Introduction
Agmatine is an endogenous amine synthesized via the
decarboxylation of L-arginine mediated by arginine decarboxylase.
Agmatine is expressed in a variety of animal organs, particularly
the brain, where it acts as a novel neurotransmitter or
neuromodulator (1).
Although the specific physiological actions of
agmatine have yet to be elucidated, numerous studies have confirmed
that agmatine significantly inhibits seizures induced by maximal
electroshock and pentylenetetrazole in rat models (2–6). The
selective reduction of N-methyl-D-aspartic acid (NMDA)
receptor-mediated activity (3,4) and
the inhibition of nitric oxide synthase activity (2,6) have
been proposed to contribute to this inhibitory effect on seizures.
However, the majority of previous studies have investigated the
effect of agmatine on acute seizures, and only a few studies have
used chronic epilepsy animal models, which are similar to the
pathological physiology of clinical epileptic patients. It was
hypothesized that agmatine may also have an anticonvulsive effect
in chronic epilepsy. Therefore, in the present study,
pentylenetetrazole-induced chronic epilepsy rat models were
employed in order to examine the anticonvulsive effects of
agmatine.
Materials and methods
Animals
A total of 50 healthy, male adult Sprague-Dawley
rats (weighing between 170 and 200 g; The Experimental Animal
Center of Wenzhou Medical College, Wenzhou, China) were used in the
present study. Convulsion was induced in the rats using
pentylenetetrazole (Sigma, St. Louis, MO, USA) as previously
described (7). Pentylenetetrazole
(35 mg/kg) was administered to rats in the agmatine pretreatment
and model control groups each morning via intraperitoneal injection
for 28 consecutive days. The present study was approved by the
ethics committee at the Medical University of Wenzhou (Wenzhou,
Zhejiang, China) and was in accordance with the Chinese laws for
animal protection.
The rats were randomly divided into the following
five groups, with 10 rats in each group: i) the saline-saline group
(normal control group), saline was injected as the negative
control; ii) the pentylenetetrazole-saline group (model control
group), the rats were treated with saline 30 min prior to
intraperitoneal injections of pentylenetetrazole; iii) the
pentylenetetrazole low-dose agmatine group (pentylenetetrazole + 20
mg/kg agmatine; Sigma); iv) the pentylenetetrazole medium-dose
agmatine group (pentylenetetrazole + 40 mg/kg agmatine); and v) the
pentylenetetrazole high-dose agmatine group (pentylenetetrazole +
80 mg/kg agmatine). Agmatine pretreatment was administered 30 min
prior to the pentylenetetrazole injections.
Behavioral observations of
convulsion
The behavior of each rat was observed for 1 h after
pentylenetetrazole injection. The seizure activity was scored
according to the following five-point scale as previously described
by Fathollahi et al (8):
stage 0, no response; stage 1, ear and facial twitching; stage 2,
convulsive wave throughout the body; stage 3, myoclonic jerks;
stage 4, turn onto their side; stage 5, turn over onto their back,
generalized tonic-clonic seizures. The convulsion grade of each rat
was recorded daily. If the rat maintained the epileptic behavior
(i.e. at stage 2) for five consecutive days, it was regarded as
kindling and the kindling rate was calculated.
Sample preparation
After the rats were decapitated, the entire brain
was rapidly removed and dissected on ice. One half of the
hippocampus was immediately flash frozen in liquid nitrogen and
stored at −80°C for subsequent reverse transcription polymerase
chain reaction (RT-PCR) experiments. The other half was immersed in
4% paraformaldehyde (Shanghai Generay Biotech Co., Ltd., Shanghai,
China) for 24 h at 4°C and then paraffin embedded. The
paraffin-embedded brain was then cut into 5-μm thick coronal
sections using a microtome. For each rat, several brain sections
were collected for subsequent experiments.
Hematoxylin and eosin staining
Two paraffin slices were selected and stained using
hematoxylin and eosin as previously described (9). In the slices, hippocampal CA3, CA1
and DG regions were examined using a light microscope (BX51M;
Olympus, Tokyo, Japan; magnification, ×10) to observe morphological
alterations of the hippocampal neurons.
Immunohistochemistry
Immunostaining was performed on the brain slices
using the Polink-2 Plus® HRP Polymer Detection System
(PV-9001; GBI Labs, Mukilteo, WA, USA) as previously described
(9). Then, the sections were
briefly dehydrated through a graded series of ethanol and incubated
with rabbit anti-mouse glial fibrillary acidic protein (GFAP)
antibody (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). The
sections were washed with phosphate-buffered saline (Shanghai
Generay Biotech Co., Ltd.) and then incubated with poly horseradish
peroxidase anti-rabbit secondary antibody (PV-9001). The
avidin-biotin complex and diaminobenzidine were used to obtain a
visible reaction product. As a negative control, the specimens in
the control experiments were processed without primary or secondary
antibodies. The immunostaining of inducible nitric oxide synthase
(iNOS) was performed in a similar manner, however, the primary
antibodies were substituted with rabbit anti-mouse iNOS antibody
(Santa Cruz Biotechnology, Inc.). A Leica microscope equipped with
a digital camera was used for the examination and imaging of the
sections (Leica, Solms, Germany).
Image analysis
To quantify the GFAP expression, the average number
of positive cells in each section were counted in a blinded manner
in five randomly selected high power fields in the hippocampal CA1
and CA3 areas (magnification, ×20), and plotted using Prism 3.0
software (GraphPad Software, Inc., San Diego, CA, USA). To quantify
the iNOS expression, Image-Pro Plus 6.0 (Media Cybernetics, Inc.,
Rockville, MD, USA) was used to analyze the iNOS
immunohistochemical images and to calculate the average light
density values (IOD/area) of each section of five randomly selected
high power fields in the hippocampal CA1 and CA3 areas
(magnification, ×40). All the sections were analyzed under the same
light intensity and magnification.
RT-PCR
RT-PCR was performed using the Quant One Step RT-PCR
kit (Tiangen Biotech, Co., Ltd., Beijing, China) according to the
manufacturer’s instructions. The thermal cycler parameters were: 4
min at 94°C followed by 30 cycles of 30 sec at 94°C, 30 sec at
58°C, 40 sec at 72°C and then 10 min at 72°C. The following
specific primers were used: NR1, forward 5′-GCTGCACGCCTTTATCTG-3′
and reverse 5′-TCCTACGGGCATCCTTGT-3′; NR2b, forward
5′-CACGGTGCCTTCAGAGTT-3′ and reverse 5′-CCTCCTCCAAGGTGACAA-3′. The
PCR products were separated using electrophoresis on a 2.0% agarose
gel. The intensity of the bands was analyzed using BioSense SC-810
Gel Documentation System (Shanghai Bio-Tech Co., Ltd., Shanghai,
China) and Gel-Pro 3.1 software (Media Cybernetics, Inc., Bethesda,
MD, USA).
Statistical analysis
Statistical analysis was performed using SPSS 180
statistical software (SPSS, Inc., Chicago, IL, USA). The values are
expressed as the mean ± standard error of the mean. Comparisons
among multiple groups were performed using a one-way analysis of
variance and a least significant difference post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Agmatine treatment reduces the severity
of pentylenetetrazole-induced chronic seizures
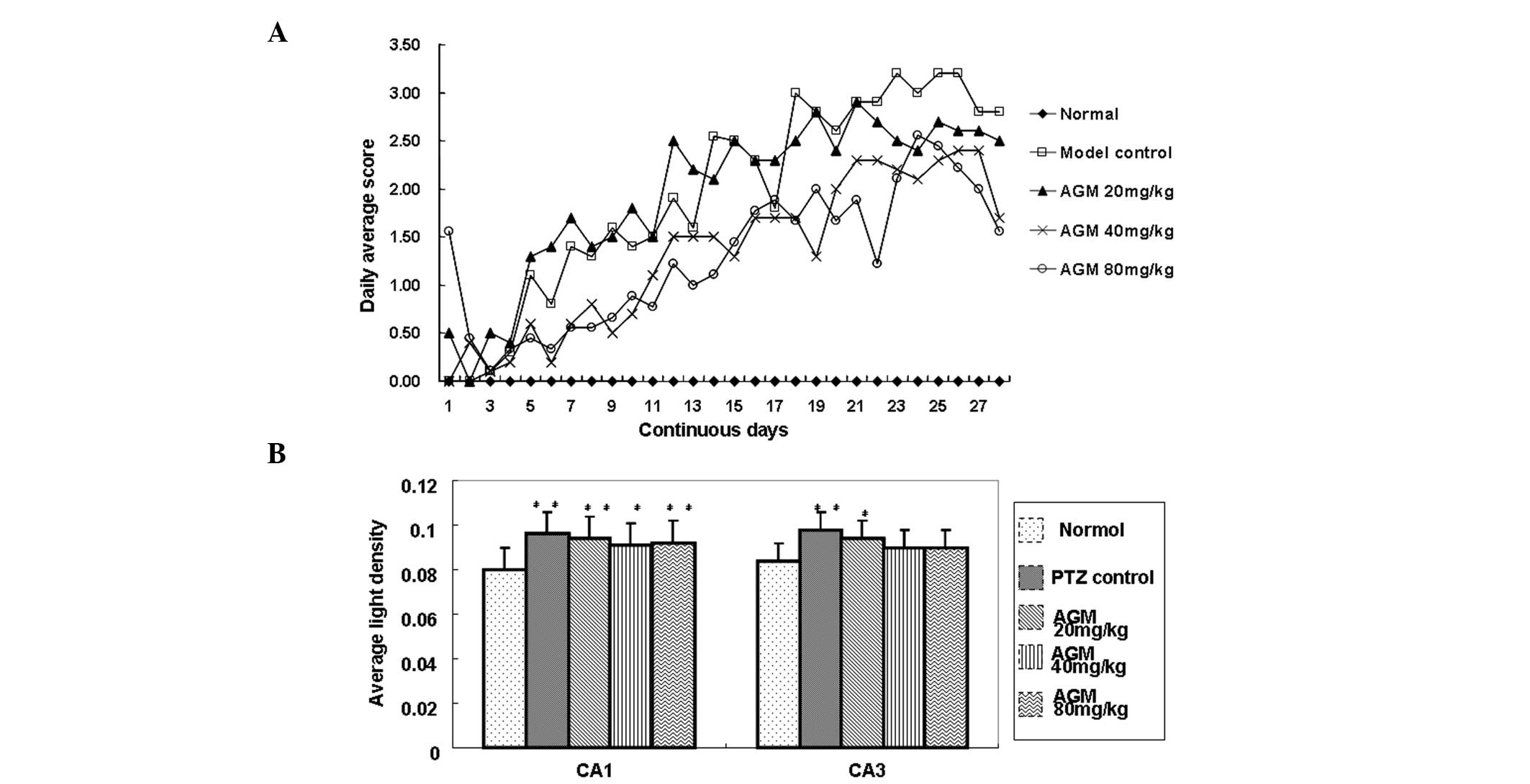
To evaluate the effect of agmatine on chronic
seizures induced by pentylenetetrazole, convulsions were measured
using a five-point scale, as previously described by Fathollahi
et al (8). Following 20
days of treatment, the majority of the rats reached a completely
kindled condition. The daily average seizure grades in the 40 and
80 mg/kg agmatine groups were significantly lower compared with
those of the model control group (P<0.001; Fig. 1A). However, no significant
difference in the kindling rate was observed among the agmatine and
model control groups (Table
I).
 | Table ISeverity and the kindling rate of rats
in each treatment group. |
Table I
Severity and the kindling rate of rats
in each treatment group.
| | | Seizure grade | |
|---|
| | |
| |
|---|
| Group | Sample (n) | Survival rate | Moderate (≤III) | Severe (≥III) | Kindling rate |
|---|
| Normal control
(A) | 10 | - | - | - | - |
| Model control
(B) | 10 | 10/10 | 5/10 | 5/10 | 10/10 |
| Agmatine 20 mg/kg
(C1) | 10 | 10/10 | 6/10 | 4/10 | 10/10 |
| Agmatine 40 mg/kg
(C2) | 10 | 10/10 | 6/10 | 4/10 | 10/10 |
| Agmatine 80 mg/kg
(C3) | 10 | 10/10 | 7/10 | 3/10 | 8/10 |
Agmatine has no effect on the expression
of iNOS
To investigate the effect of agmatine on the
expression of iNOS, the average light density (IOD/Area) values of
iNOS-positive regional expression in the CA1 and CA3 areas of the
hippocampus were obtained. The data revealed that the
pentylenetetrazole group had significantly higher values compared
with the normal group (P<0.05). A decreasing trend was observed
in the agmatine-treated rats compared with the rats in the model
control group, however, no significant difference was observed.
This suggested that iNOS activity may be increased in chronic
epileptic seizures, however, the long-term usage of agmatine does
not significantly inhibit iNOS expression (Fig. 1B).
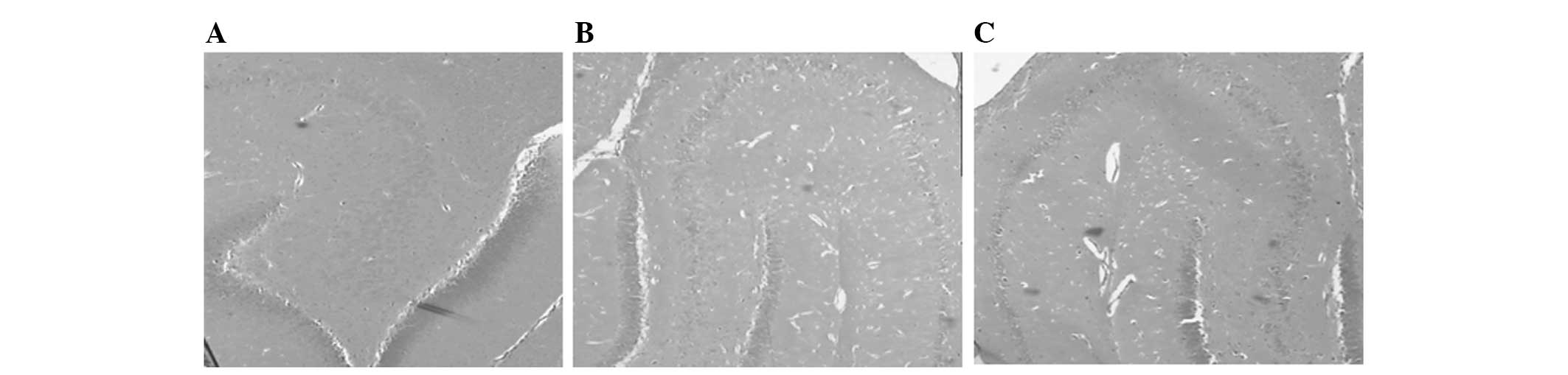
Agmatine treatment decreases cell injury
in the hippocampal area of pentylenetetrazole-treated rats
To determine whether agmatine alleviated cell injury
in the hippocampal area of pentylenetetrazole-treated rats,
hippocampal pyramidal cells were observed under a microscope. In
the normal group, hippocampal pyramidal cells exhibited regular
morphological integrity, whereas, in the model control group, cell
loss was observed and the cells were irregularly distributed and
exhibited abnormal structures, as well as wider interspaces. By
contrast, the agmatine group also exhibited neuronal loss, however,
with reduced severity, particularly in the hippocampal area
(Fig. 2). This observation
indicated that agmatine treatment partially decreased cell injury
in the hippocampal area.
Treatment with agmatine suppresses
astrocytic hyperplasia
To investigate how agmatine treatment affects
astrocytic hyperplasia, hematoxylin and eosin staining and image
analysis were performed (Fig. 3A).
In the CA1 and CA3 hippocampal regions of normal rats,
GFAP-positive cells were scattered, light brown-yellow in color and
reduced in number. By contrast, in the model control group,
GFAP-positive cells were significantly increased in number and
exhibited more intense staining, as well as thicker and extended
neurites. In the agmatine groups, GFAP expression was increased to
a certain extent, however, the number of GFAP-positive cells was
reduced and the staining was less intense compared with the model
control group. The cells were decreased in size and the neurites
were relatively thinner and shorter. The differences between the
agmatine groups and the model control group were significant
(P<0.05), in particular for the 40 and 80 mg/kg agmatine groups
(P<0.01), as shown in Fig. 3B.
These results demonstrated that agmatine suppressed astrocytic
hyperplasia.
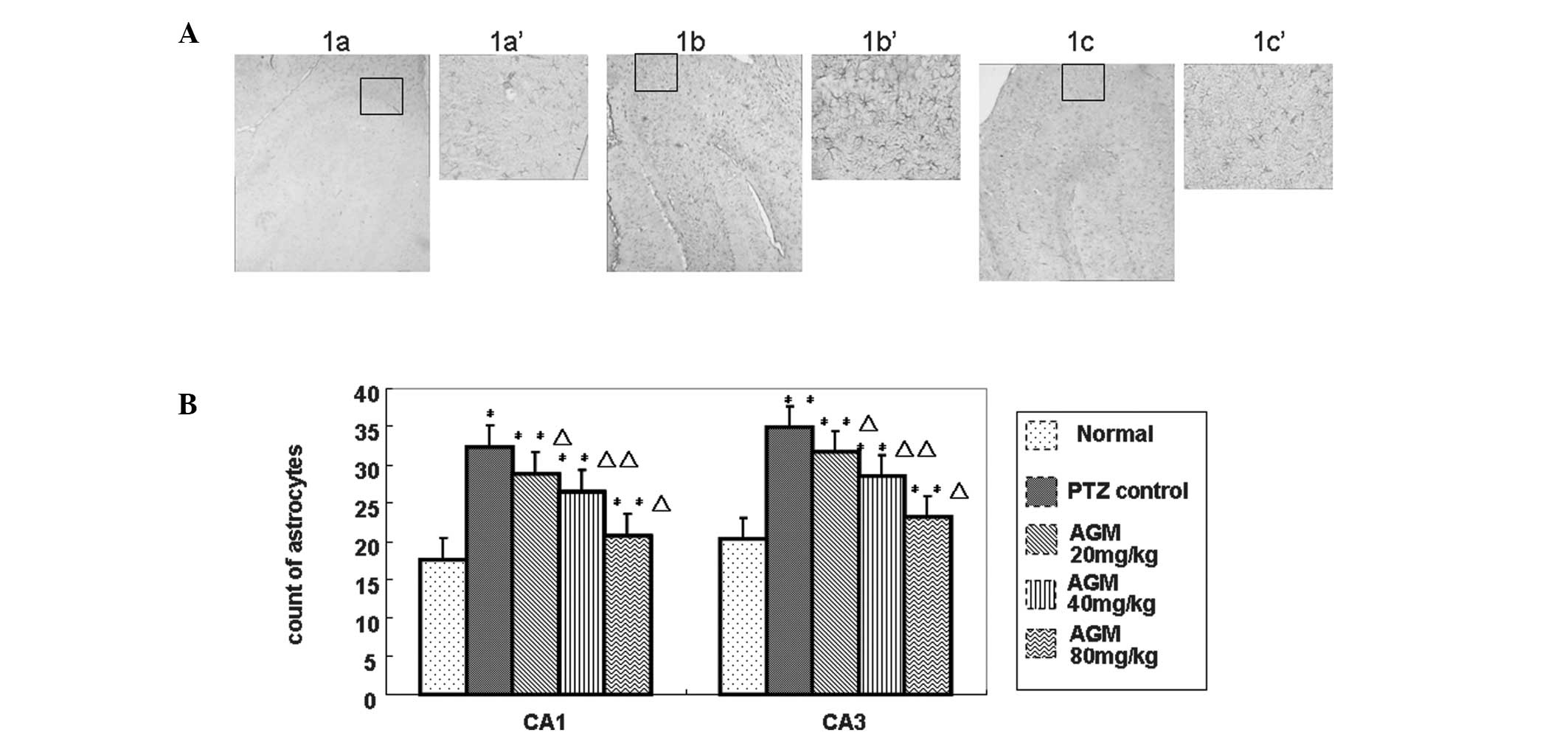 | Figure 3(A) Hyperplasia of astrocytes in the
hippocampus of agmatine-treated rats was reduced. Immunostaining of
GFAP was performed on the sections to detect increased astrocyte
expression. The expression of astrocytes in the (1a and 1a′) normal
control group and the (1c and 1c′) agmatine group was decreased
compared with the (1b and 1b′) model control group (magnification,
×4; boxed area, magnification, ×20). (B) The number of GFAP
positive cells was counted from five randomly selected microscopic
fields (magnification, ×20) and plotted. Data are presented as the
mean ± the standard deviation. *P<0.05,
**P<0.01, compared with the normal control group.
ΔP<0.05, ΔΔP<0.01, for comparisons
between the agmatine group and the model control group. AGM,
agmatine; PTZ, pentylenetetrazole; GFAP, glial fibrillary acidic
protein. |
Agmatine treatment decreases the
expression of the NMDA receptor
In order to analyze the alterations in the
expression of the NMDA receptor induced by agmatine, RT-PCR was
performed to detect NR1 and NR2b mRNA expression in the rat
hippocampus (Fig. 4A). Compared
with the model control group, the quantity of NR1 mRNA in the
agmatine groups (40 and 80 mg/kg) was significantly decreased
(P<0.01), suggesting that pretreatment with agmatine may
suppress the actions of the hippocampal NR1 (Fig. 4B). However, the low-dose agmatine
group (20 mg/kg) showed no significant difference compared with the
model control group (Fig. 4B). In
addition, no significant difference in NR2b mRNA expression was
observed among all the groups (data not shown). These results
indicated that treatment with higher concentrations of agmatine
decreased the expression of the NMDA receptor.
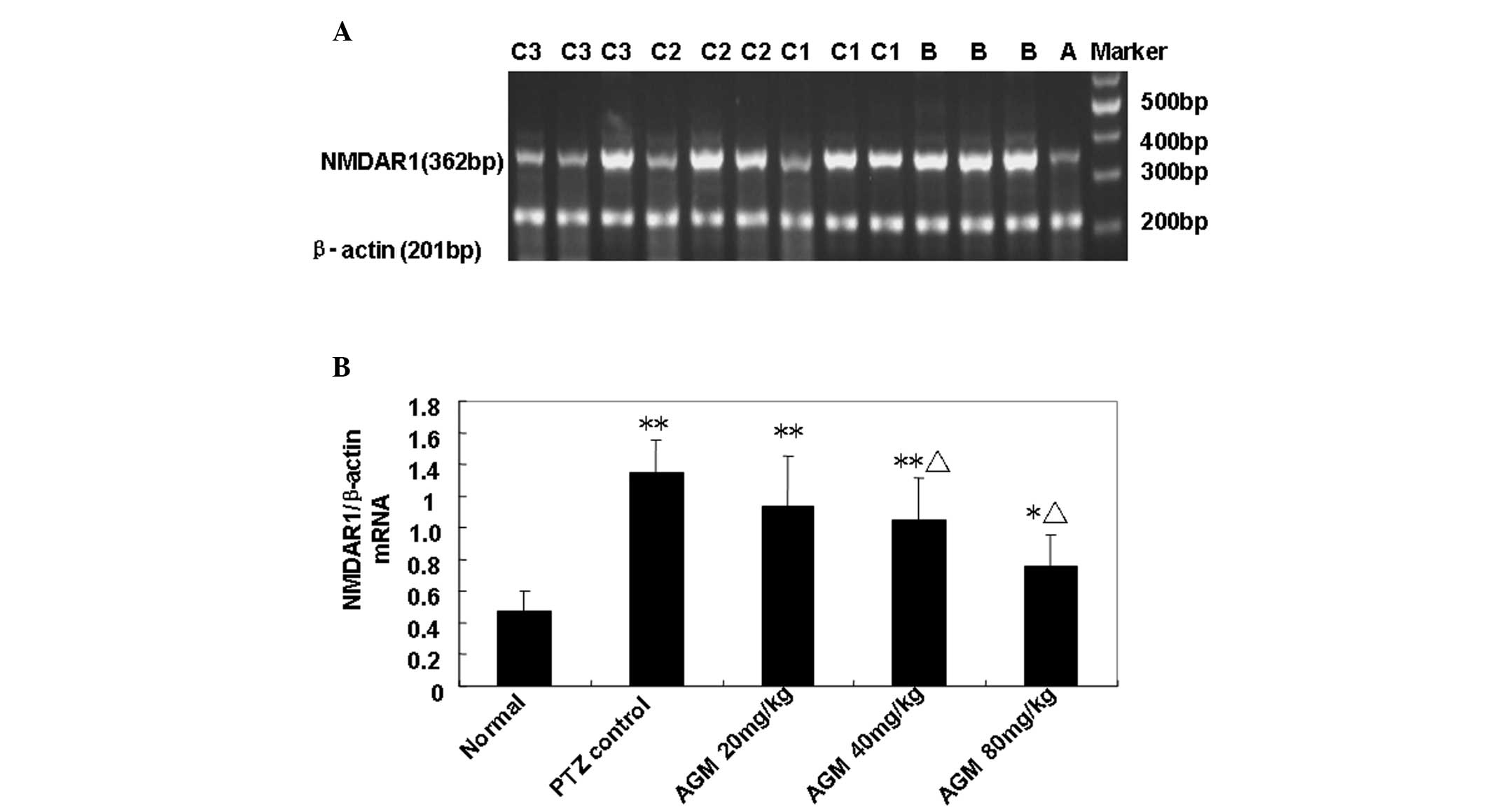 | Figure 4(A) Detection of NMDAR1 mRNA
expression in the rat hippocampus using reverse transcription
polymerase chain reaction. A, normal control group; B, model
control group; C1, C2 and C3, agmatine groups (20, 40 and 80 mg/kg,
respectively). (B) Quantification of NMDAR1 mRNA expression of the
five groups. The Y axis indicates the ratio of optical density (OD)
of the samples to the corresponding internal standard (β-actin).
Data are expressed as the mean ± standard error of the mean (n=10).
*P<0.05, **P<0.01, compared with the
normal control group. ΔP<0.01, for comparisons
between the agmatine group and the model control group. NMDAR1,
N-methyl-D-aspartic acid receptor; AGM, agmatine; PTZ,
pentylenetetrazole. |
Discussion
In the present study, it was demonstrated that
consecutive administration of agmatine provided protection against
pentylenetetrazole-induced chronic seizures in rats. These results
are consistent with previous studies that demonstrated the
inhibitory effect of agmatine in acute seizure animal models
(2,3). Furthermore, in the present study,
rats treated with agmatine exhibited significantly reduced
astrocytic hyperplasia and neurological defects in the hippocampal
area compared with rats in the model control group. Furthermore,
the expression of the NMDA1 receptor was selectively suppressed in
agmatine-treated rats.
The results from the present study are in accordance
with several previous studies demonstrating that high doses of
agmatine had marked anticonvulsive effects (3–5). In
the present study, only rats treated with a high dose of agmatine
(40 and 80 mg/kg/d) demonstrated clear inhibitory effects. This may
be due to the rapid metabolism of agmatine in the peripheral
tissues. In addition, the blood-brain barrier may also restrict the
penetration of agmatine into the brain (6). Therefore, an adequate peripheral dose
is required to produce apparent protective effects. However, this
is controversial as certain studies have found that agmatine
administered alone at doses of ≤100 mg/kg had no affect on the
threshold and provided no protection against seizures (7). In fact, in the present study repeated
administration of agmatine did not decrease the kindling rate. This
suggests that agmatine is unable to alter the threshold and this
may be associated with under-dosing, which requires investigation
in future studies.
Astrocytes are important glial cells in the brain.
Following epilepsy, the number of astrocytes increases and
alterations in morphology and function are observed. Astrocytes
have been demonstrated to be important in the mechanisms underlying
epilepsy (10). For example, it
has been demonstrated that they are involved in the maintenance of
the inflammatory state during epilepsy by releasing inflammatory
cytokines. These cytokines directly alter the excitability of the
neurons and promote mossy fiber budding of the dentate gyrus to
form an excitability loop, which may induce seizures (11). In the present study, GFAP
immunohistochemistry demonstrated that agmatine was able to
significantly reduce hippocampal astrocytic cell proliferation in a
dose-dependent manner following pentylenetetrazole-induced
seizures. This may contribute to the inhibitory effect of agmatine
on seizures.
The activation of NMDA receptors is responsible for
the development of seizures and their binding sites are upregulated
in different types of convulsant animal models. NMDA receptor
antagonists have previously been demonstrated to inhibit convulsion
(12). In addition, agmatine has
been shown to selectively modulate NMDA subunits in rat hippocampal
neurons (12) and mediate
anticonvulsive actions (3,4). In accordance with previous studies,
RT-PCR results from the present study demonstrated that the mRNA
expression of NR1 was significantly inhibited in the agmatine
groups (40 and 80 mg/kg). However, this was only observed in
animals repeatedly treated with large doses of agmatine. The reason
for this may be that only 1% of the injected agmatine reaches the
brain (4). However, the same
result was not observed for NMDA R2b mRNA expression.
Seizures are known to cause neuronal death and cell
loss may in turn increase the potential for further seizure
activity. This feedback cycle may explain the progressive and
chronic course of epilepsy (13).
Previous studies have revealed a decrease in the number of
hippocampal neurons in seizures induced by pentylenetetrazole
(14). In the present study, the
results of hippocampal morphology suggest that agmatine may
decrease cell loss in the rat hippocampus. Agmatine exhibits
seizure-suppressive and neuroprotective capabilities and may
therefore protect against convulsions on seizure-suppressive and
neuroprotective capabilities.
Several studies have also suggested that the
inhibitory effect of agmatine may be important in its
anticonvulsive properties (2,6). In
the present study, the expression of iNOS was found to increase in
the hippocampus following pentylenetetrazole administration.
However, agmatine was not found to significantly inhibit iNOS
expression.
In conclusion, the present study demonstrated that
agmatine has a protective effect against pentylenetetrazole-induced
chronic seizures and that its effective dose is relatively large
(80 mg/kg). Agmatine treatment results in decreased astrocytic
hyperplasia, neuronal cell loss and selective suppression of the
NMDA1 receptor in the hippocampus. The majority of clinical
epilepsy cases are diagnosed as long-term repeated chronic
epilepsy. Thus, further investigation regarding the function of
agmatine in chronic epilepsy is particularly important.
Acknowledgements
This study was supported by grants from the Building
Funding of Wenzhou Science & Technology Bureau Fund (grant no.
Y20070031).
References
|
1
|
Raasch W, Schäfer U, Chun J and Dominiak
P: Biological significance of agmatine, an endogenous ligand at
imidazoline binding sites. Br J Pharmacol. 133:755–780. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Demehri S, Homayoun H, Honar H, et al:
Agmatine exerts anticonvulsant effect in mice: modulation by
α2-adrenoceptors and nitric oxide. Neuropharmacology. 45:534–542.
2003.
|
|
3
|
Su RB, Wei XL, Zheng JQ, Liu Y, Lu XQ and
Li J: Anticonvulsive effect of agmatine in mice. Pharmacol Biochem
Behav. 77:345–349. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Feng Y, LeBlanc MH and Regunathan S:
Agmatine reduces extracellular glutamate during
pentylenetetrazole-induced seizures in rat brain: a potential
mechanism for the anticonvulsive effects. Neurosci Lett.
390:129–133. 2005. View Article : Google Scholar
|
|
5
|
Luszczki JJ, Czernecki R, Wojtal K,
Borowicz KK and Czuczwar SJ: Agmatine enhances the anticonvulsant
action of Phenobarbital and valproate in the mouse maximal
electroshock seizure model. J Neural Transm. 115:1485–1494. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bahremand A, Ziai P, Khodadad TK,
Payandemehr B, et al: Agmatine enhances the anticonvulsant effect
of lithium chloride on pentylenetetrazole-induced seizures in mice:
Involvement of L-arginine/nitricoxide pathway. Epilepsy Behav.
18:186–192. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Obay BD, Taşdemir E, Tümer C, Bilgin H and
Atmaca M: Dose dependent effects of ghrelin on
pentylenetetrazole-induced oxidative stress in a rat seizure model.
Peptides. 29:448–455. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fathollahi Y, Motamedi F, Semnanian S and
Zardoshti M: Examination of persistent effects of repeated
administration of pentylenetetrazol on rat hippocampal CA1:
evidence from in vitro study on hippocampal slices. Brain Res.
758:92–98. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang XS, Chen YY, Shang XF, et al:
Idazoxan attenuates spinal cord injury by enhanced astrocytic
activation and reduced microglial activation in rat experimental
autoimmune encephalomyelitis. Brain Res. 1253:198–209. 2009.
View Article : Google Scholar
|
|
10
|
Stringer JL: Repeated seizures increase
GFAP and vimentin in the hippocampus. Brain Res. 717:147–153. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ravizza T, Gagliardi B, Noé F, Boer K,
Aronica E and Vezzani A: Innate and adaptive immunity during
epileptogenesis and spontaneous seizures: evidence from
experimental models and human temporal lobe epilepsy. Neurobiol
Dis. 29:142–160. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ekonomou A and Angelatou F: Upregulation
of NMDA receptors in hippocampus and cortex in the
pentylenetetrazol-induced ‘kindling’ model of epilepsy. Neurochem
Res. 24:1515–1522. 1999.PubMed/NCBI
|
|
13
|
Bengzon J, Kokaia Z, Elmér E, Nanobashvili
A, Kokaia M and Lindvall O: Apoptosis and proliferation of dentate
gyrus neurons after single and intermittent limbic seizures. Proc
Natl Acad Sci USA. 94:10432–10437. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Oses JP, Leke R, Portela LV, et al:
Biochemical brain markers and purinergic parameters in rat CSF
after seizure induced by pentylenetetrazol. Brain Res Bull.
64:237–242. 2004. View Article : Google Scholar : PubMed/NCBI
|