Introduction
Radiotherapy is a crucial therapy for the treatment
of thorax-associated neoplasms. However, during a course of
treatment with radiotherapy, the lung tissue is inevitably damaged
by the radiation dose, which is above the biologically tolerable
threshold. Radiation-induced lung injury (RILI) is a common
complication of radiotherapy and a key dose-limiting factor, which
may reduce the probability of tumor control and affect the
patient's quality of life (1). The
pathology of RILI is complex, and includes radiation-induced
pneumonitis and fibrosis (2).
Although intensive study in the previous decades has elucidated a
number of the associated underlying mechanisms, involving, for
example, cells and cytokines, the specific mechanism remains
unclear (3).
The alveolar wall extracellular matrix (ECM)
consists of a basement membrane and an interstitial matrix. It acts
as a physical barrier to the passage of macromolecules and cells,
provides physical tissue support and anchorage sites for cells, and
serves as a substrate for cell migration and a medium for
diffusible cell signaling molecules. Matrix metalloproteinases
(MMPs) are a large family of related proteolytic enzymes that
specifically degrade ECM components. MMP2 and MMP9, also known as
gelatinases, are key enzymes in basement membrane degradation.
The alveolar epithelium is composed of type I and II
cells, which are morphologically and functionally distinct.
Alveolar epithelial type I (AE1) cells are the first type to be
affected and to undergo apoptosis when injury occurs. Alveolar
epithelial type II (AE2) cells synthesize and secrete pulmonary
surfactant, and also proliferate and transdifferentiate into AE1
cells, which play an important role in maintaining the structure
and function of the alveoli. AE2 cell injury has been associated
with the development of lung fibrosis (4), and AE2 cells from patients with
idiopathic pulmonary fibrosis and other experimental pulmonary
fibrosis diseases express high levels of epithelial-mesenchymal
transition (EMT)-associated protein markers (4,5). The
process of EMT results in a loss of epithelial polarity,
cytoskeletal reorganization, downregulation of epithelial marker
expression (e.g., E-cadherin), the acquisition of mesenchymal
markers [e.g., α-smooth muscle actin (α-SMA) and vimentin], and an
increase in cell invasiveness and migration (6). A recent study observed that
pro-surfactant protein-c (pro-SP-c) and α-SMA are co-localized in
the irradiated lung alveoli of FVB/N mice, suggesting that the AE2
cells achieved a mesenchymal phenotype (7). However, the anatomical structure of the
rodent lung differs considerably from that of humans. Furthermore,
the radiation response in the lung varies with species and strain
(8). For Bama minipigs, the
single-dose radiation dose-effect curve is comparable to that in
humans, as is the anatomical structure and physiology of the lung
(9). Therefore, the present study
aimed to determine whether AE2 cells are able to transdifferentiate
into a mesenchymal-like phenotype by dynamically observing the
protein expression levels of surfactant protein A (SP-A),
E-cadherin, α-SMA and MMPs. In addition, the present studied aimed
to elucidate the possible role of MMP2 and MMP9 in RILI using Bama
minipigs as experimental subjects.
Materials and methods
Minipigs and collection of lung
specimens
This study was approved by the Committee on the
Ethics of Animal Experiments of Guangxi Medical University (Permit
No. SCXKGUI 2009–0002; Nanning, China). All surgery was performed
under sodium pentobarbital anesthesia, and all animals were treated
humanely.
Male Bama minipigs were supplied by the Experimental
Animal Center of Guangxi Medical University. Forty minipigs, aged
2–3 months and weighing 10–15 kg, were randomly divided into
control and irradiated groups (n=20 per group). Following the
intraperitoneal administration of anesthesia, using 3% sodium
pentobarbital (30 mg/kg), minipigs in the irradiated group received
a single 15-Gy dose of right-thoracic 60Co gamma
irradiation (field, 9×10 cm; source-surface distance, 80 cm;
absorbed dose, 3 Gy/min), using a GWXJ80 60Co
radiotherapy treatment unit (Nuclear Power Institute of China,
Chengdu, China). The minipigs in the control group received sham
irradiation. Five minipigs in each group were randomly sacrificed
at 4, 8, 12 and 24 weeks post-irradiation, respectively.
Subsequently, the thorax was opened and right lung tissue was
resected under anesthesia. The upper lobe was fixed in 10% neutral
formalin for histological and immunohistochemical analysis, and the
lower lobe was snap frozen in liquid nitrogen and stored at −80°C
until use.
Histopathology and
immunohistochemistry
Lung sections (4 µm) were subjected to hematoxylin
and eosin (H&E) and Masson's trichrome staining. The sections
were deparaffinization and antigen retrieval was conducted using a
high pressure method with sections on ice in 0.01M citrate bufer.
Next, sections were incubated with mouse monoclonal α-SMA
antibodies (1:400; ab8211 Abcam, Cambridge, England) at 4°C
overnight, and then were assayed using a streptavidin-peroxidase
kit (Beyotime Institute of Biotechnology, Haimen, China), according
to the manufacturer's instructions. Positive cells displayed a
brownish-yellow cytoplasm, indicating that they were smooth muscle
cells or activated myofibroblasts that synthesize collagen and
certain components of the ECM.
Double immunofluorescence
staining
Double-color immunofluorescence analysis was used to
identify α-SMA and SP-A expression in the lung AE2 cells. Briefly,
following deparaffinization and antigen retrieval, sections were
blocked using normal donkey serum (Molecular Probes Life
Technologies, Carlsbad, CA, USA) for 20 min at 37°C, then incubated
with a mixture of anti-α-SMA (1:100, Abcam) and anti-SP-A (1:100,
Abbiotec, San Diego, CA, USA) antibodies at 4°C overnight. Nuclei
were counterstained with 4–6-diamidino-2-phenylindole (Beyotime
Institute of Biotechnology) for 8 min, and the sections analyzed
using a Nikon A1 laser scanning confocal microscope (Nikon
Corporation, Tokyo, Japan).
Western blot analysis
Lung tissue (40–80 mg) was rinsed, shredded and
homogenized in 600 µl radioimmunoprecipitation lysis buffer with 1
mM phenylmethylsulfonyl fluoride (PMSF), and the supernatant was
quantified using a bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology). Following 10% SDS-polyacrylamide gel
electrophoresis with 40 µg sample per lane, the proteins were
transferred to a polyvinylidene difluoride (PVDF) film (EMD
Millipore, Billerica, MA, USA). The film was then blocked with 1X
Tris-buffered saline with Tween 20 (TBST) for 1 h at room
temperature, and incubated with primary antibodies against GAPDH
(1:5,000; ABS16), MMP2 (1:500; MAB3308), MMP9 (1:2,000; AB6001;
Millipore), TGFβ1 (1:50; 5559-100; BioVision, Inc., Milpitas, CA,
USA), E-cadherin (1:500), vimentin (1:1,000; ab8069; Abcam) and
SP-A (1:200; Abbiotec) at 4°C overnight. After washing with
phosphate-buffered saline, the sections were incubated with donkey
anti-mouse Alexa Fluor 594-conjugated (A21202) and anti-rabbit
Alexa Fluor 488-conjugated secondary antibodies (A21206; 1:5,000;
Molecular Probes Life Technologies) at room temperature for 1 h.
After washing with 1X TBST, the film was incubated with anti-mouse
or anti-rabbit IgG horseradish peroxidase-labeled antibodies (Asbio
Technology, Inc., Guangzhou, China) at room temperature for 1 h.
Finally, the film was visualized using enhanced chemiluminescence
according to the manufacturers instructions (ECL+; Pierce
Biotechnology, Inc., Rockford, IL, USA) and scanned using an
ImageQuant LAS500 imaging system (GE Healthcare Life Sciences,
Chalfont, UK). Scanned films were analyzed semiquantitatively for
the densitomateic signal of bands on the western blot using
software included with the ImageQuant LAS500.
Gelatin zymography
Lung tissue (50 mg) was rinsed with normal saline
and shredded using operating scissors, then homogenized in 600 µl
lysis buffer (150 mM NaCl, 10 mM CaCl2, 10 g/l Triton
X-100 and 1 mM PMSF). Protein concentration was quantified using an
identical protocol to that of the western blot analysis. Following
8% SDS-polyacrylamide gel electrophoresis with 40 µg sample per
lane, the gel was stained according to the manufacturer's
instructions using an MMP zymography assay kit (Applygen
Technologies, Inc., Beijing, China). The gel was then scanned using
an Odyssey two-color infrared scanning laser imaging system and
analyzed using Odyssey software, version 2.1 (LI-COR Biosciences,
Lincoln, NE, USA).
Statistical analysis
SPSS software, version 13.0 (SPSS, Inc., Chicago,
IL, USA) was used for data processing, and data are expressed as
the mean ± standard deviation. The homogeneity of the variance was
tested for the measurement data, and one-way analysis of variance
was used for group comparisons. A least significant difference test
was used for two groups with variance homogeneity, whereas a
Games-Howell test was used for two groups with variance
heterogeneity. P<0.05 was considered to indicate a statistically
significant difference.
Results
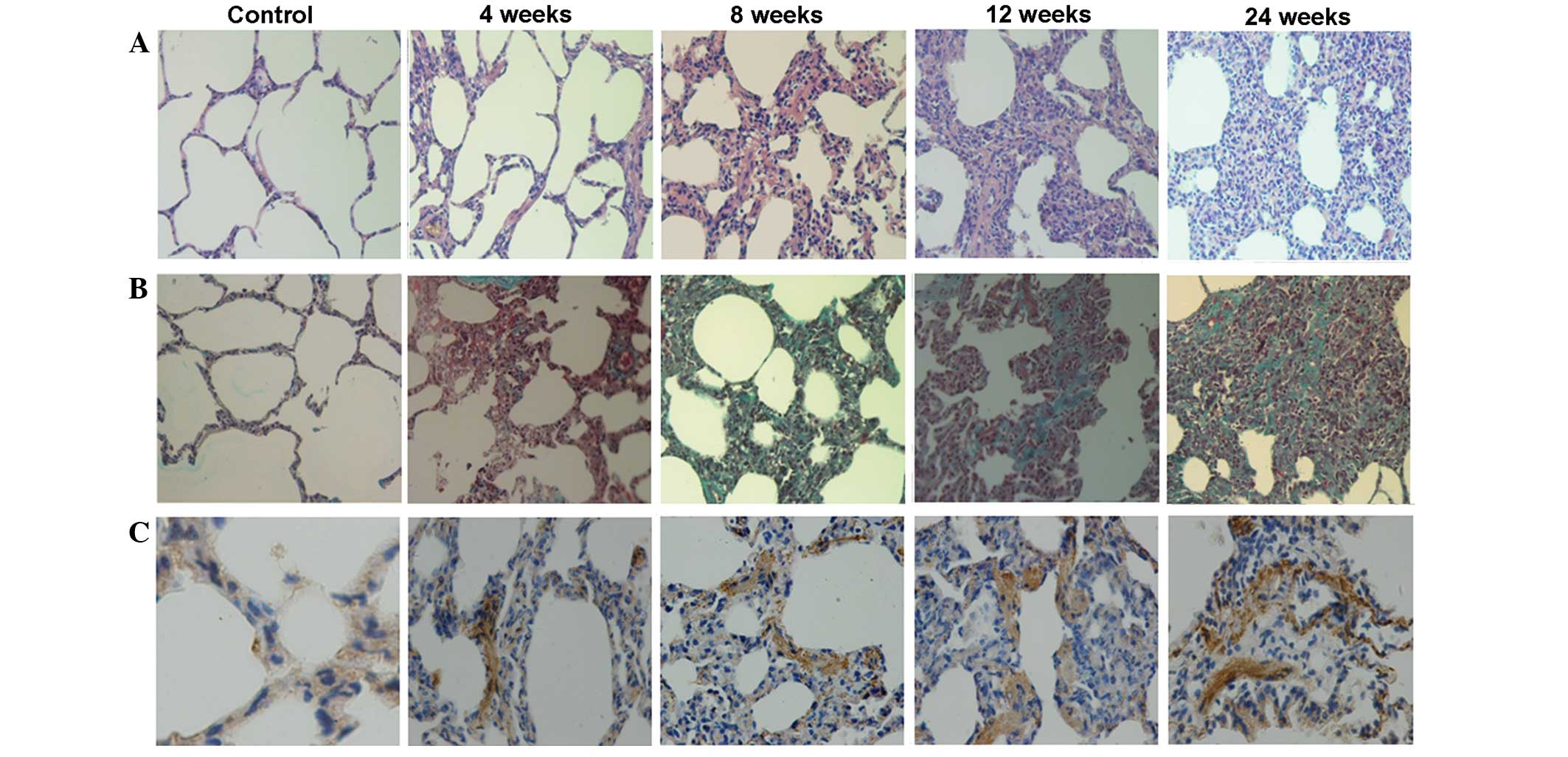
Histological changes in lung
tissue
In the control lung tissue, the structure was clear,
the alveolar cavity maintained its integrity, and there was no
inflammatory cell infiltration. By contrast, 4 weeks after
irradiation, the irradiated lung tissue showed pulmonary
interstitial and capillary hyperemia, and diffuse accumulation of
inflammatory cells. At 8 and 12 weeks after irradiation, thickened
alveolar walls, narrowed alveolar spaces, an increase in the number
of fibroblasts compared with that in the non-irradiated tissue, and
fibrous tissue hyperplasia were observed. Inflammatory cells were
also observed in the irradiated lung tissue. At 24 weeks after
irradiation, multiple fibrotic lesions were present in the
irradiated lungs (Fig. 1A). Masson's
trichrome staining indicated increasing quantities of collagen
fiber deposition in the irradiated lung tissues over time (Fig. 1B). Immunohistochemical staining for
α-SMA, a marker of smooth muscle cells/myofibroblasts, showed an
increased accumulation of myofibroblasts in the irradiated lungs
(Fig. 1C).
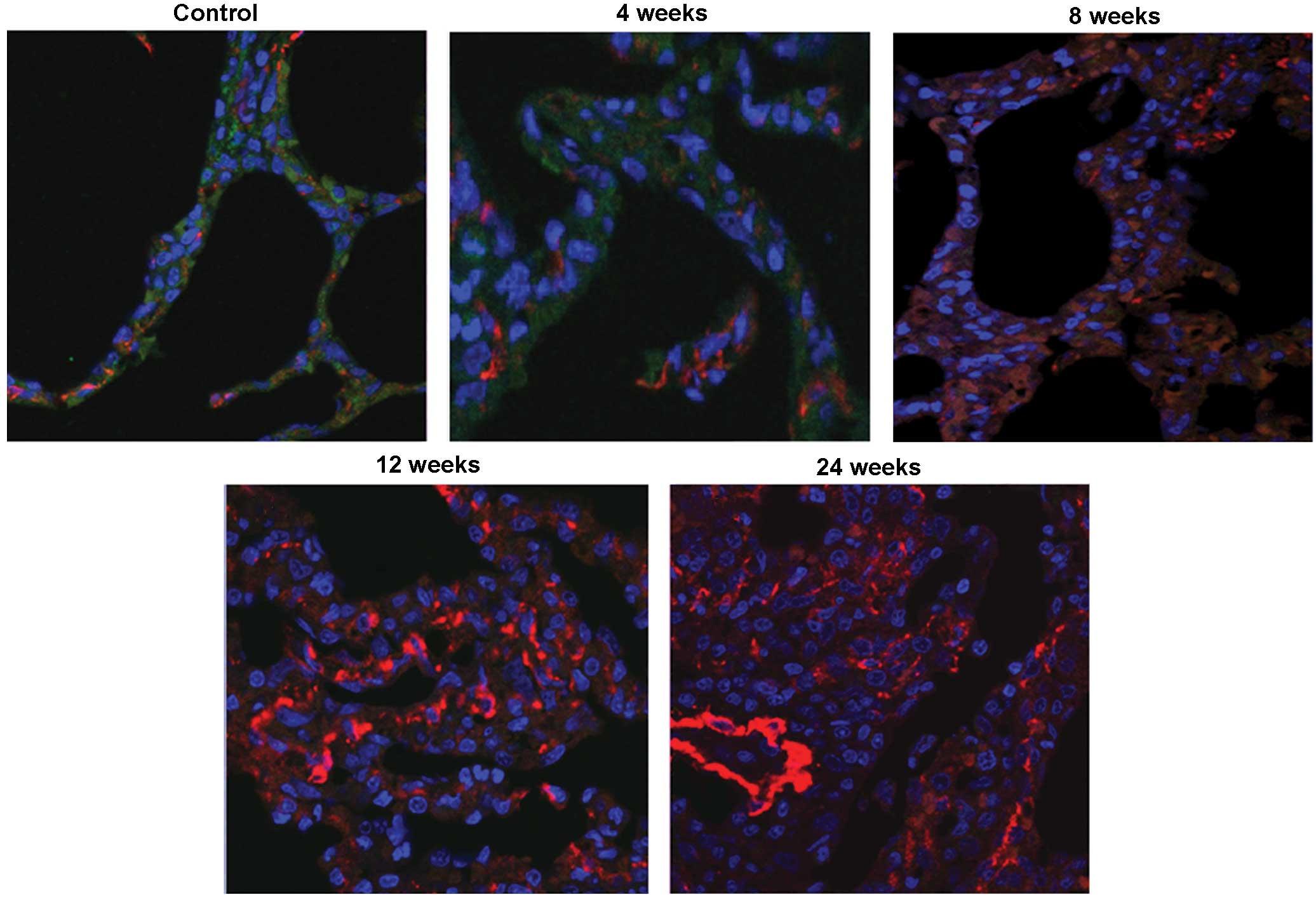
Co-localization of SP-A and α-SMA
protein in the alveoli
A previous study identified pro-SP-c and α-SMA
protein co-localization in the lungs of FVB/N mice following
irradiation, suggesting that EMT may occur in rodents with
radiation lung fibrosis (7).
Therefore, the right lung tissue of the Bama minipigs was stained
for SP-A and α-SMA protein following irradiation. The two proteins
were observed to be co-localized in the alveoli in the irradiated
lungs, and this co-localization was most evident at week 8
post-irradiation. Co-localization of SP-A and α-SMA was not
observed in the control lung alveoli (Fig. 2), suggesting that radiation caused
the trans-differentiation of AE2 cells to a mesenchymal-like
phenotype.
SP-A, TGFβ1, MMP2, MMP9, vimentin and
E-cadherin protein expression in lung tissue
SP-A protein expression levels were reduced and
TGFβ1 protein expression levels were increased following
irradiation in a time-dependent manner.
E-cadherin, a calcium-dependent adhesion molecule,
mediates the adhesion and tight junctions between epithelial cells.
E-cadherin protein expression levels in the irradiated group were
reduced in a time-dependent manner following irradiation. However,
the protein expression levels of vimentin, a mesenchymal marker,
were markedly increased following the radiation treatment.
At each time-point after irradiation, pairwise
comparisons revealed that differences between the irradiation and
control groups were statistically significant (SP-A and TGFβ1,
P<0.01; E-cadherin and vimentin, P<0.05).
Irradiation significantly increases
MMP2 and MMP9 protein expression levels
MMP9 protein expression was only moderately
increased at 12 weeks post-irradiation, but increased markedly at
24 weeks (week 24 vs. weeks 4, 8 and 12; P<0.01). For MMP2,
there was no significant difference in protein expression between
weeks 4 and 12 (P>0.05), and pairwise comparisons revealed
statistically significant differences between the groups at all
time-points (P<0.01; Fig. 3).
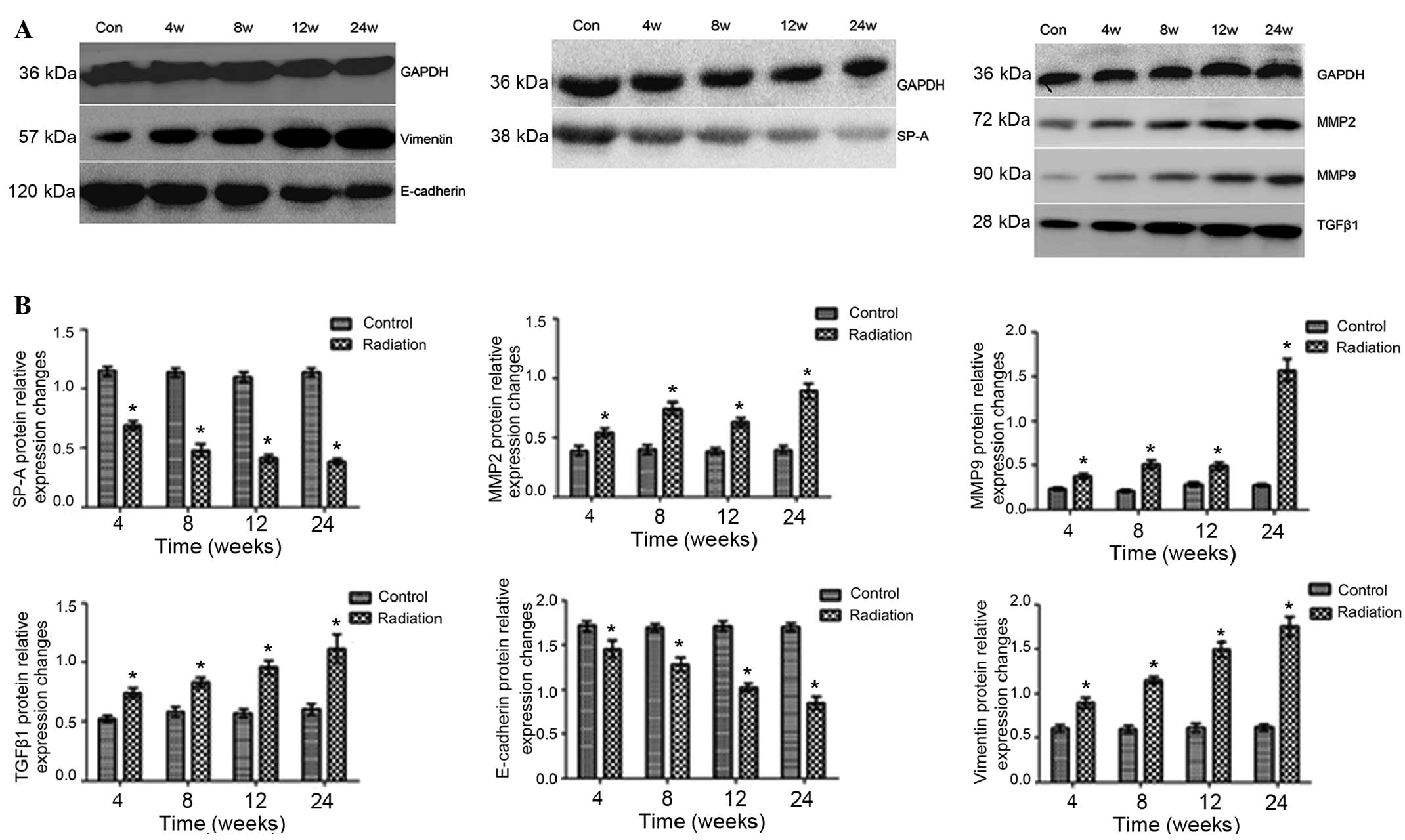 | Figure 3.Irradiation differentially modulates
SP-A, MMP2, MMP9, TGFβ1, E-cadherin and vimentin protein expression
in the lung. SP-A and E-cadherin protein expression in lung tissue
is reduced in a time-dependent manner after irradiation, while
irradiation significantly increased TGFβ1, vimentin, MMP2 and MMP9
protein expression in lung tissue. (A) Representative western
blotting results. (B) Densitometry was used to quantify the protein
in each lane. All values were normalized against GAPDH values
obtained on the same gel, with the exception of SP-A due to its
molecular weight. *P<0.01 vs. control. Data are presented as the
mean ± standard deviation. Con, control; w, week; MMP, matrix
metalloproteinase; TGFβ1, transforming growth factor β1; SP-A,
surfactant protein A. |
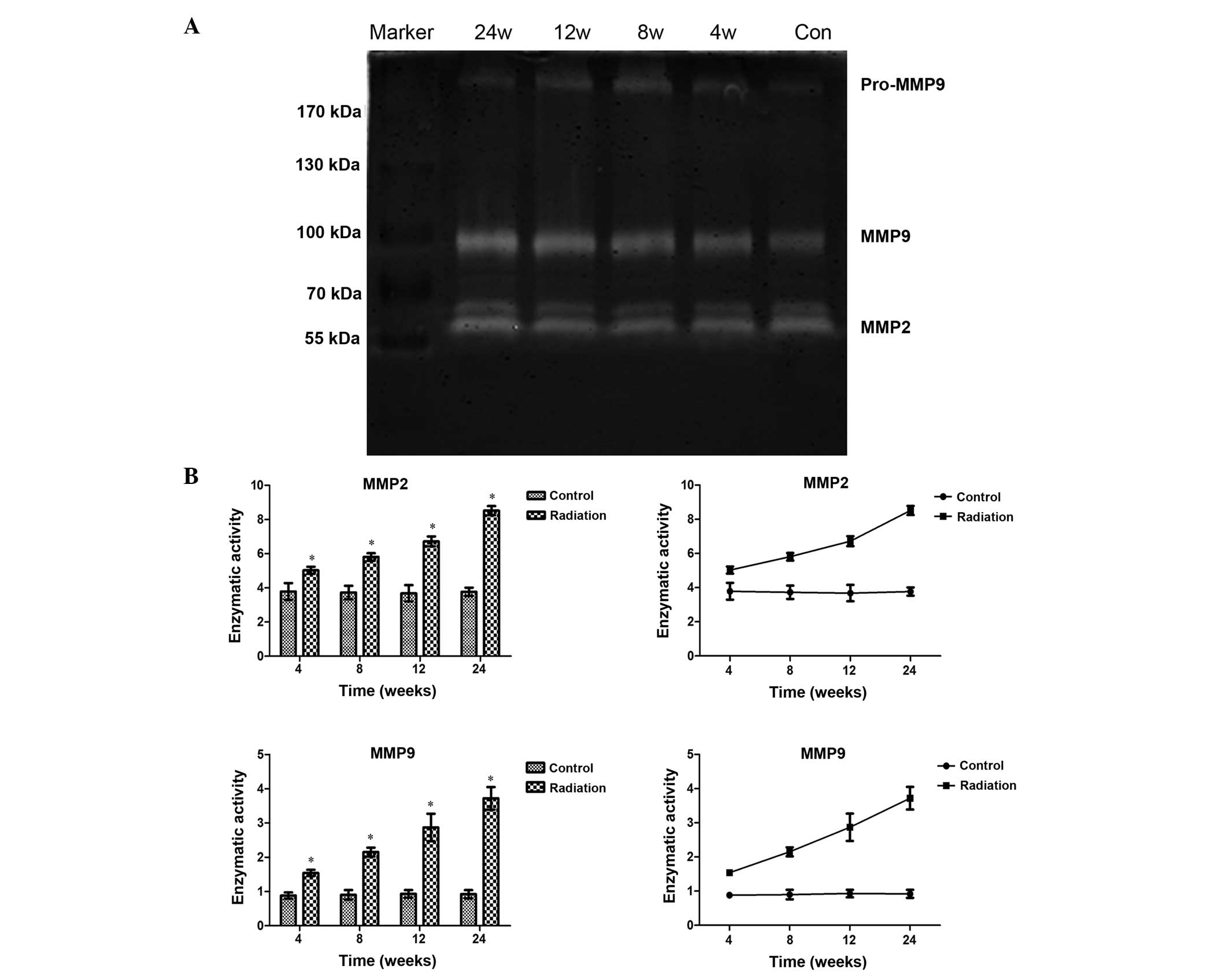
MMP2 and MMP9 gelatinase activity in
lung tissue
After irradiation, increased gelatinolytic activity
was observed in the lung tissue at each time-point compared with
that in the control lung tissue, and quantitative analysis
demonstrated the time-dependence of the increase in gelatinase
activity (Fig. 4). At each
time-point after irradiation, pairwise comparisons revealed that
differences between the irradiation and control groups were
statistically significant (MMP2, P<0.01; MMP9, P<0.05).
Discussion
Currently, rodents are typically used to establish
animal models of RILI (10,11). However, the sensitivity of the mouse
lung to RILI is strain-dependent (8), and minipigs do not exhibit this
strain-specific sensitivity (9). The
primary pathological features of radiation pneumonitis, such as
interstitial edema, capillary hyperemia, and extensive inflammatory
cell infiltration, usually appear at 1–3 months after irradiation.
Lung fibrosis may occur at 6–12 months after irradiation, and is
characterized by excessive fibroblast proliferation and massive
collagen deposition (12). In the
present study, radiation pneumonitis was observed at 4 weeks (1
month) post-irradiation in minipigs after receiving a single 15-Gy
dose of irradiation to the right thorax. At 8 weeks, thickening of
the alveolar walls, fibroblasts and inflammatory cells were
observed. At 24 weeks (6 months), multiple fibrotic lesions were
observed in the irradiated lungs (Fig.
1). These results indicate that an animal model of RILI had
been successfully established, and that radiation pneumonitis and
fibrosis were not completely separate, but that there was a certain
degree of overlap at 8 weeks after irradiation.
Alveolar cells are crucially involved in the
maintenance of lung homeostasis. AE1 cells, a terminally
differentiated type of cell, are susceptible to injury; however,
AE2 cells, are able to proliferate and differentiate, and are
considered to be the stem cells of the alveolar epithelia (13). Following damage to the alveolar
epithelium, AE2 cells are able to self-renew and transdifferentiate
into AE1 cells in order to maintain the integrity of the alveolar
epithelium. SP-A, a water-soluble surfactant protein, is secreted
by AE2 and Clara cells, and is a key innate immune molecule in the
lung. Alcorn and Wright (14)
observed that SP-A inhibited inflammatory cytokine production
through multiple mechanisms and alleviated lung injury. The
transcription regulator nuclear factor-κB is activated by SP-A, and
directly regulates MMP9 expression and indirectly induces MMP2
transcription via the transmembrane activation of p53 protein
(15). Therefore, SP-A may cause an
imbalance the inflammatory/anti-inflammatory medium if inflammatory
cytokines in the lung are increased, and affect the
protease/antiprotease balance favoring an abnormal breakdown of ECM
components. Previous studies on RILI have indicated that SP-A
levels are decreased in bronchoalveolar lavage fluid (16), but increased in serum (17), a difference that may be associated
with lung injury. In the present study, SP-A protein expression in
lung tissue was reduced in a time-dependent manner (Fig. 3), which suggests that the changes in
AE2 cells induced by irradiation lead to a reduction in the
secretion of surfactants, and promote inflammatory cytokine
production, thus influencing MMP2 and MMP9 expression. This
situation favors the development of lung injury. In addition, the
results of the present study show that the SP-A protein expression
level in lung tissue is markedly reduced at week 4 after
irradiation, which suggests that SP-A may be used as a predictor of
RILI.
A key event in fibrosis is the activation of
fibroblasts and their conversion into myofibroblasts, which is
characterized by α-SMA expression, cytokine production, and
crucially, a increased capacity to produce ECM components (18). The origins of myofibroblasts may be
divided into three categories: Recruitment and differentiation of
resident cells, bone marrow-derived ‘fibrocytes’ and EMT. A study
has demonstrated that myofibroblasts may additionally originate
from resident epithelial cells that undergo EMT (19). An in vitro study indicated
that the irradiation of RLE-6TN cells induced their transition from
an epithelial to a mesenchymal phenotype, which was mediated by the
ROS/ERK/GSK-3β/Snail pathway (20).
Similarly, α-SMA and pro-SP-c co-expression has been detected in
the AE2 cells of FVB/N mice following thoracic radiation (7). In the present study, α-SMA protein
expression levels were significantly increased after irradiation
(Fig. 1C). A significant reduction
in the expression of the epithelial cell marker E-cadherin and a
concomitant increase in the expression of vimentin, a mesenchymal
marker, were detected after irradiation (Fig. 3). In addition, double
immunofluorescence staining identified α-SMA and SP-A protein
co-localization in irradiated lung alveoli, which was highest at
week 8 after irradiation (Fig. 2),
suggesting that AE2 cells had obtained a mesenchymal phenotype.
These data indicate that an AE2-to-mesenchymal transition may occur
in the irradiated lungs of minipigs and that EMT is a mechanism
involved in RILI.
TGFβ1 is released locally from damaged parenchymal
cells and inflammatory cells, and is central to the generation of
myofibroblasts and EMT (21).
Following activation, myofibroblasts themselves begin to secrete
TGFβ1 and are thereby able to sustain their own activation through
a self-stimulatory mechanism, which facilitates the
auto-perpetuating process characteristic of fibrosis. In addition,
TGFβ1 is a potent stimulus for the production of ECM components,
such as collagen (22), and
elevation of TGFβ1 levels in the plasma of patients during the 4th
week of radiotherapy is significantly predictive of RILI (23). In the present study, the continuous
increase in the TGFβ1 protein expression in the irradiated lungs of
minipigs further indicates the key function served by TGFβ1 in the
pathogenesis of RILI.
MMPs that degrade ECM components may paradoxically
promote collagen deposition. Following lung injury, MMP2 and MMP9
are released from damaged epithelial cells, inflammatory cells or
activated myofibroblasts; they degrade and remodel the ECM, and
promote cellular migration and activate cytokines such as TGFβ1,
TNF-α and IL-1β (24,25). TGFβ1 is a key profibrogenic cytokine,
which has been implicated as a primary fibrosis trigger in various
tissues (26). This suggests that
MMP2 and MMP9 are able to facilitate EMT, and previous reports have
shown that the two proteins are able to induce EMT in renal tubular
epithelial cells (27,28). However, it is unclear whether MMP2
and MMP9 induce EMT in alveolar epithelial cells. Recent evidence
has suggested that radiation upregulates MMP expression and
activity in various tissues (29–32). The
present study indicates that thoracic irradiation induces a marked
increase in MMP2 and MMP9 protein expression and gelatinase
activity. Furthermore, baseline MMP9 expression and gelatinase
activity were only moderately elevated at week 12; however, there
was a significant increase at week 24 after irradiation (Figs. 3 and 4). These results indicate that the
MMP9-mediated method is more complex compared that of with MMP2,
and suggest that MMP9 may play a greater role than MMP2 in RILI.
The reduction of SP-A expression induced by irradiation may have
caused the MMP9 level to be relatively low at week 12 after
irradiation, and TGFβ1 may also be involved in the inhibition or
reduction of MMP9 activity. This may explain why the increase of
MMP9 protein expression and gelatinase activity differs from that
of MMP2. A previous study indicated that TGFβ1 is able to
downregulate MMP9 gelatinase activity (32), and MMP9 also facilitates the release
of TGFβ1 (25). However, the present
study showed that MMP9 protein expression and activity peaked at
week 24 after irradiation and TGFβ1 levels continually increased
(Figs. 3 and 4). Further studies may be required to
determine the influence of TGFβ1 on MMP9, and the interaction
between the two proteins.
The ECM contains gelatin and collagen, and provides
a dynamic support structure on which epithelial cells may grow. In
addition, it may influence cellular behavior, including migration,
proliferation and morphology (33).
Furthermore, ECM is a key source of cytokines, and exerts a typical
pleiotropic effect. Although the primary inducer of EMT is TGFβ1,
the significance of the ECM is becoming increasingly apparent, with
previous studies suggesting that these components (e.g., type I
collagen) influence and augment the profibrotic effects of TGFβ1
(4,34). For example, studies in various cell
types have shown that type I collagen induces EMT (34,35). A
previous study indicates that basement membrane architecture is
critical in maintaining an epithelial phenotype and that
alterations in its composition may promote phenotypic changes
(35). Therefore, the interaction
between the epithelium and the matrix facilitates the TGFβ1-induced
production of EMT (4,34). Buckley et al (36) observed that type I collagen
significantly amplified the effect of TGFβ1-induced EMT in A549
cells treated with TGFβ1. Additionally, ECM components may have an
effect on the activity of fibroblasts (37). Myofibroblasts derived from EMT have
been shown to produce ECM components, such as type I collagen
(36), indicating that
myofibroblasts affect TGFβ1-induced EMT. TGFβ1 is an effective
stimulus that promotes ECM synthesis (19), upregulates MMP2 expression (38) and downregulates MMP9 expression.
MMP9, in combination with CD44, is able to stimulate TGFβ1
production (26), while MMP2 and
MMP9 are able to degrade the ECM. This forms a positive feedback
loop and continuously promotes the development of RILI, and may
result in pulmonary fibrosis. This process a key factor in the
consistent development of irradiation-induced lung fibrosis
following irradiation. Furthermore, TGFβ1 is hypothesized to cause
fibrosis, and multiple signaling pathways are involved in
irradiation-induced lung fibrosis. Thus, further studies are
required to clearly determine the mechanisms underlying
irradiation-induced lung fibrosis.
On the basis of the present results, we propose a
mechanism for RILI in irradiated Bama minipigs. This mechanism
involves a complicated interaction among SP-A, TGFβ1, EMT and ECM
which favors the development of RILI, and the participation of MMPs
in the development of RILI by interaction with EMT molecules, as
described above. Radiation causes a reduction in E-cadherin protein
expression levels, an increase in vimentin protein expression and
co-localization of α-SMA and SP-A in alveolar epithelia, suggesting
that EMT occurs in RILI, and that MMP2 and MMP9 serve crucial
functions in EMT by regulating TGFβ1. However, there were certain
limitations to the present experiments. For example, SP-A was
selected as a marker of AE2 and SP-A was observed to decrease
following irradiation, but increased AE2-to-mesenchymal transition
was observed at week 8 post-irradiation. However, in the present
study, a reduction in the expression level and non-specificity of
SP-A possibly led to reduced EMT of AE2 cells at week 12
post-irradiation. However, on the basis of the present data, and
potentially with the support of additional future studies to
identify the effect of tissue inhibitors of MMPs and types I and IV
collagen in RILI may lead to novel preventive interventions or
improved treatments for thoracic tumor patients undergoing
radiotherapy.
Acknowledgements
This study was supported by Guangxi Science and
Technology Key Projects Fund (no. 1140003A-24).
References
|
1
|
Stone HB, Coleman CN, Anscher MS and
McBride WH: Effects of radiation on normal tissue: Consequences and
mechanisms. Lancet Oncol. 4:529–536. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cappuccini F, Eldh T, Bruder D, Gereke M,
Jastrow H, SchulzeOsthoff K, Fischer U, Köhler D, Stuschke M and
Jendrossek V: New insights into the molecular pathology of
radiation-induced pneumopathy. Radiother Oncol. 101:86–92. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Trott KR, Herrmann T and Kasper M: Target
cells in raditation pneumopathy. Int J Radiat Oncol Biol Phys.
58:463–469. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim KK, Kugler MC, Wolters PJ, Robillard
L, Galvez MG, Brumwell AN, Sheppard D and Chapman HA: Alveolar
epithelial cell mesenchymal transition develops in vivo during
pulmonary fibrosis and is regulated by the extracellular matrix.
Proc Natl Acad Sci USA. 103:13180–13185. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Willis BC, Liebler JM, LubyPhelps K,
Nicholson AG, Crandall ED, du Bois RM and Borok Z: Induction of
epithelial-mesenchymal transition in alveolar epithelial cells by
transforming growth factor-β1: Potential role in idiopathic
pulmonary fibrosis. Am J Pathol. 166:1321–1332. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Almeida C, Nagarajan D, Tian J, Leal SW,
Wheeler K, Munley M, Blackstock W and Zhao W: The role of alveolar
epithelium in radiation-induced lung injury. PLoS One.
8:e536282013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Williams JP, Brown SL, Georges GE,
HauerJensen M, Hill RP, Huser AK, Kirsch DG, Macvittie TJ, Mason
KA, Medhora MM, et al: Animal models for medical countermeasures to
radiation exposure. Radiat Res. 173:557–578. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hopewell JW, Rezvani M and Moustafa HF:
The pig as a model for the study of radiation effects on the lung.
Int J Radiat Biol. 76:447–452. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li M, Abdollahi A, Gröne HJ, Lipson KE,
Belka C and Huber PE: Late treatment with imatinib mesylate
ameliorates radiation-induced lung fibrosis in a mouse model.
Radiat Oncol. 4:662009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Molthen RC, Wu Q, Fish BL, Moulder JE,
Jacobs ER and Medhora MM: Mitigation of radiation induced pulmonary
vascular injury by delayed treatment with captopril. Respirology.
17:1261–1268. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Travis EL: The sequence of histological
changes in mouse lungs after single doses of x-rays. Int J Radiat
Oncol Biol Phys. 6:345–347. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Uhal BD: Cell cycle kinetics in the
alveolar epithelium. Am J Physiol. 272:L1030–L1045. 1997.
|
|
14
|
Alcorn JF and Wright JR: Surfactant
protein A inhibits alveolar macrophage cytokine production by
CD14-independent pathway. Am J Physiol Lung Cell Mol Physiol.
286:L129–L136. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vazquez de Lara LG, Umstead TM, Davis SE
and Phelps DS: Surfactant protein A increases matrix
metalloproteinase-9 production by THP-1 cells. Am J Physiol Lung
Cell Mol Physiol. 285:L899–L906. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hallman M, Maasilta P, Kivisaari L and
Mattson K: Changes in surfactant in bronchoalveolar lavage fluid
after hemithorax irradiation in patients with mesothelioma. Am Rev
Respir Dis. 141:998–1005. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sasaki R, Soejima T, Matsumoto A, Maruta
T, Yamada K, Ota Y, Kawabe T, Nishimura H, Sakai E, Ejima Y, et al:
Clinical significance of serum pulmonary surfactant proteins A and
D for the early detection of radiation pneumonitis. Int J Radiat
Oncol Biol Phys. 50:301–307. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wynn TA: Cellular and molecular mechanisms
of fibrosis. J Pathol. 214:199–210. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wynn TA: Integrating mechanisms of
pulmonary fibrosis. J Exp Med. 208:1339–1350. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nagarajan D, Melo T, Deng Z, Almeida C and
Zhao W: ERK/GSK3β/Snail signaling mediates radiation-induced
alveolar epithelial-to-mesenchymal transition. Free Radic Biol Med.
52:983–992. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Verrecchia F and Mauviel A: Transforming
growth factor-beta and fibrosis. World J Gastroenterol.
13:3056–3062. 2007.PubMed/NCBI
|
|
22
|
Thannickal VJ, Toews GB, White ES, Lynch
JP III and Martinez FJ: Mechanisms of pulmonary fibrosis. Annu Rev
Med. 55:395–417. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao L, Sheldon K, Chen M, Yin MS, Hayman
JA, Kalemkerian GP, Arenberg D, Lyons SE, Curtis JL, Davis M, et
al: The predictive role of plasma TGF-β1 during radiation therapy
for radiation-induced lung toxicity deserves further study in
patients with non-small cell lung cancer. Lung Cancer. 59:232–239.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Visse R and Nagase H: Matrix
metalloproteinases and tissue inhibitors of metalloproteinases:
Structure, function, and biochemistry. Circ Res. 92:827–839. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Atkinson JJ and Senior RM: Matrix
metalloproteinase-9 in lung remodeling. Am J Respir Cell Mol Biol.
28:12–24. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
López-Novoa JM and Nieto MA: Inflammation
and EMT: An alliance towards organ fibrosis and cancer progression.
EMBO Mol Med. 1:303–314. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tan TK, Zheng G, Hsu TT, Wang Y, Lee VW,
Tian X, Wang Y, Cao Q, Wang Y and Harris DC: Macrophage matrix
metalloproteinase-9 mediates epithelial-mesenchymal transition in
vitro in murine renal tubular cells. Am J Pathol. 176:1256–1270.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cheng S and Lovett DH: Gelatinase A
(MMP-2) is necessary and sufficient for renal tubular cell
epithelial-mesenchymal transformation. Am J Pathol. 162:1937–1949.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee WH, Warrington JP, Sonntag WE and Lee
YW: Irradiation alters MMP-2/TIMP-2 system and collagen type IV
degradation in brain. Int J Radiat Oncol Biol Phys. 82:1559–1566.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
StrupPerrot C, Vozenin-Brotons M-C,
Vandamme M, Linard C and Mathé D: Expression of matrix
metalloproteinases and tissue inhibitor metalloproteinases
increases in X-irradiated rat ileum despite the disappearance of
CD8a T cells. World J Gastroenterol. 11:6312–6321. 2005.PubMed/NCBI
|
|
31
|
Yang K, Palm J, König J, Seeland U,
Rosenkranz S, Feiden W, Rübe C and Rübe CE:
Matrix-Metallo-Proteinases and their tissue inhibitors in
radiation-induced lung injury. Int J Radiat Biol. 83:665–676. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Susskind H, Hymowitz MH, Lau YH, Atkins
HL, Hurewitz AN, Valentine ES, Meek AG and Zucker S: Increased
plasma levels of matrix metalloproteinase-9 and tissue inhibitor of
metalloproteinase-1 in lung and breast cancer are altered during
chest radiotherapy. Int J Radiat Oncol Biol Phys. 56:1161–1169.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Daley WP, Peters SB and Larsen M:
Extracellular matrix dynamics in development and regenerative
medicine. J Cell Sci. 121:255–264. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shintani Y, Maeda M, Chaika N, Johnson KR
and Wheelock MJ: Collagen I promotes epithelial-to-mesenchymal
transition in lung cancer cells via transforming growth factor-β
signaling. Am J Respir Cell Mol Biol. 38:95–104. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zeisberg M, Bonner G, Maeshima Y, et al:
Renal fibrosis: Collagen composition and assembly regulates
epithelial-mesenchymal transdifferentiation. Am J Pathol.
159:1313–1321. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Buckley ST, Medina C, Davies AM and
Ehrhardt C: Cytoskeletal re-arrangement in TGF-β1-induced alveolar
epithelial-mesenchymal transition studied by atomic force
microscopy and high-content analysis. Nanomedicine (Lond).
8:355–364. 2012.
|
|
37
|
Kolb M, Margetts PJ, Sime PJ and Gauldie
J: Proteoglycans decorin and biglycan differentially modulate
TGF-beta-mediated fibrotic responses in the lung. Am J Physiol Lung
Cell Mol Physiol. 280:L1327–L1334. 2001.PubMed/NCBI
|
|
38
|
Kasai H, Allen JT, Mason RM, Kamimura T
and Zhang Z: TGF-β1 induces human alveolar epithelial to
mesenchymal cell transition (EMT). Respir Res. 6:562005. View Article : Google Scholar : PubMed/NCBI
|