Introduction
Nonalcoholic fatty liver disease (NAFLD) is a
clinical pathological syndrome characterized mainly by hepatic
steatosis. The disease includes nonalcoholic fatty liver (NAFL),
nonalcoholic steatohepatitis and is associated with hepatic
cirrhosis and hepatocellular carcinoma. The pathogenesis of NAFLD
has yet to be elucidated, although the ‘two-hit’ hypothesis is
widely recognized. The initial hit refers to lipid metabolism and
insulin resistance, which induce fat accumulation and simple fatty
liver. The second hit comprises oxidative stress, inflammation and
certain other factors (1).
Oxidative stress is one of the key factors in the pathogenesis of
NAFLD. Numerous studies have demonstrated high oxidative stress
levels in patients with NAFLD (2,3). The
oxidative stress in NAFLD may be caused by hepatic triglyceride
deposition, increased levels of free fatty acids and mitochondrial
dysfunction (4). Excess fatty acid
oxidation leads to hepatic oxidative stress, a reduced antioxidant
defense ability and mitochondrial dysfunction, which consequently
increases the expression of inflammatory cytokines, such as tumor
necrosis factor-α (TNF-α). Thus, lipid peroxidation on the
mitochondrial membrane is aggravated, leading to mitochondrial
dysfunction and aggravating the liver toxicity of inflammatory
factors and apoptosis. Narasimhan et al(5) studied the changes in the oxidative
stress indices in patients with NAFLD with or without type 2
diabetes mellitus. The oxidative stress levels in patients with
NAFLD and without impaired glucose tolerance, even insulin
resistance, was observed to be significantly increased. Insulin
resistance does not initially appear in patients with NAFLD,
indicating that the function of oxidative stress is independent in
the pathogenesis of NAFLD. Therefore, regulating hepatic
inflammation and the oxidative stress level may have certain
therapeutic effects in NAFLD.
Glutamine is a type of free amino acid that accounts
for ~60% of free amino acids in the body. It is a conditionally
essential amino acid and has an important function in life
activities. Under physiological conditions, glutamine is capable of
being synthesized by the body. However, under pathological
conditions, the increasing demands from the body cause a relative
lack of glutamine, leading to a disturbance in energy metabolism
and immune suppression. In recent years, studies have shown that
glutamine is able to improve hepatic ischemia-reperfusion (6,7) and
alcohol-induced liver injury (8).
Furthermore, glutamine, which is considered to be an important
immune nutrient, is able to improve gut-derived endotoxemia
(9), has a certain resistance to
oxidative stress (10), reduces
the release of inflammatory cytokines (11) and regulates immunoreaction
(12). In the present study, rats
with NAFLD, induced by a high-fat diet, were treated with an
intervention of glutamine in the early experimental periods. The
oxidative stress levels in the rat livers were observed at
different time-points and the regulatory effect of glutamine on
oxidative stress in NAFLD was investigated.
Materials and methods
Animals
A total of 36 healthy male Sprague-Dawley (SD) rats
(160±10 g) were housed in individual stainless steel cages in an
animal room maintained at 22±2°C with 50–70% humidity and a 12-h
light/dark cycle. This study was performed in strict accordance
with the recommendations in the Guide for the Care and Use of
Laboratory Animals of the National Institutes of Health (the 8th
edition, 2011). The animal use protocol was reviewed and approved
by the Institutional Animal Care and Use Committee (IACUC) of
Fujian Provincial Hospital (Fuzhou, China).
Study protocol
The recipe for the high-fat diet and the main
reagents were as follows: 88% normal diet, 10% lard and 2%
cholesterol (Shanghai Hayes Lakes Experimental Animals Co., Ltd.,
Shanghai, China). Following 1 week of acclimation, the rats were
divided into six groups by a random number table according to the
weight in each group (n=6). The diets for the different groups were
as follows: Normal control groups (C1) and (C2), normal diet plus
saline gavage (1 ml/day); model groups (M1) and (M2), high-fat diet
plus saline gavage (1 ml/day); and glutamine treatment groups (T1)
and (T2), high-fat diet plus glutamine gavage (1 g/kg/day). The
rats were anesthetized and sacrificed on weeks 8 (C1, M1 and T1)
and 12 (C2, M2 and T2) and the liver tissues were rapidly excised.
The left lobe was stored in liquid nitrogen for further analysis,
while the right lobe was fixed in 10% neutral formalin for the
preparation of paraffin-embedded sections.
Analysis of weight and liver index
The experimental animals were weighed every weekend.
Once the rats had been sacrificed by cervical dislocation, the
livers were obtained and weighed. The liver index was calculated
using the following formula: Liver index = (liver wet weight/body
weight) ×100%.
Analysis of glutathione (GSH), TNF-α and
malondialdehyde (MDA)
Liver tissues (0.5 g) were washed with normal saline
at 4°C to remove the blood. Following this, the tissue samples were
placed in a glass homogenizer tube and 5 ml normal saline was added
at 4°C. A 10% homogenate was then prepared. Following
centrifugation at 4°C and 3,000 rpm for 15 min, the supernatant was
isolated for use. The GSH and TNF-α concentrations were measured
using enzyme-linked immunosorbent assay kits (R&D Systems,
Minneapolis, MN, USA), and the assays of the samples and standards
were simultaneously conducted, in accordance with the assay kits’
instructions. The optical density (OD) was read at 450 nm using a
microplate reader (TU-1221; Beijing Purkinje General Instrument
Co., Ltd., Beijing, China) and the concentrations of GSH and TNF-α
were calculated using the OD formula (Concentration =
[(ODtest − ODtest
blank)/(ODstandard − ODstandard blank)]
× Concentrationstandard/Molecular weight). The protein
concentration in each liver homogenate sample was measured using a
Coomassie blue protein assay kit (Dingguo Biotechnology, Beijing,
China). The MDA absorbance of the liver homogenates was measured
using thiobarbituric acid-reactive substances with MDA kits
(Nanjing Jiancheng Biotechnology Co., Ltd., Nanjing, China).
Histological examinations of the
liver
Liver tissues were fixed in 10% formaldehyde and
embedded in paraffin. The paraffin sections were subsequently cut
and processed for histological examination using hematoxylin and
eosin (H&E) and immunohistochemistry. The immunohistochemical
staining was performed using nuclear factor-κB protein 65 (NF-κB
p65) monoclonal antibody (Zhongshan Jinqiao Biotechnology Co.,
Ltd., Beijing, China) and the histological evaluation was performed
by a pathologist who was blinded to the treatment groups. The
H&E staining was graded as follows: Normal (no steatotic
cells); mild (5–33% steatotic cells); moderate (34–66% steatotic
cells); and severe (>66%steatotic cells). For the
immunohistochemical analysis, 10 fields under high magnification
(x400) were taken for observation in each slice. Scoring was
performed according to the degree of nuclear stain, based on
staining intensity and staining range. The staining intensity was
scored as follows: no stain, 0; light stain, 1; medium stain, 2;
and deep stain, 3; while staining range was scored according to the
following criteria: <5%, 0; 5–25%, 1; 26–50%, 2; 51–75%, 3; and
>75%, 4. The combined scores were then assessed and classified
as follows: <2, negative (−); 2–3, positive (+); 4–5, moderately
positive (++); and 6–7, strongly positive (+++).
Statistical analysis
SPSS 16.0 statistical software (SPSS, Inc., Chicago,
IL, USA) was used for the data analyses. Measured data are
expressed as the mean ± standard deviation. Non-parametric tests
were used for the ranked data with non-normal distribution.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Weight and liver index of rats
As shown in Table
I, the increases in body weight in the model and glutamine
treatment groups were significantly higher than those in the
control group (P<0.05); however, there were no significant
differences between the model and glutamine treatment groups
(P>0.05). The liver indices in the model and glutamine treatment
groups were significantly higher than those in the control group
(P<0.05), and the indices in the glutamine treatment group were
significantly lower than those in the model group (P<0.05).
 | Table IBody weight gain and liver indices of
rats. |
Table I
Body weight gain and liver indices of
rats.
| Group | Body weight gain
(g) | Liver index (liver
wet weight/body weight, %) |
|---|
| C1 | 276.33±19.36 | 2.47±0.16 |
| M1 | 322.50±16.40a | 3.49±0.23a |
| T1 | 299.00±15.18a | 3.25±0.19a,b |
| C2 | 332.50±21.87 | 2.50±0.14 |
| M2 | 379.00±26.45b,c | 3.86±0.19b,c |
| T2 | 375.68±14.40c | 3.50±0.21c,d |
Measurement of GSH, TNF-α and MDA levels
in the liver
Table II shows
that the TNF-α and MDA levels in the liver homogenate in the model
group were higher than those in the control group (P<0.05),
whereas the levels in the glutamine treatment group were decreased
at each time-point compared with those in the model group
(P<0.05). The GSH level in the model group was significantly
lower than that in the control group at the same time-point
(P<0.05), whereas, the level was significantly higher in the
glutamine treatment group than that in the model group
(P<0.05).
| Table IIGSH, TNF-α and MDA levels in rats. |
Table II
GSH, TNF-α and MDA levels in rats.
| Group | GSH (ng/ml) | TNF-α (ng/l) | MDA (nmol/mg
prot) |
|---|
| C1 | 0.78±0.13 | 119.39±34.81 | 0.53±0.09 |
| M1 | 0.57±0.06a | 343.83±20.61a | 1.11±0.10a |
| T1 | 0.71±0.06b | 275.93±34.12b | 0.90±0.06b |
| C2 | 0.67±0.05 | 162.68±13.50 | 0.51±0.04 |
| M2 | 0.48±0.07b,c | 427.83±53.59b,c | 1.45±0.15b,c |
| T2 | 0.59±0.05d | 308.64±25.62d | 0.98±0.22d |
Macroscopic examination of the liver
The livers in the control group were bright red and
of a normal size, with sharp edges, a smooth surface and non-greasy
sections (Fig. 1A and D). In the
model group, the liver volume had increased significantly, and the
livers were light-yellow with blunt and thick edges, rough
surfaces, and greasy sections (Fig. 1B
and E). With prolonged modeling time, the livers in the
glutamine treatment group were observed to be markedly improved
compared with those in the model group (Fig. 1C and F).
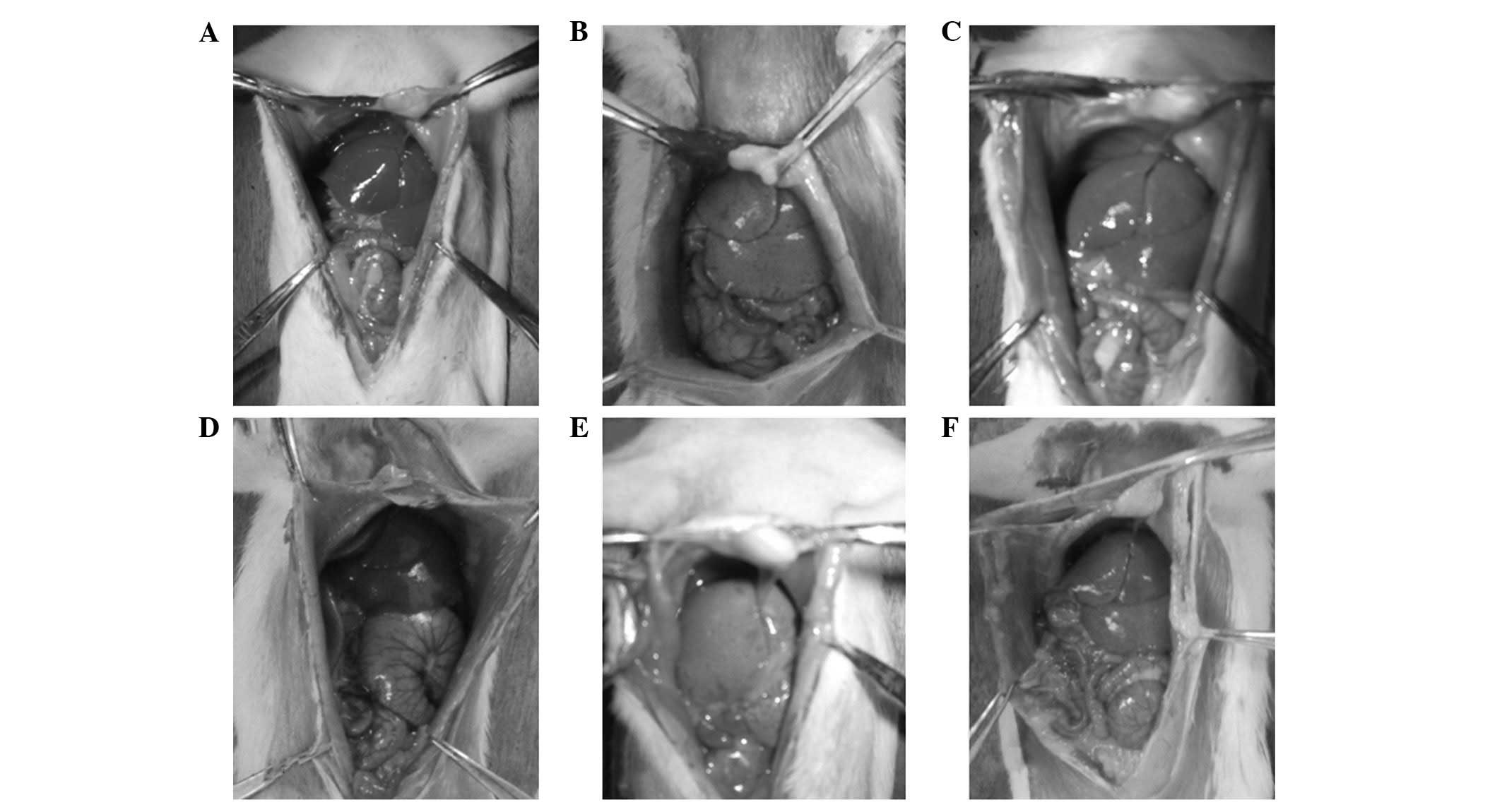 | Figure 1Macroscopic examination of the liver
in the (A) C1, (B) M1, (C) T1, (D) C2, (E) M2 and (F) T2 groups.
C1, control group at 8 weeks; C2, control group at 12 weeks; M1,
nonalcoholic fatty liver disease (NAFLD) model group at 8 weeks;
M2, NAFLD model group at 12 weeks; T1, glutamine-treated rats with
NAFLD at 8 weeks; T2, glutamine-treated rats with NAFLD at 12
weeks. |
Hepatic histopathology
On week 8, the M1 model group mainly showed
light-moderate steatosis. On week 12, the hepatocyte steatosis in
the M2 model group had intensified to severe steatosis, with no
significant changes in fibrosis. The degree of steatosis was
significantly higher than that of the control group (P<0.05).
With regard to the pathological changes, the livers in the
glutamine treatment group were observed to be significantly
improved compared with the model group under the same period
(P<0.05), and the hepatic steatosis was moderate (Fig. 2).
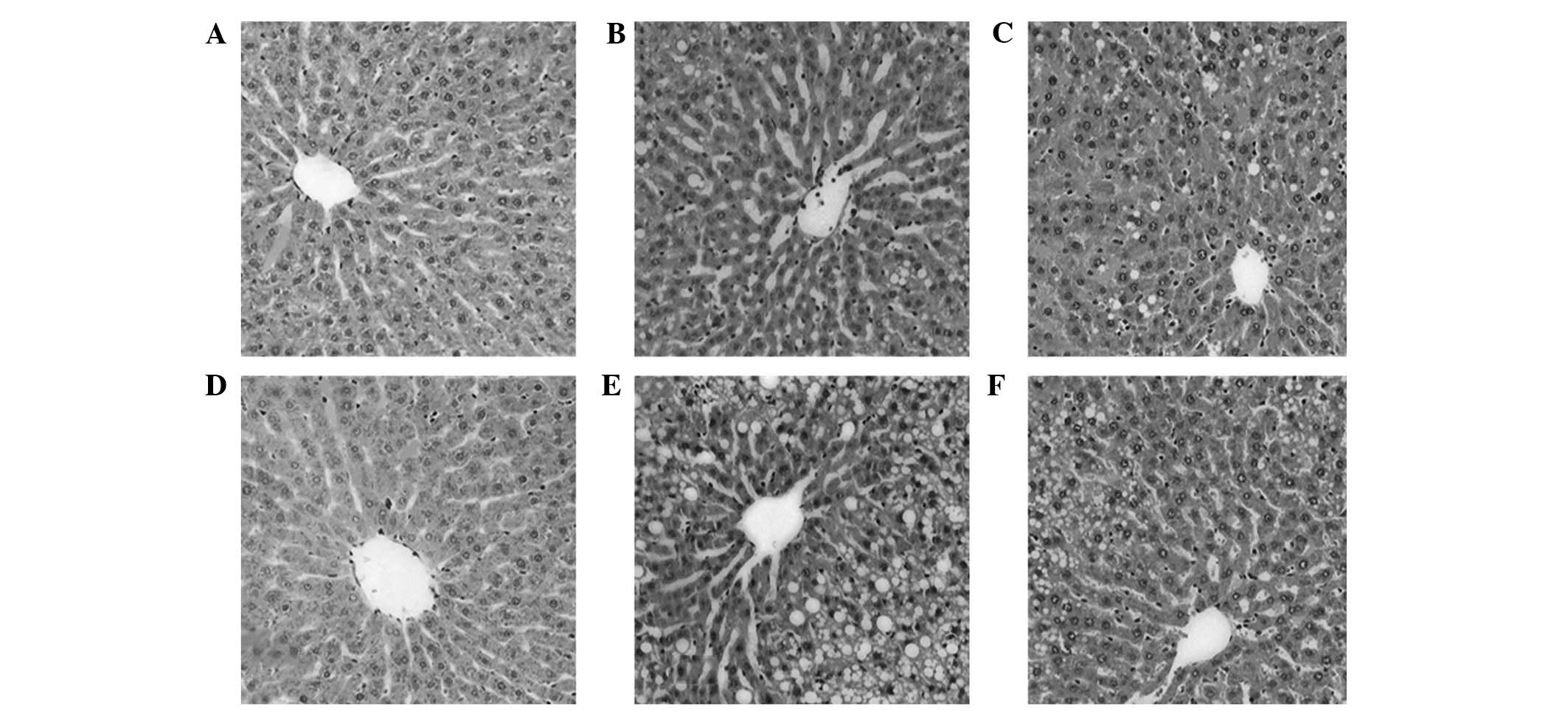 | Figure 2Pathological changes of rat liver
tissues in each group (hematoxylin and eosin staining;
magnification, ×100). (A) C1, (B) M1, (C) T1, (D) C2, (E) M2 and
(F) T2 groups. C1, control group at 8 weeks; C2, control group at
12 weeks; M1, nonalcoholic fatty liver disease (NAFLD) model group
at 8 weeks; M2, NAFLD model group at 12 weeks; T1,
glutamine-treated rats with NAFLD at 8 weeks; T2, glutamine-treated
rats with NAFLD at 12 weeks. |
Immunohistochemistry
NF-κB p65-positive cells in the rat liver tissues
were yellow-brown or dark-brown and were distributed in the nucleus
with varied intensities. On week 8, the cells of the M1 model group
were significantly more positive for NF-κB p65 than those of the C1
group in the same period (P<0.05). With prolonged modeling time,
the NF-κB p65-positivity of the cells in the M2 group further
increased (P<0.05). The number of NF-κB p65-positive cells in
the T groups was significantly lower than that of the M groups
(P<0.05; Fig. 3).
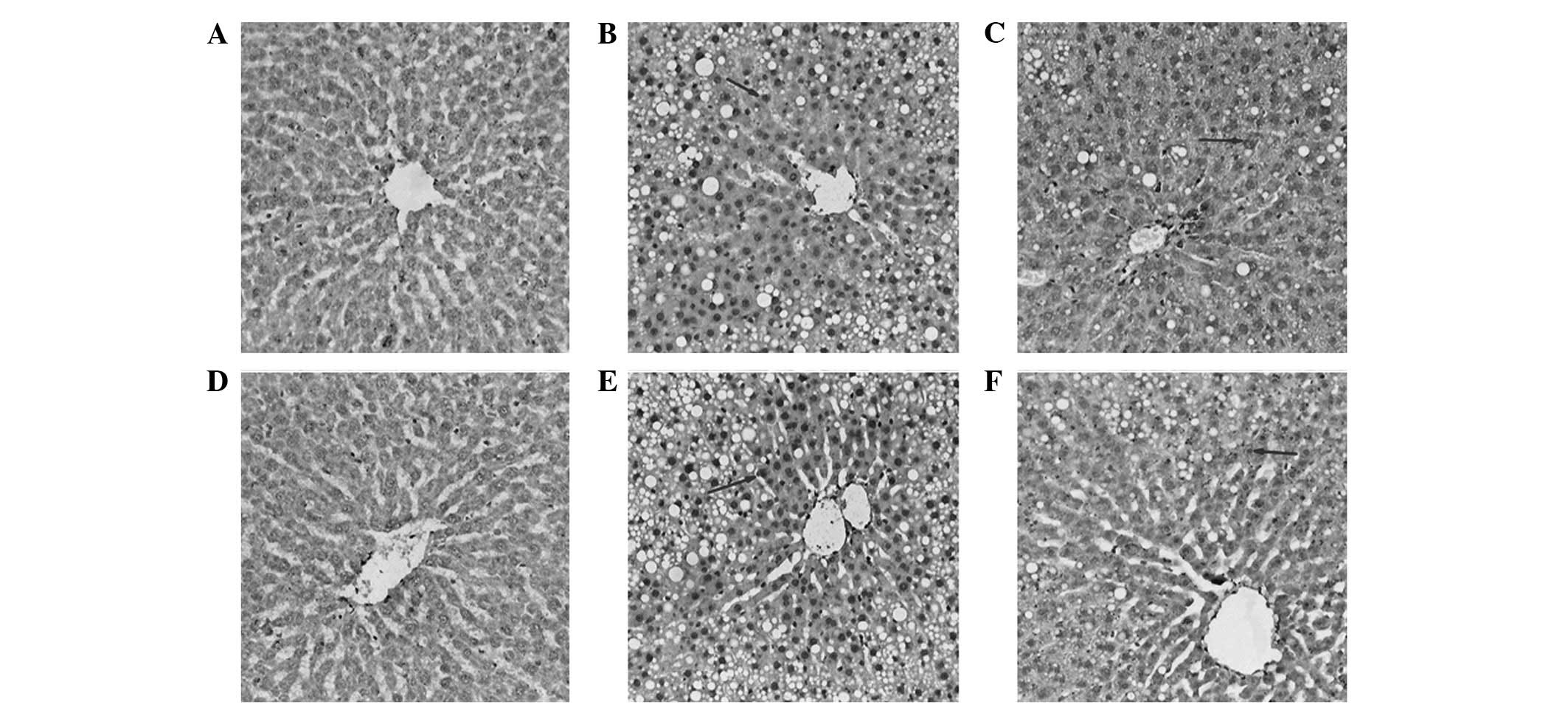 | Figure 3Expression of nuclear factor-κB
(NF-κB) in rat liver tissues (immunohistochemical staining;
magnification, ×100). (A) C1, (B) M1, (C) T1, (D) C2, (E) M2 and
(F) T2 groups. C1, control group at 8 weeks; C2, control group at
12 weeks; M1, nonalcoholic fatty liver disease (NAFLD) model group
at 8 weeks; M2, NAFLD model group at 12 weeks; T1,
glutamine-treated rats with NAFLD at 8 weeks; T2, glutamine-treated
rats with NAFLD at 12 weeks. |
Discussion
The global incidence of NAFLD has increased rapidly
in recent years, along with the development of obesity and
metabolic syndrome. In clinical practice, the prognosis for NAFLD
has changed through lifestyle control. However, the long-term
effects of the disease remain a problem. Thus, novel treatments are
required to improve hepatic steatosis and prevent the progression
of NAFLD. Glutamine is the most abundant amino acid in the blood
circulation and free amino acid pool. The product of glutamine,
GSH, is an important antioxidant that is capable of blocking
oxidative damage, while glutamine is able to reduce the release of
pro-inflammatory factors (13).
Thus, glutamine contributes to the maintenance of a stable
environment and affects the immune response and oxidative stress to
protect the organs.
Oxidative stress in NAFLD is generated from free
radicals and is a type of pathogenesis of NAFLD. Free radicals may
damage the spiral structure of DNA and affect its transcription and
replication, leading to necrocytosis. Free radicals are also able
to promote the production of pro-inflammatory mediators, such as
cytokines, and reflect the degree of oxidative stress injury
through GSH and MDA indices (14).
In the present study, the liver MDA levels in experimental model
were significantly higher than those in control group at the same
time-point (P<0.05), while the liver GSH levels in experimental
model were significantly lower than those in control group
(P<0.05). These changes were aggravated with prolonged modeling
time. With the extension of modeling time, the degree of steatosis
in the model group was aggravated gradually. These results showed
that the decreased antioxidant capacity and increased oxidative
stress levels were consistent with the aggravated pathological
changes in the liver. Glutamine treatment was able to attenuate the
changes in the GSH and MDA levels to retard the pathological
changes in the liver tissues. Endogenous glutamine is not able to
sufficiently meet the demand for GSH in the body under various
damaging conditions, such as stress and inflammation. Thus,
exogenous glutamine is added to meet the needs of the body. Yu
et al(15) demonstrated
that intravenous glutamine was able to improve GSH levels in the
serum and liver tissues, reducing chemotherapy-induced liver damage
in rats. Furthermore, Peng et al(8) indicated that in chronic ethanol-fed
rats, the GSH levels in rats treated orally with glutamine were
higher than those in the model group. The addition of glutamine to
the diet maintains the GSH concentration in the liver and reduces
alcohol-induced liver inflammation and oxidative stress levels.
Tihan et al(16)
established a rat model with reperfusion to induce oxidative injury
through abdominal hypertension. Pretreatment with a glutamine
gavage for seven days increased the serum GSH levels; however, it
reduced MDA and myeloperoxidase levels, and reperfusion-induced
oxidative damage. Our results are consistent with the results of
other studies. Decreased GSH levels have also been observed in the
livers of experimental animals and patients with NAFLD (17,18).
Patients with NAFLD experience a certain degree of
obesity and metabolic abnormalities. Excess nutrients cause
systemic low-grade inflammation. The activation of the innate
immune system has an important function in the transition process
from steatosis to NAFLD, which includes a number of inflammatory
factors, such as TNF-α. TNF-α has a key function in the cytokine
network associated with liver injury. Studies have shown that the
serum TNF-α levels and the mRNA expression of TNF-α in patients
with NAFLD are increased (19,20).
In the present study, the TNF-α levels in the liver homogenates of
the model group were significantly higher compared with the control
group (P<0.05); however, the levels in the glutamine treatment
group were lower than those in the model group (P<0.05), which
was consistent with previous studies. Tsai et al(21) fed diabetic SD rats with 1 kg forage
supplemented with 41.7 g glutamine and observed that the mRNA
expression of serum inflammatory cytokines, such as TNF-α,
interleukin (IL)-6 and transforming growth factor-β, was
significantly decreased in the glutamine intervention group
compared with the diabetic rats fed a normal diet. Glutamine has
also been shown to enhance heat shock protein (HSP) expression in
the gut and plasma, as well as to reduce the level of inflammatory
cytokines, such as IL-6 and IL-8, thus protecting against
inflammatory injury (22). Pai
et al(23) hypothesized
that a diet with glutamine was likely to maintain serum glutamine
and reduce leukocyte function-associated antigen-1 and macrophage
antigen-1 expression in rats with chronic arsenic exposure. One of
the links between leukocytes and endothelial cells is the adhesion
molecule, which is able to promote inflammation. A study showed
that glutamine supplementation was able to reduce the release of a
number of adhesion molecules, including intercellular adhesion
molecule 1 (ICAM-1) and vascular cell adhesion molecule 1 (VCAM-1)
(11). The activation of
inflammatory cells and inflammatory cytokines aggravates
mitochondrial dysfunction to form reactive oxygen species (ROS)
that, in turn, promote inflammation and aggravate NAFLD.
A close correlation exists between low-grade
inflammation in NAFLD and NF-κB activity, since NF-κB is an
important transcription factor of pro-inflammatory genes and NF-κB
has an important function in liver tissue inflammation and
oxidative stress (24). Decreased
NF-κB activation is able to mediate the reduced transcription of
downstream inflammatory factors, thereby decreasing the injury or
inflammation of the liver and reducing ROS generation to alleviate
the oxidative stress of the liver cells. However, the reduced ROS
and inflammatory factor, as the second messenger, activate NF-κB to
form a vicious circle. The abundant activation of NF-κB has been
observed in obese patients and methionine choline-deficient
diet-induced rats with NAFLD (25,26).
NF-κB may be activated in an oxidation-dependent manner. The
inhibition of its activation prevents the generation of
pro-inflammatory cytokines, such as TNF-α. TNF-α is an activation
agent of the inhibitory factor NF-κB kinase (IKK). IKK activates
NF-κB by the phosphorylation of NF-κB inhibitory factor, which
promotes TNF-α transcription (27). Thus, an inflammatory positive
feedback loop forms. The inhibition of NF-κB activation reduces the
inflammatory and oxidative stress levels. Singleton et
al(28) demonstrated that oral
glutamine reduced the secretion of inflammatory cytokines, such as
TNF-α, IL-6 and IL-18, in the lung tissues of rats with acute
respiratory distress syndrome (ARDS), and protected lung tissue by
the reducing the activation of NF-κB. Furthermore, Singleton and
Wischmeyer (29) revealed that the
addition of glutamine was able to significantly reduce NF-κB
activity and the expression of inflammatory cytokines in an HSP
gene knockout mouse. In addition, Huang et al(30) observed that glutamine was able to
reduce the release of inflammatory factor IL-8 induced by
lipopolysaccharide. In Caco-2 cells, an increased NF-κB protein
expression was observed, indicating that the regulation of
glutamine on inflammatory factors was not through the NF-κB
pathway. This inconsistent result may be due to the complex nature
of the inflammatory pathway of inflammatory factors. The
sensitivities of modulation in different pathways are different.
Elevated TNF-α levels rapidly activate NF-κB transcription
(31). In the present study, the
elevated oxidative stress and inflammatory factor levels promoted
NF-κB activation. However, the experimental sample size in this
study was small, which may mean that the results were not entirely
representative. Thus, the conclusions made in the current study
require verification in larger samples. In addition, the serum
concentration of glutamine was not analyzed. The measurement of
glutamine concentration enables the analysis of glutamine shortages
in NAFLD and glutamine concentration changes in the circulation
caused by intestinal glutamine supplementation. Other amino acids,
including arginine and glycine, may be selected as a control to
investigate whether glutamine alone has a protective effect in
NAFLD.
In conclusion, elevated levels of oxidative stress
in the liver tissues of a high-fat diet-induced rat model of NAFLD
are reduced by early intervention with glutamine, which may be
accomplished by the inhibition of the NF-κB pathway. To date, few
studies have focused on the correlation between the NF-κB pathway
and the mechanism underlying NAFLD. Further experiments, such as
hepatocyte cultivation in vitro or clinical studies, are
required to fully elucidate the correlation between the NF-κB
pathway and the mechanism of NAFLD, and to provide additional
evidence regarding the protective effects of glutamine in
NAFLD.
References
|
1
|
Day CP: Non-alcoholic fatty liver disease:
current concepts and management strategies. Clin Med. 6:19–25.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang S, Zhu H, Li Y, et al: Mitochondrial
adaptations to obesity related oxidant stress. Arch Biochem
Biophys. 378:259–268. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Madan K, Bhardwaj P, Thareja S, Gupta SD
and Saraya A: Oxidant stress and antioxidant status among patients
with nonalcoholic fatty liver disease (NAFLD). J Clin
Gastroenterol. 40:930–935. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Browning JD and Horton JD: Molecular
mediators of hepatic steatosis and liver injury. J Clin Invest.
114:147–152. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Narasimhan S, Gokulakrishnan K,
Sampathkumar R, et al: Oxidative stress is independently associated
with non-alcoholic fatty liver disease (NAFLD) in subjects with and
without type 2 diabetes. Clin Biochem. 43:815–821. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stangl R, Szijártó A, Ónody P, et al:
Reduction of liver ischemia-reperfusion injury via glutamine
pretreatment. J Surg Res. 166:95–103. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang WX, Zhou LF, Zhang L, et al:
Protective effects of glutamine preconditioning on
ischemia-reperfusion injury in rats. Hepatobiliary Pancreat Dis
Int. 10:78–82. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Peng HC, Chen YL, Chen JR, et al: Effects
of glutamine administration on inflammatory responses in chronic
ethanol-fed rats. J Nutr Biochem. 22:282–288. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kessel A, Toubi E, Pavlotzky E, et al:
Treatment with glutamine is associated with down-regulation of
Toll-like receptor-4 and myeloid differentiation factor 88
expression and decrease in intestinal mucosal injury caused by
lipopolysaccharide endotoxaemia in a rat. Clin Exp Immunol.
151:341–347. 2008. View Article : Google Scholar
|
|
10
|
Kul M, Vurucu S, Demirkaya E, et al:
Enteral glutamine and/or arginine supplementation have favorable
effects on oxidative stress parameters in neonatal rat intestine. J
Pediatr Gastroenterol Nutr. 49:85–89. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tsai PH, Liu JJ, Chiu WC, Pai MH and Yeh
SL: Effects of dietary glutamine on adhesion molecule expression
and oxidative stress in mice with streptozotocin-induced type 1
diabetes. Clin Nutr. 30:124–129. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
El-Sheikh NM and Khalil FA: L-arginine and
L-glutamine as immunonutrients and modulating agents for oxidative
stress and toxicity induced by sodium nitrite in rats. Food Chem
Toxicol. 49:758–762. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim H: Glutamine as an immunonutrient.
Yonsei Med J. 52:892–897. 2011. View Article : Google Scholar
|
|
14
|
Adams LA and Angulo P: Treatment of
non-alcoholic fatty liver disease. Postgrad Med J. 82:315–322.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu JC, Jiang ZM and Li DM: Glutamine: a
precursor of glutathione and its effect on liver. World J
Gastroenterol. 5:143–146. 1999.PubMed/NCBI
|
|
16
|
Tihan DN, Erbil Y, Seven R, et al: The
effect of glutamine on oxidative damage in an experimental
abdominal compartment syndrome model in rats. Ulus Trauma Acil
Cerrahi Derg. 17:1–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Park HJ, DiNatale DA, Chung MY, et al:
Green tea extract attenuates hepatic steatosis by decreasing
adipose lipogenesis and enhancing hepatic antioxidant defenses in
ob/ob mice. J Nutr Biochem. 22:393–400. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hardwick RN, Fisher CD, Canet MJ, Lake AD
and Cherrington NJ: Diversity in antioxidant response enzymes in
progressive stages of human nonalcoholic fatty liver disease. Drug
Metab Dispos. 38:2293–2301. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee YM, Sutedja DS, Wai CT, et al: A
randomized controlled pilot study of Pentoxifylline in patients
with non-alcoholic steatohepatitis (NASH). Hepatol Int. 2:196–201.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tilg H: The role of cytokines in
non-alcoholic fatty liver disease. Dig Dis. 28:179–185. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tsai PH, Yeh CL, Liu JJ, Chiu WC and Yeh
SL: Effects of dietary glutamine on inflammatory mediator gene
expressions in rats with streptozotocin-induced diabetes.
Nutrition. 28:288–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jang HJ, Kwak JH, Cho EY, et al: Glutamine
induces heat-shock protein-70 and glutathione expression and
attenuates ischemic damage in rat islets. Transplant Proc.
40:2581–2584. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pai MH, Chien YW, Tsai YH, Hu YM and Yeh
SL: Glutamine reduces the expression of leukocyte integrins
leukocyte function-associated antigen-1 and macrophage antigen-1 in
mice exposed to arsenic. Nutr Res. 28:544–549. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kaplowitz N and Tsukamoto H: Oxidative
stress and liver disease. Prog Liver Dis. 14:131–159.
1996.PubMed/NCBI
|
|
25
|
Videla LA, Tapia G, Rodrigo R, et al:
Liver NF-kappa B and AP-1 DNA binding in obese patients. Obesity
(Silver Spring). 17:973–979. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dela Peña A, Leclercq I, Field J, George
J, Jones B and Farrell G: NF-kappaB activation, rather than TNF,
mediates hepatic inflammation in a murine dietary model of
steatohepatitis. Gastroenterology. 129:1663–1674. 2005.PubMed/NCBI
|
|
27
|
Cai D, Yuan M, Frantz DF, Melendez PA,
Hansen L, Lee J and Shoelson SE: Local and systemic insulin
resistance resulting from hepatic activation of IKK-beta and
NF-kappaB. Nat Med. 11:183–190. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Singleton KD, Beckey VE and Wischmeyer PE:
Glutamine prevents activation of NF-kappa B and stress kinase
pathways, attenuates inflammatory cytokine release, and prevents
acute respiratory distress syndrome (ARDS) following sepsis. Shock.
24:583–589. 2005. View Article : Google Scholar
|
|
29
|
Singleton KD and Wischmeyer PE:
Glutamine’s protection against sepsis and lung injury is dependent
on heat shock protein 70 expression. Am J Physiol Regul Integr Comp
Physiol. 292:R1839–R1845. 2007.
|
|
30
|
Huang Y, Li N, Liboni K and Neu J:
Glutamine decreases lipopolysaccharide-induced IL-8 production in
Caco-2 cells through a non-NF-kappaB p50 mechanism. Cytokine.
22:77–83. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fan JG, Qian Y, Zheng XY, Cai XB and Lu
YS: Effects of pentoxifyiline on hepatic nuclear factor-kappa B
signaling pathway and insulin resistance in nonalcoholic
steatohepatitis rats induced by fat-rich diet. Zhonghua Gan Zang
Bing Za Zhi. 14:762–766. 2006.(In Chinese).
|














