Introduction
Alcoholic liver disease (ALD) is a common clinical
complication resulting from long-term alcohol abuse. Its
morphological features include alcoholic fatty liver, hepatitis and
cirrhosis. The pathogenesis of ALD is multifactorial and involves
genetic, nutritional and environmental factors, in addition to
numerous injurious factors, such as oxidative or nitrosative
stress, bacterial lipopolysaccharide (LPS) and cytokines (1,2).
Alcohol-induced sensitization of liver macrophages
by portal endotoxins/LPS is considered to be a hallmark of ALD.
Intracellular mechanisms that are associated with LPS-induced
signaling are critical in the initiation and progression of ALD.
LPS recognition by Toll-like receptor 4 (TLR4) and cell
differentiation antigen (CD14) on liver macrophages and activation
of downstream signaling pathways, which culminate in the activation
of transcription factors, such as nuclear factor (NF)-κB, lead to
increased inflammatory cytokine production in ALD. In addition,
LPS-induced mitogen-activated protein kinases (MAPKs), such as
extracellular regulated protein kinases (ERKs) contribute to liver
injury. The importance of alcohol-induced reactive oxygen species
and interactions with TLR pathways in macrophages that lead to
inflammation are becoming increasingly evident. Collectively, these
signaling pathways induce pro- and anti-inflammatory cytokines,
which are significant in ALD (3–5).
Qinggan (QG) Huoxue (HX) Recipe (R) exerts a broad
range of pharmacological effects, including partially reversible
steatosis, decreased fibrosis marker levels, decreased inflammatory
cytokine levels and resistance to lipid peroxidation. The study
group introduces the Yu-Re pathogenesis theory in ALD, which is
based on clinical epidemiology and advocates the rule of QG and HX
for the treatment of ALD (6).
Yu-Re pathogenesis is based on our previous epidemiological
investigation with 745 ALD patients. Shi-re and Yu-xue were the
basic pathomechanism of ALD, therefore the rule of QG and HX were
the basic therapy for ALD. The aim of the present study was to
characterize the regulation of the LPS-Kupffer cell (KC) signal
molecules by administering QGHXR to rats with ALD in order to
determine the experimental basis for the use of traditional Chinese
medicine in ALD therapy.
Materials and methods
Reagents
QGHXR (9 g bupleurum root, 9 g scutellaria root, 15
g red sage root, 9 g Carapax Trionycis and 15 g Radix
Puerariae), QGR (9 g bupleurum root and 9 g scutellaria
root) and HXR (15 g red sage root, 9 g Carapax Trionycis and 15 g
Radix Puerariae), were concentrated to 4.75, 1.5 and 3.25
g/ml, respectively, and processed at the Department of Pharmacy,
Longhua Hospital (Shanghai, China). In order to ensure the
uniformity and stability of herbal products, we performed the HPLC
method to detect the contents of four flavones including Puerarin,
Baicalin, Baicalein and Wogonin (7). Rabbit polyclonal anti-phosphorylated
(p)-ERK and mouse anti-rat NF-κB were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA) and anti-TLR4 was
purchased from Santa Cruz Biotechnology, Inc., (Santa Cruz, CA,
USA).
Animal preparation
Specific pathogen-free male Wistar rats (n=100;
weight, 180±10 g) were purchased from Slac Laboratory Animal
Center, Inc. (Shanghai, China). Daily general observations and
weekly body weights of the rats were recorded. The rats were
randomly divided into three groups: The blank group (n=10), the
carbon tetrachloride (CCl4) group (n=10) and the model
group (n=80). The model group was administered with a 10 ml/kg/d
dose mixture twice per day, which included 10 ml 60% alcohol, 2 ml
corn oil and 25 mg pyrazole. Intraperitoneal injections of 0.25
ml/kg of 25% CCl4 in olive oil were administered twice a
week from the second week (8); the
CCl4 group only received intraperitoneal injections. Two
rats from the model group were sacrificed each week for
histological observation and after four weeks, the model group was
divided into four subgroups: QGHXR (n=15), QGR (n=15), HXR (n=15)
and the model group (the remaining rats). The animals were
sacrificed with exsanguinating after blood samples were obtained
from abdominal aorta. The model group was administered with 10
ml/kg saline per day, the QGHXR group received 200 mg/kg QGHXR, the
QGR group received 137.5 mg/kg QGR and the HXR group received 62.5
mg/kg HXR. The blank and CCl4 groups were administered
with saline and at the end of the six weeks, all of the rats were
anesthetized with 2% pentobarbital sodium at the dosage of 2 ml/kg
and sacrificed. Blood samples and liver tissue specimens were
collected and a section of the liver was fixed for histopathology.
Another section was used for polymerase chain reaction (PCR) and
western blot analysis; the remaining tissue was stored at −80°C
until the assays were conducted. All of the rats were handled
according to the recommendations of the National Institutes of
Health Guidelines for Care and Use of Laboratory Animals. The
experimental protocol was approved by the Shanghai Medical
Experimental Animal Care Committee (Shanghai, China).
Serum alanine aminotransferase (ALT),
aspartate aminotransferase (AST) and tumor necrosis factor (TNF)-α
assays
Serum ALT and AST levels were assayed using a
Hitachi 7170S Biochemical Analyzer (Hitachi Ltd., Tokyo, Japan).
The serum TNF-α level was assayed using a mouse TNF-α enzyme-linked
immunosorbent assay kit (Yusen Biotech Inc., Shanghai, China). A
total of 50 μl standards (standard curve concentrations: 1,000,
500, 250, 125, 62.5, 32, 16 and 0 ng/ml) or serum was added to each
well in duplicate, the absorbance was measured at 450–550 nm with
Microplate Spectrophotometer (Bio-Tek Instruments Inc., Winooski,
VT, USA) and the results were calculated using a standard
curve.
Pathology and immunohistochemistry
Small segments of liver tissue were fixed in 10%
neutral buffered formalin and processed into paraffin sections for
hematoxylin and eosin (H&E) or immunohistochemical staining.
The morphological variations were observed by two pathologists who
were blinded to the experimental information. Diehl et al
(9) and Wang (10) have described the pathological
changes in fatty degeneration, apoptosis and necrosis, which are
observed by H&E staining, as well as the associated criteria.
Immunohistochemical staining was performed with a
streptavidin-biotin complex kit (Boster Biological Technologies,
Inc., Wuhan, China) for CD14 using rabbit anti-CD14 antibodies
(1:100; Boster Biological Technologies, Inc.) and CD68 with mouse
anti-rat CD68 antibodies (1:200; Bio-Rad Laboratories, Hercules,
CA, USA). The slides were visualized with 3,3′-diaminobenzidine and
positive staining was indicated by a yellow-brown color. Evaluation
of the specific positive reactions were performed using Image-Pro
Plus 6.0 (Media Cybernetics, Inc., Rockville, MD, USA) and were
presented as the integral optical density value.
Semi-quantitative reverse
transcription-PCR (RT-PCR) analysis for TNF-α, CD14 and
TLR4
Total RNA was extracted from the liver tissue using
the TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA,
USA). The complementary DNA was synthesized from 2 μg total RNA
using Moloney murine leukemia virus reverse transcriptase (Takara
Co., Ltd., Japan) and mouse TNF-α, CD14 and TLR4 mRNA
were amplified using the primers shown in Table I. The PCR analysis was conducted as
follows: 26 Cycles at 95°C for 30 sec and at 60°C for 40 sec
followed by a 1 min extension stage at 72°C. The amplification
products were electrophoretically analyzed on 1.0% agarose gel
containing 0.1 μg/ml ethidium bromide.
 | Table ISequence-specific primers of TNF-α,
CD14, TLR4 and β-actin. |
Table I
Sequence-specific primers of TNF-α,
CD14, TLR4 and β-actin.
| Gene | Product (bp) | Forward | Reverse |
|---|
| TNF-α | 379 |
5′-ATCGGTCCCAACAAGGAGGAGAAGT-3′ |
5′-TCCTTAGGGCAAGGGCTCTTGATGG-3′ |
| CD14 | 481 |
5′-TCGGCTTGTTGCTGTTGCCTTTGAC-3′ |
5′-TTCTGCGAGCCAGGTATCCGTTGTT-3′ |
| TLR4 | 500 |
5′-GGATTTTACGAATTCCACCTGTTAT-3′ |
5′-CGATACAATTCGACCTGCTGCCTCA-3′ |
| β-actin | 224 |
5′-TGTGATGGTGGGTATGGGTCAGAAG-3′ |
5′-TCACGGTTGGCCTTAGGGTTCAGAG-3′ |
Semi-quantitative western blot analysis
for TLR4, p-ERK and NF-κB
Homogenates that contained equal quantities of the
proteins (50 μg) were separated using SDS-PAGE and
electrophoretically transferred onto a nitrocellulose membrane
(Bio-Rad Laboratories). The nitrocellulose membrane was blocked
overnight with 5% non-fat dry milk in phosphate-buffered saline
with Tween-20 (0.1%, v/w) at 4°C. The membrane was incubated with
the primary antibodies, which were diluted according to the
manufacturer’s instructions for 16 h. Horseradish
peroxidase-conjugated anti-rabbit or anti-mouse IgG (Santa Cruz
Biotechnology, Inc.) served as secondary antibodies. The
immunoreactive protein was visualized by ECL Photon
Chemiluminescence Western Blotting Detection Kits (FIVEphoton
Biochemicals, San Diego, CA, USA) and western blot analysis
according to the manufacturer’s instructions.
Replications and statistical
analysis
Each result was independently replicated at least
three times and certain results were repeated multiple times, when
appropriate or required. All of the continuous variables were
expressed as means ± standard deviation, and analysis of variance
and post hoc tests were used to measure the statistical
significance of differences compared with the control. The software
package, SPSS 15.0 (SPSS Inc., Chicago, IL, USA) was used for the
data analysis and P<0.05 was considered to indicate a
statistically significant difference.
Results
Parameter observations in rat models
The rats became excited and ran around the cage
following the administration of alcohol. Subsequently, they were
unable to walk and eventually fell into a deep sleep. Furthermore,
the weight of the rats noticeably reduced and the rats became
cachectic. The daily diet, weight and energy of the rats in the
QGHXR, QGR and HXR groups were better than those observed in the
model group. The serum ALT and AST levels were upregulated to a
greater extent in the model group compared with the control group;
in addition, the related molecules in the LPS-KC pathway were
identified to be partially activated in the model group. Twenty-one
rats succumbed during the study; however, no animal experienced
>10% QGHXR-related weight loss following the treatment regimen
(data not shown).
QGHXR reduces liver injury due to the
reduction of ALT, AST and TNF-α levels in ALD rats
QGHXR reduced the serum ALT, AST and TNF-α levels by
17.98% (43.53±6.66 vs. 53.07±10.65 U/l), 37.65% (120.09±31.15 vs.
192.62±38.43 U/l) and 21.86% (24.99±4.65 vs. 31.98±4.61 pg/ml),
respectively, compared with the model group. QGR significantly
decreased the AST levels, however, it did not affect ALT and TNF-α.
HXR reduced the AST and TNF-α levels without affecting the ALT
levels (Fig. 1A). The liver
samples from the model group presented with hepatic steatosis and
inflammatory monocytic infiltrates when compared with the samples
from the blank group (Fig. 1Ba and
b) and the CCl4 group appeared to be identical to
the blank group (Fig. 1Bc). The
QGHXR treatment significantly improved the aforementioned
pathological changes compared with the QGR or HXR treatments
(Fig. 1Bd–f). Furthermore, the
RT-PCR results identified that QGHXR and HXR suppressed TNF-α mRNA
expression, however, no distinct QGR effect was evident on TNF-α
mRNA (Fig. 1C).
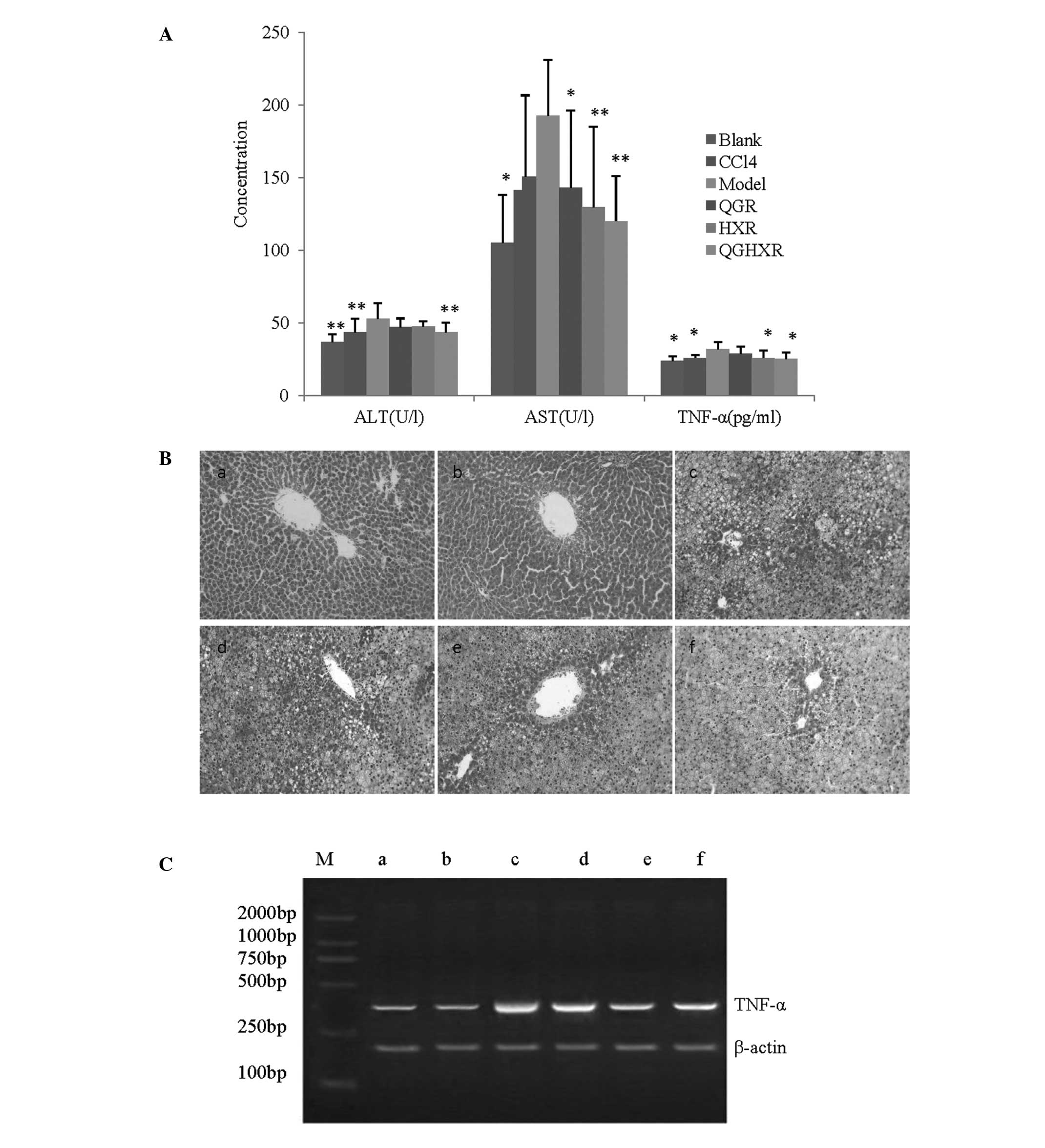 | Figure 1QGHXR improved liver injury by
reducing ALT, AST and TNF-α levels in the ALD rats. (A) Serum ALT,
AST and TNF-α levels of the ALD rats were significantly higher than
those observed in the control group. Treatment with QGHXR decreased
the ALT levels to a greater extent than QGR or HXR. Similar trends
were observed in the AST and TNF-α levels. *P<0.05
and **P<0.01 compared with the model group. (B)
Magnification, ×200. The liver in the model group (Bc) exhibited
marker hepatic steatosis and inflammatory infiltrates of monocytes
compared with the blank group (Ba). The CCl4 group (Bb)
was identical to the blank group. The QGHXR treatment (Bf)
significantly improved the pathological parameters compared with
(Bd) QGR or (Be) HXR treatment. (C) The reverse
transcription-polymerase chain reaction results revealed that QGHXR
and HXR suppressed TNF-α mRNA expression; however, QGR showed no
distinct effect on TNF-α mRNA. Lane M, marker; lane a, blank group;
lane b, CCl4 group; lane c, model group; lane d, QGR
group; lane e, HXR group; lane f, QGHXR group. CC14,
carbon tetrachloride; QGR, Qinggan Recipe; HXR, Huoxue Recipe;
QGHXR, Qinggan Huoxue Recipe; ALT, alanine aminotransferase; AST,
aspartate aminotransferase; TNF, tumor necrosis factor; ALD,
alcoholic liver disease; mRNA, messenger RNA. |
QGHXR downregulates CD68, CD14 and
TLR4 expression in ALD rat liver tissue
QGHXR and HXR inhibited KC activation (Fig. 2Aa–f) and immunohistochemistry and
western blot analysis demonstrated that QGHXR decreased the CD14
and TLR4 protein levels in the cytoplasm of the liver
(Figs. 2Ba–f and 3C). However, QGR and HXR showed no effect
on CD14 and TLR4 protein levels. In addition, the RT-PCR
results showed that the QGHXR treatment decreased the
overexpression of CD14 and TLR4 mRNA, as well as
identifying that QGHXR was more effective than HXR and QGR
(Fig. 3A and B).
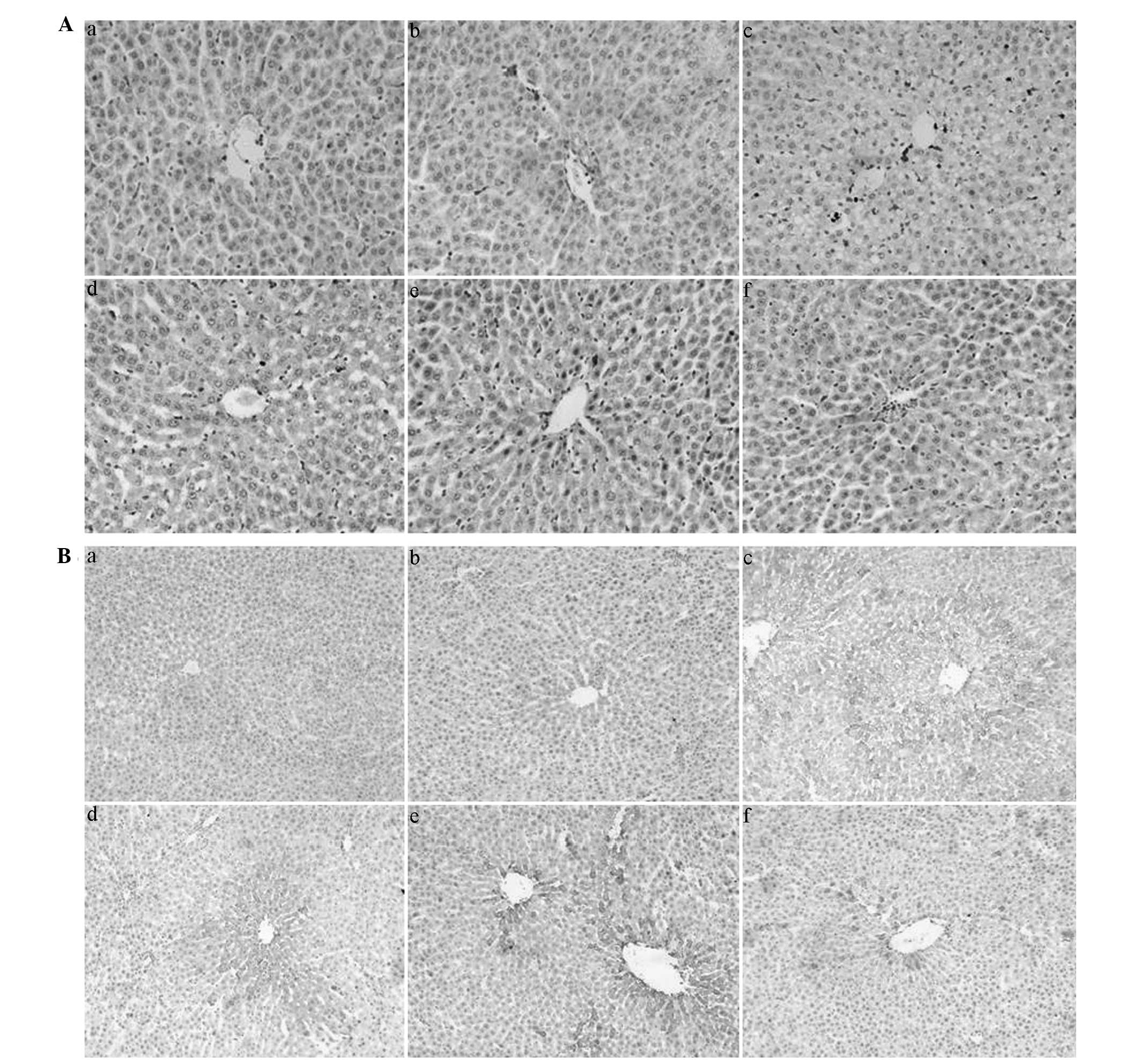 | Figure 2QGHXR downregulated CD68 and CD14
expression in the liver tissue of the ALD rats. (A) Magnification,
×400. (Aa) Marginal CD68 positive staining was observed in the
sinus hepaticus, the portal area of the hepatic lobules and in the
liver of the blank group. (Ab) No obvious change in the
CCl4 group was evident. (Ac) Evident CD68-positive
staining was observed in the liver of the model group,
concentrating in the sinus hepaticus where steatosis and
inflammatory cell infiltrates were apparent. (Af) QGHXR and (Ae)
HXR inhibited Kupffer cell activation; however, (Ad) QGR did not
show any significant effect. (B) Magnification, ×200. (Ba)
Immunohistochemistry showed a small positively stained area in the
cytoplasm of the liver from the blank group, which was
predominantly situated on the sinus hepaticus or non-parenchymal
cells around the central veins. (Bb) No obvious change in the
CCl4 group was evident. (Bc) An obvious CD14-positive
area was observed in the model group. (Bf) QGHXR decreased the CD14
expression in the model rats; however, (Bd) QGR and (Be) HXR did
not significantly effect the CD14 expression. QGHXR, Qinggan Huoxue
Recipe; CD, cell differentiation antigen; ALD, alcoholic liver
disease; CC14, carbon tetrachloride; HXR, Huoxue Recipe;
QGR, Qinggan Recipe. |
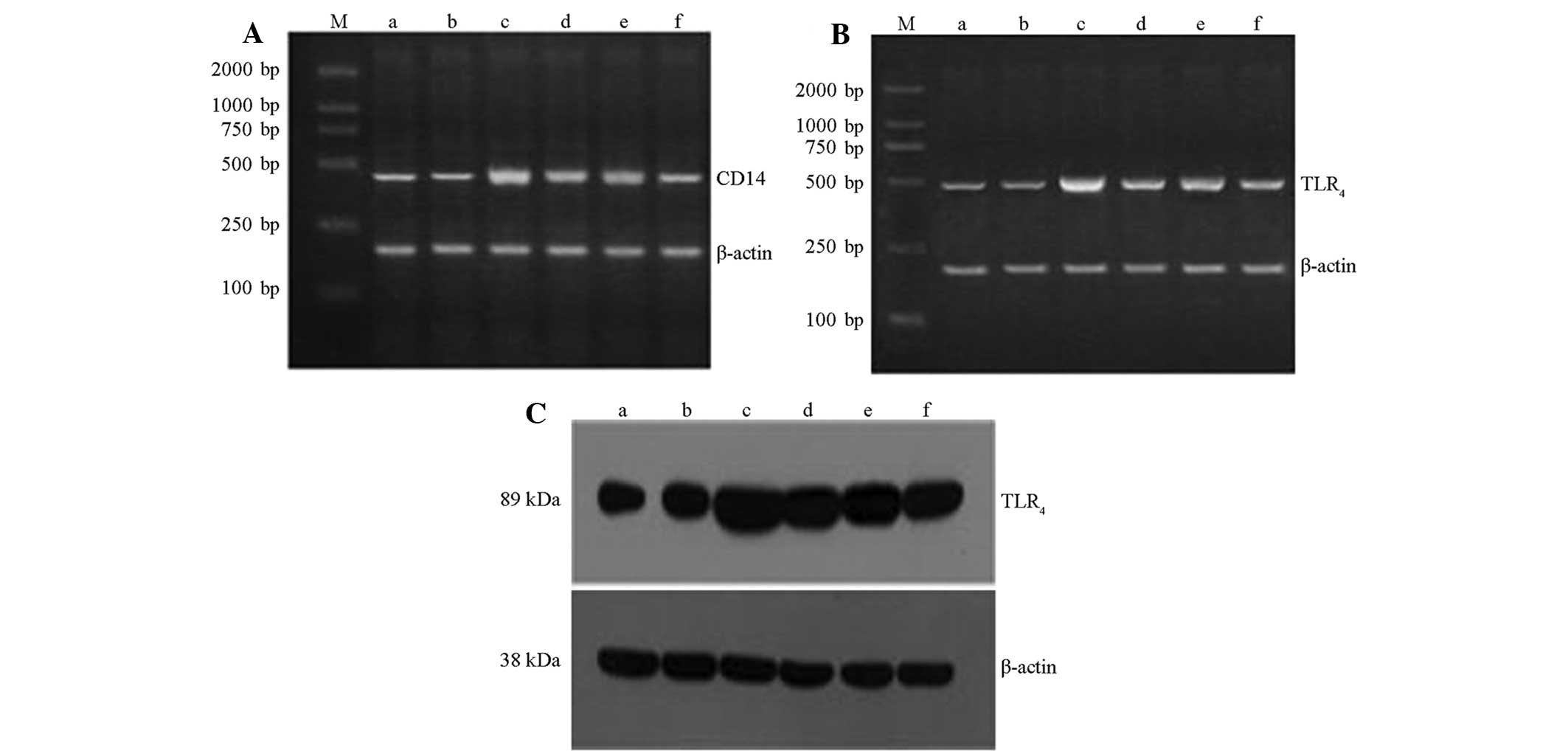 | Figure 3QGHXR downregulated CD14 and TLR4
expression in the liver tissue of the ALD rats. Reverse
transcription-polymerase chain reaction results for (A) CD14 and
(B) TLR4. (C) Western blot analysis results for TLR4. Lane M,
marker; lane a, blank group; lane b, CCl4 group; lane c,
model group; lane d, QGR group; lane e, HXR group; lane f, QGHXR
group. QGHXR, Qinggan Huoxue Recipe; CD, cell differentiation
antigen; TLR4, Toll-like receptor 4; ALD, alcoholic
liver disease; CC14, carbon tetrachloride; QGR, Qinggan
Recipe; HXR, Huoxue Recipe. |
QGHXR inhibits hepatic ERK activation and
NF-κB expression
The expression of hepatic p-ERK and NF-κB in the ALD
rats was upregulated compared with that of the normal and
CCl4 group (Fig. 4A and
B). ERK activation and NF-κB expression were inhibited in the
QGHXR group and no significant changes in p-ERK expression were
apparent. QGHXR showed greater inhibition of NF-κB expression when
compared with QGR.
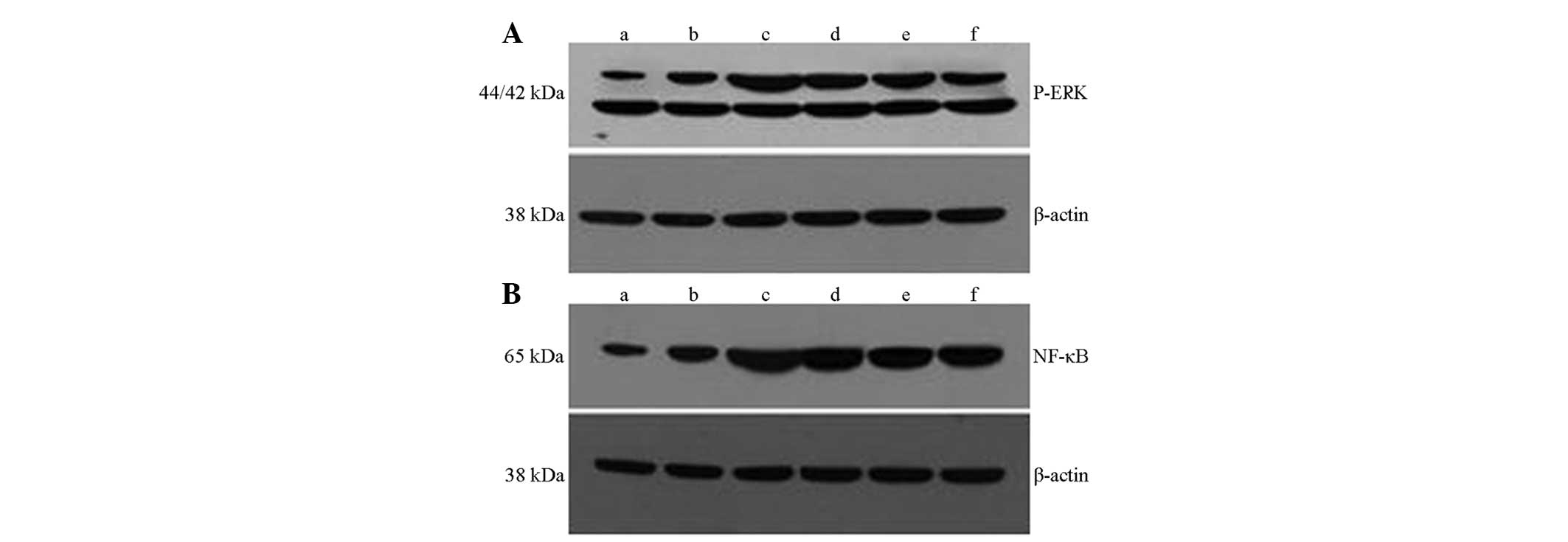 | Figure 4QGHXR inhibited hepatic ERK activation
and NF-κB expression in the liver tissue of the ALD rats. Western
blot analysis results for (A) p-ERK and (B) NF-κB. Lane M, marker;
lane a, blank group; lane b, cell differentiation antigen 14 group;
lane c, model group; lane d, Qinggan Recipe group; lane e, Huoxue
Recipe group; lane f, QGHXR group. p-ERK,
phosphorylated-extracellular regulated protein kinase; NF, nuclear
factor; QGHXR, Qinggan Huoxue Recipe. |
Discussion
There is increasing evidence indicating that Chinese
medicinal formulas may be adopted as therapeutic agents or
adjuvants in ALD treatment (6,11).
In the present study, a Wistar rat model, which has previously been
successfully adopted for the screening of therapeutic agents for
ALD (7), was used and QGHXR was
demonstrated to exert a range of pharmacological effects for
improving liver injury without extensive toxicity. The rule of QG
and HX were the basic therapy for ALD. QGHXR exerts effects on
treating ALD. Therefore we separated the whole formulation QGHXR
into QGR and HXR in order to determine their roles in ALD. In the
present study, a 0.25ml/kg of 25% CCl4 served as the
coefficient of alcohol as it reduces the model cycle time and
enhances its stability (6). The
CCl4 group was identical to the blank group except for
the marginally increased AST level (data not shown), which
indicated that alcohol is the predominant cause of liver
injury.
Chronic ethanol consumption leads to the elevation
of hepatic LPS levels that target CD14/TLR4 receptors,
which are also elevated following ethanol consumption, leading to
the production of pro-inflammatory cytokines, such as TNF-α, IL-6
and transforming growth factor-β1, and the subsequent activation of
hepatic stellate cells (HSCs) and liver fibrosis (12,13).
The following studies were conducted to investigate the underlying
mechanism of the effect of QGHXR on ALD.
The liver is composed of parenchymal cells, such as
hepatocytes and non-parenchymal cells, such as sinusoidal
endothelial cells, KCs, HSCs, dendritic cells and other
lymphocytes. Previous studies indicated that KCs were predominantly
involved in alcohol-mediated inflammation via LPS/TLR4
signaling-dependent mechanisms (14,15)
in addition to indicating that QGHXR and HXR inhibit KC
activation.
The currently accepted model of ALD indicates that
LPS promotes hepatic injury via the induction of KC activation,
resulting in the production of TNF-α and additional inflammatory
mediators (16,17). Significant evidence of the pivotal
role of TNF-α in alcohol-induced liver injury was identified
through a study, which used anti-TNF-α antibodies to prevent liver
injury in alcohol-fed rats (18)
as well as the observation that mice lacking a TNF type I receptor
did not develop ALD (19,20). QGHXR and HXR significantly reduced
TNF-α mRNA, however, QGR demonstrated no distinct effect; the TNF-α
protein expression was similar to that of TNF-α mRNA. QGHXR and HXR
protected the hepatocytes by reducing TNF-α generation and QGR
showed no significant effect on the TNF-α gene and protein
regulation, which was consistent with the results relating to the
regulation of KC activation.
Furthermore, immunohistochemistry and western blot
analysis indicated that QGHXR decreased CD14 expression in the
cytoplasm, but not QGR or HXR. CD14 is a
glycosylphosphatidylinositol-anchored protein that also exists in a
soluble form. It facilitates the transfer of LPS to the
TLR4/lymphocyte antigen 96 (MD2) receptor complex and
modulates LPS recognition (21,22).
A recent study demonstrated that the expression of cytokines, such
as TNF-α were inhibited following the addition of CD14 monoclonal
antibodies (23). In addition,
sensitivity to LPS increased 100–1,000 times following CD14 cDNA
transfection into low endotoxin-reactive cells that do not express
CD14 (24). Therefore, QGHXR
decreased the efficiency of endotoxin signal conduction, and
reduced the production of inflammatory factors and hepatocellular
injury by regulating CD14 expression.
TLR4 recognizes the LPS lipid A motif, a
suggested cofactor in the pathogenesis of ALD (25). Moreover, TLR4 is a
predominant component of the LPS recognition receptor complex,
which involves the co-receptors CD14, MD2 and the LPS binding
protein (LBP) (26). Studies in
knockout mouse models have shown that chronic alcohol feeding in
CD14-, TLR4- and LBP-deficient mice resulted in the
alleviation of alcohol-induced liver injury, indicating the
significance of the TLR4 pathway (27–29).
QGR, HXR and QGHXR significantly decreased TLR4;
however, QGHXR exhibited a superior effect compared with HXR and
QGR, which indicated that QGHXR decreased TLR4
expression and inhibited further signal transmission.
Changes in related molecules, such as p-ERK and
NF-κB, were measured to determine the influence of QGHXR on the
LPS-KC pathway. NF-κB is a central regulator of cellular stress in
all liver cell types, forms p65/p50 heterodimers in macrophages and
binds to the promoter region of various pro-inflammatory genes,
which results in gene transactivation (30). The hepatic macrophage expression of
pro-inflammatory mediators is predominantly regulated by NF-κB and
murine models of chronic alcohol administration demonstrated
increased NF-κB DNA binding in the liver (31). Chronic alcohol intake is
hypothesized to prime the liver via sustained NF-κB activation and
induction of basal and LPS-stimulated TNF-α. LPS recognition
activates the MAPK family members, including ERK1/2, p38 and c-Jun
N-terminal kinase, resulting in TNF-α production (32). In addition, chronic alcohol intake
activates LPS-induced ERK1/2 activation, which contributes to TNF-α
expression in murine hepatic macrophages (33). Recently Karki et al found
that extract of buckwheat sprouts inhibited pro-inflammatory
mediators IL-6 and TNF-α production in
lipopolysaccharide-stimulated macrophages (RAW264.7) (34). Nwozo et al evaluated the
protective effects of oils from Zingiber officinale (ginger)
and Curcuma longa (turmeric) on acute ethanol-induced fatty
liver in male Wistar rats (35).
The results of the present study showed that QGHXR and its separate
components, HXR and QGR, significantly decreased p-ERK and NF-κB
expression, thus indicating that the therapeutic effect of QGHXR on
ALD rats may be due to p-ERK and NF-κB downregulation.
In the present study, QGHXR was indicated to be a
potent sensitizer for ALD in experimental rats. QGHXR regulated the
membrane receptor, protein kinase, NF and abnormal function of the
cytokine network via the LPS-KC pathway. Chinese herbal medicine
appears to manifest its activity slowly, therefore, improving the
agent administration strategy due to an earlier administration time
and a longer therapeutic period, which may enhance the performance
of the agent.
There were certain limitations in the present study;
the exact mechanism of QGHXR protection against ALD was not
identified; thus, further investigation into the stimulating effect
is required. As a result of using rats, the ability to obtain
robust evidence was limited, therefore, large-scale multicentric
placebo-controlled prospective studies are required to verify the
results. Regardless of these limitations, the present study
provided preliminary data to support future QGHXR evaluations.
Through observation of the multi-element, multichannel and
multitarget action characteristics of Chinese medicine, QGHXR may
be screened and the formulations simplified to establish the
foundation for identifying their composition and active
components.
In conclusion, the Chinese medicinal formula, QGHXR,
is a potential treatment for ALD. The present study provided
further clarification of the mechanism for QGHXR as a treatment for
ALD via the LPS-KC pathway. Although the underlying mechanisms that
govern these effects remain undetermined, the available evidence
collectively demonstrated that QGHXR may be of therapeutic benefit
in a clinical setting, indicating its potential use as an agent for
protecting against ALD.
Acknowledgements
The present study was supported by the Supporting
Project for Elitists in the New Century of the Ministry of
Education (grant no. NCET07-0563), the National Nature Science
Foundation of China (grant nos. 81202979) and the Shanghai Leading
Academic Discipline Project (grant nos. J50305 and E3008).
References
|
1
|
Rao RK, Seth A and Sheth P: Recent
advances in alcoholic liver disease I. Role of intestinal
permeability and endotoxemia in alcoholic liver disease. Am J
Physiol Gastrointest Liver Physiol. 286:G881–G884. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Albano E: Oxidative mechanisms in the
pathogenesis of alcoholic liver disease. Mol Aspects Med. 29:9–16.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nagy LE: Recent insights into the role of
the innate immune system in the development of alcoholic liver
disease. Exp Biol Med (Maywood). 228:882–890. 2003.PubMed/NCBI
|
|
4
|
Mandrekar P and Szabo G: Signalling
pathways in alcohol-induced liver inflammation. J Hepatol.
50:1258–1266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
O’Shea RS, Dasarathy S and McCullough AJ;
Practice Guideline Committee of the American Association for the
Study of Liver Diseases; Practice Parameters Committee of the
American College of Gastroenterology. Alcoholic liver disease.
Hepatology. 51:307–328. 2010.
|
|
6
|
Ji G, Wang YQ and Cao CL: Clinical study
on treatment of alcoholic liver disease by qinggan huoxue recipe.
Zhongguo Zhong Xi Yi Jie He Za Zhi. 24:13–16. 2004.(In
Chinese).
|
|
7
|
Lu YL, Wang M and Ji G: Determination of
Puerarin, Baicalin, Baicalein and Wogonin in Qinggan Huoxue Recipe
by HPLC. Chin Pharm J. 45:299–301. 2010.
|
|
8
|
Wang L, Ji G, Zheng PY and Long AH:
Establishment of a rat model of alcoholic liver fibrosis induced by
complex factors. Zhong Xi Yi Jie He Xue Bao. 4:281–284. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Diehl AM, Goodman Z and Ishak KG: Alcohol
like disease in nonalcoholics. A clinical and histologic comparison
with alcohol-induced liver injury. Gastroenterology. 95:1056–1062.
1988.PubMed/NCBI
|
|
10
|
Wang TL: The classification, staging and
and categorization of the pathologic diagnostic criteria of
alcoholic liver disease. Zhong Hua Gan Zang Bin Za Zhi. 5:312–313.
2001.
|
|
11
|
Lv XH, Zhou LP, Liu DP, et al: Traditional
Chinese medicine Kang Xian Fu Fang I is effective for prophylaxis
and treatment of alcoholic liver disease in rats. Hepatobiliary
Pancreat Dis Int. 6:182–187. 2007.PubMed/NCBI
|
|
12
|
Enomoto N, Ikejima K, Yamashina S, et al:
Kupffer cell sensitization by alcohol involves increased
permeability to gut-derived endotoxin. Alcohol Clin Exp Res. 25(6
Suppl): 51S–54S. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wheeler MD and Thurman RG: Up-regulation
of CD14 in liver caused by acute ethanol involves oxidant-dependent
AP-1 pathway. J Biol Chem. 278:8435–8441. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hines IN and Wheeler MD: Recent advances
in alcoholic liver disease III. Role of the innate immune response
in alcoholic hepatitis. Am J Physiol Gastrointest Liver Physiol.
287:G310–G314. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Purohit V, Gao B and Song BJ: Molecular
mechanisms of alcoholic fatty liver. Alcohol Clin Exp Res.
33:191–205. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thurman RG: II. Alcoholic liver injury
involves activation of Kupffer cells by endotoxin. Am J Physiol.
275:G605–G611. 1998.PubMed/NCBI
|
|
17
|
Song Z, Zhou Z, Uriarte S, et al:
S-adenosylhomocysteine sensitizes to TNF-alpha hepatotoxicity in
mice and liver cells: a possible etiological factor in alcoholic
liver disease. Hepatology. 40:989–997. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tilg H, Jalan R, Kaser A, et al:
Anti-tumor necrosis factor-alpha monoclonal antibody therapy in
severe alcoholic hepatitis. J Hepatol. 38:419–425. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yin M, Wheeler MD, Kono H, et al:
Essential role of tumor necrosis factor alpha in alcohol-induced
liver injury in mice. Gastroenterology. 117:942–952. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Olleros ML, Martin ML, Vesin D, et al: Fat
diet and alcohol-induced steatohepatitis after LPS challenge in
mice: role of bioactive TNF and Th1 type cytokines. Cytokine.
44:118–125. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takeda K and Akira S: Toll-like receptors
in innate immunity. Int Immunol. 17:1–14. 2005. View Article : Google Scholar
|
|
22
|
Dunne A and O’Neill LA: Adaptor usage and
Toll-like receptor signaling specificity. FEBS Lett. 579:3330–3335.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Verbon A, Dekkers PE, ten Hove T, et al:
IC14, an anti-CD14 antibody, inhibits endotoxin-mediated symptoms
and inflammatory responses in humans. J Immunol. 166:3599–3605.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee JD, Kato K, Tobias PS, Kirkland TN and
Ulevitch RJ: Transfection of CD14 into 70Z/3 cells dramatically
enhances the sensitivity to complexes of lipopolysaccharide (LPS)
and LPS binding protein. J Exp Med. 175:1697–1705. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Saitoh S, Akashi S, Yamada T, et al: Lipid
A antagonist, lipid IVa, is distinct from lipid A in interaction
with Toll-like receptor 4 (TLR4)-MD-2 and ligand-induced
TLR4 oligomerization. Int Immunol. 16:961–969. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Visintin A, Mazzoni A, Spitzer JA and
Segal DM: Secreted MD-2 is a large polymeric protein that
efficiently confers lipopolysaccharide sensitivity to Toll-like
receptor 4. Proc Natl Acad Sci USA. 98:12156–12161. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Uesugi T, Froh M, Arteel GE, Bradford BU
and Thurman RG: Toll-like receptor 4 is involved in the mechanism
of early alcohol-induced liver injury in mice. Hepatology.
34:101–108. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Uesugi T, Froh M, Arteel GE, et al: Role
of lipopolysaccharide-binding protein in early alcohol-induced
liver injury in mice. J Immunol. 168:2963–2969. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yin M, Bradford BU, Wheeler MD, et al:
Reduced early alcohol-induced liver injury in CD14-deficient mice.
J Immunol. 166:4737–4742. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ghosh S: Regulation of inducible gene
expression by the transcription factor NF-kappaB. Immunol Res.
19:183–189. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wheeler MD, Yamashina S, Froh M, Rusyn I
and Thurman RG: Adenoviral gene delivery can inactivate Kupffer
cells: role of oxidants in NF-kappaB activation and cytokine
production. J Leukoc Biol. 69:622–630. 2001.PubMed/NCBI
|
|
32
|
Yao J, Mackman N, Edgington TS and Fan ST:
Lipopolysaccharide induction of the tumor necrosis factor-alpha
promoter in human monocytic cells. Regulation by Egr-1, c-Jun, and
NF-kappaB transcription factors. J Biol Chem. 272:17795–17801.
1997. View Article : Google Scholar
|
|
33
|
Shi L, Kishore R, McMullen MR and Nagy LE:
Chronic ethanol increases lipopolysaccharide-stimulated Egr-1
expression in RAW 264.7 macrophages: contribution to enhanced tumor
necrosis factor alpha production. J Biol Chem. 277:14777–14785.
2002.
|
|
34
|
Karki R, Park CH and Kim DW: Extract of
buckwheat sprouts scavenges oxidation and inhibits pro-inflammatory
mediators in lipopolysaccharide-stimulated macrophages (RAW264.7).
J Integr Med. 11:246–252. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nwozo SO, Osunmadewa DA and Oyinloye BE:
Anti-fatty liver effects of oils from Zingiber officinale
and Curcuma longa on ethanol-induced fatty liver in rats. J
Integr Med. 12:59–65. 2014.PubMed/NCBI
|














