Introduction
Atherosclerosis (AS) has come to be recognized as an
active and inflammatory process, rather than simply a passive
process of lipid infiltration or a reparative process following
endothelial injury (1–3). One of the major inflammatory cytokines
is tumor necrosis factor-α (TNF-α), a pro-inflammatory cytokine
that is released in response to a pathological condition (4). TNF-α can injure the structure of
endothelial cells and induce inflammatory responses by enhancing
the expression and secretion of adhesion molecules, including
vascular cell adhesion molecule-1 (VCAM-1), intercellular adhesion
molecule-1 (ICAM-1), endothelial cell selectin and fibronectin (FN)
(5,6), which results in leukocyte recruitment
to the endothelium and the initiation of AS. A number of chemicals
derived from plants with anti-inflammatory properties have been
reported to exert an anti-leukocyte recruitment effect (7,8). FN is a
250-kDa adhesive glycoprotein and one of the most abundant proteins
in the extracellular matrix (ECM). FN generates a scaffold that
allows the attachment of other ECM components, and large quantities
of FN have been detected in atherosclerotic plaques, suggesting
that it may play a role in the pathogenesis of AS (9,10).
Physiologically, FN plays an important role in a number of
processes, including cell adhesion, motility and tissue repair;
however, its overproduction may decrease the motility and
replication of various cell types, including endothelial cells
(11).
Oxidative stress and the production of intracellular
reactive oxygen species (ROS) have been implicated in the
pathogenesis of AS (12). ROS and
their by-products may not only be cytotoxic to cells but also play
a role in signal transduction processes, such as cell growth and
the post-translational modification of proteins, which contributes
to the formation of AS (13). The
common key point in the pathophysiology of AS is believed to be the
intracellular redox signal-induced expression of specific
inflammatory genes (14). Salvia
miltiorrhiza (S.M.), a herb that is often used in Traditional
Chinese Medicine, has been found to exert beneficial effects on the
circulatory system (15). Aqueous
extracts of S.M. that are rich in antioxidants have been described
as being effective in reducing AS in experimental studies in
vitro and in vivo (16,17). Our
previous studies have demonstrated that the main compounds of S.M.
inhibit endothelin-1 expression, stimulate nitric oxide production
(18) and attenuate plasminogen
activator inhibitor type 1 production in TNF-α-treated human
umbilical vein endothelial cells (HUVECs) (19). Furthermore, we reported that
protocatechuic aldehyde (PA, also known as
3,4-dihydroxybenzaldehyde), a compound isolated from the aqueous
extract of S.M., selectively inhibits TNF-α-induced VCAM-1 and
ICAM-1 expression and reduces monocyte adhesion to endothelial
cells (20); however, no studies
examining the effect of PA on the regulation of FN in endothelial
cells exist, to the best of our knowledge.
The aim of the present study was four-fold: i) To
examine the effect of TNF-α on FN secretion and expression in
cultured HUVECs; ii) to investigate the effect of PA on
TNF-α-induced FN expression in HUVECs; iii) to explore the effect
of PA on the TNF-α-induced activation of extracellular
signal-regulated kinase 1 and 2 (ERK1/2), c-Jun N-terminal kinase
(JNK) and p38 mitogen-activated protein kinase (p38) in HUVECs; and
iv) to investigate the activity of the key AS-related transcription
factor nuclear factor-κB (NF-κB).
Materials and methods
Reagents
PA was purchased from the Chinese National Institute
for the Control of Pharmaceutical and Biological Products (Beijing,
China). PA was dissolved in warm culture medium just before
incubation with HUVECs. Direct exposure of the PA to light and air
was avoided during the experiments. Recombinant human TNF-α, the
CellTiter Aqueous One Solution Cell Proliferation Assay (MTS) and
the Gel Shift Assay Core system were purchased from Promega Corp.
(Madison, WI, USA). Antibodies to FN (rabbit polyclonal; 1:500;
cat. no. sc-9068), phosphorylated (P-)ERK1/2 (rabbit polyclonal;
1:200; cat. no. sc-101761), ERK1/2 (rabbit polyclonal; 1:250; cat.
no. sc-292838), p38 (rabbit polyclonal; 1:400; cat. no. sc-535) and
JNK (rabbit polyclonal; 1:400; cat. no. sc-571) were purchased from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA), and the
antibodies to P-JNK (rabbit monoclonal; 1:500; cat. no. 4668) and
P-p38 (rabbit polyclonal; 1:500; cat. no. 9211) and the anti-rabbit
IgG horseradish peroxidase (HRP)-conjugated secondary antibody
(1:1,200; cat. no. 7074P2) were obtained from Cell Signaling
Technology, Inc. (Beverly, MA, USA). The FN ELISA kit was obtained
from Shanghai Sun Biotech Co., Ltd. (Shanghai, China).
Cell culture
HUVECs were obtained from Cascade Biologics, Inc.
(Portland, OR, USA), in the form of cryopreserved primary cultures,
and grown in culture flasks (Costar® Corning, New York, NY, USA) in
M200 endothelial cell growth medium (Cascade Biologics, Inc.)
supplemented with 2% low serum growth supplement (LSGS; Cascade
Biologics, Inc.) according to the manufacturer's instructions. The
growth medium was changed every other day until the cells reached
confluence. Passage 3 and 4 cells were grown in monolayers at 37°C
in a humidified atmosphere of 5% CO2 and 95% air and used for
experiments at 80% confluency. At 24 h prior to the experiments,
the control medium was removed and replaced with LSGS-free medium
containing 0.4% fetal bovine serum. For the experiments, HUVECs
were cultured in medium containing 0.4% fetal bovine serum with or
without the PA for 18 h, prior to further culture for 6 h with
TNF-α (2 ng/ml).
Assessment of cell viability and
cellular ROS generation
In order to evaluate cytotoxicity, cells were seeded
at a density of 5,000 cells/well into 96-well culture plates
(Costar) and grown for 48 h. The cells were incubated with PCA
(6.75, 13.5, 27, 54 or 108 nM) with or withour TNF-α (2 ng/ml) in
M200 media (serum-free) containing 0.4% fetal bovine serum for 24
h, and 20 µl MTS was then added to each well for further incubation
at 37°C for 2 h. The absorbance of the solubilized formazan was
read at 490 nm using a Victor 1420 Multilabel Counter instrument
(Wallac, Turku, Finland). Cells incubated in control media were
taken to be 100% viable. The generation of ROS was assessed using
the ROS-sensitive fluorescence indicator 2′,7′-dichlorofluorescein
diacetate (DCFH-DA). A total of 10 µmol/l DCFH-DA was added to the
cell culture wells in a potassium phosphate buffer for 30 min. The
fluorescence of 2′,7′-dichlorofluorescein, the oxidation product of
DCFH-DA, was measured at an excitation wavelength of 485 nm and an
emission wavelength of 530 nm on a fluorescence spectrophotometer
(Victor 1420 Multilabel Counter; Wallac).
Cell surface immunoassay and FN
secretion
To detect the expression of adhesion molecules,
ELISA was performed as previously described (19). In brief, the HUVECs were plated onto
96-well plates overnight and growth-arrested for 24 h with 0.4%
serum-containing medium. The cells were then stimulated with TNF-α
with or without PA, washed with phosphate-buffered saline (PBS) and
fixed. Anti-FN antibody (Shanghai Sun Biotech Co., Ltd.) was added
to the wells for 1 h at 37°C. Cells were washed and the expression
of FN was quantified by the addition of o-phenylenediamine
dihydrochloride in phosphate-citrate buffer. Following incubation
for 20 min at 37°C, the reaction was terminated through the
addition of 5 N H2SO4, and the absorbance of
each well was measured at 490 nm using a Multilabel reader. In
order to measure the FN secretion by the HUVECs, an ELISA kit was
used to determine the levels of soluble FN antigens in the culture
supernatant of the HUVECs, according to the manufacturer's
instructions (Shanghai Sun Biotech Co., Ltd.).
Preparation of nuclear extracts
Upon reaching 80% confluency, the HUVECs were
administered the indicated treatment and nuclear protein extracts
were prepared, as described previously (19). All nuclear extraction procedures were
performed on ice with ice-cold reagents. The cells were harvested,
washed with PBS, resuspended in Buffer A, which contained 10 mM
HEPES (pH 7.6), 10 mM KCl, 0.1 mM EDTA, 1 mM dithiothreitol (DTT)
and 0.5 mM phenylmethylsulfonyl fluoride (PMSF), and incubated for
10 min at 4°C. Following centrifugation at 300 × g for 10 min, the
pellets containing the nuclei were suspended in Buffer B, which
contained 20 mM HEPES (pH 7.6), 10 mM KCl, 1 mM EDTA, 1 mM DTT, 0.5
mM PMSF, 25% glycerol and 0.4 M NaCl, for 30 min. Nuclear proteins
were isolated by centrifugation at 12,000 × g for 20 min. The
protein concentrations were determined using a Bicinchoninic Acid
Protein Assay kit (Pierce Biotechnology, Rockford, IL, USA, and the
proteins were stored at −80°C until use in the NF-κB binding
activity assay.
NF-κB binding activity assay
The NF-κB activity in the nuclear protein (20 µg) of
treated or control HUVECs was measured using a DNA-binding ELISA
kit (TransAM™ NF-κB p65 assay; Active Motif, Carlsbad, CA, USA)
according to the manufacturer's instructions and analyzed using a
microplate absorbance reader.
Western blotting and activity assay of
mitogen-activated protein kinases (MAPKs) (ERK1/2, JNK and
p38)
Following treatment with the reagents, the cells
were washed with PBS and harvested in 200 µl lysis buffer
containing 20 mM HEPES (pH 7.9), 10 mM NaCl, 1 mM EDTA, 1 mM DTT,
0.1% Nonidet P40 (V/V) and protease inhibitor (0.1 µg/ml leupeptin,
5 µg/ml aprotinin and 0.5 mM PMSF). Cell lysates were subjected to
sodium dodecyl sulfate polyacrylamide gel electrophoresis, and
proteins were transferred to a polyvinylidene difluoride membrane
(Schleicher and Schuell Biosciences, Inc., Keene, NH, USA). The
membrane was blocked for 1 h at room temperature with Tris buffered
saline-Tween 20 (0.5%)/5% non-fat skim milk. The blots were then
incubated overnight with antibodies against FN, or the
phosphorylated or unphosphorylated forms of ERK1/2, JNK or p38 at
4°C, followed by incubation for 1 h with HRP-conjugated secondary
antibody. Immunoreactive bands were visualized using Western
Blotting Luminol Reagent (Santa Cruz Biotechnology, Inc.), and the
blots were exposed to XBT-1 film (Kodak, Xiamen, China).
Statistical analysis
All values are expressed as the mean ± standard
error of the mean of independent determinations. Statistical
analysis was performed with analysis of variance and the Tukey test
using GraphPad Prism® software (GraphPad Software, Inc., La Jolla,
CA, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Effects of PA on FN secretion and cell
surface expression
To examine the effect of TNF-α on FN secretion and
surface expression in HUVECs, the HUVECs were exposed to 2 ng/ml
TNF-α for 6, 12 and 18 h. The expression of FN was determined in
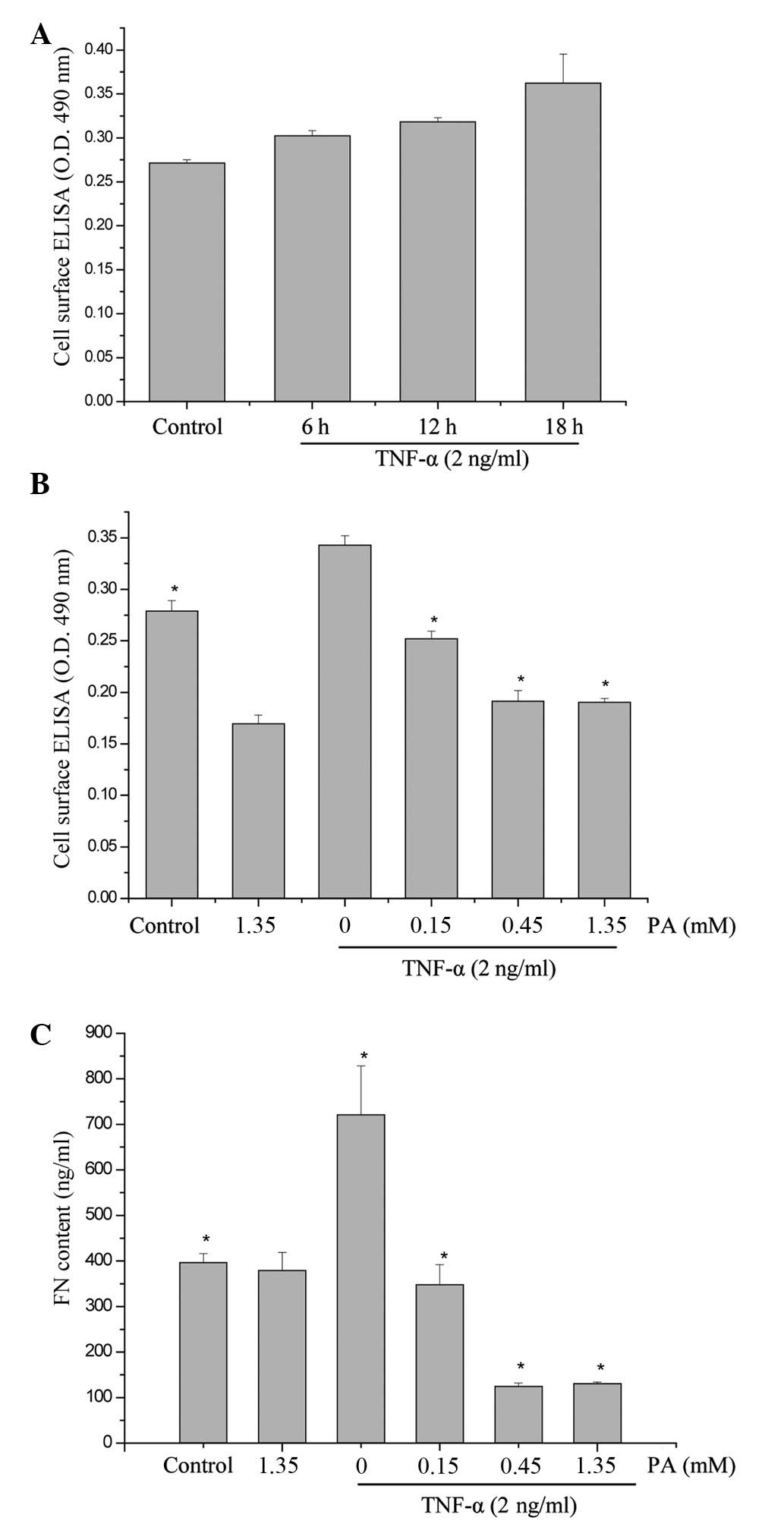
accordance with the aforementioned method. As shown in Fig. 1, the surface expression of FN
increased in a time-dependent manner (maximum at 18 h). To
investigate whether PA affected the TNF-α-induced FN secretion and
expression, the effect of various concentrations of PA on
TNF-α-induced FN secretion and expression was examined using ELISA
and cell surface ELISA. PA at doses of 0.15, 0.45 and 1.35 mM
induced a significant dose-dependent inhibition of FN protein
surface expression. FN protein surface expression was reduced to
73.4, 55.8 and 55.5% of the control, respectively, and FN secretion
was significantly reduced to 48.2, 17.4 and 18.2%, respectively
(Fig. 1). The FN expression in the
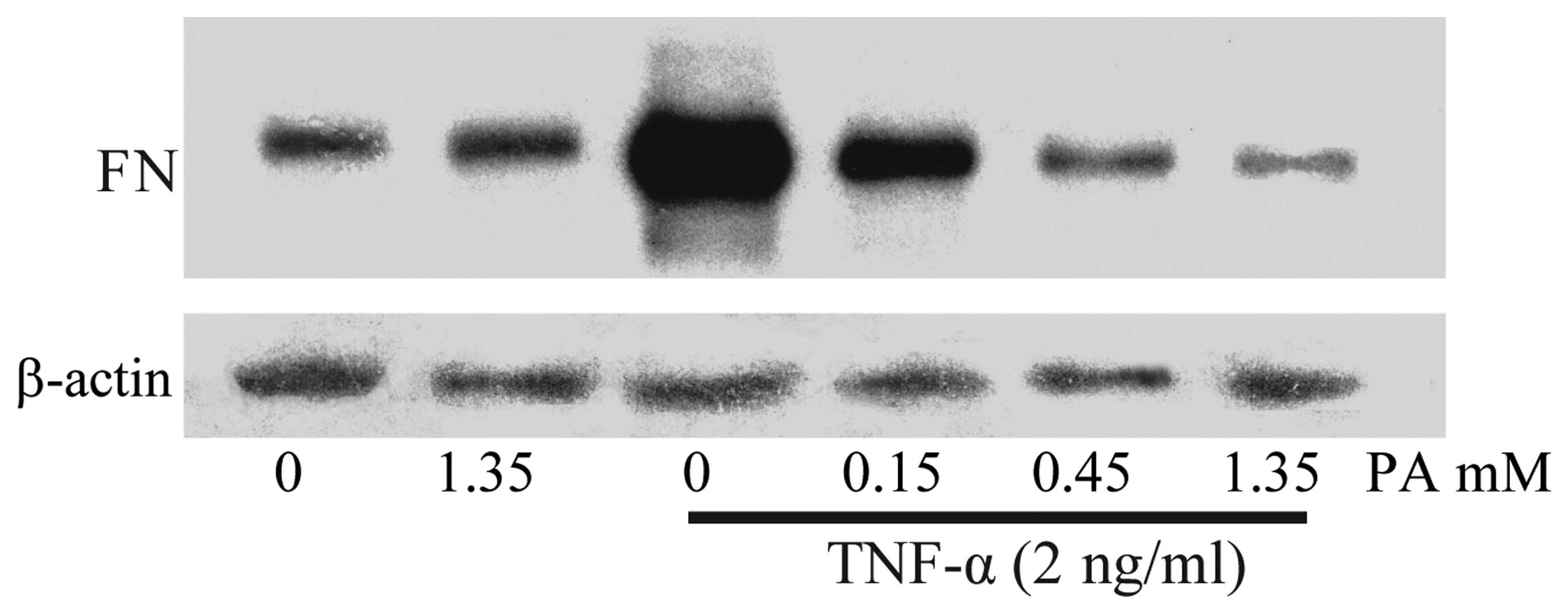
cytoplasm was also examined using western blot analysis. As shown
in Fig. 2, the FN expression in the
TNF-α-stimulated HUVECs increased notably, whereas the pretreatment
of HUVECs with PA for 18 h markedly attenuated the TNF-α-stimulated
FN expression in a dose-dependent manner.
PA attenuates TNF-α-induced ROS
production in HUVECs
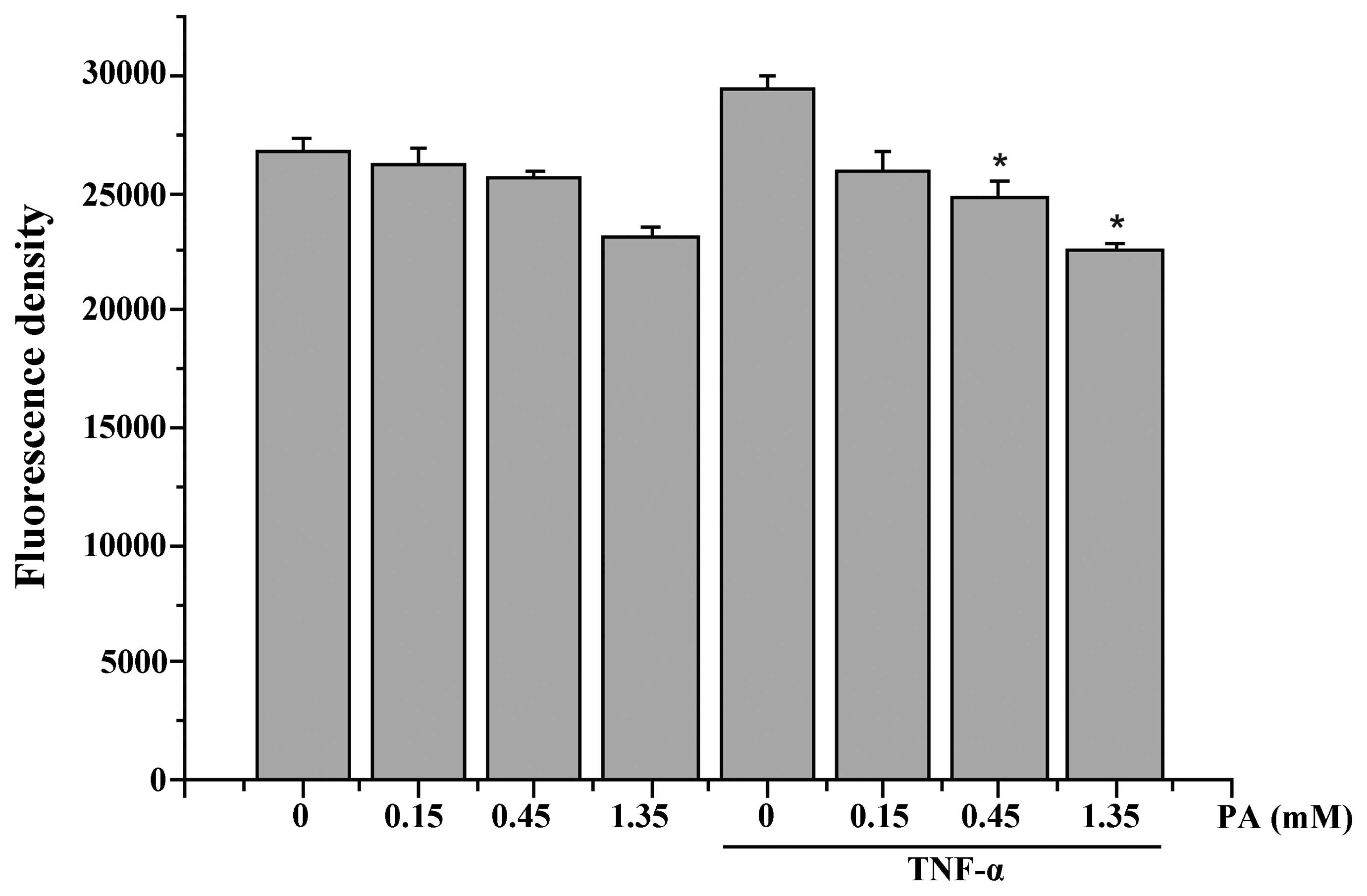
As shown in Fig. 3,
incubating the HUVECs for 15 min with 2 ng/ml TNF-α significantly
increased the intracellular ROS generation. PA was found to inhibit
the TNF-α-induced increase in ROS generation in a dose-dependent
manner (Fig. 3).
Effect of PA on TNF-α-induced ERK1/2,
JNK and p38 activation in HUVECs
To examine whether PA affects TNF-α-induced MAPK
activation, the effect of various concentrations of PA on
TNF-α-induced ERK1/2, JNK and p38 activation in HUVECs was
assessed. The cells were pretreated with PA for 60 min prior to the
addition of TNF-α (2 ng/ml) for 15 min for ERK1/2, JNK and p38
activation. TNF-α-induced JNK activation was inhibited by PA in a
concentration-dependent manner (0.15, 0.45, and 1.35 mM). By
contrast, ERK1/2 and p38 activation were not affected by PA
(Fig. 4). Western blot analysis with
anti-ERK1/2, -JNK and -p38 antibodies revealed no differences in
the total quantities of ERK1/2, JNK and p38. These findings suggest
that TNF-α-induced JNK activation, but not ERK1/2 and p38
activation, is specifically sensitive to PA in HUVECs.
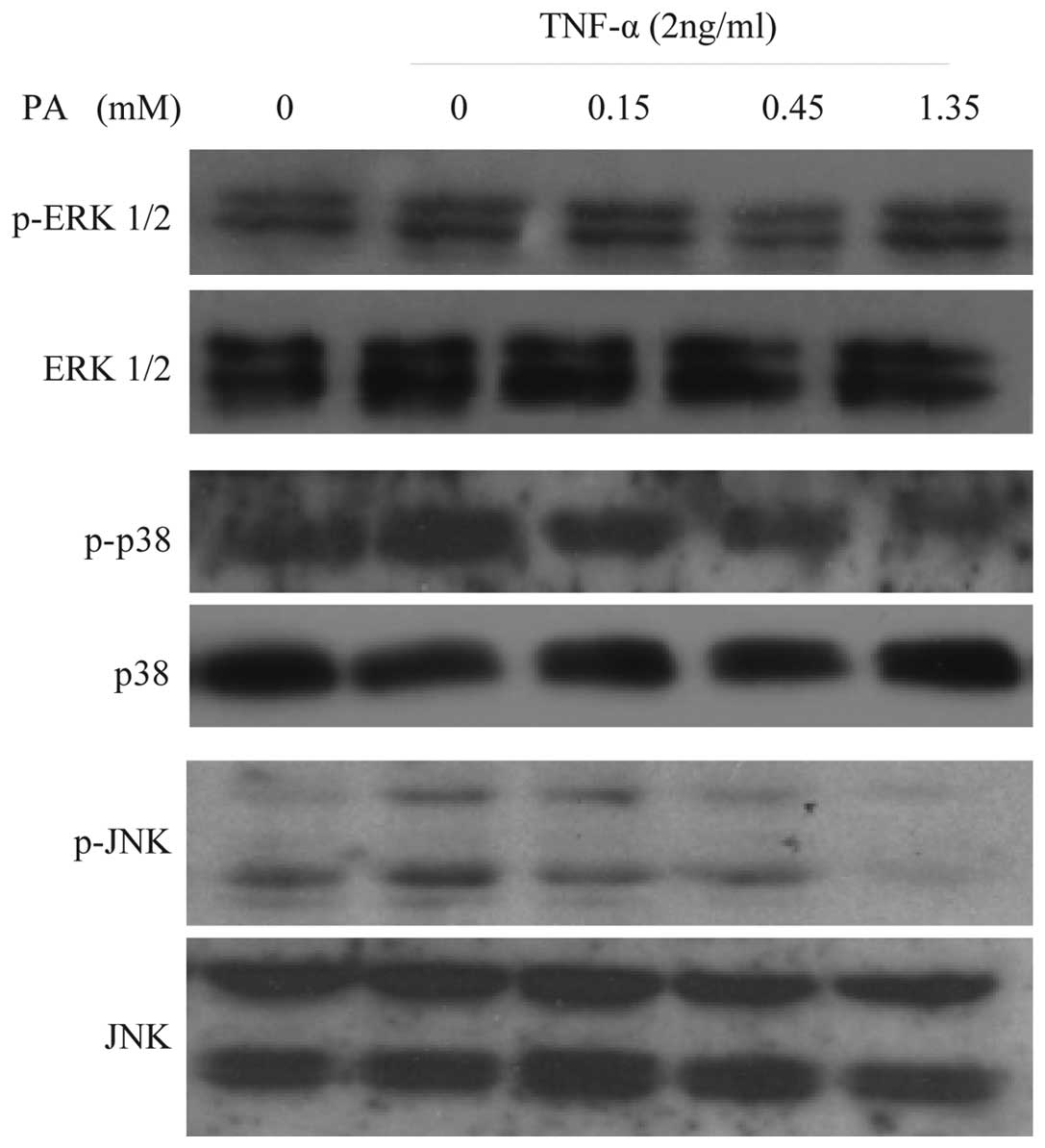 | Figure 4.PA inhibits the TNF-α-induced JNK
activation in a concentration-dependent manner in human umbilical
vein endothelial cells, but not ERK1/2 and p38 activation. Cells
were pretreated with PA at the indicated concentrations for 60 min.
The cells were then stimulated with 2 ng/ml TNF-α for 15 min for
ERK1/2, JNK and p38 activation. Cells were harvested, lysed and
used for subsequent analysis. No significant differences in the
levels of ERK1/2, JNK and p38 were observed using immunoblot
analysis with anti-ERK1/2, -JNK and -p38 antibodies. PA,
protocatechuic aldehyde; FN, fibronectin; TNF-α, tumor necrosis
factor-α; p-ERK1/2, phosphorylated extracellular signal-regulated
kinase 1 and 2; JNK, c-Jun N-terminal kinase; p38, p38
mitogen-activated protein kinase. |
PA inhibits the TNF-α-induced
transcriptional activation of NF-κB in HUVECs
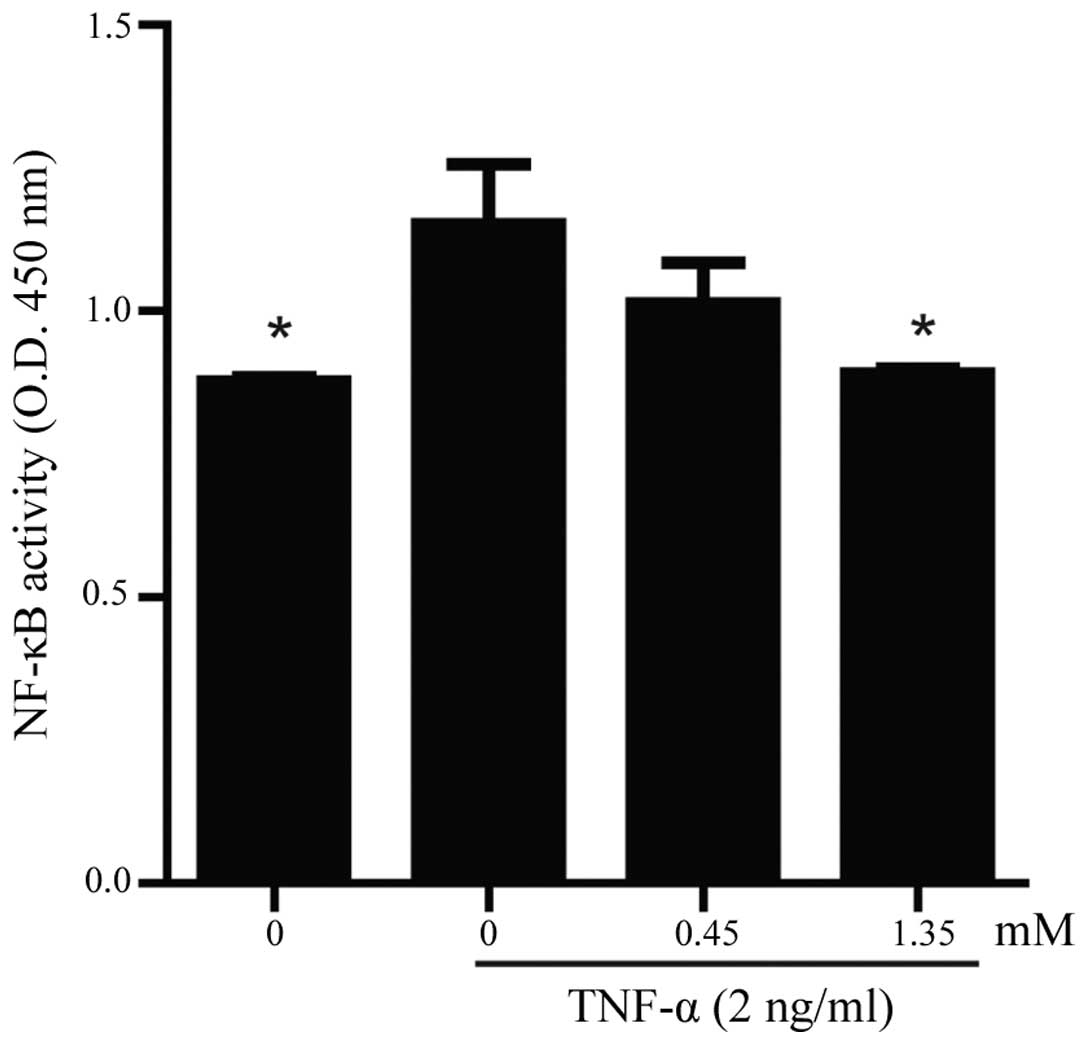
To examine the effect of PA on constitutive NF-κB
activation, an NF-κB p65 activity assay was performed using a
DNA-binding ELISA kit. As shown in Fig.
5, increased DNA binding activity for NF-κB was observed in the
TNF-α-stimulated HUVECs, whereas the pretreatment of the HUVECs
with PA for 18 h markedly attenuated the TNF-α-stimulated NF-κB
activation in a dose-dependent manner.
Discussion
S.M. has been demonstrated to be clinically
effective for the prevention and treatment of AS (21,22).
Previous studies have demonstrated that S.M. is an antioxidative,
antithrombogenic and anti-inflammatory plant (23). AS is characterized by endothelial
cell injury and dysfunction. One of the earliest events in
atherogenesis is the adhesion of monocytes to the endothelium,
followed by their infiltration and differentiation into macrophages
(2). This key step is mediated by
the interaction of monocytes with molecules expressed on the
surface of endothelial cells (24).
FN is a large, multifunctional glycoprotein that is important in
ECM organization, tissue remodeling and wound healing (25). The overproduction of FN can decrease
the motility and replication of numerous types of cells, including
endothelial cells. Inflammation or injury can trigger the
deposition of transitional ECM proteins, such as FN and fibrinogen
(FG), into the subendothelial matrix (26). In vivo, FN and FG are
deposited at AS-prone sites prior to other signs of AS (27). Cellular FN (cFN) normally makes up
<2% of the total FN present in the plasma and is synthesized
locally by endothelial cells, smooth muscle cells or fibroblasts in
response to cytokine stimulation or vascular injury (28). cFN and soluble VCAM-1 have been found
to be reliable markers of endothelial injury (29). In the present study, the effect of PA
on the expression and secretion of endothelial FN was investigated.
It was found that the surface expression, secretion and content of
FN in the cytoplasm increased significantly following the treatment
of HUVECs with TNF-α. PA treatment attenuated basal and
TNF-α-induced FN surface expression and secretion in a
dose-dependent manner.
In previous study, we reported that the aqueous
compound of S.M., PA, selectively inhibited cytokine-induced VCAM-1
and ICAM-1 expression and reduced monocyte adhesion to endothelial
cells through an antioxidative mechanism (20). In the present study, PA was found to
dose-dependently attenuate the ROS production in HUVECs with or
without TNF-α stimulation. ROS overproduction leads to oxidative
modifications of DNA, lipid oxidation, protein modification and the
activation of redox-sensitive genes (13,30,31),
including the upregulation of FN expression (32), as well as that of VCAM-1 and ICAM-1.
The earliest stages of AS are associated with the increased
attraction and adhesion of monocytes to the endothelium, which is
mediated by the adhesive molecules expressed by the activated
endothelium. FN plays an important role in the initiation and
progression of AS.
In response to TNF-α treatment, the transcription
factor NF-κB and MAPKs, including ERK, p38 and JNK, are activated
in most types of cells (33).
Activation of NF-κB and MAPKs plays an important role in the
induction of numerous cytokines and immune-regulatory proteins and
is pivotal for several inflammatory responses (34). In the present study, the mechanisms
underlying the action of PA were investigated by examining the
effects of PA on the MAPK and NF-κB pathways. It was found that PA
specifically inhibited TNF-α-induced JNK activation, but not ERK1/2
and p38 activation, in HUVECs. The effect of PA on JNK activation
by TNF-α would be expected to have important consequences for the
expression of adhesion molecules by endothelial cells (35). The most likely mechanism for the
PA-induced inhibition of TNF-α would be crosstalk between the MAPK
and NF-κB signaling pathways. At another level, each of the three
major MAPK pathways (p38, ERK and JNK) has been shown to
phosphorylate p65 or p50 in various cell types, affecting
transcriptional activity (36). To
directly assess whether NF-κB activity was attenuated in
endothelial cells treated with PA, the DNA binding activity of
NF-κB was analyzed using a commercial DNA-binding ELISA assay, and
it was found that the TNF-α-induced NF-κB activity was
significantly inhibited by PA.
In conclusion, the present study has demonstrated
that PA, the aqueous ingredient of S.M., significantly inhibits
TNF-α-induced JNK activation, but not ERK1/2 and p38 activation, in
HUVECs. Experimental data also showed that PA inhibits NF-κB
transcriptional activation and the resultant expression and
secretion of the adhesion molecule FN. These results may provide
useful insight to enhance the understanding of the pharmacological
action of S.M.
Acknowledgements
This study was supported by a grant from the China
Postdoctoral Science Foundation (no. 2004036218).
References
|
1
|
Langheinrich AC and Bohle RM:
Atherosclerosis: Humoral and cellular factors of inflammation.
Virchows Arch. 446:101–111. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ross R: Atherosclerosis - an inflammatory
disease. N Engl J Med. 340:115–126. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Montecucco F and Mach F: Atherosclerosis
is an inflammatory disease. Semin Immunopathol. 31:1–3. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ding M, Ye TX, Zhao GR, Yuan YJ and Guo
ZX: Aqueous extract of Salvia miltiorrhiza attenuates increased
endothelial permeability induced by tumor necrosis factor-alpha.
Int Immunopharmacol. 5:1641–1651. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pellegatta F, Radaelli A, Ferrero E,
Toninelli E, Vidal MJ, Chierchia SL and Zocchi MR: Inducible nitric
oxide synthase modulates fibronectin production in the EA.hy926
cell line and cultured human umbilical vein endothelial cells. J
Cardiovasc Pharmacol. 24:1014–1019. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Modur V, Zimmerman GA, Prescott SM and
McIntyre TM: Endothelial cell inflammatory responses to tumor
necrosis factor alpha. Ceramide-dependent and -independent
mitogen-activated protein kinase cascades. J Biol Chem.
271:13094–13102. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yoon JJ, Lee YJ, Park OJ, Lee SM, Lee YP,
Cho NG, Kang DG and Lee HS: Doinseunggitang ameliorates endothelial
dysfunction in diabetic atherosclerosis. Evid Based Complement
Alternat Med. 2013:7835762013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yin Y, Wan J, Li P, Jia Y, Sun R, Pan G
and Wan G: Protective effect of Xin Mai Jia ultrafiltration extract
on human umbilical vein endothelial cell injury induced by hydrogen
peroxide and the effect on the NO-cGMP signaling pathway. Exp Ther
Med. 8:38–48. 2014.PubMed/NCBI
|
|
9
|
Rohwedder I, Montanez E, Beckman K,
Bengtsson E, Dunér P, Nilsson J, Soehnlein O and Fässler R: Plasma
fibronectin deficiency impedes atherosclerosis progression and
fibrous cap formation. EMBO Mol Med. 4:564–576. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Erickson HP: Strethching fibronectin. J
Muscle Res Cell Motil. 23:575–580. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Madri JA, Pratt BM and Yannariello-Brown
J: Matrix-driven cell size change modulates aortic endothelial cell
proliferation and sheet migration. Am J Pathol. 132:18–27.
1988.PubMed/NCBI
|
|
12
|
Alexander RW: Theodore cooper memorial
lecture. Hypertension and the pathogenesis of atherosclerosis.
Oxidative stress and the mediation of arterial inflammatory
response: A new perspective. Hypertension. 25:155–161. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Patel RP, Moellering D, Murphy-Ullrich J,
Jo H, Beckman JS and Darley-Usmar VM: Cell signaling by reactive
nitrogen and oxygen species in atherosclerosis. Free Radic Biol
Med. 28:1780–1794. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Griendling KK, Sorescu D and Ushio-Fukai
M: NAD(P)H oxidase: Role in cardiovascular biology and disease.
Circ Res. 86:494–501. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zheng CS, Xu XJ, Ye HZ, Wu GW, Xu HF, Li
XH, Huang SP and Liu XX: Computational pharmacological comparison
of Salvia miltiorrhiza and Panax notoginseng used in the
therapy of cardiovascular diseases. Exp Ther Med. 6:1163–1168.
2013.PubMed/NCBI
|
|
16
|
Chen YH, Lin SJ, Ku HH, Shiao MS, Lin FY,
Chen JW and Chen YL: Salvianolic acid B attenuates VCAM-1 and
ICAM-1 expression in TNF-alpha-treated human aortic endothelial
cells. J Cell Biochem. 82:512–521. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen YL, Yang SP, Shiao MS, Chen JW and
Lin SJ: Salvia miltiorrhiza inhibits intimal hyperplasia and
monocyte chemotactic protein-1 expression after balloon injury in
cholesterol-fed rabbits. J Cell Biochem. 83:484–493. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou Z, Wang SQ, Liu Y and Miao AD:
Cryptotanshinone inhibits endothelin-1 expression and stimulates
nitric oxide production in human vascular endothelial cells.
Biochim Biophys Acta. 1760:1–9. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou Z, Liu Y, Miao AD and Wang SQ:
Salvianolic acid B attenuates plasminogen activator inhibitor type
1 production in TNF-alpha treated human umbilical vein endothelial
cells. J Cell Biochem. 96:109–116. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou Z, Liu Y, Miao AD and Wang SQ:
Protocatechuic aldehyde suppresses TNF-alpha-induced ICAM-1 and
VCAM-1 expression in human umbilical vein endothelial cells. Eur J
Pharmacol. 513:1–8. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zeng Y, Song JX and Shen XC: Herbal
remedies supply a novel prospect for the treatment of
atherosclerosis: A review of current mechanism studies. Phytother
Res. 26:159–167. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin TH and Hsieh CL: Pharmacological
effects of Salvia miltiorrhiza (Danshen) on cerebral
infarction. Chin Med. 5:222010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu WY and Wang YP: Pharmacological actions
and therapeutic applications of Salvia miltiorrhiza depside
salt and its active components. Acta Pharmacol Sin. 33:1119–1130.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Faggiotto A and Ross R: Studies of
hypercholesterolemia in the nonhuman primate. II. Fatty streak
conversion to fibrous plaque. Arteriosclerosis. 4:341–356. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Potts JR and Campbell ID: Structure and
function of fibronectin modules. Matrix Biol. 15:313–321. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sechler JL, Corbett SA, Wenk MB and
Schwarzbauer JE: Modulation of cell-extracellular matrix
interactions. Ann NY Acad Sci. 857:143–154. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Orr AW, Sanders JM, Bevard M, Coleman E,
Sarembock IJ and Schwartz MA: The subendothelial extracellular
matrix modulates NF-kappaB activation by flow: A potential role in
atherosclerosis. J Cell Biol. 169:191–202. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kosmehl H, Berndt A and Katenkamp D:
Molecular variants of fibronectin and laminin: Structure,
physiological occurrence and histopathological aspects. Virchows
Arch. 429:311–322. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Powers RW, Majors AK, Cerula SL, Huber HA,
Schmidt BP and Roberts JM: Changes in markers of vascular injury in
response to transient hyperhomocysteinemia. Metabolism. 52:501–507.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mann GE, Bonacasa B, Ishii T and Siow RC:
Targeting the redox sensitive Nrf2-Keap1 defense pathway in
cardiovascular disease: Protection afforded by dietary isoflavones.
Curr Opin Pharmacol. 9:139–145. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Genestra M: Oxyl radicals, redox-sensitive
signalling cascades and antioxidants. Cell Signal. 19:1807–1819.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee HB, Yu MR, Song JS and Ha H: Reactive
oxygen species amplify protein kinase C signaling in high
glucose-induced fibronectin expression by human peritoneal
mesothelial cells. Kidney Int. 65:1170–1179. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu ZG: Molecular mechanism of TNF
signaling and beyond. Cell Res. 15:24–27. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Baeuerle PA and Baltimore D: NF-kappaB:
Ten years after. Cell. 87:13–20. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ip YT and Davis RJ: Signal transduction by
the c-Jun N-terminal kinase (JNK)-from inflammation to development.
Curr Opin Cell Biol. 10:205–219. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jones WK, Brown M, Wilhide M, He S and Ren
X: NF-kappaB in cardiovascular disease: Diverse and specific
effects of a ‘general’ transcription factor? Cardiovasc Toxicol.
5:183–202. 2005. View Article : Google Scholar : PubMed/NCBI
|