene. Mol Cell Biol. 1:871–883. 1981.
|
|
12.
|
Gibbs PE and Dugaiczyk A: Origin of
structural domains of the serum-albumin gene family and a predicted
structure of the gene for vitamin D-binding protein. Mol Biol Evol.
4:364–379. 1987.PubMed/NCBI
|
|
13.
|
Fanali G, Ascenzi P, Bernardi G and Fasano
M: Sequence analysis of serum albumins reveals the molecular
evolution of ligand recognition properties. J Biomol Struct Dyn.
29:691–701. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Bujacz A: Structures of bovine, equine and
leporine serum albumin. Acta Crystallogr D Biol Crystallogr.
68:1278–1289. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Luo Z, Shi X, Hu Q, Zhao B and Huang M:
Structural evidence of perfluorooctane sulfonate transport by human
serum albumin. Chem Res Toxicol. 25:990–992. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Curry S, Mandelkow H, Brick P and Franks
N: Crystal structure of human serum albumin complexed with fatty
acid reveals an asymmetric distribution of binding sites. Nat
Struct Biol. 5:827–835. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Sugio S, Kashima A, Mochizuki S, Noda M
and Kobayashi K: Crystal structure of human serum albumin at 2.5 A
resolution. Protein Eng. 12:439–446. 1999. View Article : Google Scholar
|
|
18.
|
Maruyama T, Katoh S, Nakajima M and
Nabetani H: Mechanism of bovine serum albumin aggregation during
ultrafiltration. Biotechnol Bioeng. 75:233–238. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Carter DC and Ho JX: Structure of serum
albumin. Adv Protein Chem. 45:153–203. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Phillips A, Shaper AG and Whincup PH:
Association between serum albumin and mortality from cardiovascular
disease, cancer, and other causes. Lancet. 2:1434–1436. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Nicholson JP, Wolmarans MR and Park GR:
The role of albumin in critical illness. Br J Anaesth. 85:599–610.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Singh-Zocchi M, Andreasen A and Zocchi G:
Osmotic pressure contribution of albumin to colloidal interactions.
Proc Natl Acad Sci USA. 96:6711–6715. 1999.PubMed/NCBI
|
|
23.
|
Stewart AJ, Blindauer CA, Berezenko S,
Sleep D and Sadler PJ: Interdomain zinc site on human albumin. Proc
Natl Acad Sci USA. 100:3701–3706. 2003. View Article : Google Scholar
|
|
24.
|
Hu W, Luo Q, Wu K, et al: The anticancer
drug cisplatin can cross-link the interdomain zinc site on human
albumin. Chem Commun (Camb). 47:6006–6008. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Vorum H: Reversible ligand binding to
human serum albumin. Theoretical and clinical aspects. Dan Med
Bull. 46:379–399. 1999.PubMed/NCBI
|
|
26.
|
Chuang VT, Kragh-Hansen U and Otagiri M:
Pharmaceutical strategies utilizing recombinant human serum
albumin. Pharm Res. 19:569–577. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Fasano M, Curry S, Terreno E, et al: The
extraordinary ligand binding properties of human serum albumin.
IUBMB Life. 57:787–796. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Buckow R, Wendorff J and Hemar Y:
Conjugation of bovine serum albumin and glucose under combined high
pressure and heat. J Agric Food Chem. 59:3915–3923. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Xiao J, Wu M, Kai G, Wang F, Cao H and Yu
X: ZnO-ZnS QDs interfacial heterostructure for drug and food
delivery application: enhancement of the binding affinities of
flavonoid aglycones to bovine serum albumin. Nanomedicine.
7:850–858. 2011. View Article : Google Scholar
|
|
30.
|
Yang M, Hoppmann S, Chen L and Cheng Z:
Human serum albumin conjugated biomolecules for cancer molecular
imaging. Curr Pharm Des. 18:1023–1031. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Zhi J, Teller SB, Satoh H, Koss-Twardy SG
and Luke DR: Influence of human serum albumin content in
formulations on the bioequivalency of interferon alfa-2a given by
subcutaneous injection in healthy male volunteers. J Clin
Pharmacol. 35:281–284. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Otsubo E and Takei T: Characterization of
the surface activity of a synthetic surfactant with albumin. Biol
Pharm Bull. 25:1519–1523. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Basrur V, Yang F, Kushimoto T, et al:
Proteomic analysis of early melanosomes: identification of novel
melanosomal proteins. J Proteome Res. 2:69–79. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Angal S and Dean PD: The effect of matrix
on the binding of albumin to immobilized Cibacron Blue. Biochem J.
167:301–303. 1977.PubMed/NCBI
|
|
35.
|
Leatherbarrow RJ and Dean PD: Studies on
the mechanism of binding of serum albumins to immobilized cibacron
blue F3G A. Biochem J. 189:27–34. 1980.PubMed/NCBI
|
|
36.
|
Metcalf EC, Crow B and Dean PD: The effect
of ligand presaturation on the interaction of serum albumins with
an immobilized Cibacron Blue 3G-A studied by affinity gel
electrophoresis. Biochem J. 199:465–472. 1981.PubMed/NCBI
|
|
37.
|
Malik A, Rudolph R and Sohling B: A novel
fusion protein system for the production of native human pepsinogen
in the bacterial periplasm. Protein Expr Purif. 47:662–671. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Girard M, Bietlot HP, Mousseau N, Cyr TD
and Ethier JC: Use of capillary electrophoresis for the
characterization of human serum albumin heterogeneity. Biomed
Chromatogr. 12:183–184. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Francis GL: Albumin and mammalian cell
culture: implications for biotechnology applications.
Cytotechnology. 62:1–16. 2010. View Article : Google Scholar : PubMed/NCBI
|